
Abstract
There is increasing evidence that even low levels of blast cause brain injury, but little is known about their thresholds and mechanisms. Exposure of rats to 10–60 kPa blasts elevate intracranial pressure (ICP) in a dose-dependent manner and impair cognitive function. We have evaluated a prophylactic measure against these brain injuries in a rat animal model, consisting of feeding them processed cereal. This type of feed is known to ameliorate disturbances in secretion of body fluids and to have anti-inflammatory effects. In humans, intake of processed cereals is effective against intestinal diarrhea and also reduces the symptoms of Ménière's disease. Rats were given either standard laboratory feed or processed cereal feed for 2 weeks before exposure to blast in a shock tube. The ICP was monitored at different time points up to 1 week after exposure to a 60-kPa blast, and for up to 24 h after exposure to a 30-kPa blast. Maximal ICP elevation was reached at 10 h in both groups. In the group of rats on standard feed exposed to 60 kPa, an ICP increase of 145% was noted at 10 h, and the corresponding increase in the rats fed processed cereal feed was only 50%. In rats exposed to a 30-kPa blast, those fed standard feed and processed cereal feed demonstrated increases of ICP of 80% and 40%, respectively. Cognitive function as measured by the Morris water maze was assessed in other groups of rats at 2 days after exposure to 10- or 30-kPa blasts. Their performance was significantly impaired at both exposure levels in rats on standard feed, but no functional impairment was seen in rats fed processed cereal feed.
Introduction
C
We recently showed, in a rat model, that exposure to low levels of blast overpressure (10–60 kPa) impairs cognitive function and elevates intracranial pressure (ICP) in a dose-dependent manner (Säljö et al., 2009). Here we use the same model to investigate whether the consumption of processed cereal feed can prevent the development of the effects of blast on ICP and cognitive function. These feeds also have positive clinical effects on conditions involving disturbed fluid balance, such as diarrhea (Bjorck et al., 2000) and Ménière's disease (Hanner et al., 2004, 2009). This study shows that blast-induced ICP elevations and cognitive impairments are prevented by the consumption of processed feed for 2 weeks prior to mild to moderate blast exposure. The simplicity and safety of this prophylactic intervention makes it a good candidate for the prevention of blast-mediated brain injury.
Methods
Animals
Male Wistar rats with a body weight of 230–330 g (B&K Universal, Stockholm, Sweden) were housed 3–4/cage at a constant room temperature of 20 ± 1°C and humidity of 55%, with a day/light cycle of 6/18 h. The rats were kept in the animal facility for 2 weeks prior to blast exposure to establish the effects of the two types of feeds and to acclimatize them to the environment and human handling. During this time they were provided food and water ad libitum. A total of 180 rats were used: 136 for ICP measurements and 44 for the assessment of cognitive function. Both types of assessments were performed by the same investigator, who was blinded to group assignment. The study was approved by the Ethics Committee for Animal Experiments, Umeå, Sweden, and was performed according to the Guidelines for the Care and Use of Laboratory Animals.
Feeds
The animals were given either standard laboratory feed (2016 Teklad Global 16% protein diet; Harlan Laboratories Inc., Indianapolis, IN), or feed consisting in part of hydrothermally processed cereal (Specially Processed Cereal [SPC®]; Lantmännen AS-Faktor AB, Stockholm, Sweden). This constituted 20% of the feed, and the remaining 80% was a standard rodent diet (Labfor R36; Lantmännen, Kimstad, Sweden). The 20% contribution was judged to be similar to the intake of SPC by humans as a supplemental nutrient (Bjorck et al., 2000; Hanner et al., 2004, 2009). The concentrations of nutrients, vitamins, and minerals in the two types of feed are shown in Table 1. The final concentrations of the 20%/80% feed mixture were calculated from the product descriptions for Labfor R 34 and SPC. A comparison of the concentrations of sugars and amino acids cannot be made with the available product descriptions. However, the effect of the hydrothermal processing on the concentrations of certain sugars and amino acids can be found in the article by Björck and associates (2000). The processing increases the level of glucose from 0.3 to 5.3 mg/g. For fructose, sucrose, and maltose, the pre-processing levels of 0.3, 11.4 (mean value), and 0 mg/g, are increased to 3.1, 65.7, and 9.0 mg/g, respectively. The concentrations of the amino acids histidine, glutamic acid, lysine, tryptophan, and isoleucine were also raised considerably by processing (Björck et al., 2000).
The product description for the standard feed is taken from the information sheet of Harlan Laboratories. The product description for the processed cereal feed is taken from the information on the product AS-Faktor SPC-Flakes® and the information sheet of Labfor Lantmännen. The calculations for the processed cereal feeds were made by the authors. For full addresses of the companies, see Materials and Methods.
Blast exposure
The rats were anaesthetized with an IP injection of diazepam (1 mL/kg body weight Stesolid Novum; A/S Dumex, Copenhagen, Denmark), sodium pentobarbital (60 mg/kg), and saline, mixed 2:1:1 before blast exposure. They were equipped with earplugs and allowed to breathe spontaneously. Blast exposure was generated with a 3.1-m-long, 0.2-m-diameter shock tube (Säljö et al., 2008). The rat was placed inside the tube 0.25 m from the opening of its expansion compartment with the long axis of its body parallel to the tube and the head facing the origin of the blast. The rat was firmly fixed by its abdomen to a horizontal shelf secured with a vertical support. The pressures were selected such that they had small but significant effects on the rats' ICP (Säljö et al., 2009). Furthermore, exposure at 30 kPa seems to be the lowest dose that causes effects on the histological neuropathology in pigs (Säljö et al., 2008). Pulses with a Pmax of 10, 30, and 60 kPa as measured in the air in close proximity to the head were generated with pressures of 0.2, 0.6, and 1.2 atm, respectively, and at a duration of 4–6 msec. The compression compartment was 1.6 m long and its volume was 49 L. A cellophane membrane, separating the compartments of the shock tube, was perforated to rupture at the desired pressure. All animals were exposed three times within 20 min at the same pressure. The controls received anesthesia and were not exposed to blast.
Intracranial pressure
Two groups of rats given standard laboratory feed were exposed to 30 kPa and 60 kPa (Table 2). Two other groups were given the processed cereal feed were also exposed to 30 kPa and 60 kPa blasts (Table 2). Control (unexposed) rats were fed standard feed or processed cereal feed without blast exposure. The standard laboratory feed and processed cereal feed groups were then randomly assigned to subgroups to investigate the time course of ICP changes (Table 2). In the 60-kPa groups, the ICP was recorded at 0.5, 2, 6, 10, and 24 h, and 2 and 7 days after exposure, while ICP was recorded only at 2, 10, and 24 h in the 30-kPa groups (Table 2).
Calculated using two-way ANOVA.
The rats were anesthetized and their heads were fixed in a stereotactic device to reduce skull movement. A miniature optic probe (diameter 0.42 mm, Samba 201/202 system; Samba Sensors AB, Västra Frölunda, Sweden) was protected from direct contact with the brain tissue by a 1.1-mm (outer diameter) piece of Venflon tubing. The probe was inserted through a 1.3-mm hole drilled in the skull 2 mm behind the bregma and 2 mm from the midline, and into the brain parenchyma to a depth of 6 mm from the surface of the skull. The Venflon tubing was secured to the bone with biological glue (Histoacryl® Blue; B. Braun, Aesculap, AG & Co. KG, Tuttlingen, Germany). The sensor at the end of the probe was connected to a control unit (Samba 201/202) and a personal computer with software for processing and analysis. The pressure transducers were calibrated before each ICP measurement. A steady-state ICP level was reached 2–3 min after insertion of the sensor. The ICP was then recorded for 10 min to ascertain the mean value. During this period, the difference between the lowest and highest pressures recorded was <1 mm Hg. Each rat with blast exposure was used for ICP measurement at a single time point, and the sham-injured rats were included at all time points. The rats were euthanized immediately afterward, and the brains were inspected for injuries and proper placement of sensor. All ICP measurements were performed by the same investigator.
Cognitive function
Cognitive outcome was assessed with the Morris water maze (MWM; Morris, 1984). Six groups of rats were used for analysis of the effects of exposure to 10-kPa and 30-kPa blasts (Table 3). The exposed rats fed standard feed were compared to the exposed rats fed processed cereal feed. Cognitive outcomes were not assessed after exposure to 60-kPa blasts.
The rats were exposed at 10 or 30 kPa on the day 7 and were tested 2 days after blast exposure.
The MWM procedure is described in an article by Säljö and colleagues (2009). The maze pool was a water-filled circular plastic tank (22 ± 1°C) with a diameter of 1.6 m. A 0.1-m acrylic glass platform was submerged 10 mm below the water's surface in one of the pool's four quadrants. The time to find the platform (latency), path length, and swimming speed were recorded by an overhead camera connected to an externally-located computer (HVS Image software and the 2020 Plus Tracking system; HVS Image Ltd., Buckingham, U.K.). The training sessions were performed once daily for 6 days and were begun at 8
Statistical analysis
The statistical analyses were performed with SPSS 15.0 (SPSS, Inc., Chicago, IL), including the software Sample Power 2. Data are presented as means ± SEM. The MWM acquisition trials of the exposed animals on standard or processed cereal feed and unexposed animals on standard feed were analyzed with a paired sample t-test before and after exposure. The ICP measurements were analyzed using the independent sample t-test. The two blast exposure levels were compared with respect to ICP values at the post-injury time points of 2 and 10 h and 1 day (for 30 kPa), and 0.5, 2, 6, and 10 h, and 1, 2, and 7 days (for 60 kPa). Two-tailed levels of significance were used.
Results
Animals
The increase in body weight of the animals fed the processed cereal feed was similar to that of the animals on standard feed. Furthermore, weight gain appeared to be unaffected by the blast exposure in rats on both types of feed.
Intracranial pressure
The mean value in the control groups ranged between 5.8 and 6.6 mm Hg (Table 2). In the standard-feed animals exposed at 30 kPa, the mean ICP had increased by 90% at 10 h (p < 0.0001; i.e., the peak value in the series). The corresponding peak value in the animals on processed cereal feed showed an increase of 60% (Table 2). In rats exposed to 60 kPa, the group on standard feed had an ICP elevation of 145% at 10 h, while those on processed cereal feed had increases of only 50% (p < 0.0001; Table 2). In the rats on standard feed, the ICP elevation was 40% at 0.5 h, and the processed cereal feed did not reduce this initial ICP elevation (Fig. 1).
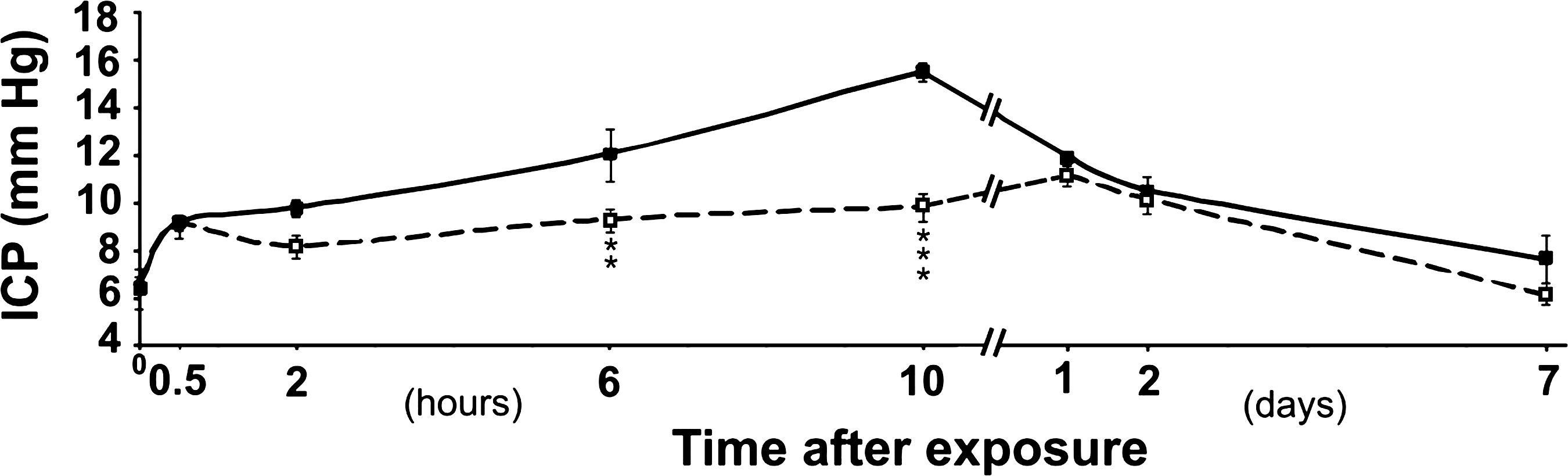
Time course of intracranial pressure (ICP) changes seen after exposure of rats to 60-kPa blasts. Means are shown of rats on standard feed (solid squares) and rats on processed cereal feed (open squares). Bars indicate standard error of the mean. Control levels are given at time point zero. The ICP of the animals on the two types of feed differed significantly (***p < 0.0001; **p < 0.02).
Morris water maze
The training period led to a similar decrease in time to find the platform in all groups (i.e., from approximately 50 sec on day 1 to around 10 sec on day 6; Table 3 and Fig. 2). In the control (unexposed) groups on standard feed, the latency to platform decreased further on the re-test on day 9, while the latency increased by 60–120% (p < 0.01) in the groups exposed to 10-kPa blasts, and 20–150 % (p < 0.01) in the groups exposed to 30-kPa blasts. In rats given processed cereal feed and exposed to 10 and 30 kPa, the latencies to platform decreased on the re-test in a fashion similar to the controls, and differed significantly from those of exposed rats on standard feed (p < 0.01). The total path length was variable and paralleled the changes in latencies, while the swimming speed was not appreciably affected, even after exposure to 30-kPa blasts.

Performance in the Morris water maze. All rats were exposed to blast overpressure at 10 kPa on day 7. Animals on standard feed are shown with solid squares, and animals on processed cereal feed are shown with open squares. Bars indicate standard error of the mean. The 6-day training period led to a similar decrease in both groups with respect to time to find the platform. In the re-test on day 9, the animals on processed cereal feed had further shortened their time to find the platform, while the animals on the standard feed had a prolonged time to find the platform. The performance on day 9 differed significantly (** p < 0.01) between the groups.
Discussion
In a recent report, we showed a dose-dependent rise of ICP and a reduction in cognitive function in rats exposed to levels of blast overpressure with a Pmax as low as 10 kPa (Säljö et al., 2009). After exposure of rats to 60 kPa, the ICP response consisted of a smaller initial peak at 0.5 h, and a main peak at 10 h. At this time, the ICP had been elevated to 16 mm Hg from the control level of 6 mm Hg. In severe, non-blast-related TBI in rats, the ICP is elevated to 25–47 mm Hg (Jamali et al., 1998; Rooker et al., 2003).
Interspecies differences with respect to absolute peak levels and time course of ICP elevations (Stocchetti et al., 2007) do not allow for direct extrapolation to humans. The current medical therapeutic options aimed at reducing elevated ICP are less than satisfactory with regard to outcomes, and are not indicated at an ICP below 20–25 mm Hg (Jantzen, 2007; Orban and Ichai, 2007; Westhout et al., 2008). In combat conditions individuals may have increased ICP, but it remains below this critical level. The clinical significance and the short-, medium-, and long-term effects of ICP below treatment thresholds are uncertain. If mild or modest elevations in ICP are found to be of significance in the physical, psychological, and social functioning of patients with blast-related mild TBI, then prophylaxis leading to an ICP reduction of 40–100% would be highly beneficial.
This study also provides clues about the mechanism of blast-related TBI. In clinical blast-related moderate and severe TBI, brain edema has been identified by imaging studies (Armonda et al., 2006). In a blast-induced mild TBI pig model, we have shown histological changes consistent with edema (Säljö et al., 2008). We hypothesize that the impaired cognitive function in the blast-exposed rats is secondary to brain edema. There were no indications that motor function, at least as judged by the swimming speed, was involved. Here we show that a processed cereal diet acts to prevent the impairment of cognitive function as measured by the MWM, and to significantly ameliorate the ICP elevation. While the feed blocked the main peak of ICP elevation, it did not affect the initial peak (Fig. 1). This suggests that the feed specifically reduces brain edema, since the initial increase in ICP is thought to be vascularly mediated and reversible (Engelborghs et al., 1998; Fritz et al., 2005).
TBI research involves measurement of cognitive impairment (Parton et al., 2005), which often is correlated with injuries to neuronal cell groups, particularly in the hippocampus (Hamm et al., 1993; Sanders et al., 1999). Cognitive impairment may, however, occur without apparent neuronal loss or injury (Lyeth et al., 1990). TBI treatments comprised of administration of substances acting on neurotransmitter systems (Kokiko and Hamm, 2007) may provide better neuronal survival (Statler et al., 2006). Less specific treatments, such as those targeting brain edema, are also protective. As a rule, diets or supplements administered otherwise contain protective substances. Supplementation with morphine (Zohar et al., 2006), melatonin (Ozdemir et al., 2005), progesterone (Jones et al., 2005), magnesium (Hoane et al., 2008), nicotinamide (Hoane et al., 2006), and choline (Guseva et al., 2008) improve outcomes after TBI. In other cases, secondary effects of the diet may be at work, but their protective mechanisms are often unknown. Dietary curcumin, having numerous pharmacological activities, counteracts reductions in the levels of molecules involved in energy homeostasis (Sharma et al., 2009). The beneficial effect of lactate (Rice et al., 2002) may be of a similar nature.
At least part of the TBI protection achieved with ketogenic diets (Appelberg et al., 2009; Hu et al., 2009; Wu et al., 2004, 2007) is due to the increase in the levels of brain-derived neurotrophic factor (Wu et al., 2008). Other factors that attenuate brain edema (Doczi, 1993; Rosenberg and Estrada, 1995; Vajda et al., 2001) are effective against ICP elevations (Lorenzo et al., 1989), and are active in organs other than the brain (Clerico et al., 1992; Halperin et al., 1985; Halperin et al., 1988; Hamlyn et al., 1991).
The processed cereal diet used to induce this activation is effective against post-weaning diarrhea in pigs (Lonnroth et al., 1988), and is also beneficial in humans with inflammatory bowel disease (Bjorck et al., 2000) and Ménière's disease (Hanner et al., 2004, 2009). The composition of the diet does not differ radically from a standard diet with respect to the available nutritive values, vitamins, and minerals. However, the process substantially raises the concentration of certain sugars and amino acids. Amino acid concentrations are also adequately supplied by the standard diet. The levels of various sugars are not provided in the products' documentation, making it impossible to make exact comparisons, though the effect on sugar levels is well known (Björck et al., 2000). The important properties of the processed cereal diet may well be outside the realm of the analytical protocols used to assess them. The diet itself does not contain any of the active compounds, but it stimulates their production. At least part of the activity of the diet can be reproduced by a feed containing certain sugars and amino acids (Lange and Lonnroth, 2001). Both sugars and amino acids stimulate the production of a partly purified protein, referred to as antisecretory factor, which regulates fluid balance (Lange and Lonnroth, 2001). A cloned peptide was shown to be an active region of this protein (Johansson et al., 1997, 1995). This peptide also suppresses the raised ICP seen in rats with experimental herpes simplex encephalitis (Jennische et al., 2008). Fluid-regulating activity has also been ascribed to a fatty acid amide, erucamide, as determined by a study of rats with cholera toxin–induced diarrhea (Hamberger and Stenhagen, 2003). Fatty acid amides have been the subject of considerable scrutiny as a new class of bioactive lipids. Oleamide, a fatty acid amide similar to but slightly smaller than erucamide, induces physiological sleep (Mendelson and Basile, 1999), and another fatty acid amide, anandamide, demonstrates analgesic properties (Walker et al., 1999).
While the effects of the processed cereal feed on blast-induced ICP elevation are significant, the precise mechanisms remain unknown. Furthermore, the significance of subclinical elevations of ICP in humans is unknown. The therapy may have limitations with regard to the timing of administration (i.e., pre-exposure or very early post-exposure), and have treatment applicability only in less severe TBI. On the other hand, cereals can be consumed with ease, are part of the daily diet in a number of countries, and have proven safety and efficacy in humans afflicted with other conditions. Finally, the currently available medical therapies for mild TBI are limited. This study also expands our knowledge about the potential mechanisms at work in the evolution of blast-induced mild TBI. Further investigations focusing on edema formation, fluid transport, and inflammatory pathways and their regulation may prove to be promising.
Footnotes
Acknowledgments
The authors want to express their gratitude to Svante Höjer, Samba Sensors AB, Västra Frölunda, Sweden, for sharing with us his knowledge of pressure sensors and pressure recording; to Stefan Lange, Department of Infectious Medicine, Sahlgrenska Academy, Göteborg, Sweden; and Lantmännen AS-Faktor AB, Stockholm, Sweden, for providing advice and the cereal feed. This study was supported by grants from the Swedish Armed Forces/FMV and Svenska Militärläkareföreningen.
Author Disclosure Statement
No competing financial interests exist.