
Abstract
Neurectomy and botulinum toxin A (BoNT-A) injection cause denervated muscle atrophy, but questions remain about their clinical utility. We investigated time-series alterations of rat muscle weight, functional deficits, signaling pathways, and microscopic structures, to gain an understanding of the clinical implications. Between 2008 and 2009, the maximal calf circumference of patients for calf reduction either by neurectomy or BoNT-A injections were recorded for study. A rat skeletal muscle model was established through repeated or dose-adjusted BoNT-A injections and neurectomy. The survival, apoptosis pathways, functional deficits, and microscopic structures were investigated using Western blot, sciatic functional index (SFI), and transmission electron microscopy (TEM), respectively. The rat muscle weight ratio of the BoNT-A group had recovered to 89.3 ± 3.8% by week 58, but it never recovered in the neurectomy group. Muscle weight reduction by BoNT-A not only depended on the dose, but additive effects were also obtained through repeated injections. Rat SFI demonstrated rapid recovery in both groups. Molecular expressions showed a coherent and biphasic pattern. p-Akt and apoptosis-inducing factor (AIF) were upregulated significantly, with a peak at 8 weeks in the neurectomy group (p < 0.01), but cleaved caspase-9 and caspase-3 showed no significant changes in either group. TEM findings showed irreversible and reversible inner-structure disruption and sarcomere discontinuity in the neurectomy and BoNT-A groups, respectively. We demonstrated that denervation induced lasting muscle weight and structural changes of different degrees. Muscle weight reduction by BoNT-A was related to frequency and dose. AIF-mediated caspase-independent apoptosis was significantly different for neurectomy and BoNT-A injection.
Introduction
T
If neurectomy and BoNT-A induce irreversible and reversible muscle mass changes, respectively, how can the differences between the two groups be explained from a molecular point of view? Past reports have analyzed the signaling pathways and morphology (Jeng et al., 2009; Shen et al., 2006; White et al., 2000). In neurectomy, the contractive structures (myosin and actin syntheses) are reduced, and this in turn results in decreased myofibril size (Davatz et al., 2007). Molecular expression changes may increase (such as HSP70) or decrease (such as cytochrome c) as the muscle weight declines monotonically. The expression of cell adhesion molecules (CAMs) is also upregulated after nerve transection, before downregulating again (Hatoko et al., 2004). In other words, three molecular expression patterns are present after denervation: increase, decrease, and biphasic. Denervation also triggers signaling pathways in neurons and muscles with two opposing consequences: cell apoptosis and survival. Comparison of both signaling pathways may clarify the differences between the two groups. Fifteen years ago the Tsichlis laboratory discovered that Akt/protein kinase B (PKB) serine/threonine kinase was a target of phosphoinositide 3-kinase (PI3-K) (Franke et al., 1995). The principal survival-signaling pathway was PI3-K/Akt (Namikawa et al., 2000; Schwaiger et al., 2000). Activation of Akt was sufficient to prevent muscle atrophy, while the kinase activity of Akt was essential for IGF-1-induced hypertrophy (Bodine et al., 2001; Song et al., 2005). Apoptosis, a highly regulated systematic set of events resulting in cellular self-destruction without damaging the surrounding tissue, then emerged as the extrinsic and intrinsic pathways (Tang et al., 2000). In previous studies researchers reported that the intrinsic mitochondrial pathway was responsible for myonuclear apoptosis involving both cysteine aspartate-specific protease (caspase)-dependent and -independent apoptosis, depending on age, the nature of the atrophying condition, and the muscle type (Marzetti et al., 2010). The caspase-dependent pathway was initiated by the release of cytochrome c, which first activates the initiator caspases (i.e., caspase-9), and then the final effector caspase-3. On the other hand, caspase-independent apoptosis plays a major role in the denervation mitochondrial pathway. It is carried out by apoptosis-inducing factor (AIF), a flavoprotein located within the mitochondrial intermembrane space that possesses NADH-oxidase activity (Ferreira et al., 2008; Vahsen et al., 2004). AIF can translocate directly into the nucleus and induce DNA fragmentation, bypassing the caspase-dependent pathway altogether. In addition, regulatory factors such as the anti-apoptotic (i.e., Bcl-2) or pro-apoptotic (i.e. Bax) proteins regulate the responses to denervation (Chiu et al., 2003). Siu and Always (2005) found that denervation produced an increase in the Bax-to-Bcl-2 ratio. Bax is inserted into the outer mitochondrial membrane and creates mitochondrial permeability transition pores (mPTP) through which apoptotic factors stored in the intermembrane compartment are released. We endeavored to investigate the differences between these two denervation procedures in terms of the survival, apoptosis, and regulatory molecules, to see if they revealed any specific patterns. This may offer us clues for developing effective methods for prolonging the effect of BoNT-A.
In this study, we recorded the patients' data after both types of denervation procedures, then introduced a rat model to investigate the dynamic alterations of muscle weight, functional deficits, signaling pathways, and microscopic structures, that emerged in denervated rat skeletal muscle, as well as their human implications.
Methods
BoNT-A injection and neurectomy procedures on humans
From March 2008 to January 2009, we examined patients that had calf reduction (for aesthetic purposes) either by BoNT-A injections (n = 60, 20 ng/each calf; Allergan, Inc., Irvine, CA), or by selective neurectomy of the medial gastrocnemius and total soleus muscle (n = 70). Maximal calf circumference (cm) was recorded at 0 (pre-operative), 4, 8, and 26 weeks of denervation. The inclusion criteria in the study were as follows: (1) patients exhibited subjective calf enlargement or had a calf circumference greater than 34 cm; (2) no adjunctive procedures (e.g., liposuction) had been performed; and (3) patients agreed to a minimum follow-up period of 6 months.
Rat experimental designs for BoNT-A injection and neurectomy
Adult female CD® (SD) IGS rats of different ages weighing 186–220 g were purchased from BioLASCO Taiwan Co., Ltd. (Taipei, Taiwan). The animals were maintained in a 12-h/12-h light/dark cycle at a room temperature of 23° with standard laboratory rat chow and water provided ad libitum. All aspects of the animals' care followed the guidelines established by the Taipei Medical University Ethical Committee for Laboratory Animals. Both denervation procedures were performed on the rats. In the neurectomy group (n = 30), the left hindlimb was denervated by selectively excising 2 mm of the nerve to the gastrocnemius muscle. The other hindlimb (n = 5) served as a control without nerve resection in some rats to prevent muscle compensation bias. The gastrocnemius muscles from the neurectomized leg and the control leg were dissected and weighed over at six time points (0, 1, 4, 8, 12, and 26 weeks of denervation). In the BoNT-A group (n = 5/each time point), two doses of BoNT-A (1.5 and 1 ng/kg body weight, n = 5/each time point) were injected directly into the gastrocnemius muscle. An equivalent volume of saline was injected into the contralateral gastrocnemius muscle as control. The animals were euthanized at the stated post-denervation time points. Repeated BoNT-A injections were performed to study the additive effects on muscle weight reduction, with a time interval of 4 weeks between the first and second injections. The muscle weight was recorded at 0, 1, 4, 8, 12, 26, 30, 34, 42, and 58 weeks after the BoNT-A injections. The muscle was also used for Western blot analyses.
Functional assessment by the sciatic functional index (SFI)
Ten rats in both the full-dose BoNT-A and neurectomy groups were serially recorded for SFI evaluation. The walking-track analysis was based on observation of abnormal gait produced by both denervation procedures. The walking track featured a 10 × 60-cm track with 10-cm-high walls that led into a darkened box. Five walking-track trials were made for each rat at each time point, with the first three to familiarize the animal with the track. On the final two trials, the rats' hind feet were dipped in photographic developer and partly exposed photographic paper was placed on the track floor. The paper was later fixed and analyzed. Walking-track analysis was performed for each animal at 0, 1, 4, 8, 12, and 26 weeks of denervation. Several measurements were taken from the footprints: (1) distance from the heel to the third toe (i.e., the print length [PL]); (2) distance from the first to the fifth toe (i.e., the toe spread [TS]); and (3) distance from the second to the fourth toe (i.e., the intermediary toe spread [ITS]). All three measurements were taken for the experimental (E) and normal (N) sides. The SFI was calculated as previously described by Bain and colleagues (1989), according to the following equation:
The SFI ranges from near 0 for normal nerve function to −100 for total dysfunction.
Western blot analyses
Rat gastrocnemius muscle receiving full-dose BoNT-A and neurectomy was homogenized in ice-cold buffer B (20 Mm Tris-HCl [pH 7.5], 0.25 M sucrose, 50 mM β-mercaptoethanol, 1 mM phenylmethylsulfonyl fluoride, 200 μg/mL of leupeptin, 5 mM EDTA, 2 mM EGTA, and 10 mM benzamidine) using a homogenizer. Tissues were centrifuged at 100,000g for 30 min at 4°C, and supernatants were used as the cytosolic fraction. To obtain membrane fractions, the pellets were washed and extracted with buffer B containing 1% Triton X-100. Protein samples were subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The separated proteins were electrophoretically transferred to a 0.22-μm nitrocellulose membrane (Hybond-C Super or Hybond-ECL; Amersham Life Science, Buckinghamshire, U.K.) at 4°C overnight using a transfer blot apparatus (Bio-Rad Laboratories, Inc., Hercules, CA). Nonspecific sites were blocked with 5% skim milk in TTBS for 1 h at room temperature. The membranes were incubated for 2 h at room temperature, and were sequentially diluted to 1:2500 with anti-rabbit Akt, Bax, Bcl-2, cleaved caspase-3, AIF (Cell Signaling Technology, Inc., Beverly, MA), PI3-K (Santa Cruz Biotechnology, Santa Cruz, CA), and GAPDH (Santa Cruz Biotechnology). The membranes were washed and incubated for 1 h with a 1:2000 dilution of secondary antibody (alkaline phosphatase-conjugated goat anti-rabbit IgG). After three washings, antibody-reactive bands were visualized by an enhanced chemiluminescence detection system (Amersham Life Science). The blots were scanned using a GeneGnome bio imaging system (Syngene, Frederick, MD).
Transmission electron microscopy (TEM)
The muscle samples were cut longitudinally with the knife edge parallel to the muscle fiber, and osmicated using 1% osmium tetroxide in 0.1 M cacodylate buffer (pH 7.2). The samples were dehydrated in an ethanol series and embedded in Epon 812 using standard procedures. Ultrathin sections were cut and double-stained with uranyl acetate and lead citrate, and then examined under a Hitachi H-600 electron microscope (Hitachi, Tokyo, Japan).
Statistical analysis
All statistical calculations were performed using SPSS 15.0 (SPSS Inc., Chicago, IL). The differences in molecular expressions, muscle size, and functional deficits between post-denervation and normal values were investigated using a non-parametric method (Kruskal-Wallis and Mann-Whitney U tests). The SFI differences between the full-dose BoNT-A and neurectomy groups were analyzed using one-way ANOVA. The statistical significance was set at p < 0.01.
Results
Time-series study of botulinum toxin and selective neurectomy on human calves and rat muscle weight
The difference between pre- and post-operative maximal calf circumference (Δ) in the neurectomy group gradually decreased over time in the form of exponential decay. Maximal reduction was reached at 8 weeks (p < 0.01) then approached an asymptotic horizontal line. In the BoNT-A group a smaller change reached a trough value at 4 weeks, before subsequently returning to near-normal values (Fig. 1).
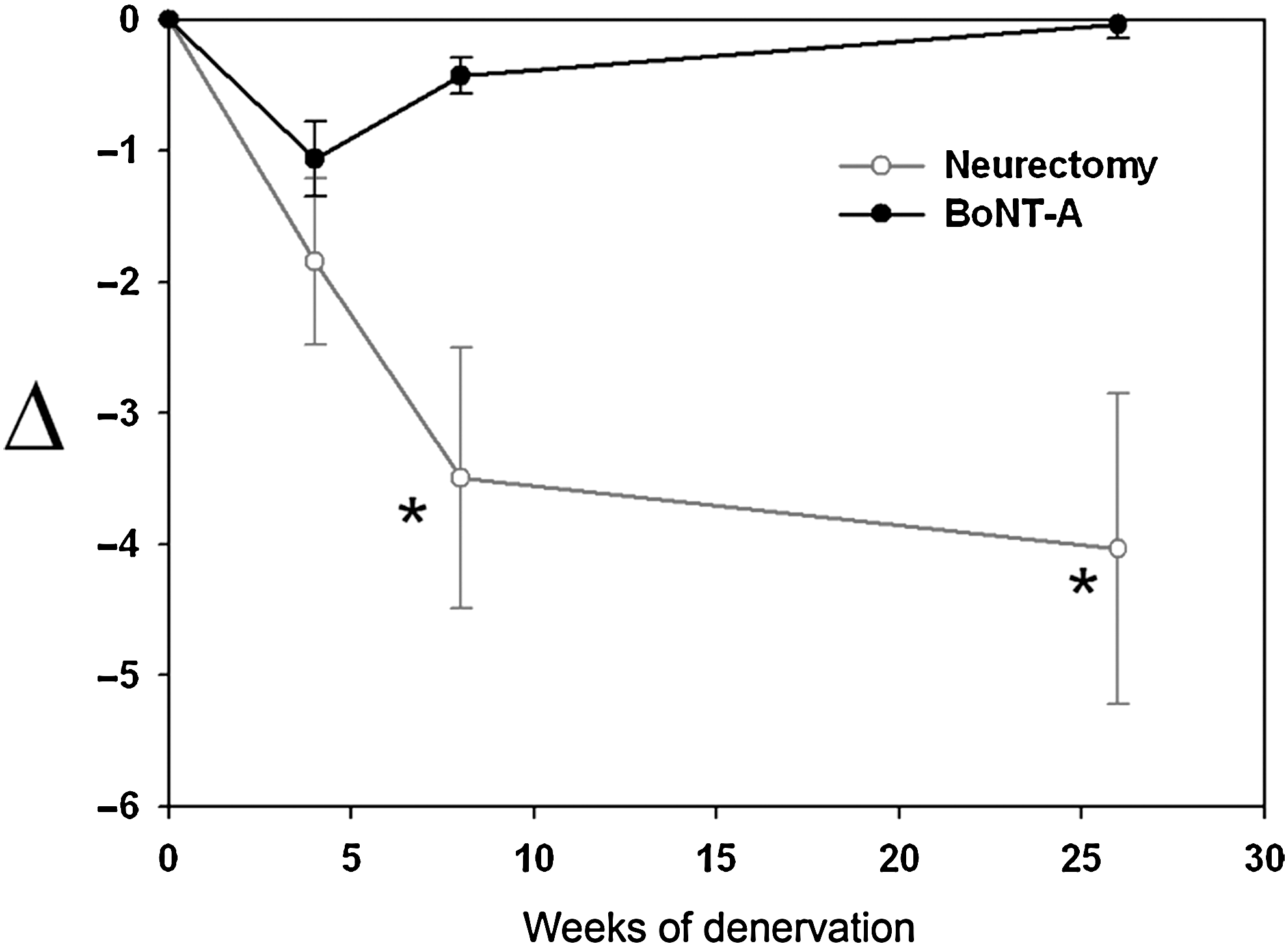
Human maximal calf circumference after botulinum toxin A (BoNT-A) injection and neurectomy. The pre- and post-treatment difference of human maximal calf circumference (Δ) after BoNT-A injection showed a maximal value at 4 weeks, then returned to normal levels at 26 weeks. Change after selective neurectomy revealed an gradual increase, with a statistically significant difference at 8 and 26 weeks (*p < 0.01).
Next, gastrocnemius muscle specimens from rats that received BoNT-A injection and neurectomy at different time points underwent weight evaluation (Fig. 2A). Neurectomy induced rapid loss of denervated rat skeletal muscle before 8 weeks of denervation, then approached 11.6 ± 2.1% of the skeletal muscle mass until 26 weeks (Fig. 2B). BoNT-A induced a V-shape recovery curve of muscle weight, with a trough seen at 4 weeks after each injection. The muscle weight ratio after a single full-dose BoNT-A injection recovered incompletely, to 89.3 ± 3.8% of original weight after 58 weeks. The muscle weight ratio with repeated full- and half-dose BoNT-A injections decreased to 72.7 ± 3.7% and 83.7 ± 4.5%, respectively, 26 weeks after the first injection, then after the second injection to 58.3 ± 6.1%, and to 70.3 ± 8.2% at 58 weeks. Dose-related results showed that lower-dose BoNT-A injection resulted in less muscle mass reduction.
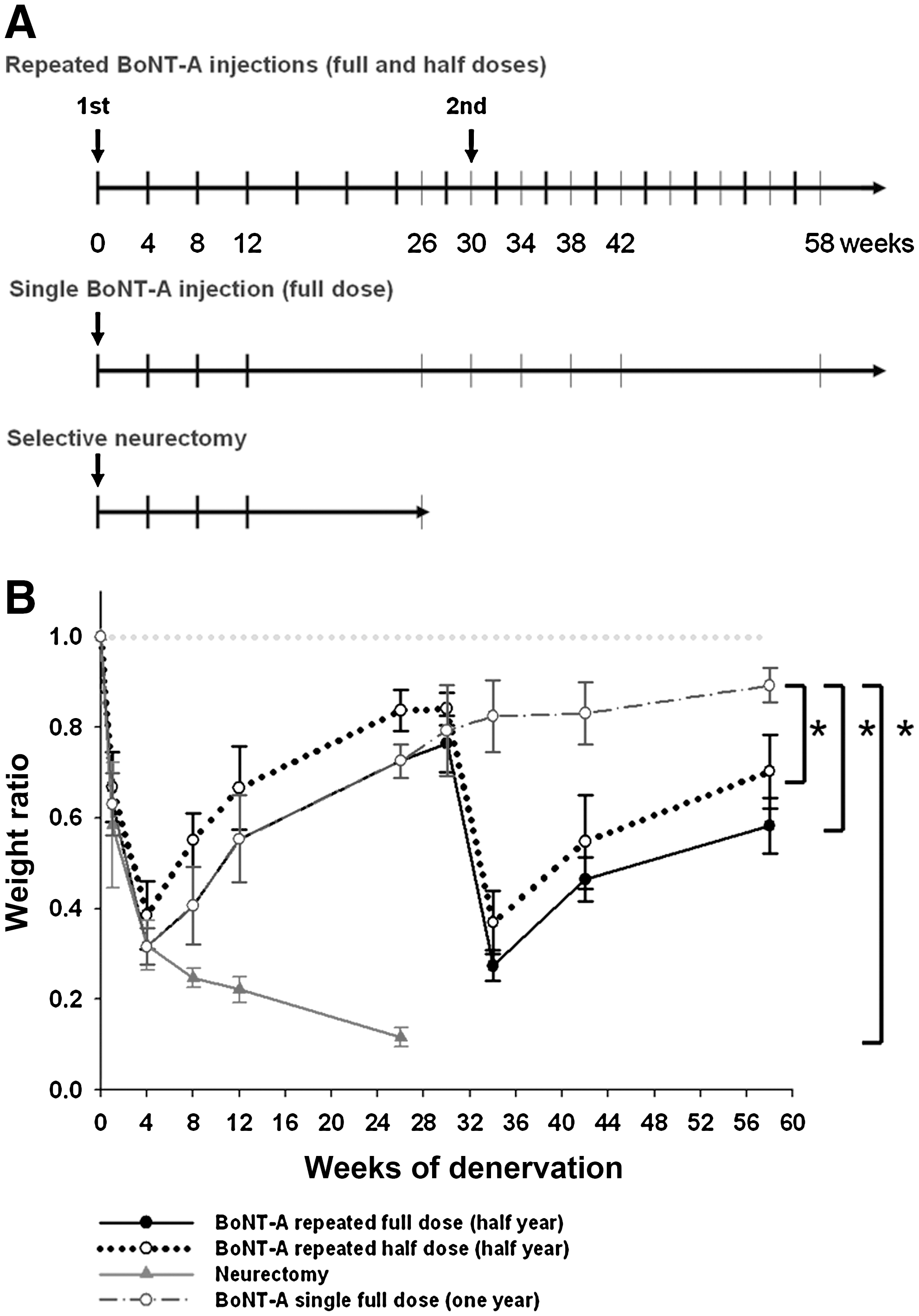
(
Functional assessment of BoNT-A and selective neurectomy in rats
Serial changes in SFI are depicted in Figure 3. SFIs of rats that received full-dose BoNT-A showed maximal functional deficit at around 1 week of denervation (p < 0.01), before soon returning to normal values. SFIs in the neurectomy group revealed immediate significant reductions in motor function that reached a plateau between 4 and 8 weeks, and then showed little improvement. There was a residual functional deficit (SFI = −19.6 ± 2.1%) at 26 weeks. Analysis by repeated one-way ANOVA showed that functional recovery of rats receiving BoNT-A appeared to be better than that of rats receiving neurectomy over time for SFI (group effects: p < 0.01 for interaction of groups over different time lines). The SFIs of rats that received BoNT-A and neurectomy showed no statistically significant difference after 8 and 26 weeks, respectively, compared to normal levels.
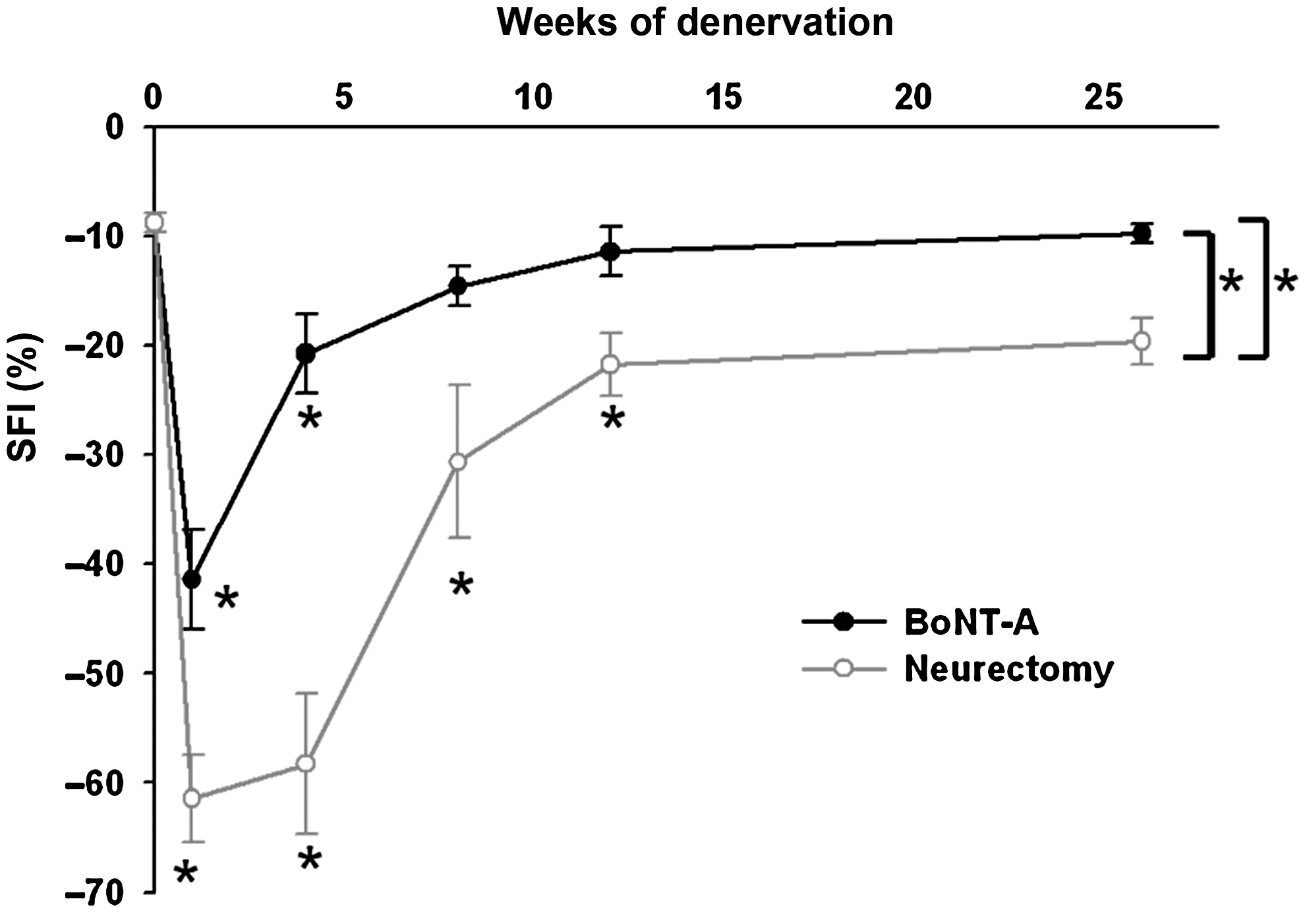
Sciatic functional index (SFI) after botulinum toxin A (BoNT-A) injection and neurectomy. Both groups showed immediate motor functional deficits. The SFI of rats that received full-dose BoNT-A soon returned to near-normal levels within 8 weeks, but SFI in the neurectomy group gradually recovered until 26 weeks of denervation. The SFIs of rats that received BoNT-A appeared to be better than those of rats that received neurectomy (*p < 0.01 compared to normal controls).
Dynamic expression profiles of survival- and apoptosis-signaling pathways
Quite surprisingly, long-term molecular expression of survival and apoptosis signaling showed clear time changes. The survival and apoptosis signaling in the BoNT-A group showed no significant changes compared to the neurectomy group. Survival signaling pathways (expression of PI3-K and p-Akt proteins) after denervation compared with normal controls revealed a compensatory increase, with a peak at 4 (BoNT-A group) and 8 (neurectomy group) weeks of denervation, before decaying to the trough level. p-Akt peak levels in the neurectomy group rose approximately 28.6-fold compared to normal innervated muscle at 8 weeks of denervation (p < 0.01), and persisted for a longer period of time compared than in the BoNT-A group until 26 weeks (Fig. 4A).
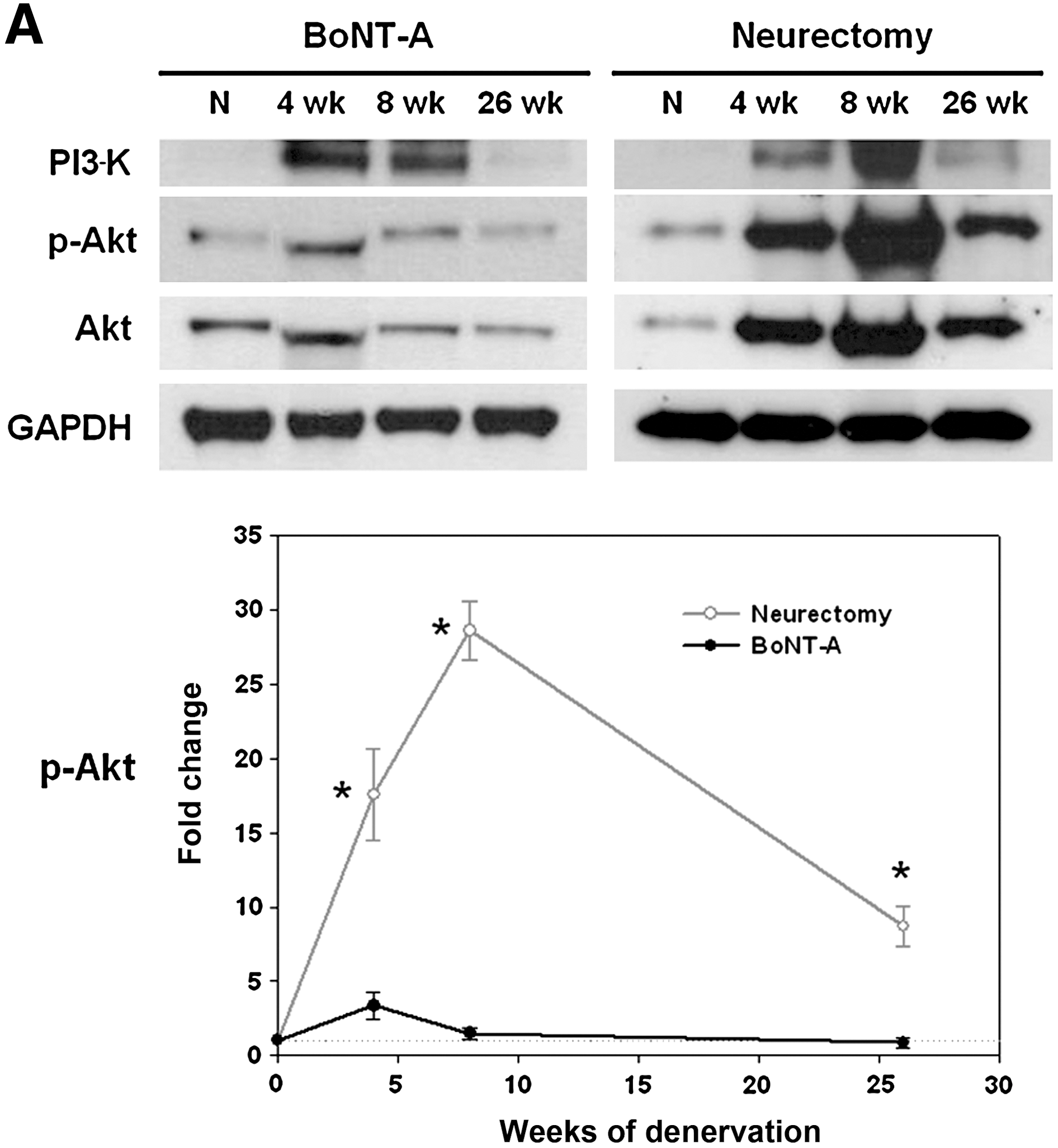
Survival and apoptosis signaling. (
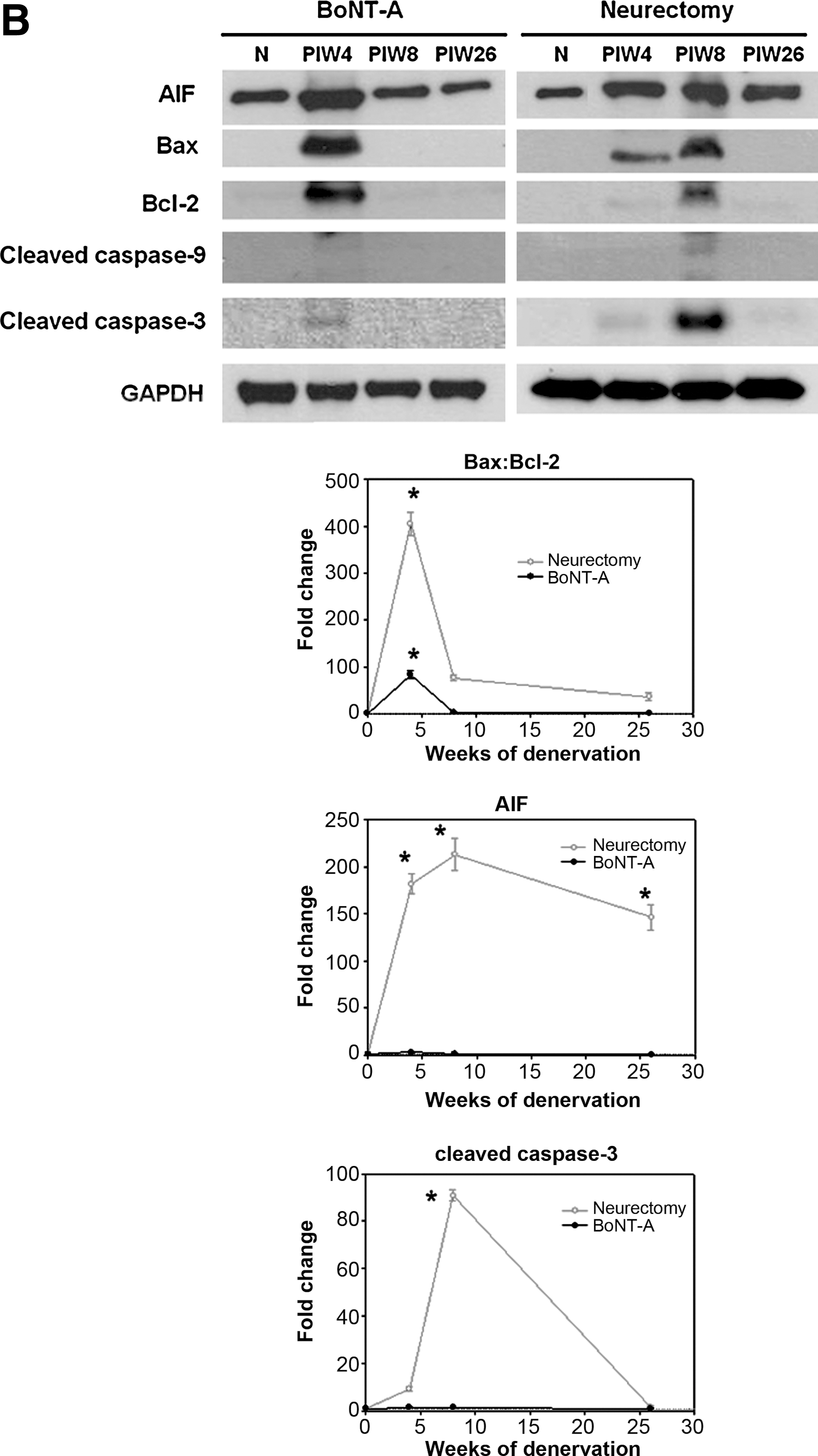
In addition, Western blots for apoptosis in the neurectomy group generally showed a stronger and more lasting increase than the survival-signaling pathway. For the mitochondrial caspase-dependent apoptosis pathway, the Bax:Bcl-2 (pro-apoptotic:anti-apoptotic protein) ratio increased, approaching a peak level, then quickly declined to near-normal levels (Fig. 4B). The maximal Bax:Bcl-2 ratio in the BoNT-A and neurectomy groups showed approximately 405.2- and 83.3-fold increases, respectively (p < 0.01). Active (cleaved) caspase-9 showed no significant elevation in either group. Cleaved caspase-3 levels showed no significant changes in the BoNT-A group. In the neurectomy group there was a ∼ 140.4-fold increase, but it then quickly declined to very low levels. Conversely, AIF for the mitochondrial caspase-independent pathway demonstrated persistently high levels over time, with a peak ∼ 213.1-fold increase seen at 8 weeks. A ∼ 146.5-fold increase remained at 26 weeks in the neurectomy group, compared to little change seen in the BoNT-A group (p < 0.01). It should be noted that the AIF:p-Akt ratio remained at around 16.9-fold, even at 26 weeks in the neurectomy group, but did not increase in the BoNT-A group.
Transmission electron microscopy
TEM showed that neurectomy induced irreversible progressive alterations of myofibrils involving the staggering and disorientation of z-discs with sarcomere disarray and inter-fiber widening (Fig. 5). BoNT-A by comparison caused mild destruction of sarcomeres at 8 weeks, but the normal structure was almost totally restored at 26 weeks.
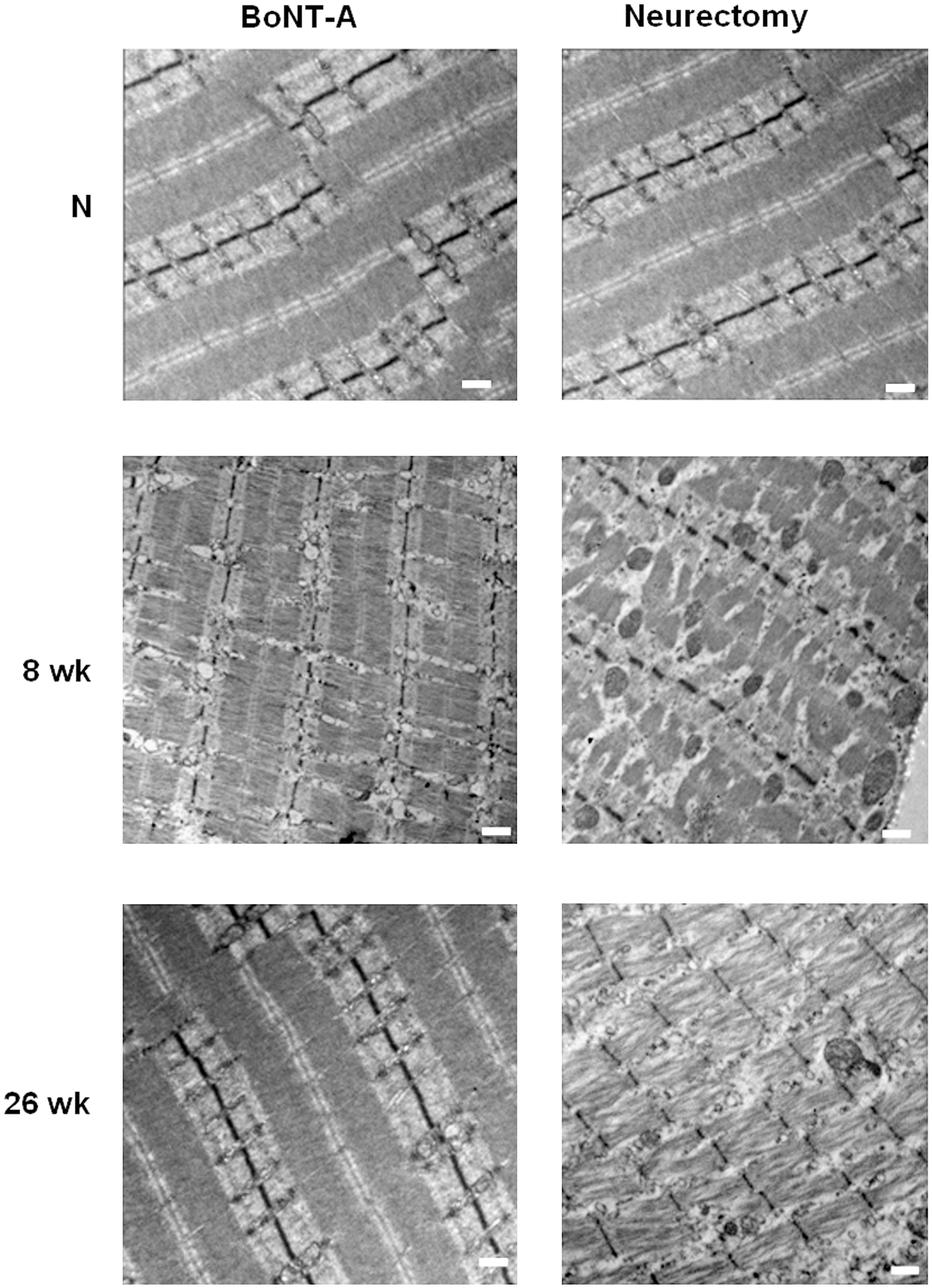
Transmission electron microscopy images. (Top) Normal sarcomere structures of rat muscle. (Middle) Irregular arrangement of z-discs was seen in the both groups at 8 weeks, but destruction of inner structures was less apparent in the botulinum toxin A (BoNT-A) group than in the neurectomy group. (Bottom) Neurectomy induced severe disruptions of the sarcomeres with widening of inter-fiber spaces seen at 26 weeks. The microscopic structures in the BoNT-A group ultimately returned to near-normal levels (scale bar = 500 nm).
Discussion
Denervation is a physiological or pathological response to nerve damage due to congenital anomalies, peripheral nerve injuries, disuse, or iatrogenic procedures. Previous research and our results from rat muscle showed that neurectomy decreased muscle weight irreversibly and monotonically in the form of exponential decay. There was still a small amount of muscle left after neurectomy, however, and this was verified by data from human patients receiving calf reduction by selective neurectomy (Tsai et al., 2008). BoNT-A injection has also become one of most frequently used aesthetic procedures in different medical subspecialties. The manufacturer's guidelines for BoNT-A injection indicate that when used for treating cervical dystonia, symptoms show improvement within the first 2 weeks, then they begin to re-emerge at approximately 8–10 weeks. Data in the literature show that the effects of BoNT-A last for 3–6 months (Ma et al., 2004). The consensus is that the effects of BoNT-A are short-term and reversible. Results of our study indicate that maximal effects of rat muscle weight reduction were reached 4 weeks after a single-dose BoNT-A injection, with incomplete recovery to 89.3 ± 3.8% of original muscle weight after 58 weeks, a result in agreement with the data of Lee and associates (2007). Additive effects were verified through repeated full- and half-dose BoNT-A injections, which produced muscle weight reduction and incomplete recovery to 58.3 ± 6.1% and 70.3 ± 8.2%, respectively, at 58 weeks. This observation surprisingly showed that there were some lasting changes in the BoNT-A group, despite the fact that muscle recovery is generally thought to occur with regard to NMJ stabilization, remodeling, and myogenesis after BoNT-A injection (Rogozhin et al., 2008; Shen et al., 2006). In short, higher frequency and doses of BoNT-A injections increased and prolonged the effect of muscle atrophy. BoNT-A injection for human calf reduction also showed a clear difference in human maximal calf circumference at 4 weeks, although calf circumference was regained later.
Patients are concerned with the length of time needed to recover after nerve injury or a denervation procedure. The extent of motor function deficits, which involve coordinated, complex, and multiple muscle-group interactions, is closely correlated with the number and the degree that denervated muscles are influenced, rather than being dependent on a single muscle. Analysis of motor function in both rat groups showed that immediate functional deficits of different degrees were present. SFIs in the BoNT-A group returned to near-normal levels with no statistically significant differences seen at 8 weeks, as shown in Figure 3. The timing of functional recovery was similar to that of bite force recovery after human masseter muscle hypertrophy post-BoNT-A injection (Ahn and Kim, 2007; Kim et al., 2009). However, the SFI of the neurectomy group at 26 weeks was −19.6 ± 2.1%, and thus there remained a residual functional deficit, even if selective neurectomy was only performed on one muscle. Neurectomy not only results in irreversible muscle mass reduction, but also induces a lasting functional deficit, as previously shown in the study by Tsai and associates (2008). Our study also revealed that denervation first results in loss of motor function, followed by muscle mass reduction. For clinical purposes, ideally only the motor function or the muscle volume should be targeted. To create iatrogenic motor function loss (i.e., for wrinkle relief), denervation should focus as completely as possible on the muscle that performs a specific function; on the other hand, denervation should target a specifically-innervated muscle as little as possible, in order to reduce muscle mass with minimal functional deficit. In short, the degree of the motor function deficit depends on the extent and number of denervated muscles.
To examine the reversible and irreversible effects on muscle weight in both denervation groups, the survival and apoptosis pathways were investigated. Molecular expression showed clear changes in both the BoNT-A and neurectomy groups, with peak elevations at 4 and 8 weeks, respectively, before declining again, a biphasic pattern. In the past, those performing short-term denervation studies incorrectly concluded that molecular expression was only up- or downregulated. Our novel results show that the main difference in myonuclear apoptosis between the two groups was for AIF-mediated caspase-independent apoptosis. The Bax:Bcl-2 ratio, which is related to the mitochondrial caspase-dependent apoptosis pathway, was elevated, with a peak at 4 weeks seen in both groups, after which this effect persisted only in the neurectomy group. Cleaved caspase-9 showed no changes in either group. Cleaved caspase-3 only increased at 8 weeks in the neurectomy group, but this effect quickly disappeared. This meant that caspase-dependent apoptosis did not significantly influence denervation changes over the long term. AIF, however, was strongly elevated, an effect that persisted even at 26 weeks in the neurectomy group. Survival signaling (PI3-K and p-Akt) showed no significant changes in the BoNT-A group, but was initially elevated before declining after 8 weeks in the neurectomy group. In a comparison of both the survival and apoptotic changes, the AIF:p-Akt ratio revealed that AIF apoptosis overrode survival signaling to induce irreversible neurectomy effects. Apoptosis resulted in the elimination of individual myonuclei (myonuclear apoptosis) and the related portion of sarcoplasm, without demise of the entire fiber. In other words, denervation may reduce the size of myofibers without completely destroying the multi-nucleated fibers, enabling muscle weight recovery similarly to a balloon being deflated or inflated. The mitochondria (subsarcolemmal and intermyofibrillar) are considered the primary centers for the integration of apoptotic signaling and the induction of apoptosis in denervation muscle atrophy. Adhihetty and colleagues (2007) reported that the mitochondrial content of rat skeletal muscle decreased after neurectomy. Our results verified that the Bax:Bcl-2 ratio is increased after neurectomy, resulting in the opening of the mPTP and release of mitochondrial AIF, causing direct DNA fragmentation and bypassing the caspase-dependent pathway. However, BoNT-A injection showed no significant survival or apoptotic changes. This might explain why BoNT-A induced reversible muscle mass changes, while neurectomy caused irreversible myofiber changes. The results of several other studies also confirmed that targeted deletion of AIF protects against mitochondrial oxidative phosphorylation, apoptosis, and may even prolong the life span (Pospisilik et al., 2007; Wissing, 2004). Thus AIF affects the reversibility of muscle weight reduction in response to denervation. Moreover, the TEM findings showed that there was progressive destruction of microstructures (such as z-discs and contractile elements) seen in the neurectomy group over time, but not in the BoNT-A group. Neurectomy induced the separation and widening of inter-fiber spaces, along with the destruction of myofiber elements (actins and myosins).
In summary, maximal muscle weight reductions after BoNT-A and neurectomy appeared at 4 and 8 weeks, respectively. The amount of muscle atrophy induced by BoNT-A increases with higher frequency and dose. Muscle atrophy of differing degrees and levels of persistence was seen after neurectomy and BoNT-A injection. Both groups also recovered complex motor function to a certain extent. In-depth investigation of molecular expression demonstrated a biphasic pattern (an initial elevation, then a decline). More importantly, AIF-mediated caspase-independent apoptosis appeared to result in lasting muscle atrophy after neurectomy, while there were no significant changes in apoptosis or survival signaling seen in the reversible muscle atrophy occurring after BoNT-A injection. These findings may be of help to those seeking ways of prolonging the effects of BoNT-A, or of minimizing the negative sequelae of nerve injuries.
Footnotes
Acknowledgment
This research project was supported by the Taipei Medical University Hospital (TMUH) under grant no. TMU98-AE1-B10.
Author Disclosure Statement
No competing financial interests exist.