
Abstract
Assessment of motor abilities after sciatic nerve injury in rodents, in particular mice, relies exclusively on walking track (footprint) analysis despite known limitations of this method. Using principles employed recently for video-based motion analyses after femoral nerve and spinal cord injuries, we have designed and report here a novel approach for functional assessments after sciatic nerve lesions in mice. Functional deficits are estimated by angle and distance measurements on single video frames recorded during beam-walking and inclined ladder climbing. Analyses of adult C57BL/6J mice after crush of the sciatic, tibial, or peroneal nerve allowed the identification of six numerical parameters, detecting impairments of the plantar flexion of the foot and the toe spread. Some of these parameters, as well as footprint functional indices, revealed severe impairment after crush injury of the sciatic or tibial, but not the peroneal nerve, and complete recovery within 3 weeks after lesion. Other novel estimates, however, showed that complete recovery is reached as late as 2–3 months after sciatic nerve crush. These measures detected both tibial and peroneal dysfunction. In contrast to the complete restoration of function in wild-type mice (100%), our new parameters, in contrast to the sciatic functional index, showed incomplete recovery (85%) 90 days after sciatic nerve crush in mice deficient in the neural cell adhesion molecule (NCAM). We conclude that the novel video-based approach is more precise, sensitive, and versatile than established tests, allowing objective numerical assessment of different motor functions in a sciatic nerve injury paradigm in mice.
Introduction
N
Another valuable research paradigm is the femoral nerve model characterized by preferential motor reinnervation of the motor quadriceps branch, and less so of the sensory saphenous branch (Brushart, 1988, 1993; Madison et al., 1996). Bifurcation of the nerve into two major branches allows studies on mechanisms, determining the molecular selectivity of reinnervation. We have recently devised a video-based method for analysis of motor functions after femoral nerve injury in mice, which we have designated single-frame motion analysis (SFMA) (Irintchev et al., 2005). The method is based on measurements of specific angles, the foot-base and heels-tail angle, at defined step cycle phases during beam-walking and on limb length measurements during voluntary reaching pursuits with the hind limbs using selected frames of ordinary video recordings. The reliability and sensitivity of this quantitative evaluation approach have been validated in studies on nerve regeneration in knockout mice (Eberhardt et al., 2006; Guseva et al., 2009; Malin et al., 2009) and for assessment of treatment strategies for improvement of functional recovery (Ahlborn et al., 2007; Mehanna et al., 2009b; Simova et al., 2006).
In the sciatic nerve lesion paradigm, the most commonly used model in which motor and sensory axons supplying a large number of lower leg muscles are severed, walking track (footprint) analysis has been the method of choice for more than 25 years in studies using rats (de Medinaceli et al., 1982; see Varejão et al., 2004, for a review of the method and modifications). The method is based on measurement of footprints of walking rats, and sciatic nerve function is estimated by the sciatic functional index or, in case of lesions of the tibial and peroneal branches of the sciatic nerve, by the tibial and peroneal functional indices, respectively. A disadvantage of the footprint analysis is that it is highly dependent on the quality of the footprints and the locomotion speed. More importantly, the precision and thus the discriminative power of the method are limited (Munro et al., 1998; Schiaveto de Souza et al., 2004; Shenaq et al., 1989; Urbanchek et al., 1999; Valero-Cabré and Navarro, 2002; Varejão et al., 2003a; Yu et al., 2001). Awareness of the insufficiencies of the track analysis has led researchers to seek novel approaches, and several kinematical parameters, such as gait-stance duration, ankle angle, and toe-out angle, have been proposed for evaluation of sciatic nerve function in rats (Santos et al., 1995; Varejão et al., 2003a,b; Walker et al., 1994; Yu et al., 2001). Application of these techniques has been limited, most likely because of the expensive technical equipment required and lack of clear advantages over the traditional footprint analysis (Varejão et al., 2004). More recently, a sophisticated computerized approach, the CatWalk gait analysis, has been tested in mice and rats with sciatic nerve injury (Bozkurt et al., 2008; Deumens et al., 2007; Vogelaar et al., 2004). This method uses video recordings of walking trials along a glass runway and allows simultaneous measurements of numerous static and dynamic gait parameters. The CatWalk is sensitive in the sense that it reveals, in contrast to the walking track analysis, long-lasting gait deficits after injury (Bozkurt et al., 2008; Vogelaar et al., 2004). However, the high cost of the equipment and the need for extensive training of the animals, including food restriction and rewarding, are limitations to this approach.
Compared with rats, the experience with walking track analyses in mice is limited. The calculations of functional indices for sciatic, tibial, and peroneal injuries have been modified for the mouse (Inserra et al., 1998), and it has been shown that this functional assay can detect differences in recovery after injuries of different severity (i.e., crush, nerve transection without repair, and transection followed by epineural suture) (Yao et al., 1998). Walking track analysis has revealed differences in functional regeneration between knockout and wild-type mice (Bannerman et al., 2008; Boivin et al., 2007; Narciso et al., 2009; Taylor et al., 2001; Yao et al., 1999). However, the precision of the method is questionable, even more so than for the rat, considering the small body size of the mouse. Alternative methods have not been created or evaluated.
We attempted to adapt our single-frame motion analysis developed for the femoral nerve to the sciatic nerve paradigm in the mouse. In pursuing this goal, we were encouraged by the successful translation of this approach to a compression spinal cord injury model in the mouse (Apostolova et al., 2006; Chen et al., 2007; Jakovcevski et al., 2007; Lee et al., 2009; Mehanna et al., 2009a). Here we report a battery of parameters that are measurable using frames of video recordings of beam-walking and inclined ladder climbing, allowing a precise evaluation of motor functions.
Methods
Experimental design
Three sets of experiments were performed. In experiment 1, 3-month-old female C57BL/6J mice were subjected to unilateral sciatic nerve crush and video recorded during beam-walking and inclined ladder climbing before injury and 1, 3, 5, 7, and 10 days after crush. The purpose of this pilot experiment was to detect injury-induced changes in locomotion and define parameters for quantitative assessment of deficits. In experiment 2, female C57BL/6J mice were subjected to either tibial, peroneal, or sciatic nerve crush and motor deficits were estimated using walking track analysis and a battery of video-based parameters over a 4-week observation period. The aims of this experiment were to assess the reproducibility in measurement and the sensitivity of individual parameters with respect to different types of injury, and to determine course and degree of functional recovery on a short-term time scale. In experiment 3, we compared recovery from sciatic nerve crush in neural cell adhesion molecule-deficient (NCAM-/-) mice and wild-type (NCAM+/+) littermates over a 3-month time period. The rationale behind this experiment was to test the potential of the traditional approach, walking track analysis, and of the novel parameters to detect deficits in motor recovery in a knockout mouse model. Sufficient evidence suggests that NCAM and the glycan α2,8 polysialic acid (PSA), a carbohydrate carried by the membrane-associated and soluble forms of NCAM (Bock et al., 1987; Olsen et al., 1993; Rutishauser, 2008) are involved in neural regeneration. Expressed abundantly by Schwann cells and axons in the peripheral nervous system during embryonic development, PSA-NCAM expression decreases in adulthood. After nerve injury in adult rodents, PSA-NCAM is upregulated in axons, Schwann cells, and skeletal muscle fibers (Covault et al., 1986; Daniloff et al., 1986; Olsen et al., 1995; Sanes et al., 1986). Recent work (Franz et al., 2008) has suggested that increased PSA-NCAM expression after nerve injury is necessary for preferential targeting of regenerating motor axons to skeletal muscle as opposed to skin, a phenomenon known as preferential motor reinnervation (Brushart, 1988, 1993). The ablation of NCAM in mice causes, in the absence of nerve injury, abnormalities in the structure and function of motor endplates (Rafuse et al., 2000), which could lead, after nerve injury, to deficient maturation of synaptic transmission at regenerated endplates (Badke et al., 1989). In support of this notion, Chipman and colleagues (2010) recently showed that the process of reinnervation in NCAM-/- mice is compromised with time after injury due to instability of the newly formed endplates leading, by 3 months after injury, to deficits in neurotransmitter release, withdrawal of axons from reinnervated endplates, and loss of contractile muscle force.
In all experiments, the size of the animal groups was intentionally limited to 7–8 mice in order to more stringently test, as compared with larger groups (>10 animals) typically used in regeneration research, the sensitivity of the methods. The peroneal group in experiment 2 was smaller still (n = 5), because two of the mice initially ascribed to this group were too anxious during beam walking and ladder climbing and were excluded prior to operation.
Animals and surgical procedures
We used female C57BL/6J and female NCAM-deficient (NCAM-/-) mice and their wild-type (NCAM+/+) littermates (C57BL/6J genetic background; 12 backcrosses) at the age of 3 months. The generation of NCAM-deficient mice has been described (Cremer et al., 1994). All animals were bred at the central animal facility of the Universitätsklinikum Hamburg Eppendorf. The mice were treated according to the German law on protection of experimental animals. For surgery, the animals were anesthetized by intraperitoneal injections of 0.4 mg kg−1 fentanyl (Fentanyl-Janssen, Janssen, Neuss, Germany), 10 mg kg−1 droperidol (Dehydrobenzperidol, Janssen) and 5 mg kg−1 diazepam (Valium 10 Roche, Hoffmann–La Roche, Grenzach-Wyhlen, Germany). The right sciatic nerve was exposed by skin incision and opening of the fascial plane between the gluteus musculature. Crush injury at the level of trochanter major was performed with Dumont forceps #5 (Fine Science Tools, Heidelberg, Germany) by manually pressing the nerve between the forceps tips for 15 sec. In two groups of C57BL/6J mice (experiment 1 and 2, n = 5 and 7, respectively) and in NCAM-/- and NCAM+/+ mice (experiment 3, n = 8 and 7, respectively), both the tibial and the peroneal branches of the sciatic nerve were simultaneously injured. In two other groups of C57BL/6J mice (experiment 2), the two sciatic nerve branches were carefully separated by blunt dissection and only the tibial branch or the peroneal branch (n = 7 and 5, respectively) were damaged. After the injury, the skin wound was closed with 6-0 sutures (Ethicon, Norderstedt, Germany) and the animals were kept in a warm room (35°C) for several hours to prevent hypothermia. Complications of sciatic nerve injuries in rodents like autotomy (self-mutilation), skin ulcerations, and limb contractures were not observed in any of the operated animals during the period of postoperative observation.
Walking track analysis
The plantar surface of the hind paws was moistened with black ink (Tinte 401, Pelikan, Schindellegi, Switzerland), and the mice were led to walk along a wooden beam (see below) on which a stripe of graph paper (1000 × 38 mm) was fixed using adhesive tape. Print length (PL) and toe spread (TS) were measured for at least four left and four right footprints per animal and time point using digitized images of the prints and ImageTool 2.0 software (University of Texas, San Antonio, TX;
Single-frame motion analysis
Beam-walking test
The beam-walking test was performed as described previously (Irintchev et al., 2005). Mice were accustomed, in 3–4 trials, to beam walking prior to operation. In this test, the animal walks unforced from one end of a horizontal beam (length 1000 mm, width 40 mm) toward its home cage located at the other end of the beam. For all mice, rear, left-, and right-side views of walking trials were captured once prior to operation and at different time-points after surgery with a video camera (A602fc, Basler, Ahrensburg, Germany) at 100 frames per sec and stored on a personal computer in audio video interleaved (AVI) format using SIMI-Motion 7.0 software (SIMI Reality Motion Systems, Unterschleissheim, Germany). The video sequences were examined using VirtualDub 1.6.15, a video capture/processing utility (software, written by Avery Lee, available at
Ladder-climbing test
After the beam-walking test, the animals were video recorded during unforced climbing up an inclined ladder as described previously for spinal cord injured mice (Apostolova et al., 2006). The ladder was made of a 4-mm-thick frame cut from a Resopal® plate (Resopal, Groß-Umstadt, Germany; 96 long × 12 cm wide with central incision of 88 × 6 cm) to which 43 round wooden rungs (100 × 2 mm) were glued at equal intervals (2 cm). The ladder was fixed in an inclined position (55°) using a Plexiglas platform. The mice were placed at the bottom rungs of the ladder, and climbing was video recorded from a position “below” the ladder (i.e., viewing the ventral aspect of the animals).
Statistical analysis
One-way analysis of variance (ANOVA) for repeated measurements with Dunnett's or Tukey's post hoc tests was used to compare mean values. The threshold value for acceptance of differences was 5%. All data are presented as mean values with standard errors of mean.
Results
Definition of video-based parameters
In experiment 1, we video recorded mice (n = 5) prior to and 1 to 10 days after sciatic nerve crush to analyze changes in locomotion on the beam and the inclined ladder and to define parameters for quantitative estimation of motor deficits. Sciatic nerve injury in mice and rats causes impairment of the foot plantar flexion, that is, the ability to increase the angle between foot and leg (ankle extension), controlled by the tibial nerve (Bain et al., 1989). As a result, stepping with the injured leg changes from digitigrade (on the toes) to plantigrade (setting the whole length of the foot on the ground). In footprint analyses, this deficit is estimated by the increase of the print length after injury. Our experiments indicated that this deficit can be reliably estimated by measurements of angles between the foot axis and the horizontal plane at toe-off position during beam walking, the foot-base and the lateral foot-base angle (Fig. 1A–D). In addition, we noticed that after take-off during beam walking, in the mid-swing phase, the toe spread on the injured side is reduced compared with intact animals (Fig. 2A–D). This deficit, measured by the toe-spread angle, could reflect both tibial and peroneal dysfunction (Bain et al., 1989). Impairment of plantar flexion was also visible during ladder climbing. During the stance (weight-bearing) phase, impaired plantar flexion resulted in “hanging” of the heel below the rung while, in contrast, the heel was invisible prior to injury (Fig. 3A–D). As a measure of this abnormality we defined the ladder stance angle, which, in addition to plantar flexion, reflects to a small degree changes in the toe spread (Fig. 3C,D). Two other parameters were devised to estimate changes in toe spread during ladder climbing. Upon placement of the foot on a rung, the toes of uninjured mice are spread and firmly grasp the rung (Fig. 4A,C). After injury, the spread is decreased and the grasping is absent (Fig. 4B,D). The parameter defined for estimation of this injury-induced alteration is designated ladder toe spread and is measured as absolute length in mm. Finally, we noticed that while approaching the rung, the toes of uninjured mice were first spread (Fig. 5A) followed by an immediate and transient adduction (decrease of the spread, Fig. 5C) prior to grasping the rung (Fig. 4C). The transient change in the spread during target reaching, estimated by the ladder-spread index, declines after injury (Fig. 5A,C vs. Fig. 5B,D).
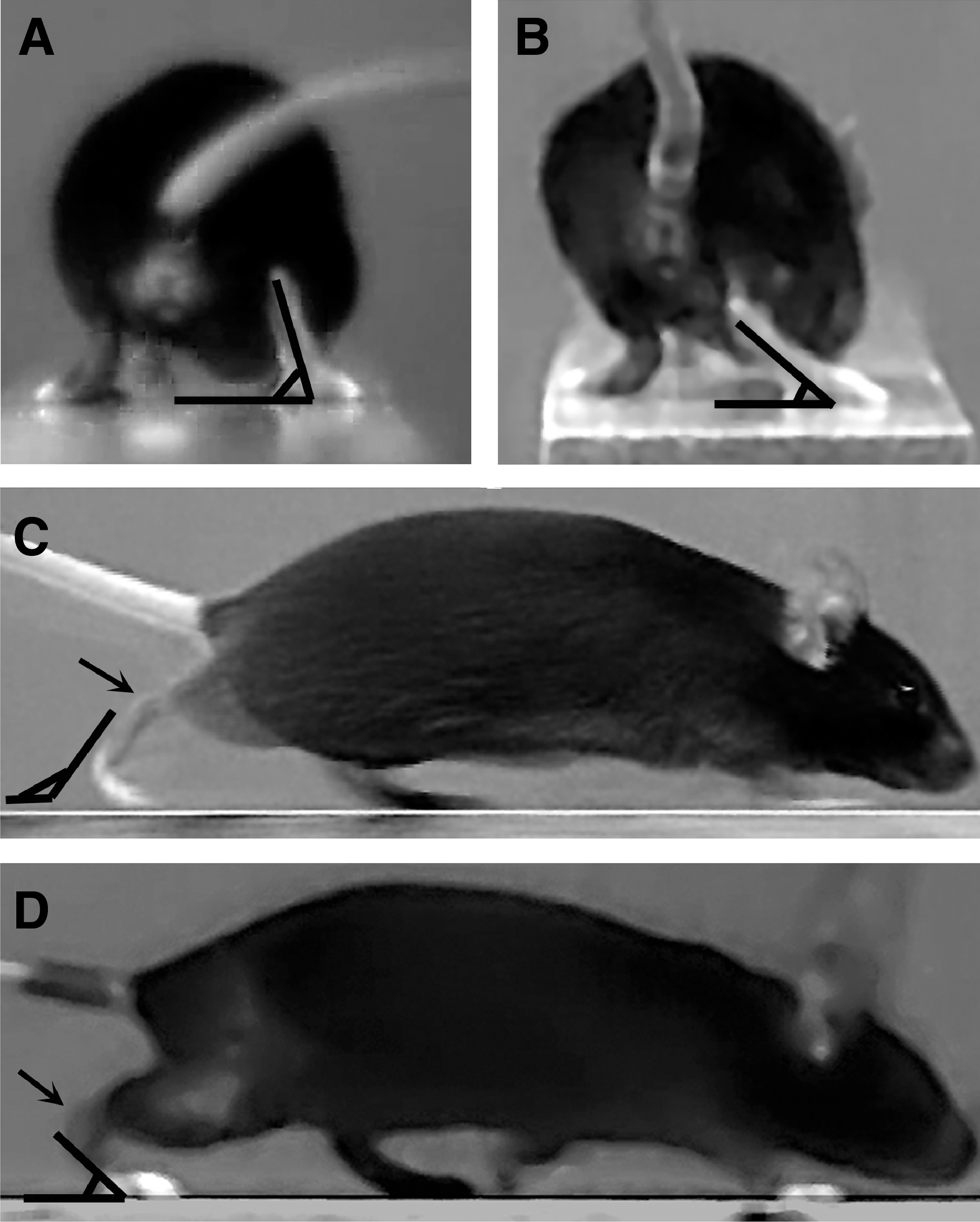
Foot-base angle and lateral foot-base angle (FBA and LFB) definitions. Shown are video frames recorded during beam-walking trials prior to injury (
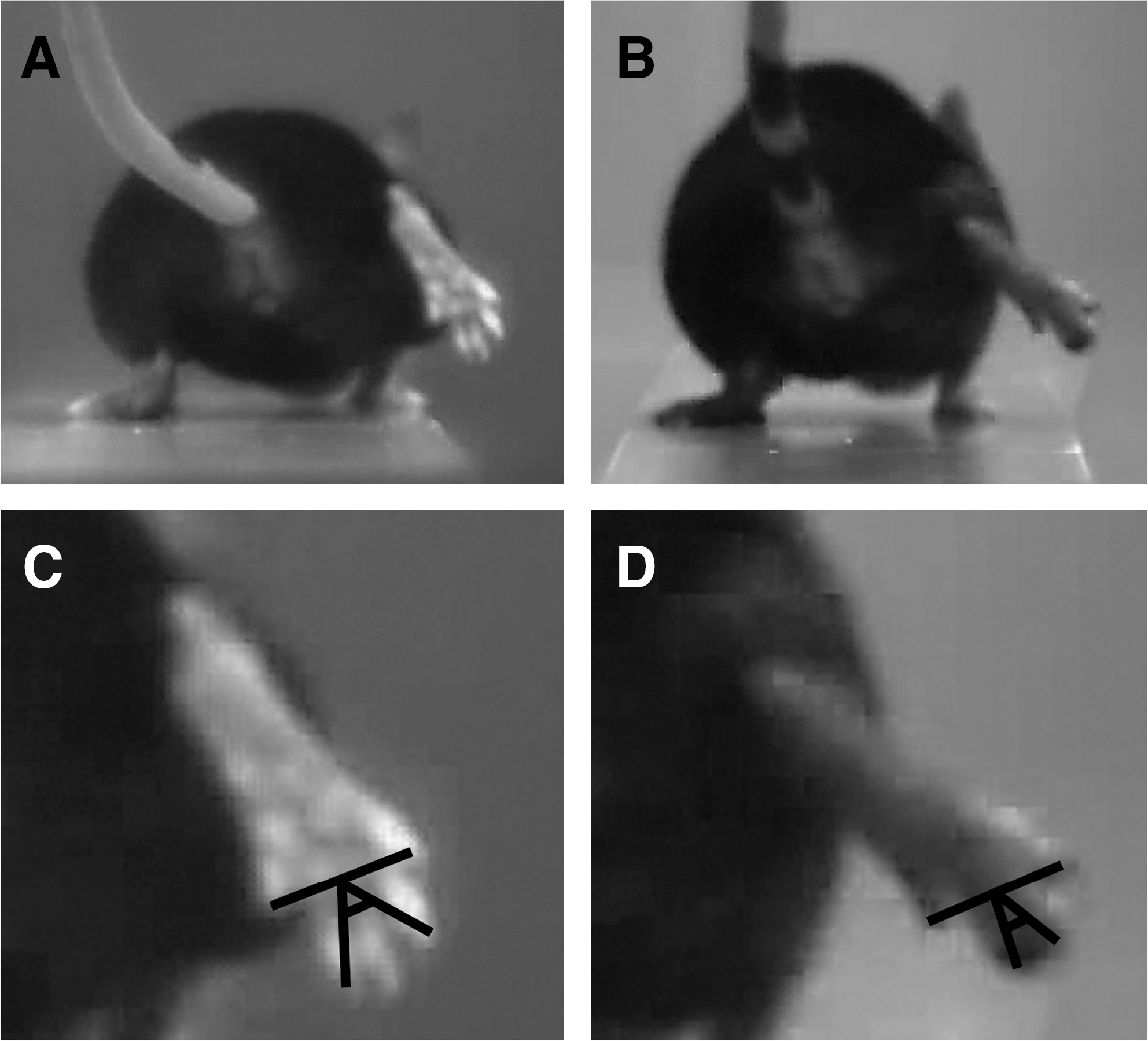
Toe-spread angle (TSA) definition. Shown are video frames recorded during beam-walking trials prior to injury (
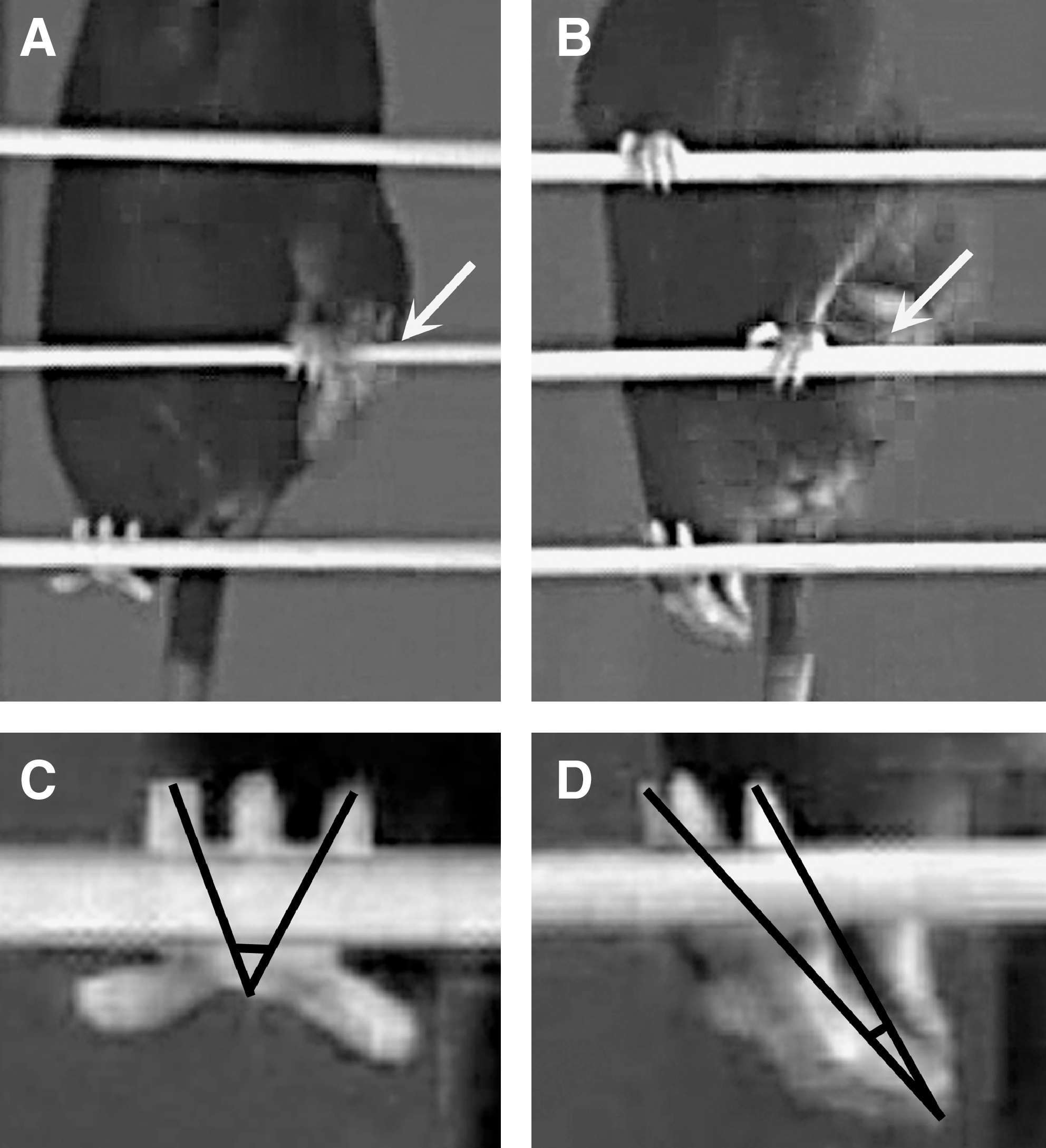
Ladder stance angle (LSA) definition. Shown are video frames recorded during inclined ladder climbing prior to injury (

Ladder toe-spread (LTS) definition. Shown are frames recorded during inclined ladder climbing prior to injury (

Ladder spread index (LSI) definition. Shown are video frames recorded during inclined ladder climbing prior to injury (
Because of zooming and camera movements, calibration of the video recordings (i.e., knowledge of pixel/absolute length ratios in the x- and y-axis required for classical motion analyses) was not possible. This was carefully considered in selection and definition of parameters. Four of our measures are independent of calibration because they are all based on angle measurements. However, these parameters differ in that two of them, the foot-base angles, are angles between two lines, a simple definition suitable for estimating alterations in foot placement with respect to the plane of locomotion. The other two estimates, the toe spread angle and the ladder stance angle, are three-point angles. This approach is better than two-line angle measurements in these cases because the only two lines that can be used, the axis of the second and fourth toes, are either poorly definable (ladder stance angle, toes only partially visible, Fig. 3C,D) or form too small an angle in intact mice (toe spread angle, Fig. 2C) to allow reliable estimates of changes (decline) at different time-points after injury. Angle measurements are unsuitable for estimation of toe spread during ladder climbing. Therefore, we use length measurements for the ladder toe spread and the ladder spread index. The calibration problem is solved in different ways for the two parameters. The ladder toe spread is normalized to the length of the rung on which the paw is placed and can thus, knowing the rung length, be easily calculated in absolute units, specifically, millimeters. The ladder spread index is a ratio of two length measurements in two successive video frames and thus is independent from paw distance from the camera.
During beam and ladder trials, mice mostly move at low to moderate speed and usually traverse the beam or ladder with several stops. When step cycles immediately preceding or following stops were excluded and the definitions of the gait parameters were precisely fulfilled, our measurements were very reproducible. In cases of fast movements (fast walking, running, jumping over several rungs), our criteria for measurement of the parameters were not fulfilled and such episodes were not analyzed. Also, all parameters are defined so that measurement errors due to camera movements and zooming while following the mice during walking or climbing are negligible. Our initial observations suggested that a suitable initial time-point of observation was 5 days after injury, a time-point when, on the one hand, the animals had gotten used to the sudden loss of function and pain or discomfort from the skin wound had been minimized and, on the other hand, muscle reinnervation had not occurred.
Walking track analysis
In experiment 2, we first assessed the sensitivity of the walking track analysis with respect to different types of injury in C57BL/6J mice, crush of the tibial or peroneal nerve only, or simultaneous crush of these two nerves (sciatic nerve crush). As expected based on previous data on rats and mice, sciatic nerve crush resulted in a significant decline in the sciatic functional index, from approximately 0 prior to injury to −100 at 5 days after the lesion (Fig. 6A). This rapid decline was followed by a similarly fast increase of the index and, compared with the preoperative value, no statistical difference was detectable already 14 days after injury. Interestingly, similar results were obtained when tibial and peroneal, instead of sciatic functional indices were calculated from the paw print values of the same animals (Fig. 6D,G). Significant gait disturbances were also detected 5 days after tibial nerve crush using all three functional indices (Fig. 6B,E,H). The sciatic index appeared to be more sensitive than the tibial and peroneal indices as it revealed differences up to day 21 after lesion (Fig. 6B) compared to differences at 5 and 7 days only found using the other two parameters (Fig. 6E,H). Finally, none of the three functional indices was significantly changed, compared with preoperative values, 5 to 28 days after peroneal nerve crush (Fig. 6G,F,I). These results show that functional indices in the mouse detect primarily tibial nerve dysfunction (plantar flexion). All three indices revealed a rapid, within 2–3 weeks, recovery from tibial and sciatic nerve injuries, while they were unable to detect functional impairments after peroneal nerve crush at any time point after injury.
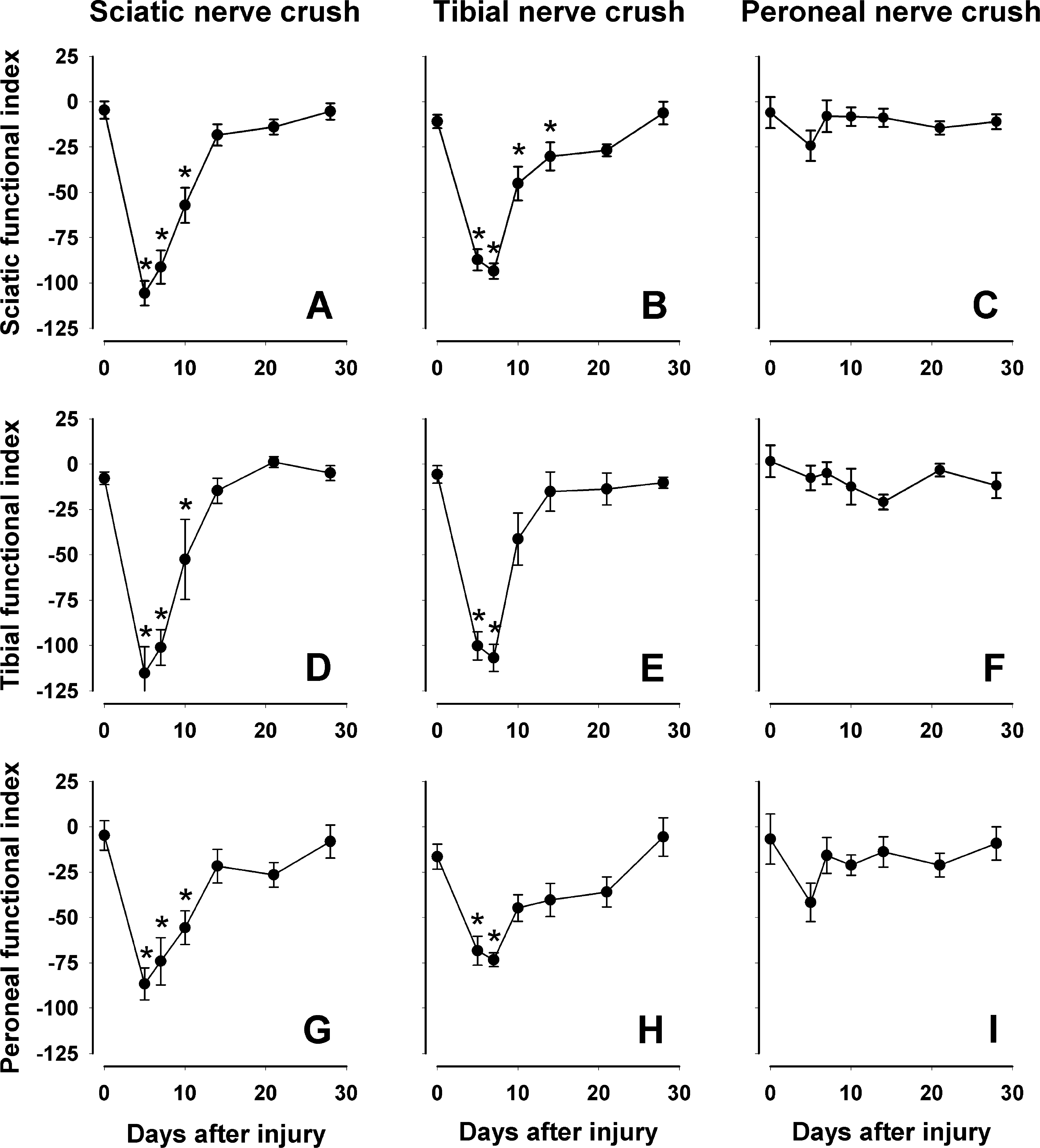
Walking track analysis. Comparison of different functional indices (sciatic,
Beam-walking analysis
In parallel to footprint measurements, we analyzed three beam-walking parameters for the animals subjected to sciatic, tibial, or peroneal nerve crush (experiment 2). Five days after sciatic and tibial, but not peroneal nerve crush, the foot-base angle and the lateral foot-base angle, measured using rear- and side-view video frames, respectively (Fig. 1A–D), declined significantly as compared with values at day 0 (Fig. 7A–C). Both parameters returned to normal values within 21 days after sciatic and tibial nerve injuries (Fig. 7A,B,D,E). Thus, similar to the functional indices, these two parameters detect basically tibial nerve (plantar flexion) dysfunction and show that this dysfunction is rapidly overcome, within 3 weeks, after injury. The third parameter, the toe spread angle, was significantly reduced at 5 versus 0 days for all three types of injury and did not recover to preoperative values within the 28-day observation period (Fig. 7G–I). We conclude that the toe spread angle estimates tibial and peroneal nerve functions, in contrast to the foot-base angles, and in particular, the control of toe movements. Also, this parameter appears to be more sensitive as it reveals functional deficits for longer time periods after injury compared with foot-base angles and functional indices.
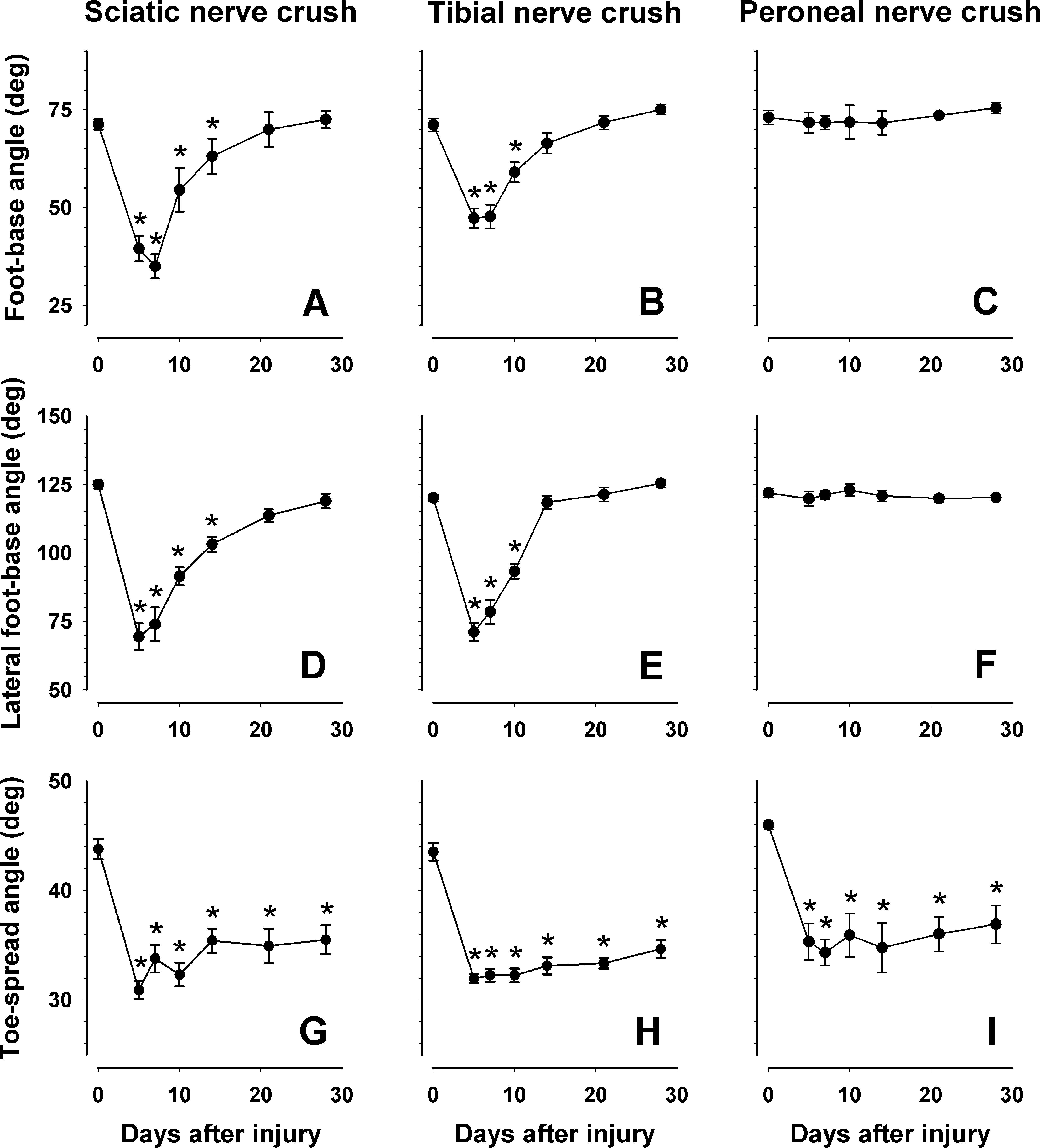
Beam-walking analysis. Comparison of different parameters (foot-base angle,
Ladder-climbing analysis
The C57BL/6J mice used for walking tests (experiment 2) were also analyzed during inclined ladder climbing. One of the parameters used, the ladder stance angle, showed significant alterations and lack of recovery 5 to 28 days after sciatic and tibial nerve crush but no changes after peroneal nerve injury (Fig. 8A–C). These findings were expected since this parameter was designed as a measure of plantar flexion ability. Compared with plantar flexion-related parameters for ground locomotion (foot-base angles for beam walking), which recovered to normal within 21 days after sciatic and tibial nerve injuries (Fig. 7A,B), the ladder stance angle was still significantly reduced 28 days after injury compared with the preoperative value (Fig. 8A,B). This difference between the beam and ladder parameters can be explained by a different load on the injured limb. Both foot-base angles are measured using frames in which the ipsilateral foot is at take-off position, a position at which body support is provided by the stance of the contralateral leg. In contrast, the ladder stance angle is measured at end-stance when the load is high and additionally increased as compared with stance during beam walking because of the up-right position of the body and, thus, reduced body-weight support by the forelimbs.
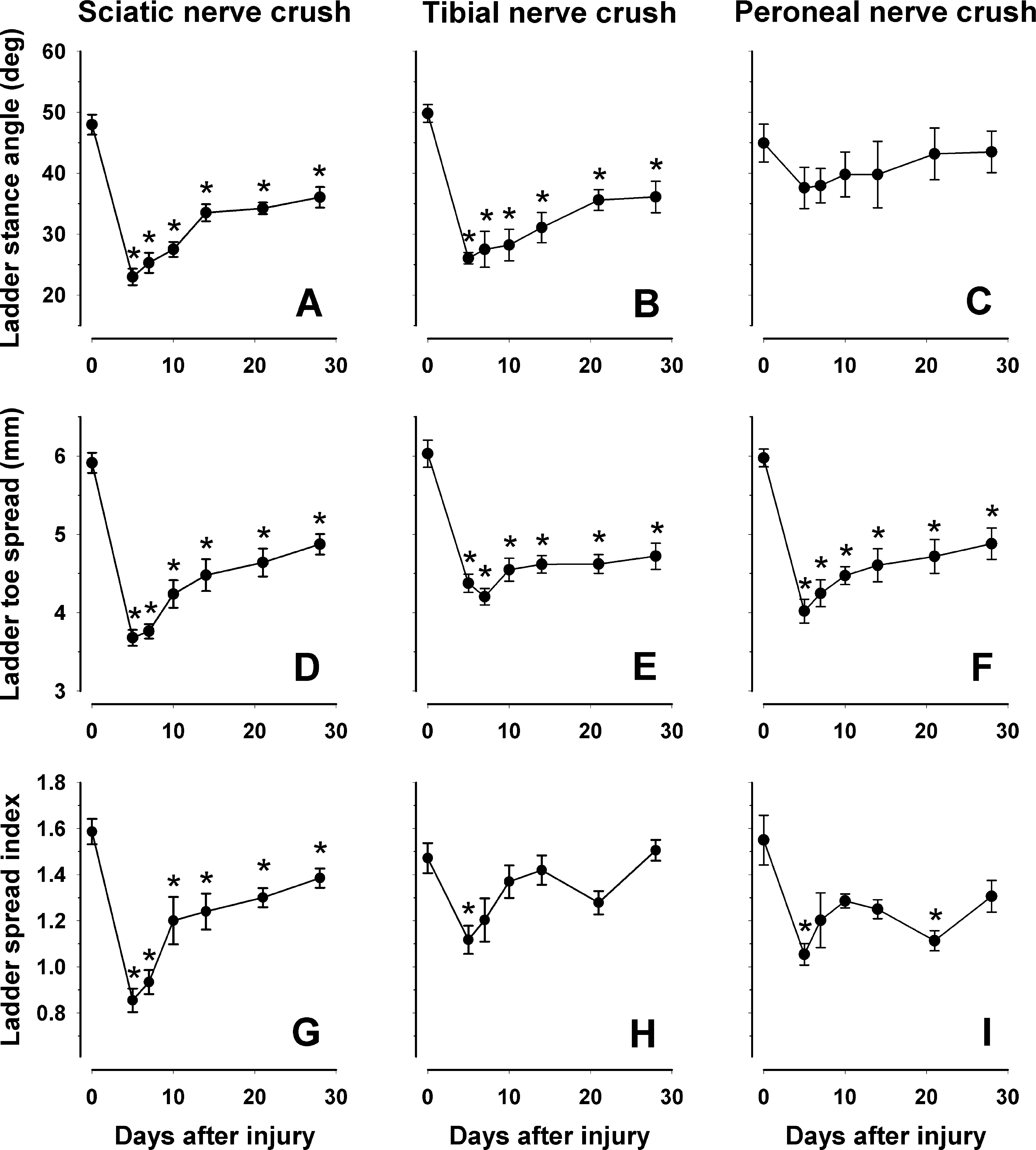
Ladder climbing analysis. Comparison of different parameters (ladder stance angle,
The second parameter, ladder toe spread, is a parameter expected to reveal dysfunction of both the tibial and the peroneal nerve. Consistent with this notion, we found a significant decline of the spread 5 days after sciatic, tibial, and peroneal nerve injuries compared with day 0 (Fig. 8D–F). Similar to the toe-spread angle estimated during beam walking (Fig. 7G–I), the ladder toe spread did not recover during the 28-day observation period (Fig. 8D–F). Significant decline and deficient recovery after sciatic nerve injury was also found for the ladder spread index, which, in contrast to other spread-related parameters, is evaluated during the end phase of a target-oriented movement (Fig. 8G). Since the same parameter changed less after tibial and peroneal nerve injury compared with sciatic nerve injury (Fig. 8H,I), we assume that both nerves contribute to the control of toe spreading prior to target reaching. In general, compared with ankle-related parameters (foot-base and ladder stance angles), the estimates of toe spreading revealed delayed recovery after nerve crush and allowed detection of peroneal dysfunction.
Worse functional outcome of sciatic nerve crush in NCAM-/- versus NCAM+/+ mice
In experiment 3, we analyzed NCAM-/- and NCAM+/+ mice over a 90-day period after sciatic nerve crush using the parameters employed in the short-term study of C57BL/6J mice described above. We pursued three goals. The first one was to reproduce the results of experiment 2 using the NCAM+/+ mice on an inbred C67BL/6J genetic background. At 28 days after sciatic nerve crush, complete functional recovery in NCAM+/+ mice was revealed by the sciatic functional index (Fig. 9A), the foot-base angle (Fig. 9B) and the lateral foot-base angle (not shown) while deficits were found for the toe spread angle (Fig. 9C), the ladder stance angle (Fig. 9D), the ladder toe spread (Fig. 9E), and the ladder spread index (Fig. 9F). Thus, for all parameters studied, the estimated outcome at 28 days after injury was similar to that in the first experiment (Figs. 6A; 7A,D,G; and 8A,D,G). These results indicate the reproducibility of the method in the context of this study.
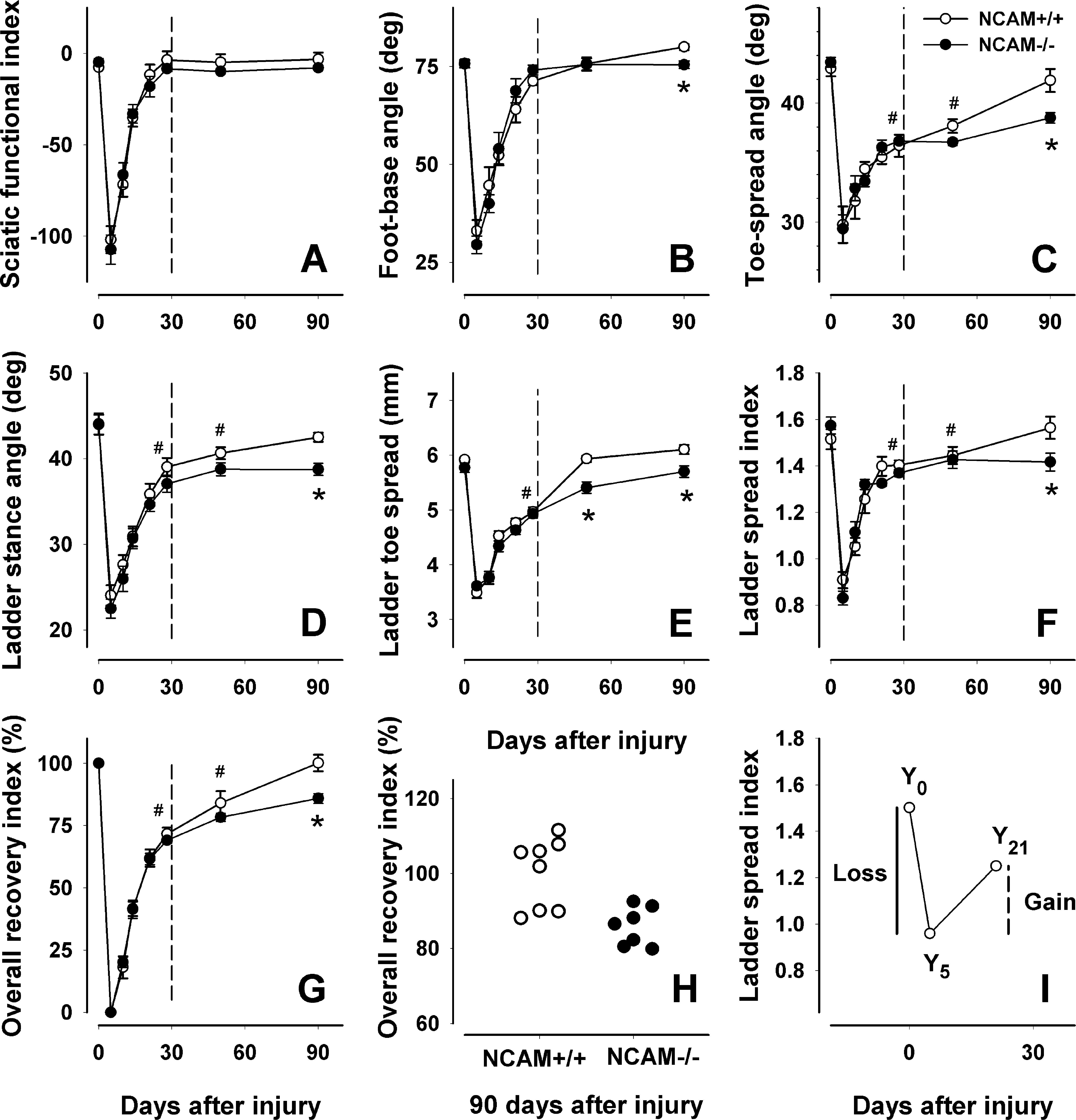
Worse outcome of sciatic nerve crush in NCAM-/- mice versus NCAM+/+ littermates. Shown are mean values ± SEM of sciatic functional indices (
The second aim was to elucidate if functional deficits persisting during the 28-day follow-up, as detected by some of our novel parameters, would disappear at longer time periods after injury. The analysis of NCAM+/+ mice showed complete recovery to normal (preoperative) values of all parameters studied at 90 days after crush (Fig. 9A–G). However, at 50 days, some parameters were still significantly different from the values at 0 (Fig. 9C,D,F). Thus, the general conclusion is that, expectedly, functional recovery after sciatic nerve crush is complete. However, complete recovery in mice requires much longer time, 3 months, than could be expected from previous results (Baptista et al., 2007; Boivin et al., 2007; Narciso et al., 2009; Yao et al., 1998).
Finally, we compared NCAM-/- and NCAM+/+ mice during the 90-day follow-up period. Comparisons of the values at 0 and 5 days did not show differences between the genotypes (Fig. 9A), indicating that NCAM-/- mice have normal gait prior to injury and are impaired to the same degree as NCAM+/+ mice after nerve crush. Using the sciatic functional index, we did not find differences in the time course and degree of recovery during the 90-day period between the genotypes (Fig. 9A), suggesting that NCAM-/- mice, similar to NCAM+/+ littermates, recover completely. Using our new parameters, we also could not detect genotype-related differences between 5 and 50 days (Fig. 9B–F) with the exception of the ladder toe spread at 50 days (Fig. 9E). However, the outcome at 90 days, as estimated by all novel parameters, was worse in NCAM-/- than NCAM+/+ mice (Fig. 9B–F). As indicated by the overall recovery indices, the NCAM+/+ mice recovered their motor abilities to normal (100%), while the degree of recovery in NCAM-/- mice only reached 85% (p < 0.05 compared with NCAM+/+ mice, t test, Fig. 9G,H). The conclusion from these results is that NCAM deficiency does not influence the initial phases of reinnervation and functional recovery but causes late deficits and, thus, less favorable outcomes 3 months after nerve crush injury compared with wild-type mice.
Discussion
The present study shows that footprint analysis is an insensitive method for assessment of sciatic nerve dysfunction in the mouse. As a better alternative, we propose a battery of video-based parameters, which, although unconventional in terms of kinematics, reflect specific aspects of motor incapability after damage of the sciatic nerve or its major branches (Table 1). The new method is sensitive, easily reproducible and quantitative, and allowed us to demonstrate that complete functional recovery after sciatic nerve crush in the mouse is achieved unexpectedly late, by 3 months after injury. We were also able to detect a small but significant deficit, as compared with wild-type mice, in the outcome of sciatic nerve lesion in mice deficient in the cell adhesion molecule NCAM, a molecule involved in a variety of processes ranging from neurite outgrowth to synaptogenesis and synaptic function (see below).
The claim that our method is more sensitive than footprint analysis requires a critical consideration. This statement is partially incorrect since we reached the same conclusion when we compared recovery after sciatic nerve crush in C57BL/6J mice using the sciatic functional index, on the one hand (Fig. 6A), and two of our parameters, the foot-base angle and the lateral foot-base angle, on the other hand (Fig. 7A,D), that is, complete recovery is achieved within 3 weeks after injury. The statement is, however, correct as far as novel parameters, in particular ladder parameters (Fig. 7A,D,G), showed incomplete recovery within 4 weeks. A reasonable explanation of what at first seems to be a contradiction can be provided if we consider the functions that individual parameters estimate. In our experiments, the sciatic functional index appeared to estimate tibial (plantar flexion), but not peroneal dysfunction (Fig. 6A–I). Similarly, the foot-base angles measured impairment of plantar flexion (Table 1). Thus, the conclusion based on three parameters is that impairment of plantar flexion during ground locomotion is overcome within 3 weeks after injury. However, plantar flexion during ladder climbing, as estimated by the ladder stance angle (Fig. 9D, Table 1), remained impaired for at least 50 days. Assuming that body weight load is greater in the stance phase used for measurements of the ladder stance angle than it is for the foot-base angles, we can conclude that the ability of full body weight support returns to normal late after nerve crush. Late recovery was also found using estimates of the toe spread, parameters detecting both peroneal and tibial nerve dysfunction (Table 1). The delayed, as compared with ankle extension, recovery of toe-related parameters could be due to the more distal position of most muscles controlling toe movements than muscles enabling ankle extension, and thus longer reinnervation times. However, this notion cannot explain delays of weeks. Therefore, we propose that the long-lasting deficits in toe-spread control reflect dysfunctions in sensory control of movement (i.e., in reflex pathways). This explanation is supported by the finding that, after initial recovery of motor and sensory functions, rats with sciatic nerve crush develop a neuropathic pain-like syndrome that appears to impede paw loading for at least 2 months after injury (Vogelaar et al., 2004). The general conclusion from these considerations is that the potential power of our method is not greater precision of measurements compared with walking track analysis but a more versatile analysis based on simultaneous use of different parameters estimated during different types of movement. This complex analysis also provides a more secure basis for conclusions drawn from a given experiment. The finding of significant differences between NCAM-/- and NCAM+/+ mice in five different parameters 90 days after injury is a strong argument in favor of the statement that the mutation has an effect on functional recovery.
Reproducibility is an essential requirement for a novel method. The only argument in favor of reproducibility here is the fact that we had similar findings in three groups of mice, C57BL/6J, NCAM-/-, and NCAM+/+ mice with sciatic nerve crush. We are, nevertheless, convinced that the method can be reproduced under different conditions in different laboratories. This optimism is based on our previous experience with single-frame motion analyses after femoral nerve and spinal cord injuries showing that precise definitions of parameters and their strict application are guarantees for reproducibility of results by different investigators. An example with respect to this notion in this study is the foot-base angle used also for functional analyses after femoral nerve injury. The preoperative values for three groups of mice (C57BL/6J, NCAM-/-, and NCAM+/+) are very similar to each other and to previously published values for intact C57BL/6J mice (Irintchev et al., 2005). Interestingly, while the foot-base angle increases after femoral nerve injury, sciatic nerve injury causes a decrease of the angle compared with uninjured mice. This difference can be explained by the fact that the same parameter estimates two different functions in the two injury paradigms, knee extension versus plantar flexion, respectively.
An important characteristic of the method described here is its simplicity. It is simple with respect to the equipment required. Although here we used a video camera producing 100 rather than the usual 25 frames per second and a motion analysis software for video capture, the minimum and sufficient requirements for single-frame motion analyses are a conventional camcorder, a computer, free software packages (VirtualDub and ImageTool), a self-made beam, and a self-made ladder (Apostolova et al., 2006; Irintchev et al., 2005). The method is also simple in the sense that no special knowledge is required. The only requirement is to strictly apply the criteria for selecting video frames and the definitions for angle and distance measurements. Not least, the approach relies on the performance of tasks (beam walking and ladder climbing) that are not stressful and do not require special training of the mice. The total time required for recording two to three beam trials and a similar number of ladder trails performed by one mouse is short, less than 5 min, and the time required for analyses is also reasonably short, roughly estimated to be 10 min for selection of frames and measurements of one parameter per mouse and time-point.
Based on the hypothesis that ablation of NCAM would have functional consequences, we compared recovery in NCAM-/- and NCAM+/+ mice. NCAM is involved in functionally important signal transduction mechanisms (Doherty et al., 1990; Niethammer et al., 2002; Schuch et al., 1989; Walmod et al., 2004; Walsh and Doherty, 1997) and promotes neurite outgrowth and fasciculation (Doherty et al., 1990; Rutishauser and Edelman, 1980), muscle innervation during development (Landmesser et al., 1988), endplate function in the adulthood (Rafuse et al., 2000), and upregulation in axons, Schwann cells, and muscle fibers after peripheral nerve injury (Covault et al., 1986; Daniloff et al., 1986; Olsen et al., 1995; Sanes et al., 1986). Considering this knowledge and the finding that in vivo application of polyclonal antibodies against NCAM delays endplate reinnervation in wild-type mice (Langenfeld-Oster et al., 1994), we expected to find delayed recovery in NCAM-/- compared with NCAM+/+ mice as a result of impaired axonal regrowth and synapse formation. We found, however, differences between NCAM-/- and NCAM+/+ mice were only detectable at the latest time-point studied, 90 days after crush. Thus, the absence of NCAM does not appear to impair the functional regeneration in the first 2 months after sciatic nerve crush, which can be attributed to redundancy (i.e., compensation by other molecules with partially similar functions) or to elimination of inhibitory NCAM-receptor interactions. Our conclusion that functional recovery after sciatic nerve crush in NCAM-/- mice is initially normal is in line with recent evidence that NCAM deficiency does not impair early synapse formation and muscle force recovery after tibial nerve crush (Chipman et al., 2010). The failure of complete recovery in NCAM-/- mice at 3 months after injury can be explained by instability of the newly formed endplates leading to a progressive development of synaptic dysfunction and axonal withdrawal from reinnervated endplates (Chipman et al., 2010). Thus, using the novel motion analysis approach, we could confirm conclusions on the impact of NCAM ablation on peripheral nerve regeneration in mice based on analysis using different methodology (morphological analyses, muscle tension recordings, endplate electrophysiology). Although in general good agreement, our results and these of Chipman and colleagues (2010) differ significantly with respect to the degree of functional impairment in NCAM-/- mice observed at 3 months after injury. While in vitro tension recordings have shown a dramatic decline of soleus muscle force in NCAM-/- mice, by approximately two-thirds compared with control values in NCAM+/+ mice, here we demonstrate only small differences, on average 15%, in degree of recovery between the genotypes. This quantitative difference does not necessarily reflect different sensitivities of the methods. Rather, we attribute the discrepancy to analysis of a single slow-twitch muscle in vitro using muscle tension recordings as opposed to performance of many different, fast- and slow-twitch muscles that may be affected to different degrees by the mutation in the limb of the freely moving animal. Finally, we must point out that our results also demonstrate the sensitivity of the new approach since significant differences between the genotypes 90 days after sciatic nerve crush were found using the novel parameters but not the sciatic functional index.
In conclusion, we believe that we have devised a method of functional analysis of sciatic nerve function in the mouse that will be useful for future research. The evidence provided in support of this notion and the simplicity of the method should encourage critical tests and applications of this approach by other researchers.
Footnotes
Acknowledgments
We are thankful to Fabio Morellini for providing NCAM-deficient mice. MS was supported by the New Jersey Commission for Spinal Cord Research.
Author Disclosure Statement
No competing financial interests exist.