
Abstract
Traumatic brain injury (TBI) remains the most common cause of death in persons under age 45 in the Western world. Recent evidence from animal studies suggests that supplementation with omega-3 fatty acids (O3FA) improves functional outcomes following focal neural injury. The purpose of this study is to determine the benefits of DHA supplementation following diffuse axonal injury in rats. Four groups of 10 adult male Sprague-Dawley rats were subjected to an impact acceleration injury and then received 30 days of supplementation with either 10 mg/kg/d or 40 mg/kg/d of docosahexaenoic acid (DHA). Serum fatty acid levels were determined from the isolated plasma phospholipids prior to injury and at the end of the 30 days of DHA supplementation. Following sacrifice, brainstem white matter tracts underwent fluorescent immunohistochemical processing for labeling of β-amyloid precursor protein (APP), a marker of axonal injury. Dietary supplementation with either 10 mg/kg/d or 40 mg/kg/d of DHA for 30 days results in significantly (p < 0.05) increased DHA serum levels of 123% and 175% over baseline, respectively. Immunohistochemical analysis reveals significantly (p < 0.05) decreased numbers of APP-positive axons in animals receiving dietary supplementation with DHA, 26.1 (SD 5.3) for 10 mg/kg/d, and 19.6 (SD 4.7) for 40 mg/kg/d axons per mm2, versus 147.7 (SD 7.1) axons in unsupplemented animals. Sham-injured animals had 6.4 (SD 13.9) APP positive axons per mm2. Dietary supplementation with DHA increases serum levels in a dose-dependent manner. DHA supplementation significantly reduces the number of APP-positive axons at 30 days post-injury, to levels similar to seen those in uninjured animals. DHA is safe, affordable, and readily available worldwide to potentially reduce the burden of TBI.
Introduction
T
While various studies have given hope that pharmacological or physiological interventions might reduce axonal injury, to date over 30 major clinical studies have failed to show a significant effect in reducing morbidity and mortality or improving outcomes of TBI (Doppenberg et al., 1997). Advances have been made in the treatment strategies for the clinical management of the acute injury, and critical care and rehabilitation medicine have greatly improved both survival and functional outcomes. The science of nutritional supplementation has undergone significant changes, from the early goals of simply delivering necessary calories, to current regimens that provide specific amino acid and fatty acid combinations to maximize the healing process. Docosahexaenoic acid (DHA), the principal constituent of omega-3 fatty acids (O3FA) of neural tissue, has demonstrated significant anti-inflammatory properties, and is the precursor of neuroprotection-1. Recent evidence from animal studies suggests that supplementation with O3FA (particularly eicosapentaenoic acid [EPA] and DHA) improves functional outcomes following focal neural injury (Wu et al., 2004, 2007; Mills et al., E-pub ahead of print). Additional recent evidence from animal models of ischemia demonstrate improvements in outcome and reductions in infarct volume with DHA supplementation (Belayev, 2009). We hypothesized that DHA supplementation following diffuse axonal injury in rats would ameliorate secondary mechanisms of injury, and result in a decreased number of injured axons, as measured using immunohistochemical analysis of β-amyloid precursor protein (APP)-positive axons.
Methods
Marmarou impact acceleration injury model in rats
Four groups of 10 (n = 40) of adult male Sprague-Dawley rats were subjected to a Marmarou impact acceleration injury resulting in reproducible TBI. Rats weighing 350–400 g received isoflurane-induced anesthesia, and were subsequently maintained on inhaled isoflurane using a modified medical anesthesia machine. Lidocaine 1% local anesthetic was injected subcutaneously along the planned incision site. Buprenorphine was used for post-operative analgesia. Body temperature was controlled during the approximately 10-min procedure using a heating blanket, and adequate sedation was confirmed by evaluation of the response to heel tendon pinch. The animals were shaved and prepared in sterile fashion for surgery, followed by subcutaneous injection of local anesthetic into the planned incision site. A 3-cm midline incision in the scalp was made, and the periosteal membranes were separated, exposing the bregma and the lambda. A metal disk 10 mm in diameter and 3 mm thick was attached to the skull with cyanoacrylate centered between the bregma and the lambda. The animal was placed prone on a foam bed with the metal disk directly under an acrylic glass tube. A 450-g brass weight was dropped a single time through the tube from a height of 2 m, striking the disk. The animal was then ventilated on 100% oxygen while the skull was inspected, the disk removed, and the incision repaired. When the animal recovered spontaneous respirations, anesthesia was discontinued and the animal was returned to its cage for post-operative observation (Marmarou et al., 1994). All procedures involving live animals were approved by the Institutional Animal Care and Use Committee of West Virginia University, and were performed according to the principles of the Guide for the Care and Use of Laboratory Animals, published by the Institute of Laboratory Resources, National Research Council (National Institutes of Health publication 85-23-2985).
DHA supplementation and serum level monitoring
The four groups received dietary supplementation with DHA (Martek Inc., Columbia, MD) for 30 days starting on post-injury day 1. Two of the four groups received dietary supplementation with DHA daily in the following amounts: group 1, 10 mg/kg/d, and group 2, 40 mg/kg/d. These dosages were selected based on our previous studies of O3FA supplementation, and reflect typical human equivalent dosing. Group 3 served as an unsupplemented control, and group 4 underwent sham injury and received no supplementation. Each group received rat chow ad libitum and were housed in the small-animal vivarium under veterinary staff supervision. Fatty acid blood testing was performed prior to injury and at the end of the 30 days of DHA supplementation by analyzing the isolated serum phospholipids (including DHA, EPA, and arachidonic acid) from 50-μL blood samples using a previously described method (Holub and Skeaff, 1987) (Nutrasource Diagnostics, University of Guelph, Guelph, Ontario, Canada).
Tissue preparation and immunohistochemical labeling
Following 30-day post-injury survival, the animals were euthanized with a lethal dose of 0.5 mL ketamine and 0.5 mL xylazine. The animals were immediately perfused transcardially with 200 mL cold 0.9% saline to wash out all blood. This was followed by 4% paraformaldehyde infusion in Millonig's buffer for 40 mins. The entire brain, brainstem, and rostral spinal cord were removed and immediately placed in 4% paraformaldehyde for 24 h. Following the 24-h fixation period, the brain was blocked by cutting the brainstem above the pons, cutting the cerebellar peduncles, and making sagittal cuts lateral to the pyramids. The resulting tissue containing the corticospinal tracts and the medial lemnisci, areas previously shown to yield traumatically-injured axons, was then sagittally cut on a vibratome into 50-μm-thick sections. The tissues underwent temperature-controlled microwave antigen retrieval using previously described techniques (Stone et al., 1999). The tissues were pre-incubated in a solution containing 10% normal serum and 0.2% Triton X in PBS for 40 min.
For APP labeling, the tissue was incubated in polyclonal antibody raised in rabbit against APP (#51-2700; Zymed Laboratories, Carlsbad, CA) at a dilution of 1:200 in 1% NGS in PBS overnight. Following incubation in primary antibody, the tissue was washed three times in 1% NGS in PBS, then incubated in a secondary anti-rabbit IgG antibody conjugated with Alexa 488 fluorophore (A11008; Molecular Probes, Eugene, OR), diluted at 1:200 for 2 h. The tissue underwent a final wash in 0.1 M phosphate buffer, and then was mounted using an anti-fade agent and cover-slipped. The slides were sealed with acrylic and stored in the dark in a laboratory refrigerator (Mills et al., 2003).
Fluorescent microscopy and image analysis
The tissue was examined and images were acquired using a laser scanning confocal microscope system with an argon 488 excitation laser and a 40 × objective lens. Ten digital images were obtained from the tissue of each animal, and images were then randomized. Individual injured axons were independently counted and the data were stored in a spreadsheet. The counts were converted to density per square millimeter with the formula axon count per image/image area. Differences between group means were determined using paired t-tests and considered significant if the probability value was less than 0.05.
Stereological quantification of axonal injury
A stereological method was used to determine an unbiased estimate of the number of APP-positive axons per cubic millimeter in the corticospinal tract and medial lemniscus. The optical fractionator technique utilitizing StereoInvestigator 7.0 (MBF Bioscience, Williston, VT) and a Nikon Eclipse (Nikon, Melville, NY) microscope with 4 × and 20 × objectives was performed. Sagittal APP-stained specimens were examined with low magnification and regions of interest (ROI) were drawn incorporating the corticospinal tract and medial lemniscus. The software then selected random 50-μm2 counting frames with depth of 15 μm, and APP-positive axons were marked. The volume of the ROI was determined using the Cavalieri method, the volume of the sum of the counting frames was calculated, the sum total of injured axons within the counting frames was calculated, and an estimate of the number of APP-positive axons per cubic millimeter was calculated.
Results
Impact acceleration model and serum fatty acid levels
The mortality rate in this model of TAI was 0%. Animals tolerated daily oral supplementation without any observed untoward effects.
Supplementation for 30 days after the brain trauma with DHA at a dosage of either 10 mg/kg/d or 40 mg/kg/d resulted in increased levels of serum DHA of 123% and 176% over initial levels, respectively (Fig. 1). Animals receiving no supplementation had a 7% decrease in DHA. Serum EPA levels likewise increased in supplemented animals 104% and 313%, respectively, for the 10-mg and 40-mg doses, while unsupplemented animals showed a 59% decrease. The arachidonic acid:eicosapentaenoic acid ratio (AA:EPA), a marker of inflammation, decreased 72% and 109%, respectively, in the 10-mg and 40-mg groups, while it increased 65% in unsupplemented animals.
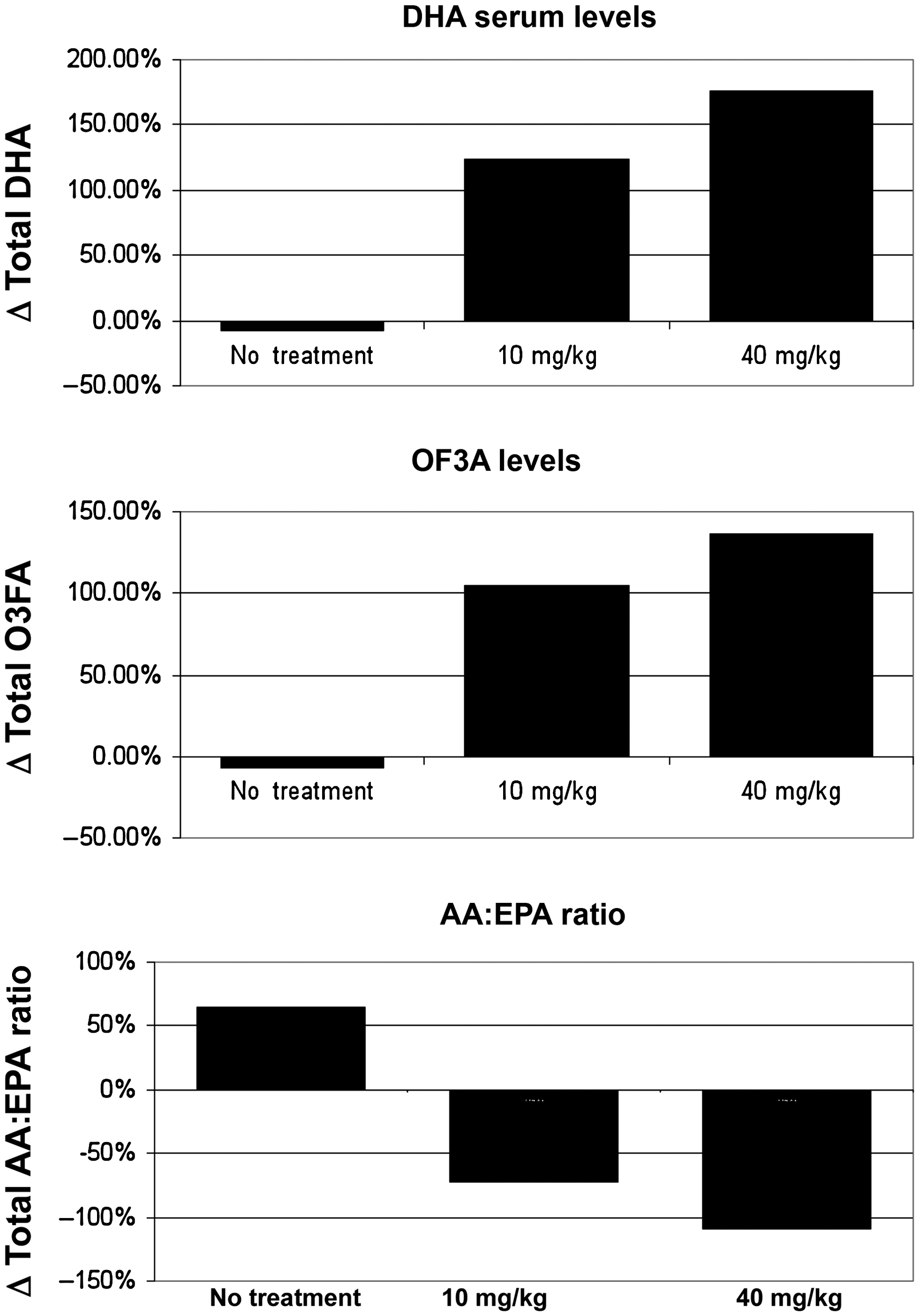
Oral supplementation with either 10 mg/kg/d or 40 mg/kg/d of DHA for 30 days increased serum levels of docosahexaenoic acid (DHA) and total omega-3 fatty acids (O3FA). The arachidonic acid:eicosapentaenoic acid ratio (AA:EPA), a marker of systemic inflammation, was significantly lower in animals receiving supplementation than in unsupplemented animals.
Immunohistochemical analysis of APP-positive axons
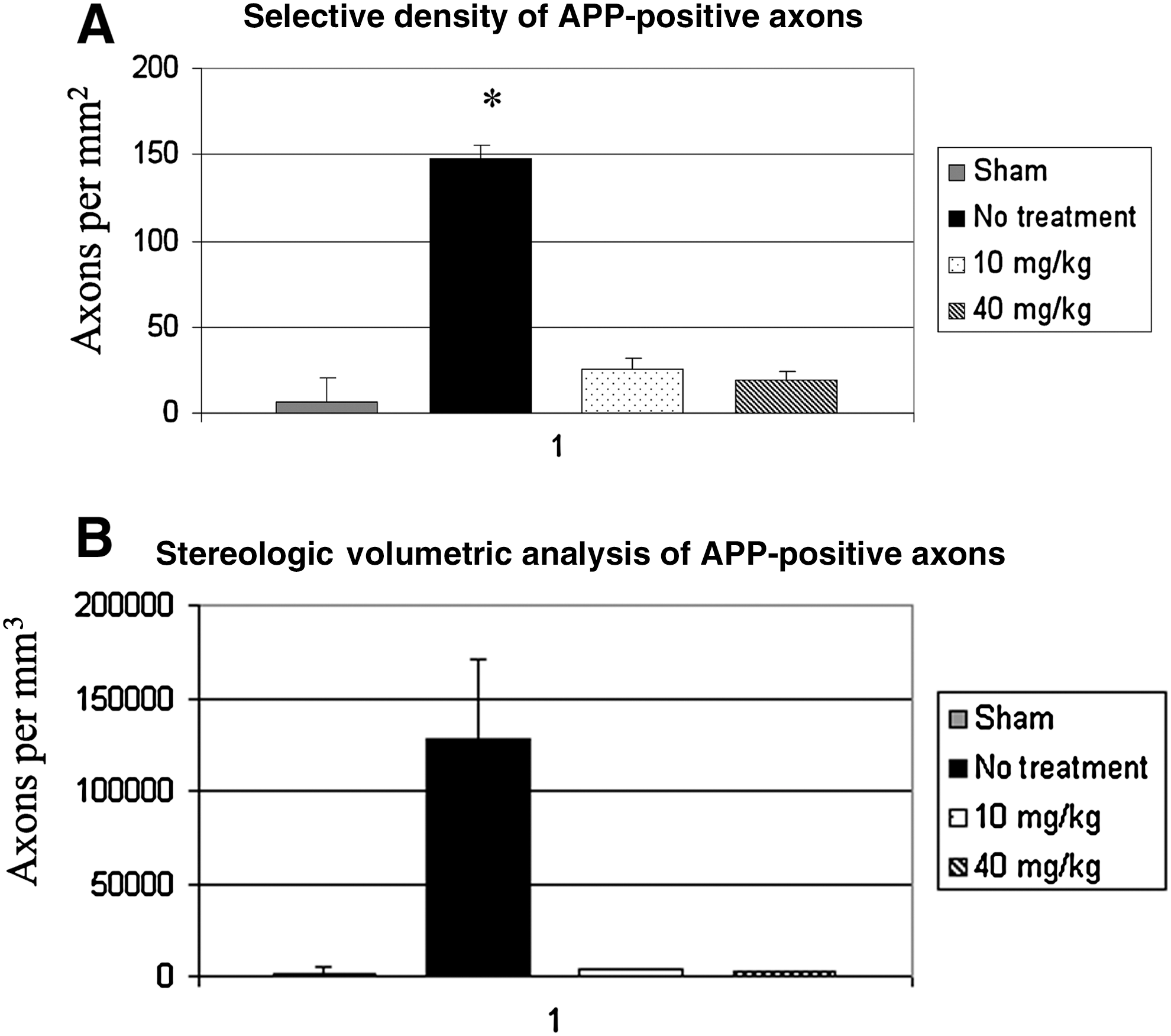
In sham-injured animals, axons throughout the medullary corticospinal tract and medial lemnsci demonstrated a paucity of labeling for APP. These rare labeled axons did not demonstrate vacuolization, swelling, or breakdown, characteristics typical of TAI (Fig. 2). In comparison, evaluation of axons of animals receiving no supplementation at 30 days post-injury demonstrated focal labeling of APP within swollen contiguous and terminal axon segments, consistent with previous findings suggestive of impaired axoplasmic transport in TAI. Following microscopic digital image acquisition from multiple areas within the corticospinal tract and medial lemnsci from multiple tissue slices, counting of APP-positive axons was performed, and the results were converted to density per square millimeter. This demonstrated a significant quantitative difference of 147.7 (SD 7.1) axons in unsupplemented animals, versus sham-injured animals, which had 6.4 (SD 13.9) APP-positive axons/mm2 (Fig. 3).

(
(
In animals receiving either 10 mg/kg/d or 40 mg/kg/d of DHA, axons throughout the corticospinal tract and medial lemnisci demonstrated only rare APP-positive axons, similarly to sham-injured animals. However, in comparison to sham-injured animals, the rare APP-positive axons were more likely to demonstrate morphologic characteristics of injury, primarily swelling and disconnection. Quantitative analysis revealed significantly (p < 0.05) decreased numbers of APP-positive axons in animals receiving dietary supplementation with DHA, 26.1 (SD 5.3) and 19.6 (SD 4.7) axons/mm2 respectively, in the 10-mg and 40-mg groups, versus 147.7 (SD 7.1) axons/mm2 in unsupplemented animals.
Immunohistochemical analysis of caspases-3-positive axons
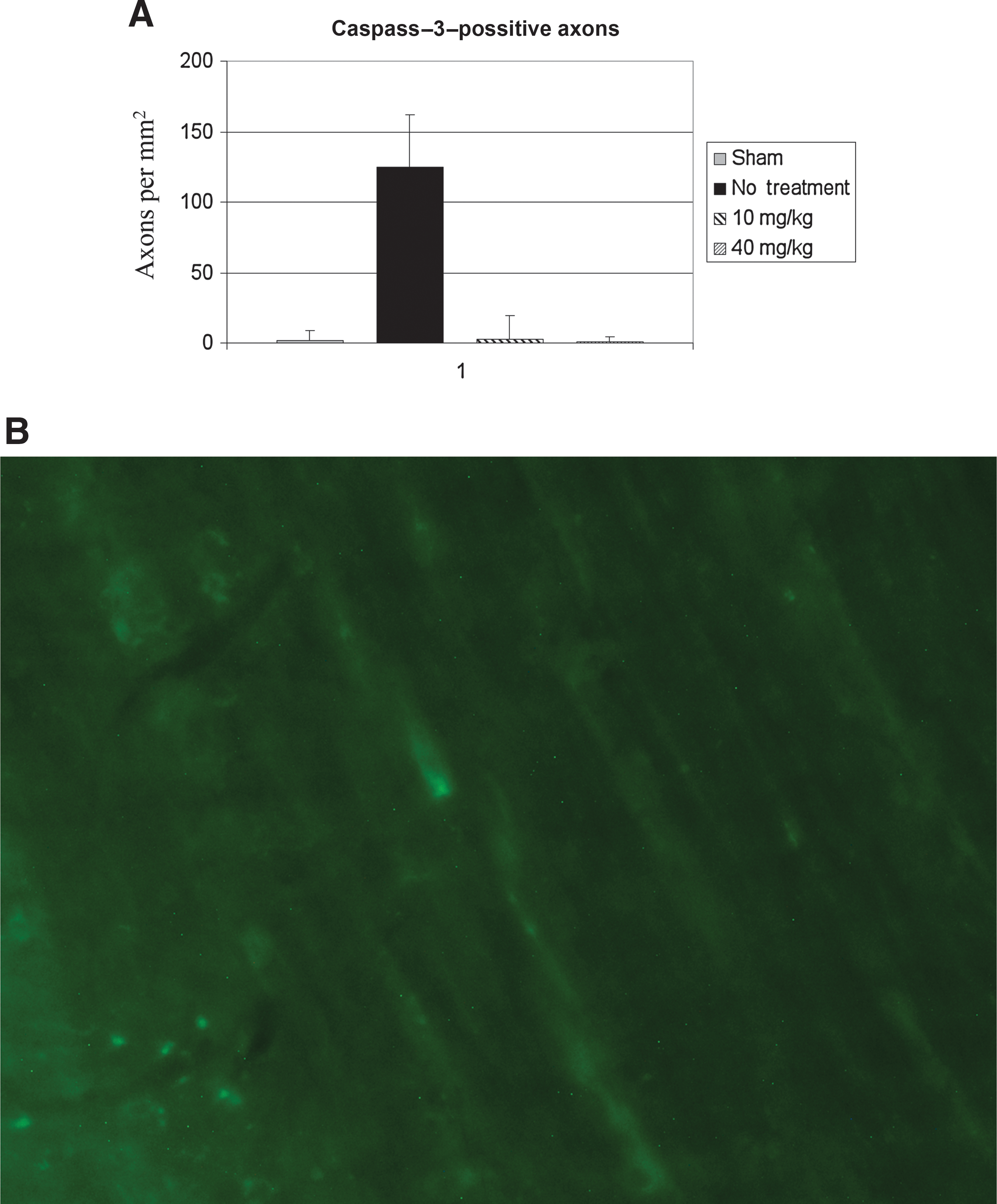
Following microscopic digital image acquisition from multiple areas within the corticospinal tract and medial lemnsci from multiple tissue slices, counting of caspases-3-positive axons was performed, and the results were converted to density per square millimeter. This demonstrated a significant quantitative difference of 124.9 (SD 37.0) axons in unsupplemented animals, versus sham injured animals, which had 1.7 (SD 7.5) caspases-3-positive axons/mm2 (Fig. 4). In animals receiving either 10 mg/kg/d or 40 mg/kg/d of DHA, axons throughout the corticospinal tract and medial lemnisci demonstrated only rare caspases-3-positive axons, similarly to sham-injured animals. Quantitative analysis revealed significantly (p < 0.05) decreased numbers of caspases-3-positive axons in animals receiving dietary supplementation with DHA, with 2.8 (SD 1.7) and 0.9 (SD 3.7) axons/mm2 respectively, in the 10-mg and 40-mg groups, versus 124.9 (SD 37.0) axons/mm2 in unsupplemented animals.
(
Stereologic analysis of APP-positive axons
The stereologic optical fractionator method was utilized to determine an unbiased estimate of the number of APP-positive axons per cubic millimeter within the corticospinal tract and medial lemnsicus. This demonstrated a significant quantitative difference of 128,048 (SD 42,206) in unsupplemented animals, versus sham-injured animals, which had 1648 (SD 1847) APP-positive axons/mm3 (Fig. 4). In animals receiving either 10 mg/kg/d or 40 mg/kg/d of DHA, stereologic analysis revealed significantly (p < 0.05) decreased numbers of APP-positive axons, with 3720 (SD 1924) and 2892 (SD 3221) axons/mm3, respectively, versus 128,048 (SD 42,206) axons in unsupplemented animals.
Discussion
This study demonstrates that DHA alone, given after TAI, is neuroprotective. Oral supplementation with either 10 mg/kg/d or 40 mg/kg/d of algae-derived DHA for 30 days following an impact acceleration injury resulted in significantly decreased numbers of injured axons as measured by APP staining. Likewise, a significant decrease in active caspase-3-positive axons provides additional evidence for the neuroprotective and injury-ameliorating effects of DHA.
Serum response
Analysis of the serum phospholipids at the end of the supplementation period after the traumatic insult showed a dose-response effect by an increase in the total of both EPA and DHA levels compared to sham animals, whereas there was a decrease in the combined EPA and DHA serum levels in the non-supplemented animals. These results are consistent with those of previous studies in both rats and humans, that demonstrated uptake and retroconversion of DHA into EPA (Brossard et al., 1996; Vidgren et al., 1997). Interestingly, there was an increase in serum AA levels in the non-supplemented group, although a non-significant increase was seen in the supplemented animals. As a result, the AA:EPA ratio, an indicator of systemic inflammation, was significantly increased in the non-supplemented animals compared to the supplemented animals. This would have the effect of activating leukocytes into neutrophils and macrophages that could more easily enter into the brain. Dosage with 10 mg/kg/d or 40 mg/kg/d reflect typical human equivalent DHA supplemental dosing with between 1 and 3 g/d.
Mechanisms of traumatic brain injury
Conventional theory has held that TAI involves immediate axonal tearing through the direct action of forces associated with the traumatic insult. More recently, experiments employing anterograde tracers have revealed that TAI is a progressive event, involving a focal impairment of axoplasmic transport, leading to axonal swelling and ultimately disconnection following TBI (Raghupathi et al., 2000). The cellular mechanisms of injury include mitochondrial disruption, loss of calcium homeostasis, and activation of apoptotic cascades (Buki et al., 2000; Eldadah and Faden, 2000; Sullivan et al., 2005; Wang et al., 1999). Similar changes may occur with spinal cord injury (Lang-Lazdunski et al., 2003; Murphy et al., 1994; Xu et al., 2004).
The concept of the secondary phase of TBI includes what are now recognized as ongoing abnormalities in glucose utilization and cellular metabolism, as well as membrane fluidity, synaptic function, and structural integrity (Aoyama et al., 2008; Hovda, 2007). In general, axon membranes are injured, leakage occurs, and axonal transport is interrupted, in a progressive process. This concept is reinforced by recent autopsy findings in professional athletes participating in contact sports, showing multi-focal areas of damaged neurons and their processes that were remarkable for tau antibody staining, representing numerous times and differing regions of injury from multiple concussions (Omalu et al., 2005, 2006).
Omega-3 fatty acids
The primary O3FAs are EPA and DHA. Prior work from our lab with the same head-injury model has shown that dietary supplementation with a fish oil concentrate rich in EPA and DHA significantly reduces the number of injured axons (Mills et al., E-pub ahead of print). The current experiment is the first to analyze the effects of immediate post-injury treatment with an algae-derived DHA-only dietary additive. DHA constitutes the primary O3FA in brain synaptosomal plasma membranes and synaptic vesicles. Human brain O3FA content consists of 97% DHA, while the retina contains 93% (Martinez, 1992). Neuronal membranes are composed largely of phospholipids, and DHA stimulates phosphatidylserine and phosphatidylethanolamine production, as well as neurite outgrowth triggered by nerve growth factor (Cao et al., 2004). During fetal development and the first few years of life, the DHA content of the human brain exponentially rises, while dietary deficiency of DHA causes the brain to retain DHA longer when other organs are depleted. Unlike AA, DHA appears to be retained by neuronal membrane phospholipids, as opposed to astroglial cells, which readily release it, probably reflecting a trophic or anti-apototic effect important for neuronal survival (Carver et al., 2001; Martinez 1994; Salem et al., 2001). TBI can cause acute and long-lasting disturbances in brain phospholipid metabolism, and degradation of neuronal and astrocytic membrane phospholipids has been documented (Wu et al., 2004), thus contributing to the evolving secondary injury.
Neuronal and cellular response to traumatic brain injury
Modulation of the inflammatory cascade by O3FA has been proposed as a critical neuroprotective mechanism. Arachidonic acid, a primary omega-6 fatty acid in the plasmalemma, is metabolized by cyclooxygenase (COX) and lipoxygenase (LOX) enzymes to pro-inflammatory eicosanoids, such as the two-series prostaglandins and thromboxanes, prostaglandin E2 and leukotriene B4 (Needleman et al., 1979). These eicosanoids enhance vascular permeability, increase local blood flow, increase infiltration of leukocytes, and enhance production of proinflammatory cytokines such as tumor necrosis factor-α, interleukin-1 (IL-1), and IL-6. In contrast, O3FA can decrease COX activity and inhibit the formation of proinflammatory eicosanoids and cytokines (Calder, 2003; Lonergan et al., 2004).
Several mechanisms have been proposed to explain how O3FA may play a neuroprotective role, including reductions in excitotoxicity, modulation of calcium and potassium channels, activation of gene transcription, and formation of neuroprotectin-1 and resolvins. In various ischemia models, it has been demonstrated that DHA has a neuroprotective effect on hippocampal injury, by reduction of lipid oxidative damage through inhibition of prostaglandin synthesis, a protective mechanism against glutamate-induced neurotoxicity (Hogyes et al., 2003; Wang et al., 2003), and a reduction of the reactive oxygen species seen in TBI that reduce levels of brain-derived growth factor (BDGF), synapsin I, and cAMP responsive element-binding protein (CREB), which unchecked can result in synaptic dysfunction and cognitive impairment (44). The neuroprotective effect of DHA may involve activation of gene transcription through retinoid receptor signaling (Jump, 2002), and the formation of mediators such as 10,17S-docosatriene (neuroprotection-D1), an endogenous compound with antioxidant (Bazan, 2005; Sarsilmaz et al., 2003), as well as anti-inflammatory (Mukherjee et al., 2004) effects.
O3FA have significant affinity for two-pore potassium channels, such as TWIK-related potassium channel (TREK), and TWIK-related arachidonic acid-stimulated potassium channel (TRAAK; Emsley et al., 2003; Lauritzen et al., 2000). Both EPA and DHA have also been demonstrated to inhibit calcium channels (Danthi et al., 2005). This could potentially avert apoptosis of damaged neurons and their projecting fibers. EPA increases the levels of resolvins, thus further decreasing the intensity of the inflammatory process (Schwab et al., 2007; Dusart and Schwab, 1994). A recent finding shows reduction in the levels of silent information 2 (Sir2) and energy metabolic markers following O3FA supplementation. Sir2 is believed to have neuroprotective properties in a traumatically stressed environment by reducing oxidative stress in the hippocampus. Sir2 can detoxify reactive oxygen species and modulate brain energy metabolism to ensure optimal neuronal survival (Wu et al., 2007; Songur et al., 2004). In hippocampal neurons, DHA has been shown to have neurite growth-promoting effects (Calderon and Kim, 2004).
Nutritional supplementation
Emerging modern research indicates a potentially greater role for dietary supplementation for treating patients with MTBI. Several mechanisms, such as those highlighted above, have been proposed as the mechanisms of action of O3FA, all potentially interacting at the cellular level and reducing the effects of traumatic insults on neurons and their projections. The findings of this study, that DHA alone may have equally dramatic results as those seen in our prior work utilizing fish oil concentrate containing both EPA and DHA, have several implications. Among the most important are that DHA alone may have sufficient properties to provide significant amelioration of effects of TBI, including the additional finding that retroconversion from DHA to EPA occurs. likely contributing to the beneficial effects of the former. This may provide several advantages, including that DHA may be obtained from an algal source, it can be produced in limitless supply in a controlled laboratory environment, and it is known to be free of possible contamination. Compared to marine sources, there is elimination of many issues, including procurement and purity.
The numerous proposed mechanisms of action of O3FA demonstrate that the biochemical pathways may either be unknown or multiple in their effects of reducing traumatic axonal damage. However, the most likely manner in which these positive effects are mediated appears to involve stabilization of the cellular environment, to reduce reactive oxygen species, to modulate continued energy production, and to promote axonal injury and repair mechanisms. Our findings are consistent with previous research that has demonstrated that O3FA are protective against cellular injury (Wu et al., 2007; Blondeau et al., 2002).
O3FA preparations have previously been shown to be safe and well tolerated in patients in several disease states (Schlanger et al., 2002). Further research is needed to ascertain the exact mechanisms, dosages, and preferred administration schedule for the prevention of mTBI, including a pre-treatment protocol that is currently underway in our laboratory. DHA concentrates like those used in this experiment are derived from algal sources, and are commercially available for clinical use to treat TBI. Administration of DHA after brain trauma as a neuroprotective and injury-ameliorating treatment deserves consideration and further clinical investigation as a promising, inexpensive, and innovative new approach in TBI management.
Conclusion
Dietary supplementation with DHA increases serum levels of these same fatty acids. DHA supplementation significantly reduces the number of APP-positive axons at 30-days post-injury to levels similar to those seen in uninjured animals. DHA is safe, affordable, and readily available worldwide to potentially reduce the burden of traumatic brain injury.
Footnotes
Acknowledgments
This work was supported by a grant from Martek, Inc. The authors would like to recognize Rebecca Kessler for assistance with manuscript preparation.
Author Disclosure Statement
This work was financially supported by Martek Biosciences, Inc., for which Dr. Bailes serves as a consultant.