
Abstract
Neurotrophin-3 (NT3) is an important neurotrophic factor for spinal cord injury (SCI) repair. However, constant exchange of cerebrospinal fluid often decreases the effective dosage of NT3 at the targeted injury site. In the present study, a recombinant collagen-binding NT3 (CBD-NT3), consisting of a collagen-binding domain (CBD) and native NT3, was constructed. Linear rat-tail collagen (LRTC) was used as a physical carrier for CBD-NT3 to construct a LRTC/C3 system. The collagen-binding ability of CBD-NT3 was verified, and the bioactivity of CBD-NT3 was assayed with neurite outgrowth of dorsal root ganglia (DRG) explants and DRG cells in vitro. After complete spinal cord transection in rats, LRTC/CBD-NT3 or the LRTC/NT3 system was transplanted into the injury site. Hindlimb locomotion recovery was closely observed using the Basso-Beattie-Bresnahan (BBB) locomotor rating scale and the grid walk test. Significant improvement was observed in the LRTC/CBD-NT3 group. The results of regenerating nerve fiber and anterograde tracing of biotinylated dextran amine (BDA)-labeled corticospinal tract (CST) fibers demonstrated axonal regeneration of LRTC/CBD-NT3 in the injured spinal cord. Serotonin fiber regrowth also illustrated the effectiveness of LRTC/CBD-NT3. Thus, collagen-binding NT3 with LRTC may provide an effective method for treating SCI.
Introduction
R
Collagen could serve as a physical bridge to connect the rostral and caudal portions of the injured spinal cord, and could be used as a vehicle to deliver NT3. In previous work, we made a linear collagen material that retained mechanical strength and fiber organization (Lin et al., 2006). In this study we explored another type of fibrous collagen.
Traditionally, the binding affinity between collagen and NT3 has been low, and it usually takes a large amount of NT3 to achieve a satisfactory therapeutic effect. Such high doses may be unsafe, and diffusion of NT3 into other tissues may cause adverse effects. To increase the binding ability of NT3 to collagen, we constructed a recombinant fusion protein consisting of NT3 and a collagen-binding domain (CBD). The CBD domain was a polypeptide TKKTLRT, which demonstrated an ability to bind to type I collagen in a study by De Souza and associates (1994). We hypothesized that the combination of the bioactive collagen-binding NT3 and a collagen scaffold could guide axonal regeneration and retain the growth factor at the site of SCI, and effectively promote axonal regeneration and locomotor functional recovery.
Methods
Plasmid construction and protein production
Total cDNA of human fetus brain was used as the template for amplifying the native human NT3 by PCR. A coding sequence for the collagen-binding domain of TKKTLRT was linked to NT3, constructing the coding sequence for CBD-NT3 (Fig. 1A). The coding sequence for NT3 or CBD-NT3 was inserted into pET-28a. Either NT3 or CBD-NT3 was induced with 1 mM isopropyl β-D-thiogalactopyranoside (IPTG) in the BL21 strain of E. coli at 37°C for 5 h. The recombinant protein of NT3 or CBD-NT3 was accumulated in the inclusion body and solubilized in 8 M urea containing 0.4% β-mercaptoethanol. After refolding, the protein containing His-tag was purified with nickel chelate chromatography (Amersham Biosciences, Piscataway, NJ). The expression and purification of NT3 and CBD-NT3 were determined by SDS-PAGE and Western blot, respectively.

(
Preparation of linear rat-tail collagen
The collagen used in our study was made from the white aponeurosis of rat tail. The residual muscles, connective tissues, and fat were all removed, and the collagen was washed repeatedly in distilled water. They were further treated with 1% tri(n-butyl)phosphate (TnBP; Sigma-Aldrich, Munich, Germany) in 50 mM Tris-HCl buffer (pH 8.0) at 4°C for 2 days to remove the cellular components, and extracted in 50 mM Tris-HCl buffer (pH 8.0, 1 M NaCl) at 4°C for 6 h to remove soluble proteins and other cellular elements. Then the samples were rinsed in distilled water several times in order to remove the residual agents, then they were freeze-dried.
Collagen-binding assay
The binding ability of CBD-NT3 to collagen was measured by a modified ELISA assay. The type I collagen prepared from rat tail tendon was added to a 96-well plate. The plate was blocked with fetal bovine serum for 2 h at room temperature. Serial dilutions of NT3 and CBD-NT3 were added to the wells and incubated at 37°C for 2 h, then unbound proteins were removed by washing extensively with PBS. Anti-polyhistidine monoclonal antibody (Sigma-Aldrich, St. Louis, MO) was used as the primary antibody, and alkaline phosphatase (ALP)-conjugated goat-anti-mouse IgG (Sigma-Aldrich) was used as the secondary antibody. The remaining proteins were detected by 2 mg/mL para-nitrophenylphosphate (pNPP; Sigma-Aldrich) in ALP buffer (100 mM Tris-HCl, 100 mM NaCl, and 10 mM MgCl2, pH 9.6) for 30 min at 37°C. The results were quantified at 405 nm using an ELISA reader (Molecular Devices, Sunnyvale, CA). A calibration curve was constructed for each sample to quantitate the total amounts of proteins retained by the collagen. The results were analyzed on a Scatchard plot, and the Kd of each protein was determined using the least-square method.
Biological activity of the proteins
E15 rat dorsal root ganglia (DRG) explants and DRG cells were utilized for detecting the biological activity of the renatured NT3 and CBD-NT3. DRG cells were obtained from E15 rat embryos and digested with 0.08% trypsin (Invitrogen, Carlsbad, CA) for 15 min. Three DRG explants/well, and cells at a density of 105 DRG cells/well were cultured in serum-free DMEM/F12 medium (HyClone, Rochester, NY), supplemented with N2 (Sigma-Aldrich) in polylysine-coated 48-well plates (Costar; Corning, Corning, NY). After stimulation with 0.01, 0.1, or 1 μM renatured NT3 or CBD-NT3, the cells were maintained for 3 days, immunostained with β-tubulin type III (monoclonal TuJ1, 1:500; Sigma-Aldrich) as the primary antibody for neurons at 4°C overnight, and then they were incubated with a secondary antibody, anti-mouse IgG, and Hoechst 33342 (1:500; Sigma-Aldrich) to stain the nuclei. The number of surviving neurons and the length of the neurites were calculated. The other cells were cultured under identical conditions, except NT3 and CBD-NT3 served as a control. All conditions were established in triplicate. The assays were assessed in duplicate by two independent assessors.
Bioactivity assay on the collagen in vitro
The effect of CBD-NT3 was tested by bioactivity assay on the collagen in vitro. The collagen was prepared from rat tail tendon as described above. Then 100 μL/well of serial dilutions of NT3 and CBD-NT3 (1, 10, and 100 μM) were added to the 48-well plates coated with collagen and incubated at 37°C for 2 h. Then the plates were washed extensively with PBS. DRG cells were seeded at a density of 105 cells/well under the same conditions as those described above, and 3 days later the cells were immunostained with β-tubulin type III and Hoechst 33342 as described above. Cells cultured under identical conditions but without growth factors served as a blank control.
Surgical procedures
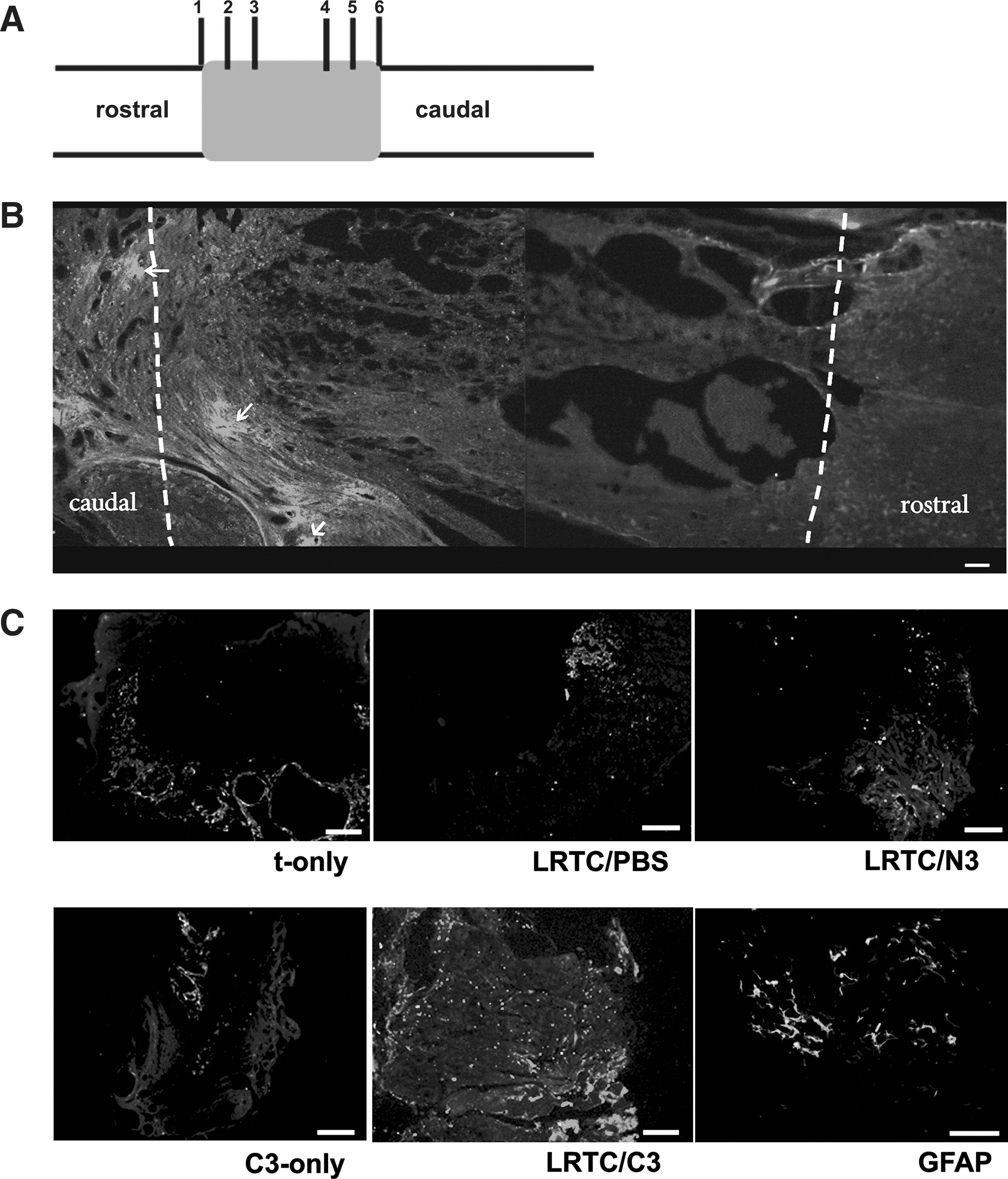
Seventy-five adult female Wistar rats (160–220 g) were used in this study. The procedures used were performed according to local guidelines on the ethical use of animals, and the National Institutes of Health “Guide for the Care and Use of Laboratory Animals” (NIH publication no. 80-23, revised 1996). After anesthesia by IP administration of 3.6% chloral hydrate (1 mL/100 g body weight), the dorsal aspect was shaved, and the rats received a dorsal laminectomy at T8–T10 to expose the spinal cord. The spinal cord was transected using microscissors, and the cord stumps were lifted to ensure complete transection. The lesion of the cord was dissected and formed a 2-mm-long cavity. Then LRTC bundles 2 mm long by 2 mm in diameter (50 bundles/rat) loaded with growth factor were implanted in the cavity. Because our model requires CBD-NT3 with LRTC (LRTC/C3 group; 15 rats), several control groups were also examined: LRTC loaded with PBS (LRTC/PBS group; 15 rats); LRTC loaded with NT3 (LRTC/N3 group; 15 rats); CBD-NT3 only (C3-only group; 10 rats); transected-only animals (t-only group; 10 rats); and sham-operated animals (sham group; 10 rats). During surgery the body temperature of each rat was maintained at 37°C. The surgical site was sutured in layers, and penicillin and sodium chloride were applied.
Extensive post-operative care was given after the rats were returned to their cages. Bladder expression was carried out twice a day as necessary until the bladder emptying reflex was re-established. Intramuscular injection of penicillin was continued after surgery twice a day for 3 days, and the cage floors were kept clean to reduce the incidence of infection. Food and water were available ad libitum.
Locomotion function
Functional recovery of the hindlimbs was assessed using the Basso-Beattie-Bresnahan (BBB) locomotor rating scale (Basso et al., 1995, 1996). The scale's scoring ranges from 0–21, with 0 representing complete paralysis, and 21 representing normal locomotion. This evaluation was performed for 4 min for each rat at 1, 2, 3, 4, 5, 6, 7, 8, 12, and 16 weeks post-injury by a researcher blinded to study group.
Grid walk testing was begun 1 month after surgery, when the injured rats began to show weight-bearing and plantigrade steps. The rats were allowed to climb across a 70-cm-high, 45°-sloped grid with holes 10 × 10 mm in size; Ramón-Cueto et al., 2000). Two side walls were used to keep the rats from falling off. Footfalls through the gird and a lack of plantigrade stepping were both considered errors. The total number of steps that a rat used to cross the grid and the number of errors were counted. All trials were videotaped and analyzed off-line frame by frame by a researcher blinded to study group. Any external sensory stimulation, such as via tail pinch, invalidated the test.
Anterograde tracing
Eighteen days before the rats were perfused, some animals were randomly chosen and the anterograde tracer biotinylated dextran amine (BDA, 10% wt/vol; Molecular Probes, Eugene, OR) was injected into eight sites(0.5 μL per site) with a glass micropipette-tipped Hamilton syringe. The sites were 1.0 mm deep in the dorsal cortical surface, 0.5 and 2 mm posterior to the bregma, and 1.0 and 2.0 mm from the midline (the sagittal suture). Sections of the lesioned spinal cord, as well as the rostral and caudal stumps, were treated with 0.3% Triton X-100 for 15 min, and then incubated in avidin-HRP (Molecular Probes), for detection of BDA-labeled axons.
Histological analysis
Four months after surgery, the rats were deeply anaesthetized and perfused with saline followed by 4% paraformaldehyde. The entire spinal cord was gently harvested and post-fixed in 4% paraformaldehyde at 4°C. A 1-cm section of spinal cord tissue, including the lesion site as well as the rostral and caudal stumps, was immersed in 20% sucrose buffer for 24–48 h at 4°C, then 30% sucrose buffer for 24 h, after which the cord was embedded in embedding compound. Serial longitudinal and transverse sections (20 μm thick) of the spinal cord were cut with a cryostat, mounted on gelatin-subbed slides, and kept at −20° C until further processing. The sections were blocked in 10% normal goat serum in PBS at room temperature for 2 h to prevent nonspecific staining. An anti-neurofilament IgG antibody (monoclonal, 1:200; Chemicon International, Temecula, CA) was used to show neuron-specific intermediate filaments, and a polyclonal anti-GFAP antibody (1:200; Abcam, Cambridge, MA) was used to localize astroglial cells and glial scarring. After incubation with primary antibody at 4°C for 24 h, the sections were incubated with the fluorescent secondary antibodies anti-mouse IgG (1:100; Invitrogen), and anti-rabbit IgG (1:200; Invitrogen), and examined with a fluorescence microscope (Leica DM IRE2; Leica Microsystems, Wetzlar, Germany). Motor neurons and fibers were stained with anti-serotonin primary antibody (5-HT, polyclonal, 1:200; Chemicon) at 4°C for 24 h, then the sections were incubated with anti-rabbit HRP-conjugated secondary antibody(1:100; Sigma-Aldrich) at 37°C for 2 h, and soaked in diaminobenzidine (DAB) solution until staining was evident. Negative controls using PBS instead of the primary antibody were performed to determine the specificity of the antibody. Ten or 20 random areas of each sample were counted to evaluate for positive nerve fibers, astroglial cells, or glial scars, and motor-related fibers. The percentage of positive areas/total tissue area were calculated using Image-Pro Plus software.
Statistical analysis
Data are presented as mean values ± standard deviation. The statistical significance of the differences in parameters was determined using two-tailed Student's t-test.
Results
Expression and purification of NT3 and CBD-NT3
The monomer molecular weights of NT3 and CBD-NT3 were 15.49 and 17.69 kD, as calculated according to the amino acid sequences. Most of the proteins were in inclusion bodies, and they needed to be refolded and then purified. Their expression and purification were determined using SDS-PAGE (Fig. 1B and C), and Western blot (Fig. 1D), under the same conditions.
The structure of linear rat-tail collagen (LRTC)
The prepared LRTC is shown in Figure 2A. It has a fibrous structure to support cell growth and extension along its surface. Bundles of LRTC were used as scaffolds to transplant into the cavity at the injury site. This SEM image shows the microstructure of LRTC (Fig. 2B).
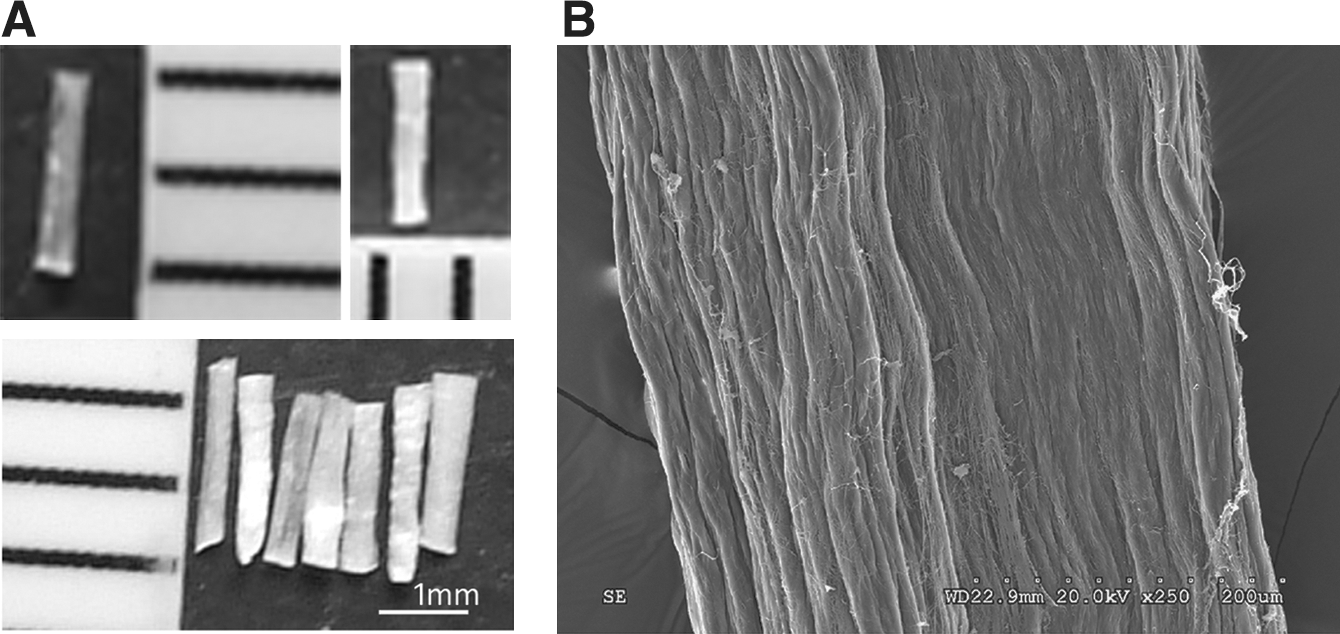
The structure of rat-tail collagen. (
CBD-NT3 binds specifically to type I collagen in vitro
The collagen-binding abilities of NT3 and CBD-NT3 were assessed by ELISA assays. According to the optical density at 405 nm, the amount of retained CBD-NT3 on the collagen was significantly higher than that of NT3, at solution concentrations ranging from 0.15–10 μM (Fig. 3A). The amounts of retained NT3 and CBD-NT3 were calculated by Scatchard analysis based on the standard curve. The curve shows the dissociation constant Kd values (Fig. 3B). The Kd values for the binding of NT3 and CBD-NT3 to collagen were 0.86 and 0.35 μM, respectively. The lower Kd value of CBD-NT3 confirmed that it could bind specifically to collagen.
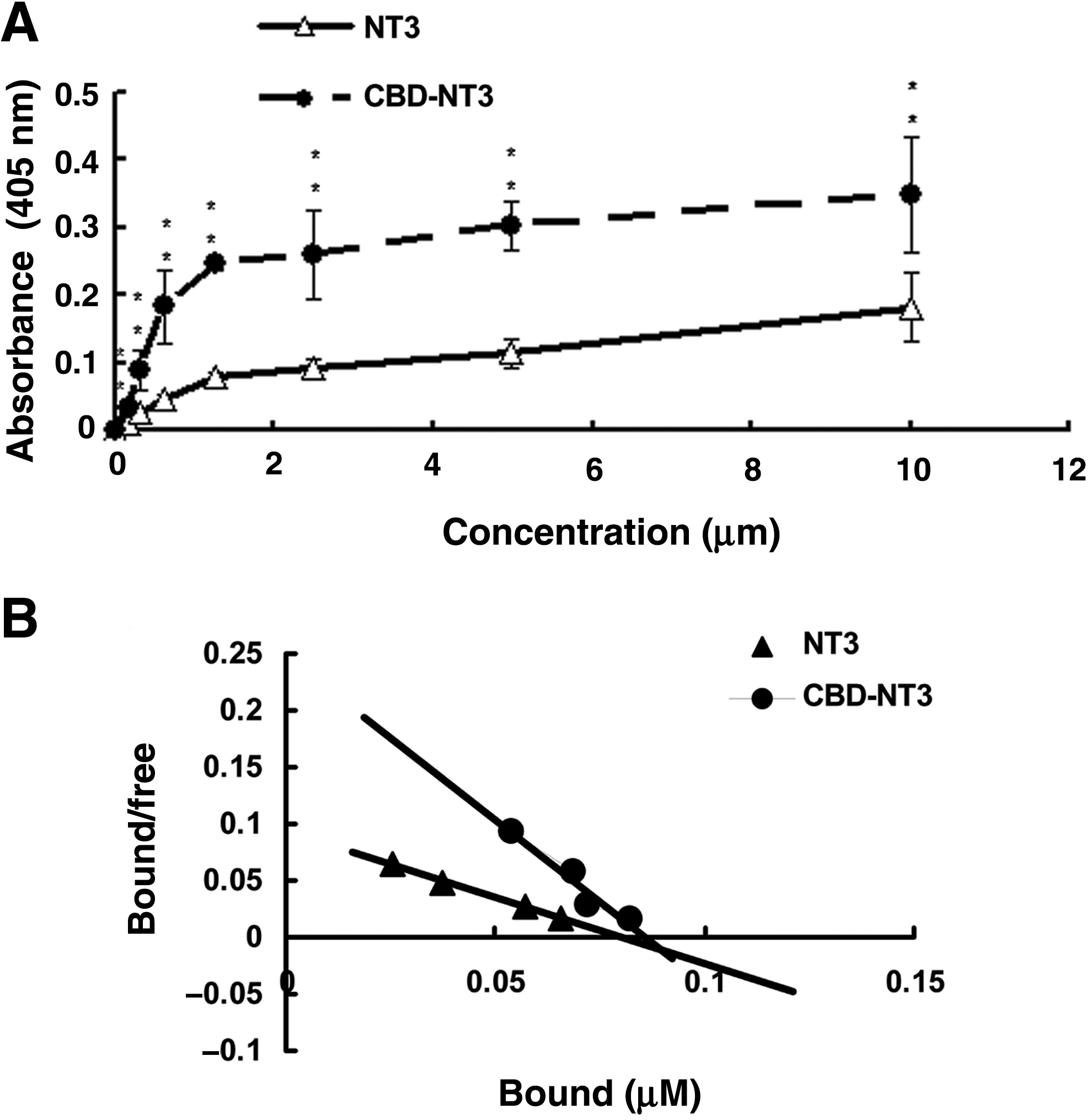
Collagen-binding ability and biological activity of recombinant NT3 and CBD-NT3 in vitro. (
NT3 and CBD-NT3 showed identical bioactivity on DRG
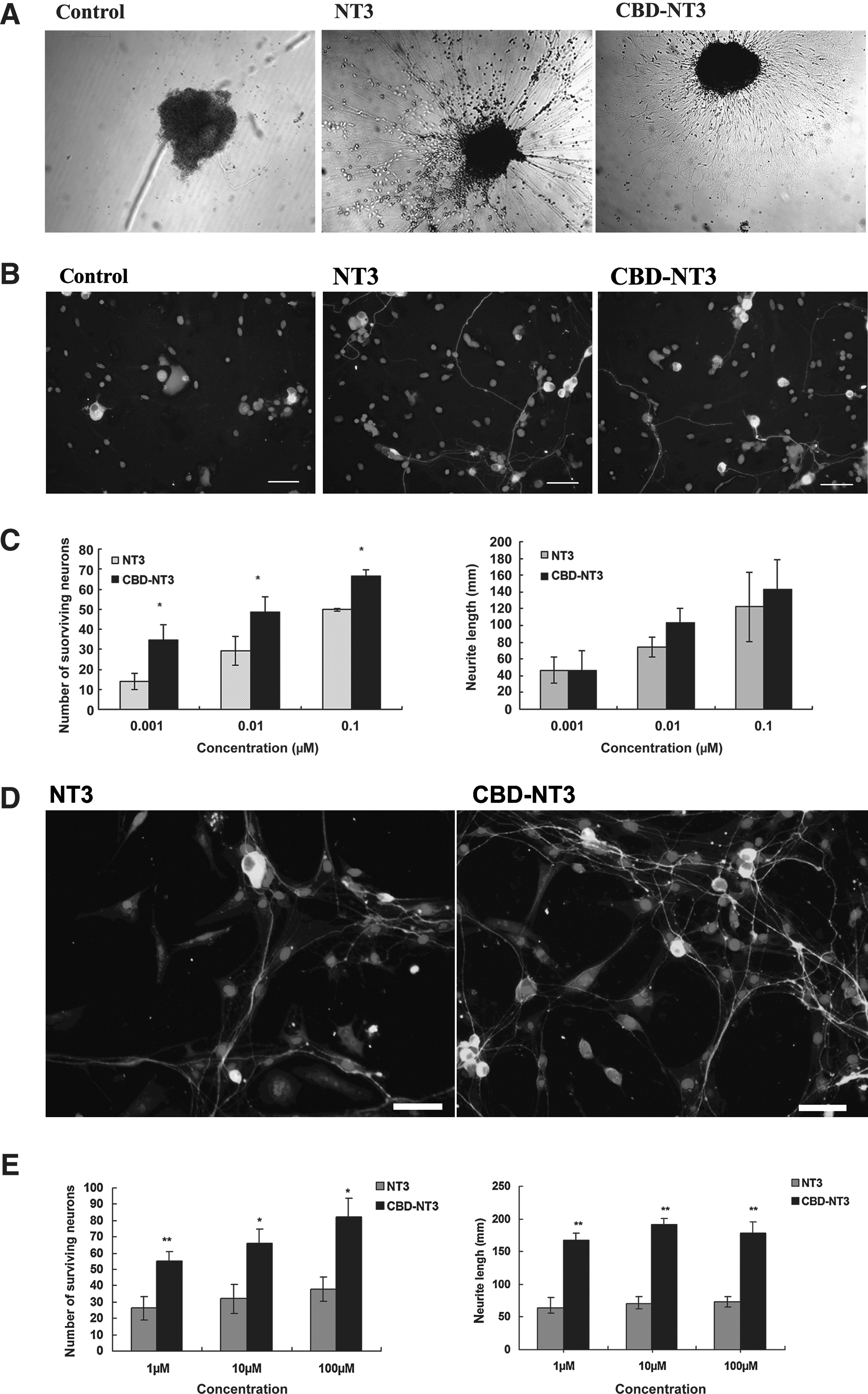
The biological activity of NT3 and CBD-NT3 were investigated with the lumbar DRG explants and DRG cells. Both NT3 and CBD-NT3 enhanced neurite outgrowth of the DRG explants (Fig. 4A). The DRG explants in the control animals displayed only minimal outgrowth (Fig. 4A). To allow quantitative comparison of the bioactivity of NT3 and CBD-NT3, DRG cells were used, and the cells were fixed and immunostained with β-tubulin type III to show neurons and neurites. Hoechst 33342 was used for detecting nuclei (Fig. 4B). NT3 or CBD-NT3 added to the DRG cell cultures increased the neurite outgrowth response with increasing doses. Both NT3 and CBD-NT3 were able to induce neurite growing at concentrations of 0.01, 0.1, and 1 μM, and there was no significant difference between the two groups (Fig. 4C). The surviving neurons in the CBD-NT3 group were significantly more numerous than those in the NT3 group (p < 0.05) (Fig. 4C).
(
CBD-NT3 promoted DRG outgrowth on collagen in vitro
We also examined the biological activity of NT3 and CBD-NT3 on collagen using DRG cells. At concentrations of 1, 10, and 100 μM, CBD-NT3 maintained more cells and promoted longer neurites than did NT3 (p < 0.05; Fig. 4D and E).
LRTC/C3 promoted better locomotor recovery
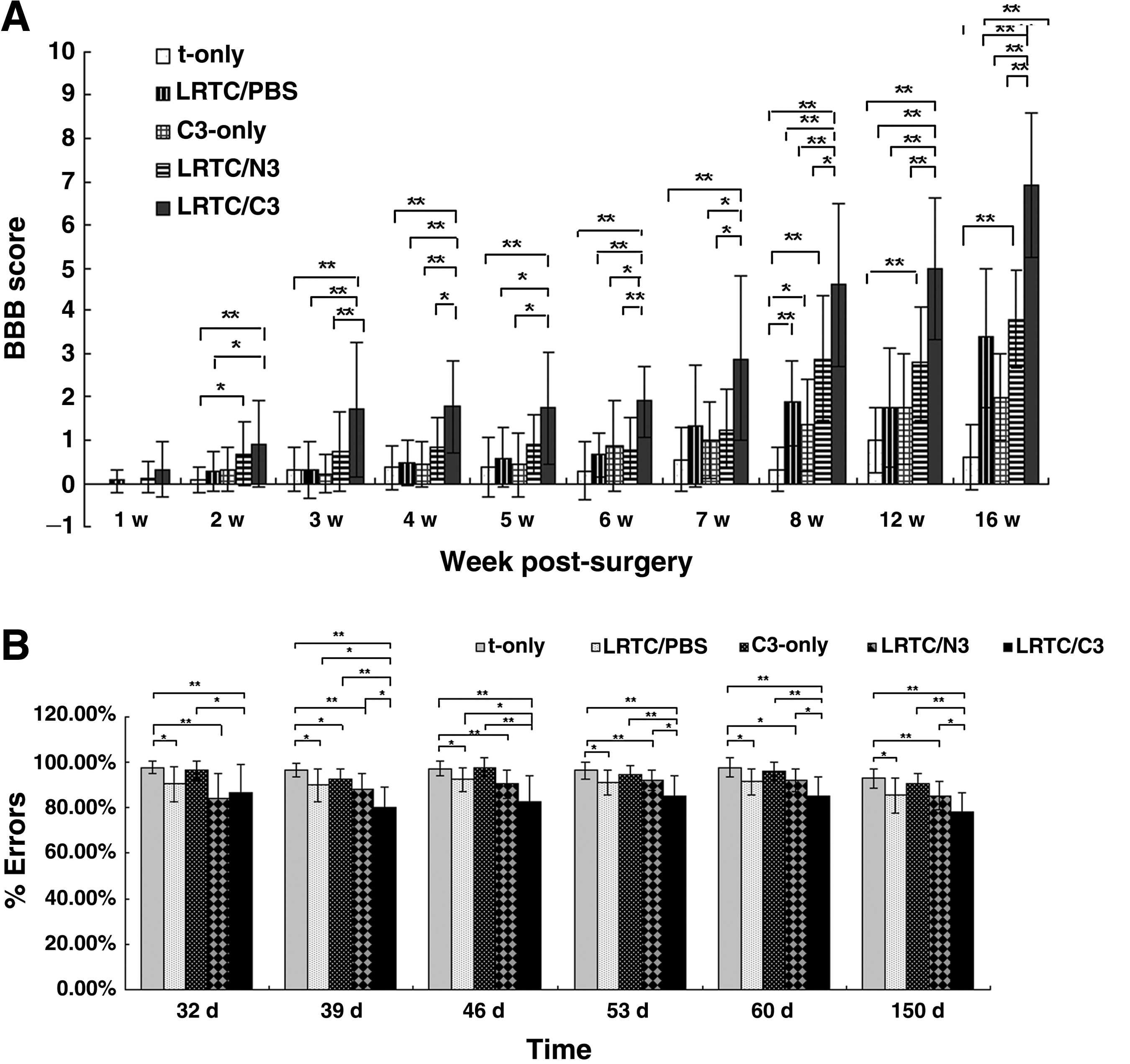
BBB scores were determined by comparative analysis of the scores over time (Fig. 5A). Some rats died due to urinary tract infections during the observation period. The sham-operated animals were nearly normal during the entire study period. At week 1, the BBB scores of the treatment groups were nearly 0, and there were no statistically significant differences among the groups. This demonstrated that the SCI model was successful. Significant differences were seen at week 2. The average scores of the LRTC/C3 group were higher than those in the t-only group (p < 0.01), and the LRTC/PBS group (p < 0.05). From week 3 onward, the animals in the LRTC/C3 group performed significantly better than those in the t-only, LRTC/PBS, and C3-only groups. From the week 4 post-surgery onward, the BBB scores of the LRTC/C3 animals were statistically significantly higher than those of LRTC/N3 animals.
(
The grid-walk test was used as an over-ground measure of motor function. Sham-operated animals showed normal performance during the entire study period. For the first 3 weeks, performance on weight-bearing stepping were nearly identical for the t-only, LRTC/PBS, LRTC/N3, C3-only, and LRTC/C3 groups. Some differences began to emerge starting about 30 days post-surgery (Fig. 5B). The t-only animals showed only a few weight-bearing steps during climbing, and their performance was significantly worse than that of the LRTC/PBS, LRTC/N3, and LRTC/C3 animals. The percentage of errors for the LRTC/C3 rats decreased dramatically from 32 days post-surgery until the rats were killed. Their error rates were statistically significantly lower than those of the t-only, C3-only, LRTC/PBS, and LRTC/N3 groups (Fig. 5B).
LRTC/C3 promoted the regeneration of nerve fibers
Regeneration of nerve fibers was estimated using NF200 immunostaining. The NF200-stained sections were 200 μm away from the border of the lesion and the rostral stump (Fig. 6A). There were only a few irregular regenerating nerve filaments at the lesion site in t-only, LRTC/PBS, and C3-only rats (Fig. 7A). In LRTC/N3-treated rats, stained bundles in isolated areas appeared in the lesioned spinal cord (Fig. 7A). However, there were significantly more dispersed filaments in LRTC/C3-treated rats, and the filaments were much less irregular (Fig. 7A). LRTC/C3 animals had statistically significantly more regenerating nerve fibers than t-only (p < 0.01), LRTC/PBS (p < 0.01), LRTC/N3 (p < 0.01), and C3-only animals (p < 0.05; Fig. 7B).
(
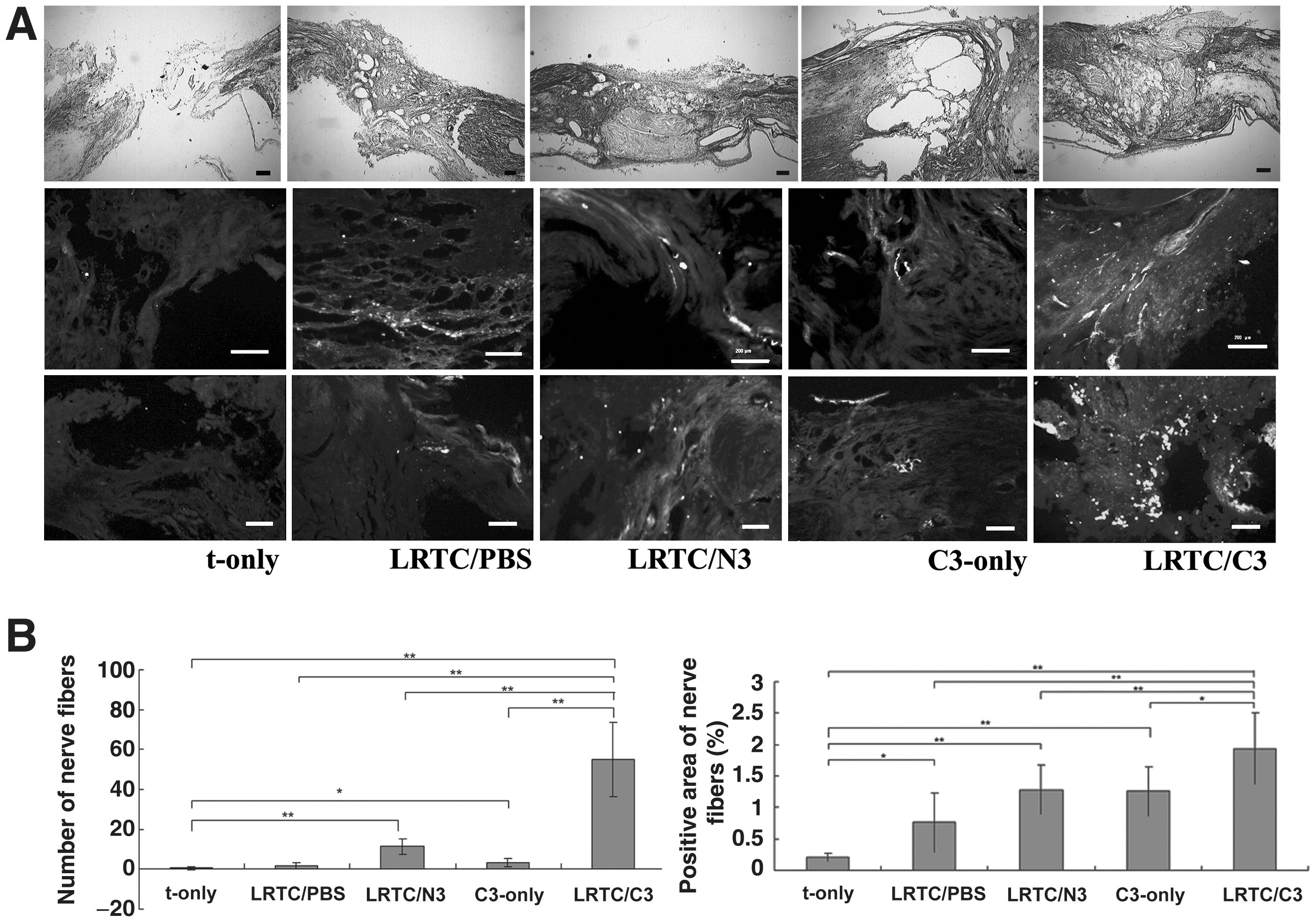
(
LRTC/C3 promoted regenerating nerve fibers that grew beyond the GFAP-positive glial scar
Generally, GFAP immunostaining revealed that the lesion cavity was surrounded by glial scars with few in-growing neuronal fibers after SCI. Thus it showed that the neuronal fibers stopped growing at the lesion's edge. We could see a dense GFAP-positive glial scar at the interface of the lesion and the spared tissue (Fig. 6B). A number of regenerated fibers were seen growing beyond the GFAP-stained lesion area in LRTC/C3 animals (Fig. 6C), but few were visible in the LRTC/PBS and LRTC/N3 animals (Fig. 6C). There were very few nerve fibers seen growing in the t-only and C3-only groups (Fig. 6C). These sections were taken 400 μm from the border of the lesion and the rostral stump (Fig. 6A).
LRTC/C3 enhanced corticospinal tract axon growth across the transplant
To investigate whether axons could traverse the transplant, BDA-labeled corticospinal tract (CST) axons were investigated. The sections were taken 400 μm from the caudal stump and the lesion interface (Fig. 6A). The fibers appeared wider and some had a beaded appearance, which may be characteristic of regenerating fibers (Fig. 8C). There were obvious differences among the groups in the lesioned and caudal portions (Fig. 8A), yet the differences were indistinguishable in rostral stump sections (data not shown). The numbers of BDA-positive fibers in the lesions and in the caudal stumps, as well as the percentages of BDA-positive areas were quantified (Fig. 8B). In the sections from LRTC/C3-treated rats, the percentages were significantly higher than those in the t-only (p < 0.01), LRTC/PBS (p < 0.01), LRTC/N3 (p < 0.05), and C3-only groups (p < 0.05).
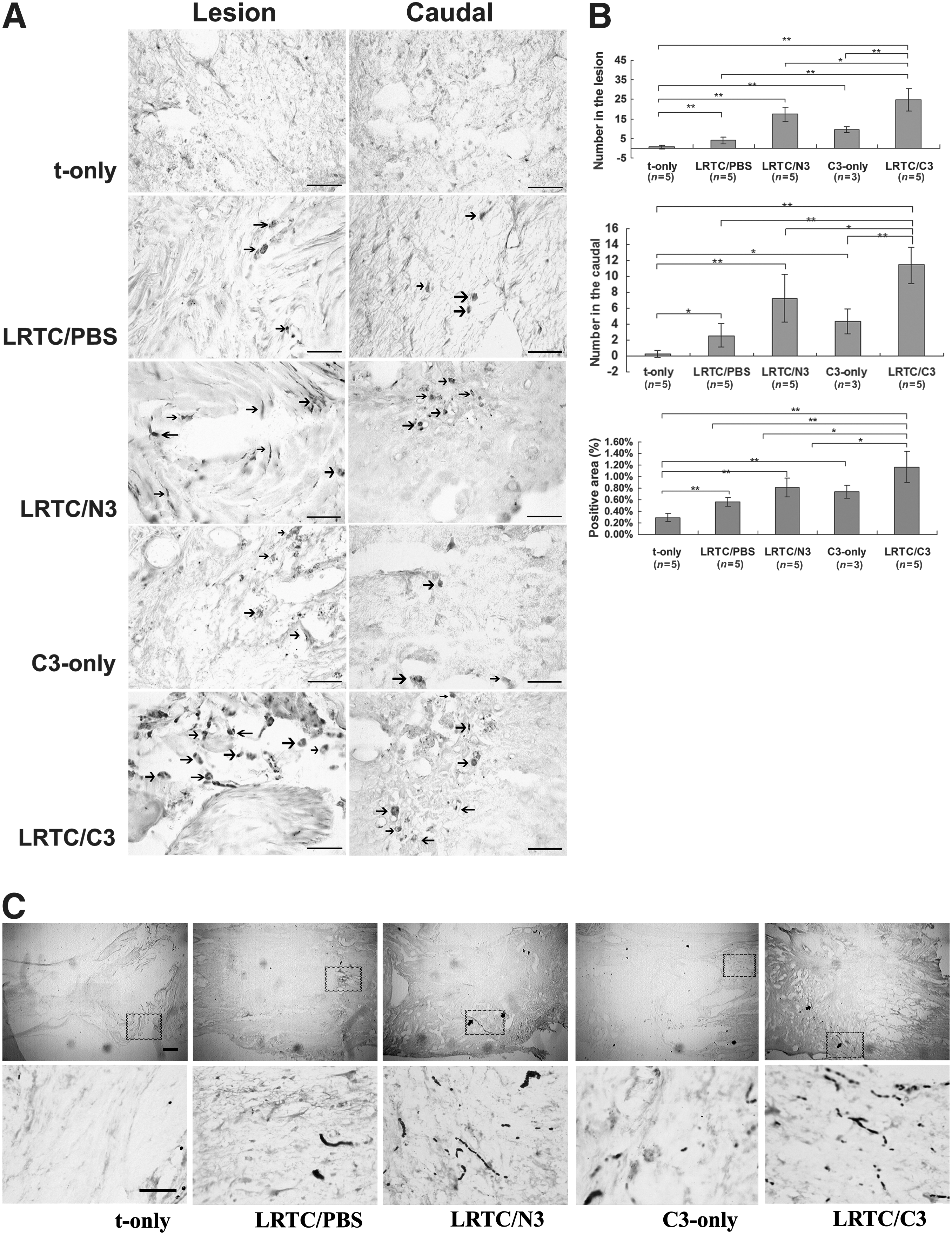
Detection and quantification of biotinylated dextran amine (BDA)-labeled corticospinal tract (CST) fibers. (
LRTC/C3 enhanced more 5-HT fiber regeneration
5-HT fibers, which are related to motor recovery, were examined and quantified with serotonin immunostaining. The sections were taken 200 μm from the interface of the lesion and the caudal stump (Fig. 6A). The 5-HT fibers seen in the LRTC/C3 group were much more distinct than in the other groups (Fig. 9A). Thus, there was significantly more regrowth of 5-HT fibers in the LRTC/C3 group than in the t-only (p < 0.01), LRTC/PBS (p < 0.01), LRTC/N3 (p < 0.05), and C3-only groups (p < 0.01; Fig. 9B).
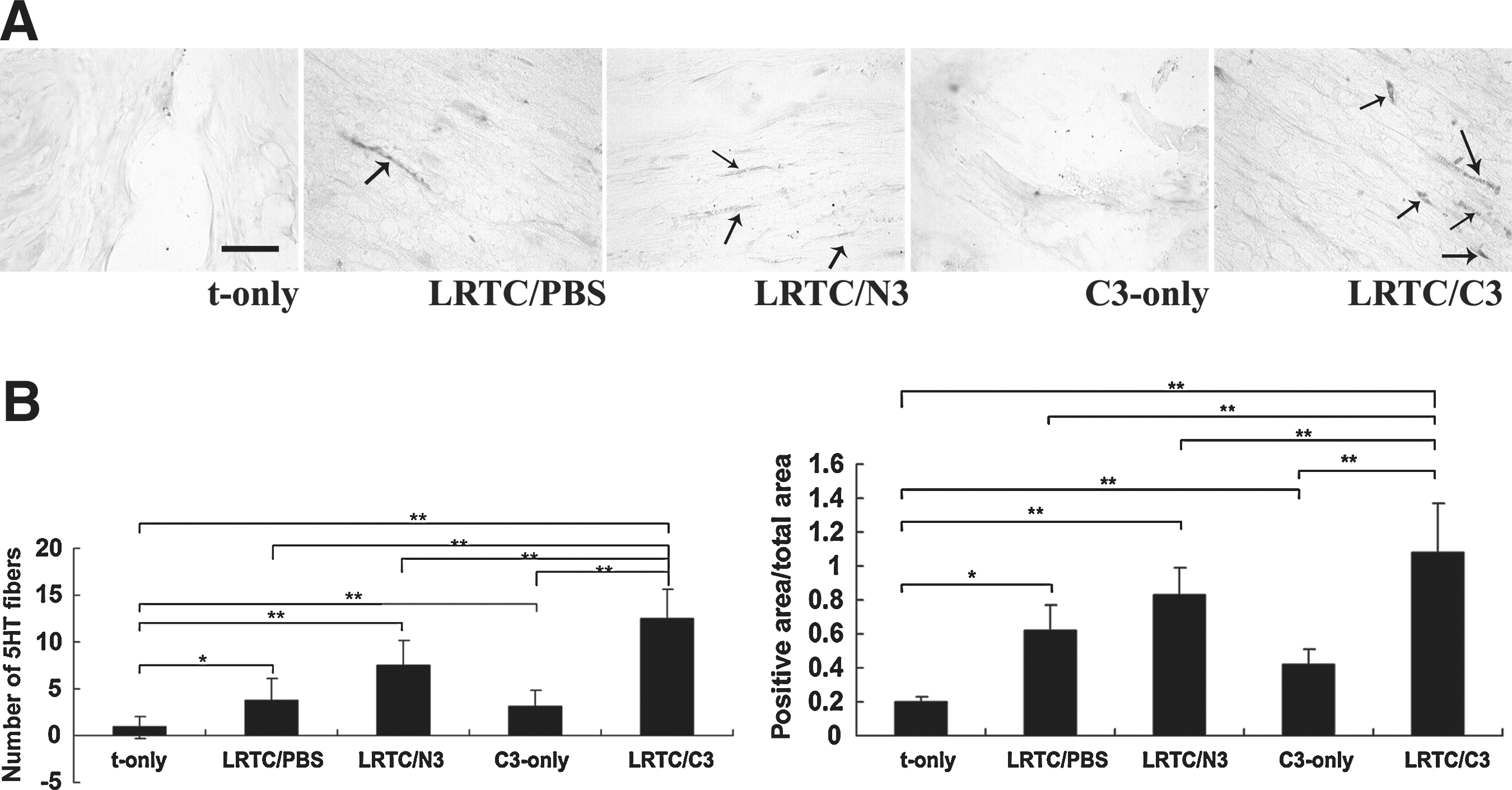
Raphe spinal axons and the quantitative assessment of serotonin (5-HT) axon growth through the transplant and spinal cord. (
Discussion
In the present study, we demonstrated the efficacy of the collagen scaffold LRTC with collagen-binding NT3 for SCI repair. Collagen has been used before as a vehicle for local application of NT-3 (Houweling et al., 1998; Schnell et al., 1994). However, the extracted collagen has generally lost mechanical strength and fiber structure. Linear collagen has a strong structure, and thus is suitable for nerve guidance, and in the past has been made from bovine aponeurosis (Lin et al., 2006). In our experiment, linear collagen prepared from rat-tail was used as a physical bridge, and provided an effective biological delivery system for NT3.
NT3 is an important neurotrophic factor for the regeneration of neural tissue (Bregman et al., 1995; Grill et al., 1997; Kobayashi et al., 1997; Rabchevsky et al., 2000; Ramu et al., 2007). However, direct application of NT3 is difficult because the constant exchange of cerebrospinal fluid removes NT3 (Taylor et al., 2004). The diffusion of NT3 and the large amount of NT3 thus required may weaken its effect, and may adversely affect the body. Therefore a major challenge is to increase the affinity between collagen and NT3. We utilized the collagen-binding peptide TKKTLRT (de Souza et al., 1994) to reconstruct CBD-NT3. In this way, CBD-NT3 could specifically bind to LRTC.
We demonstrated the binding ability of CBD-NT3 to collagen. The improved binding and lower Kd of CBD-NT3 promoted strong binding, as we expected. We then investigated whether the fused CBD-NT3 retained the bioactivity of the NT3. The tests on DRG explants and DRG cells demonstrated that the bioactivity of CBD-NT3 was not adversely affected. Moreover, CBD-NT3 was shown to promote cell survival. One possible reason for this was that the glia in the DRG secreted collagen to help recruit CBD-NT3 locally.
We chose a completely transected spinal cord model to evaluate the effects on SCI repair, thus avoiding the possibility that repair resulted from any remaining compensatory fibers. It has been reported that either collagen or NT3 can promote regeneration in SCI rats (Bregman et al., 1995; Grill et al., 1997; Kobayashi et al., 1997; Lee et al., 2001; Liu et al., 1998, 2001; Rabchevsky et al., 2000; Ramu et al., 2007; Stang et al., 2005; Yoshii et al., 2004). Thus NT3 plus collagen may have a synergistic effect to stimulate fiber regeneration, and thus have better therapeutic efficacy.
In our experiments, we analyzed locomotor function using the BBB scale and the grid-walk test. The BBB scale has seen widespread use (Basso et al., 1995, 1996; Buddeberg et al., 2004; Loy et al., 2002; Muir and Webb, 2000; Schucht et al., 2002). The 21-point scale is used to classify rat locomotion, ranging from normal to complete paralysis. Most of the rats in the LRTC/C3 group scored from 5–9 on the BBB scale. The scores of nearly all of the control animals remained at 0–2 during the entire 16-week study period, but to our surprise, one or two control rats attained scores of 4 or 5. Given that no axons remained after transection, there is no explanation for this phenomenon, except for the great plasticity present in spinal networks (Edgerton et al., 2001; Fouad and Pearson, 2004; Pearson, 2000; Rossignol et al., 2002). In the experiments using the BBB scale and the grid-walk test, animals in the LRTC/PBS, LRTC/N3, and LRTC/C3 groups performed significantly better than those in the other groups. There was a statistically significant difference between the LRTC/N3 and LRTC/C3 groups, indicating the potential therapeutic efficacy of the combination of LRTC with collagen-binding NT3. The performance of C3-only animals was worse, reflecting the fact that the collagen scaffolds were required for the treatment to be effective.
Fiber regeneration was demonstrated by immunohistochemical staining of nerve fibers. The animals in the LRTC/PBS, LRTC/N3, and C3-only groups had significantly enhanced axonal growth compared to the t-only rats. LRTC/C3 was able to stimulate statistically significantly more fiber regrowth. This combination may thus provide a more favorable environment for nerve fiber regeneration.
This phenomenon was also demonstrated in the experiment in which we examined BDA-labeled CST axons. The results indicate that more fibers were induced to grow beyond the transplant in LRTC/C3 animals. The significant difference seen between t-only animals and the LRTC/PBS, LRTC/N3, and LRTC/C3 animals demonstrates the importance of LRTC, but the best results were seen in the LRTC/C3-treated animals. These results are consistent with the rats' locomotor performance, again emphasizing the potential of the collagen/CBD-NT3 system. As the regenerating fibers seen in the LRTC/C3 group were more dispersed and branched, we postulate that the complex regenerating axons were able reconnect with one another, and that this plays a significant role. Thus, we believe that collagen-binding with NT3 may have a better reparative effect than native NT3, as it is immobilized by the LRTC. NT3 has been reported to induce and guide the regrowth of CST axons in injured spinal cord (Grill et al., 1997; Houweling et al., 1998; Jeffery and Fitzgerald, 2001; Houweling et al., 1998; Ruitenberg et al., 2005; Schnell et al., 1994; von Meyenburg et al., 1998; Zhou et al., 2003). Moreover, given the agreement of our BDA-labeled CST results with those of locomotor function recovery, we postulate that locomotor function recovery may be associated with the regrowth of CST axons.
5-HT fibers originate in the brainstem nuclei and descend throughout the spinal cord, and they show strong growth after SCI. 5-HT is an important neurotransmitter that influences the rhythmic pattern of hindlimb locomotion (Barbeau and Rossignol, 1991; Chen et al., 2007), and increases the excitability of motor neurons (Lee et al., 2007). Some researchers have confirmed a positive correlation between 5-HT fiber growth and the degree of functional recovery (Chen et al., 2007; Coumans et al., 2001; Piantino et al., 2006; Tobias et al., 2003). Calcium channels in adult rat motor neurons are also facilitated by 5-HT, more evidence of a relationship between functional improvement and 5-HT fiber regeneration (Li et al., 2007). In our investigations, the animals in the LRTC/PBS, LRTC/N3, and LRTC/C3 groups all saw improvements, but the LRTC/C3 group had the largest degree of 5-HT fiber regeneration. We believe that LRTC/C3 provides the best microenvironment for 5-HT fiber regeneration, and thus induces better recovery of locomotion.
Conclusion
In this study, we demonstrated that CBD-NT3 binds specifically to collagen, and that NT3 is thus retained, with no loss of bioactivity. Using a complete-transection rat spinal cord model, we have demonstrated that collagen binding to NT3 promotes axonal regeneration and functional restoration of the injured spinal cord.
Footnotes
Acknowledgments
This research was supported by the Natural Science Fund of China (no. 30930032, U0632008, 30270491, 30772232, and 30500526), and by the Chinese Academy of Sciences (KSCX2-YW-R133).
Author Disclosure Statement
No competing financial interests exist.