
Abstract
Traumatic injury to the central nervous system (CNS) is accompanied by the spreading damage of secondary degeneration, resulting in further loss of neurons and function. Partial transection of the optic nerve (ON) has been used as a model of secondary degeneration, in which axons of retinal ganglion cells in the ventral ON are spared from initial dorsal injury, but are vulnerable to secondary degeneration. We have recently demonstrated that early after partial ON injury, oxidative stress spreads through the ventral ON vulnerable to secondary degeneration via astrocytes, and persists in the nerve in aggregates of cellular debris. In this study, we show that diffuse transcranial irradiation of the injury site with far red to near infrared (NIR) light (WARP 10 LED array, center wavelength 670 nm, irradiance 252 W/m−2, 30 min exposure), as opposed to perception of light at this wavelength, reduced oxidative stress in areas of the ON vulnerable to secondary degeneration following partial injury. The WARP 10 NIR light treatment also prevented increases in NG-2-immunopositive oligodendrocyte precursor cells (OPCs) that occurred in ventral ON as a result of partial ON transection. Importantly, normal visual function was restored by NIR light treatment with the WARP 10 LED array, as assessed using optokinetic nystagmus and the Y-maze pattern discrimination task. To our knowledge, this is the first demonstration that 670-nm NIR light can reduce oxidative stress and improve function in the CNS following traumatic injury in vivo.
Introduction
Oxidative stress is an important contributor to damage following injury to the CNS, and can be modulated by treatment with far red to near infrared (NIR) light. We have recently demonstrated that the oxidative stress marker manganese superoxide dismutase (MnSOD) spreads through the ventral ON vulnerable to secondary degeneration via astrocytes, as early as 5 min after partial ON injury (Fitzgerald et al., 2010), and remains in the nerve in aggregates of cellular debris (Fitzgerald et al., 2009a). MnSOD is a marker of increased production of the free radical superoxide, which it catalytically converts to H2O2. It serves as a general marker for oxidative stress (Aucoin et al., 2005), is associated with RGC death following injury due to excess H2O2 production (Nguyen et al., 2003; Levkovitch-Verbin et al., 2000), and is rapidly upregulated following spinal cord injury (Earnhardt et al., 2002). Oxidative stress can occur as a consequence of excessive accumulation of Ca2+ ions (Park et al., 2004; Tezel, 2006). Spreading of calcium signals through the astrocyte syncytium and to adjacent neurons and glia has also been demonstrated in a variety of in vitro and in vivo systems (Halassa and Haydon, 2010; Hamilton et al., 2008, 2010; Lobsiger and Cleveland, 2007; Scemes et al., 2000). Altered communication between astrocytes and oligodendrocytes via gap junctions may result in oligodendrocyte dysfunction and eventual myelination deficits with important functional consequences (Nave, 2010; Park et al., 2004; Williams et al., 2007). Furthermore, disruptions in the ionic balance may be transmitted from astrocytes directly to axons at their contact points at nodes of Ranvier (Butt et al., 1994), leading to degeneration of neurons despite the maintenance of myelin sheaths (Nave, 2010). We have recently shown that the density of myelinated axons falls in areas of ON vulnerable to secondary degeneration 28 days after partial ON injury, suggesting that myelin is indeed vulnerable; furthermore, myelin loss is accompanied by continued loss of visual function (Fitzgerald et al., 2009a; Selt et al., 2010). It is likely that containment of early oxidative stress will be required in order to control later disruption to myelin and loss of function.
Cytochrome c oxidase is a key component of the electron transport chain. It is required for ATP biosynthesis and is often disrupted in inherited and acquired impairments of mitochondrial function, resulting in oxidative stress (Sadun, 2002). Cytochrome c oxidase is a primary photoacceptor of light in the red to NIR region of the spectrum (Eells et al., 2004; Karu, 2003). In a number of in vitro models, it has recently been demonstrated that direct or indirect inhibition of cytochrome c oxidase by exposure of neurons to potassium cyanide, rotenone, or tetrodotoxin can be reversed by treatment with NIR light at a wavelength of 670 nm, indicating that increasing cytochrome c oxidase activity with 670-nm light reduces oxidative stress (Desmet et al., 2006; Liang et al., 2008; Wong-Riley et al., 2005; Ying et al., 2008). NIR light treatment reduced formation of reactive oxygen species in response to potassium cyanide, with resultant decreases in apoptosis (Liang et al., 2006). NIR light treatment also protected neurons and stimulated NGF-induced neurite outgrowth in conditions of oxidative stress induced by H2O2 exposure (Giuliani et al., 2009). Furthermore, cytochrome c oxidase activity was increased in untreated neurons exposed to NIR light (Desmet et al., 2006). In vivo, NIR light treatment, as opposed to perception of light at this wavelength, has been shown to prevent rotenone-induced decreases in visual function, with accompanying increases in cytochrome c oxidase and superoxide dismutase activity (Rojas et al., 2008). NIR light treatment also attenuates the toxic effects of methanol-derived formate on retinal function (Eells et al., 2003). Other beneficial effects of NIR light treatment include improvements in working memory (Michalikova et al., 2008), improvements in chicken hatchling survival rates (Yeager et al., 2005), accelerated wound healing, and the prevention of oral mucositis (Conlan et al., 1996; Desmet et al., 2006). In this study, we assess 670-nm light delivered by LED array, as a treatment for secondary degeneration following partial ON transection. It is important to note that 670-nm light is referred to as red, far red, and NIR light in the literature (Eells et al., 2004; Rojas et al., 2008; Yeager et al., 2005), and here we use the term NIR to describe our 670-nm treatments. We demonstrate a reduction in oxidative stress in the linked astrocytic network of ventral ON vulnerable to secondary degeneration, which is accompanied by improvements in visual function.
Methods
Animals
Female PVG hooded rats (160–190 g) were bred at the Animal Resources Centre (Murdoch, W.A.), housed under a standard 12-h light/dark cycle, and fed standard rat chow and water ad libitum. Procedures conformed to “Principles of Laboratory Animal Care” (National Institutes of Health publication no. 86-23, revised 1985), and were approved by The University of Western Australia's Animal Ethics Committee. The rats were anesthetized with xylazine (ilium xylazil, 10 mg/kg IP; Troy Laboratories, Glendenning, N.S.W., Australia) in combination with ketamine (Ketamil, 50 mg/kg; Troy Laboratories) and euthanized with Euthal (sodium pentobarbitone, 850 mg/kg, and sodium phenytoin, 125 mg/kg IP). Nine groups of animals were used (see Fig. 1 for experimental design).

Schematic representation of the experimental design for the three series of experiments conducted in the current study. Animals that had undergone partial optic nerve (ON) transection with or without near infrared (NIR) light treatment using the WARP 10 LED array are referred to as PT + NIRWARP10, and PT no NIRWARP10, respectively (or PT + NIRVET75 and PT no NIRVET75 for animals treated with the VET 75 LED array). Unoperated control animals are referred to as Unoperated no NIRWARP10 or Unoperated no NIRVET75 (PT, partial ON transection; ES, unoperated eyelid suture; ESR, unoperated eyelid suture removed; OKN, optokinetic nystagmus; Y, Y-maze pattern discrimination testing; S, sacrifice and perfusion; IHC, immunohistochemical analysis; EM, ultrastructural analysis by electron microscopy).
Partial optic nerve transection
The procedure has been described previously (Fitzgerald et al., 2009a; Levkovitch-Verbin et al., 2003). Briefly, the skin overlying the skull was incised along the midline, and the parenchyma of the right ON was exposed approximately 1 mm behind the eye by making a slit in the dura mater. A controlled 200-μm cut (approximately one-quarter of the ON width) was made in the dorsum of the ON using a diamond radial keratotomy knife, with the depth determined by the protrusion of the blade beyond a surrounding guard. Care was taken not to stretch the ON or damage major ophthalmic blood vessels. The sheath was closed, the skin was sutured (4-0 silk), and the animals recovered on a warming blanket. The rats were treated with an antibiotic and an analgesic subcutaneously (neomycin, 10 mg/kg in sterile PBS; carprofen, 2.8 mg/kg), and a topical antibiotic was applied to the skin wound (Tricin; Jurox Pty. Ltd., Rutherford, N.S.W., Australia). No postoperative infections were observed. We have previously shown that animals undergoing sham partial ON transection do not differ significantly from unoperated animals (Fitzgerald et al., 2009a). Unoperated rats were therefore used for anesthesia and handling controls.
Near infrared light treatment
Following partial ON transection, the animals were treated with 670-nm NIR light 3 cm above the surface of the entire head, using one of two LED arrays (WARP 10 or VET 75; Quantum Devices Inc., Barneveld, WI). Both treatment regimens commenced immediately following the partial ON transection (day 1; Fig. 1). Both devices delivered light with a center wavelength of 670 nm, and a full-width at half-maximum bandwidth of approximately 22 nm. Each array was positioned parallel to, and at a distance of 3 cm from, the dorsal surface of the rat's head. The WARP 10 LED array delivered a surface irradiance of 252 W/m−2 for 30 min per treatment. The VET 75 LED array delivered a surface irradiance of 1859 W/m−2 for 3 min per treatment, providing a more intense exposure for a shorter time. The spectral irradiance (300–900 nm) of both arrays was measured using a calibrated USB4000 CCD spectroradiometer (Ocean Optics, Dunedin, FL) and a 3-m-long, 200-μm-diameter quartz optical fiber fitted with a CC-3-UV cosine corrector irradiance probe.
We also quantified the amount of light reaching the injury site by recording in vivo the spectral irradiance at different intracranial locations: under the ON, behind the eye, and under the center of the brain. Access to the ventral surface of the ON was obtained by removing the lower jaw, palate, and overlying musculature of a freshly euthanized rat. An opaque black plastic shield positioned beneath the upper jaw prevented light other than that passing through the eye and/or cranium from reaching the irradiance probe. A single animal was used for the irradiance measurements. Three spectra were measured at each probe location and averaged; the standard deviations in total (550–750 nm) irradiance at the different measurement locations ranged from 10–28%, and primarily reflect subtle differences in probe position between subsequent scans within a set.
The WARP 10 light-emitting diode (LED) array was used to deliver NIR light for a total of 30 min per treatment. Choice of treatment time with the WARP 10 array was based on previous in vivo studies demonstrating beneficial effects (Rojas et al., 2008). In the first experiment, treatments were administered immediately following partial ON transection, and again 24 h later, immediately before euthanasia (PT + NIRWARP10 group, n = 3). The length of the treatment required daily anesthesia with xylazine and ketamine (see above). The non-treatment group therefore comprised partial ON-transected rats that were anesthetized twice over the 24-h period, but received no subsequent NIR light treatment (PT no NIRWARP10 group, n = 3). Unoperated control animals were similarly anesthetized (Unoperated no NIRWARP10 group, n = 3).
An additional experiment on a separate group of partial ON-transected rats was performed to assess the effects of NIR light treatment delivered each day for 6 consecutive days (PT + NIRWARP10 group, n = 8). The non-treatment group comprised animals receiving partial ON transection, but no subsequent NIR light treatment (PT no NIRWARP10 group, n = 6), but which were anesthetized daily for 6 days as described above. Unoperated control animals were similarly anesthetized daily (Unoperated no NIRWARP10 group, n = 6).
We also tested the VET 75 LED array, which was used to deliver NIR light to another group of animals each day for 6 consecutive days following partial ON transection. Treatment using the VET 75 array was administered for only 3 min per day (PT + NIRVET75 group, n = 6), thereby avoiding anesthesia, but delivering approximately 75% of the same light dose as the WARP 10 array per treatment. The rats were wrapped in a towel and held such that the head remained 3 cm under the surface of the array. The non-treatment groups therefore comprised animals receiving partial ON transection that were held but not exposed to NIR light (PT no NIRVET75 group, n = 6). Unoperated control animals were also similarly handled daily (Unoperated no NIRVET75 group, n = 3).
Behavioral assessments of visual function
Visual function of animals in the treatment and non-treatment groups was examined by assessing optokinetic nystagmus and Y-maze pattern discrimination after 6 days of NIR light treatment with the WARP 10 LED array and optokinetic nystagmus alone following NIR light treatment with the VET 75 LED array (Fig. 1).
Optokinetic responses
The uninjured left eye was closed with 6-0 silk suture under anesthesia on the day after the last NIR light treatment (day 7; Fig. 1), and again on day 27 (for the VET 75 array groups). Eyelid suture ensured that all responses were due to the visual ability of the experimental eye following partial ON transection with or without NIR light treatment. Optokinetic nystagmus is an accepted first-line screening test of visual function, and was conducted as described previously (Abdeljalil et al., 2005; Fitzgerald et al., 2009a). A normal optokinetic nystagmus comprises a smooth pursuit, with the head moving at the same speed as stripes inside a rotating drum, followed by a rapid realignment movement or fast reset phase (Cahill and Nathans, 2008; Hogie et al., 2008). The smooth pursuit and fast reset phases were analyzed separately by a single investigator blinded to the treatment identity of the animals (groups: PT + NIRWARP10, n = 8; PT no NIRWARP10, n = 6; Unoperated no NIRWARP10, n = 6, and PT + NIRVET75, n = 4; PT no NIRVET75, n = 6; Unoperated no NIRVET75, n = 3). Numbers of responses by animals receiving partial ON transection with or without NIR light using the WARP 10 array (8 days), or VET 75 array (8 and 28 days), were analyzed using one-way analysis of variance (ANOVA), and Dunnett's post-hoc test (significance value p ≤ 0.05). The analysis included responses from unoperated control animals receiving daily anesthetic (Unoperated no NIRWARP10, 8 days), and 3 min of daily handling (Unoperated no NIRVET75, 8 and 28 days).
Y-maze pattern discrimination task
The rats were habituated to handling for 5 consecutive days to minimize stress, and were then pre-trained for the 3 weeks prior to partial ON transection (Fig. 1). Food intake was limited for the duration of training and testing, with weight monitored daily to ensure animal health. The floor of the Y maze was covered with wood shavings identical to that in the subjects' housing. A laminated card displaying a visual stimulus was placed at the end of each arm. The visual stimuli consisted of laminated squares of paper (10 × 15-cm each), with either vertical black and white stripes at 0.25, 0.5, 0.75, or 1.0 cycle per degree (cpd), or a solid 50% grey square of the same luminance as the striped pattern. Correct choices were immediately rewarded by feeding the animal a treat, and after each attempt, the rats were returned to the starting position by hand for the next trial. Training consisted of 30–40 attempts daily. The location of the striped visual stimulus was randomly allocated to the arms before each trial, but kept constant for all rats. Individuals were scored on their choice after all four paws had crossed a line 30 cm from the visual stimulus. Subjects were deemed to have made a non-choice if they had not crossed a line after 60 sec. Accuracy (percentage of correct choices) was recorded and criterion performance was set at 80%. After each trial, the wood shavings were mixed in order to disrupt any olfactory information. The stripe size on the visual stimuli was decreased until the animals could reach criterion at 1.0 cpd.
Only rats that had reached criterion performance during pre-training were tested in the Y-maze pattern discrimination task (groups: PT + NIRWARP10, n = 5; PT no NIRWARP10, n = 4; Unoperated no NIRWARP10, n = 5). Testing was conducted for 3 days by investigators blinded to the treatment identity of the animals, starting on day 8 after partial ON surgery (Fig. 1). Up to three sets of 40 tests were conducted on each rat per day. Stripe sizes were decreased once the animal had reached criterion performance, until all animals had been tested at 1.0 cpd on day 10. One animal in group PT no NIRWARP10 did not reach criterion at 0.75 cpd, but was also tested at 1.0 cpd on day 10. The percentage of correct choices at 1.0 cpd on day 10 were recorded and analyzed using one way ANOVA, followed by Dunnett's post-hoc test (significance was set at p ≤ 0.05).
Immunohistochemistry
Immunohistochemical analysis was conducted on ONs at the injury site from 3–4 animals per group, following treatment with the WARP 10 array and behavioral assessments (10 days) (Fig. 1). After transcardial perfusion with 0.9% saline, then 4% paraformaldehyde in 0.1 M phosphate buffer (pH 7.2), the ONs were dissected and post-fixed in 4% paraformaldehyde overnight, cryoprotected by immersion in 15% sucrose in PBS overnight, embedded in tissue embedding medium, and cryosectioned transversely (16 μm) before mounting on microscope slides. If not processed immediately, the slides were stored at −80°C. ON sections at the injury site were air dried, rehydrated in PBS, and incubated overnight at 4°C in primary antibodies (diluted in PBS + 0.2% Triton-X100) for detecting: manganese superoxide dismutase (MnSOD, 1:500; Stressgen, San Diego, CA); astrocyte glial fibrillary acidic protein (GFAP, GA5, 1:500; Sigma-Aldrich, St. Louis, MO); β-III tubulin (TUJ1, 1:500; Covance Inc., Princeton, NJ); resident microglia/macrophages (Iba-l, 1:1000; Wako Chemicals USA, Inc., Richmond, VA); activated microglia/macrophages (ED-1, 1:500; Serotec, Raleigh, NC); oligodendrocyte precursor cells (OPCs, NG-2, 1:500; Abcam, Cambridge, MA); oligodendrocytes (olig-2, 1:250; Chemicon International, Temecula, CA); or myelin basic protein (MBP, 1:150; Abcam). Antibody binding was visualized following 2-h of incubation with anti-rabbit (Alexa Fluor 488, 1:400; Molecular Probes, Eugene, OR), anti-mouse (Alexa Fluor 546, 1:400; Molecular Probes), or anti-goat (for MBP, Alexa Fluor 488, 1:400; Molecular Probes) secondary antibodies. The slides were cover-slipped using Fluoromount-G (Southern Biotechnology Associates, Birmingham, AL), and viewed using fluorescence microscopy. All sections for each antibody were immunostained at the same time to ensure uniformity of immunohistochemical procedures, and the images were captured at constant exposure times. Images were viewed at low magnification in order to simultaneously visualize changes in the dorsal and ventral ON. Control sections stained only with secondary antibodies were included in all experiments, and showed no or only minimal fluorescence (data not shown).
Image intensity of MnSOD immunoreactivity in all astrocytes from a standardized area of ventral ON was quantified using ImageJ analysis software, by tracing around GFAP-immunopositive astrocytes and determining the intensity of MnSOD using dual-label immunohistochemistry. At least 20 astrocytes were analyzed for each animal. Total numbers of: MnSOD-immunopositive aggregates, Iba-1-immunopositive resident microglia/macrophages; ED-1-immunopositive activated microglia/macrophages, NG-2-immunopositive OPCs, and olig-2-immunopositive oligodendrocytes were counted in the dorsal and/or ventral halves of the transverse ON sections at the injury site, as appropriate. The injury site was identified by reference to adjacent sections stained immunohistochemically with β-III tubulin sections (data not shown). Numbers of cells in the dorsal or ventral halves of ON sections or image intensities were statistically analyzed using ANOVA and Bonferroni/Dunn or Dunnett's post hoc tests, requiring a significance value of p ≤ 0.05.
Electron microscopy
Ultrastructural analysis was undertaken for 3 animals per group, following treatment using the WARP 10 array (30 min/day for 6 days, assessed at day 10; Fig. 1). The ON containing the injury site was post-fixed in 1% osmium (ProSciTech, Kirwan, Queensland, Australia) for 90 min with shaking, and processed using a Lynx tissue processor into Araldite Procure mixture (ProSciTech). Embedded ON segments were cured for 24 h, serially cross-sectioned (1 μm) in three sets at 50-μm intervals, and stained with toluidine blue to identify the approximate injury site. Ultrathin sections were also cut, and low-power photographs of entire sections were taken to ensure identification of the lesion site. Three high-power images at 5800 × magnification were taken from central ON vulnerable to secondary degeneration, the images were saved as TIF files, and all axons within the images were assessed using ImageJ analysis software to determine the minimum axon diameter, axon area, and the minimum fiber diameter (i.e., including both the axon and the myelin sheath). At least 150 axons were assessed for each animal. The results were expressed as axon areas (μm2) and G ratios, and were analyzed using one-way ANOVA and Dunnett's post-hoc tests (significance value of p ≤ 0.05). G ratios were calculated by dividing the minimum diameter of each axon by the minimum diameter of the fiber, including both the axon and the myelin sheath.
Results
NIR light treatment with the WARP 10 LED array reduced secondary oxidative stress
The total NIR irradiance (550–750 nm) delivered by the WARP 10 LED array, reaching the ventral surface of the ON through the overlying tissue was 1.75 W/m−2 (approximately 0.7% of the light hitting the dorsal surface of the head). The total irradiance penetrating to the ON through the eyeball and orbit was 1.54 W/m−2 (0.6%), and through the central brain and skull was 0.3 W/m−2 (0.1%).
The effects on oxidative stress of NIR light treatment using the WARP 10 array were assessed in the ventral ON vulnerable to secondary degeneration at two time points after partial ON transection. First, in the early phase of oxidative stress 24 h after partial ON transection, when MnSOD has been shown to be increased in hypertrophic astrocytes (Fitzgerald et al., 2010), and second, 10 days after partial ON transection, when MnSOD is present in immunoreactive aggregates (Fig. 1).
Two NIR light treatments with the WARP 10 array, one immediately after partial ON transection and the second at 24 hours after injury and immediately before euthanasia, prevented early increases in MnSOD immunoreactivity in ventral astrocytes immunopositive for GFAP (24 h, PT + NIRWARP10 versus PT no NIRWARP10; p ≤ 0.05; Fig. 2). Values were not significantly different from those in unoperated controls (p > 0.05; Fig. 2). The high intensity of MnSOD in the dorsal ON precluded analysis at this time point.
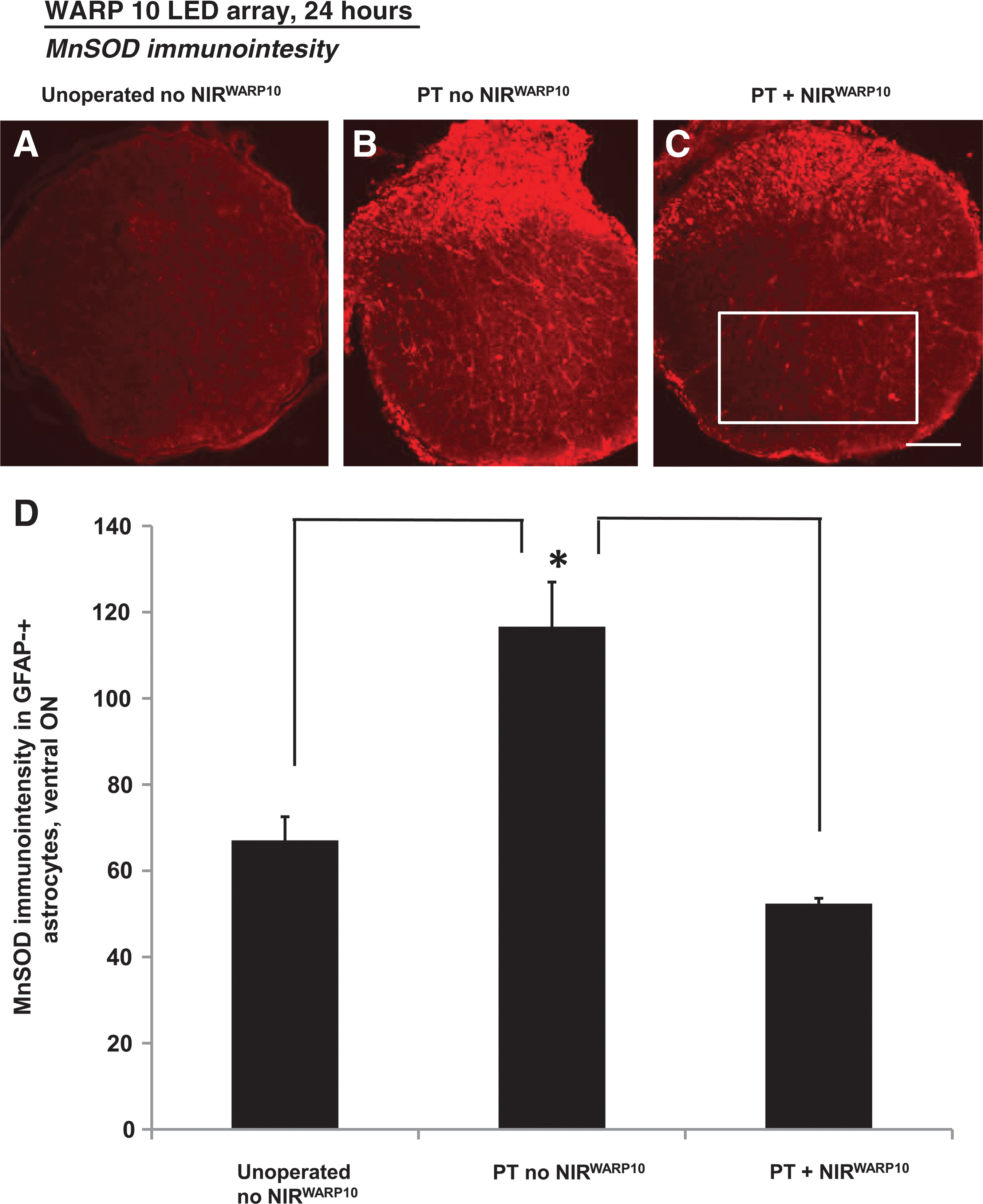
Representative images and quantification of optic nerve (ON) sections stained immunohistochemically for manganese superoxide dismutase (MnSOD) following partial ON transection at 24 h, with (
Ten days after partial ON transection, immunoreactivity of MnSOD in GFAP-immunopositive astrocytes of ventral ON was not above background and further analysis was not undertaken. However, as in previous studies (Fitzgerald et al., 2009a, 2010), partial transection resulted in significantly increased numbers of MnSOD-immunoreactive aggregates (not in astrocytes) in the ventral ON vulnerable to secondary degeneration (mean ± SEM: PT no NIRWARP10 = 7.0 ± 2.0, Unoperated no NIRWARP10 = 0.0 ± 0.0; p ≤ 0.05). NIR light treatment with the WARP 10 array (30 min/day for 6 days) reduced numbers of MnSOD-immunoreactive aggregates in the ventral ON to levels that were not significantly different from those in unoperated controls (mean ± SEM: PT + NIRWARP10 = 3.0 ± 0.82; p > 0.05). In contrast, 10 days after partial ON transection, the increased numbers of MnSOD-immunoreactive aggregates in the dorsal ON affected by the primary injury were not significantly altered by NIR light treatment (data not shown; p > 0.05).
NIR light treatment with the WARP 10 LED array did not affect secondary microglia/macrophage infiltration
Similarly to previous studies (Fitzgerald et al., 2009a, 2009b), partial ON injury (PT no NIRWARP10) resulted in loss of axonal profiles at the dorsal injury site, and diffuse axonal loss in the ventral ON vulnerable to secondary degeneration 10 days after injury, as demonstrated by β-III tubulin immunohistochemistry (data not shown). Diffuse axonal loss of β-III tubulin-immunopositive axonal profiles in the ventral ON was also observed following treatment with the WARP 10 array (data not shown).
We have previously demonstrated that partial ON injury results in increased numbers of ED-1-immunopositive activated microglia/macrophages and Iba-1-immunopositive resident microglia macrophages in the ventral ON vulnerable to secondary degeneration, by 7 days after injury (Fitzgerald et al., 2009a). We have confirmed and extended this result in the current study, demonstrating significantly increased numbers of both ED-1- and Iba-1-immunopositive cells in the ventral half of the ON at 10 days after injury, (p ≤ 0.05; Table 1). NIR light treatment with the WARP 10 array did not reduce the numbers of ED-1- or Iba-1-immunopositive cells in the ventral ON (PT + NIRWARP10 versus PT no NIRWARP10; p > 0.05; Table 1).
Numbers of ED-1-immunopositive activated microglia/macrophages and Iba-1-immunopositive resident microglia/macrophages were not altered with NIR light treatment using the WARP 10 LED array, 10 days following partial ON transection. Data are expressed as (mean ± standard error of the mean) numbers of immunopositive cells per ventral or dorsal half of the transverse section of the ON at the injury site or the equivalent, in animals subjected to partial ON transection with (PT + NIRWARP10) or without (PT no NIRWARP10) NIR light treatment with the WARP 10 LED array, for 30 min per day for 6 consecutive days, compared to unoperated control animals (Unoperated no NIRWARP10). Significant differences between numbers of ED-1- and Iba-1-immunopositive cells in the dorsal and ventral ON regions following partial ON transection, with or without NIR light treatment, compared to unoperated controls are indicated (*p ≤ 0.05).
NIR, near infrared; PT, partial transection; ON, optic nerve; LED light-emitting diode.
Similarly, 10 days after partial ON transection, numbers of both ED-1- and Iba-1-immunopositive cells were significantly increased in the dorsal ON (affected by the primary injury), but NIR light treatment with the WARP 10 array did not reduce the numbers of ED-1- or Iba-1-immunopositive cells in the dorsal half of the ON (PT + NIRWARP10 versus PT no NIRWARP10; p > 0.05; Table 1).
NIR light treatment with the WARP 10 LED array prevented secondary increases in NG-2-immunopositive OPCs
At 10 days following partial ON transection, the number of NG-2-immunopositive OPCs in the ventral ON vulnerable to secondary degeneration was increased (p ≤ 0.05; Fig. 3). NIR light treatment using the WARP 10 array significantly reduced the number of NG-2-immunopositive OPCs in the ventral ON (PT + NIRWARP10 versus PT no NIRWARP10; p ≤ 0.05) to values not significantly different from unoperated controls (p > 0.05; Fig. 3).
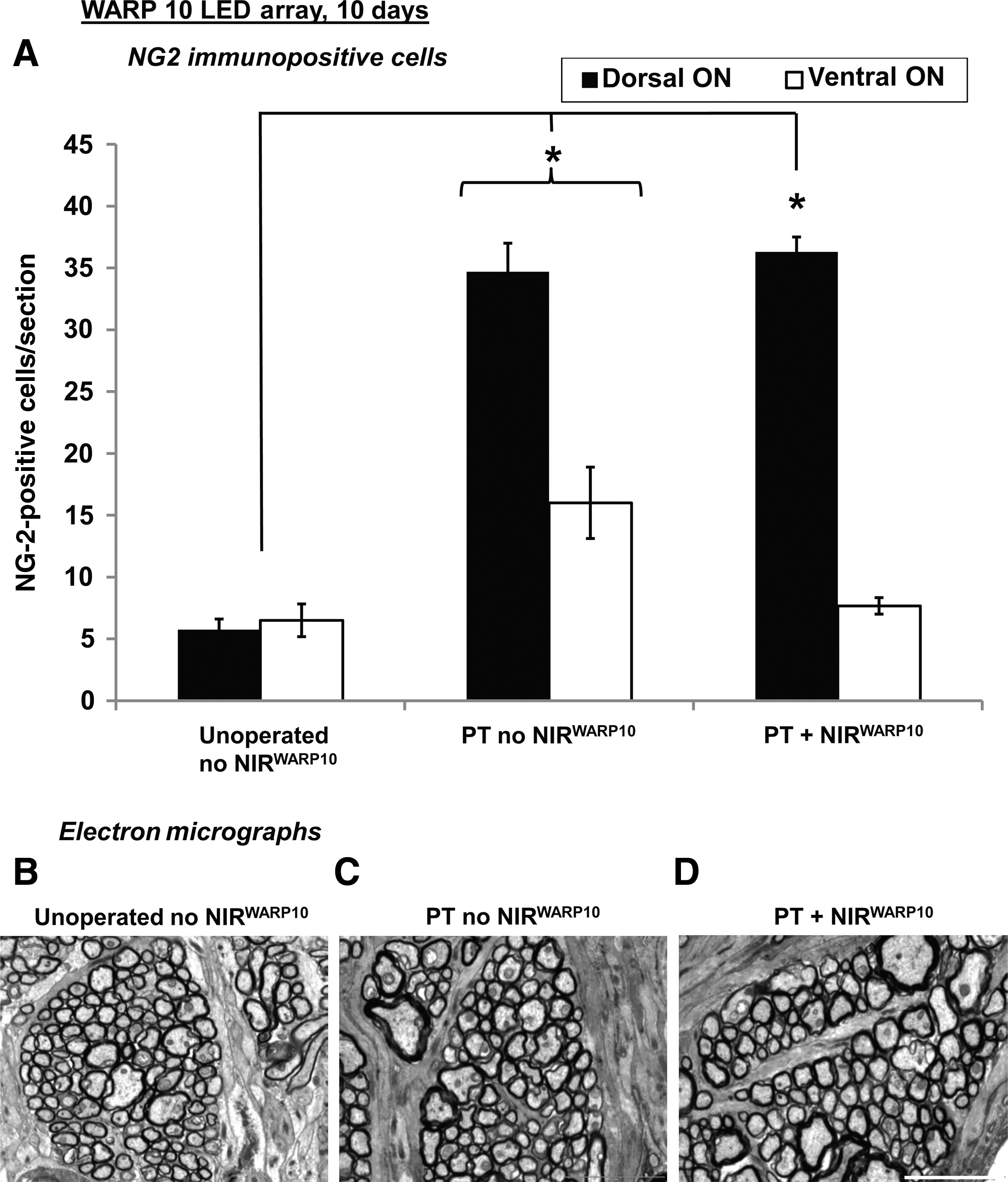
Similarly, at 10 days following partial ON transection, numbers of NG-2-immunopositive OPCs in dorsal halves of the ON were significantly increased (p ≤ 0.05; Fig. 3). However, NIR light treatment with the WARP 10 array had no effect on numbers of NG-2-immunopositive OPCs affected by the primary injury (PT + NIRWARP10 versus PT no NIRWARP10; p > 0.05; Fig. 3).
Ten days after partial ON transection, NIR light treatment using the WARP 10 array had no effect on axon diameter or G ratios in the central ON vulnerable to secondary degeneration, as assessed by ultrastructural analyses using electron microscopy (PT + NIRWARP10 versus PT no NIRWARP10; p > 0.05; Table 2, Fig. 3). However, it is important to note that the partial ON injury resulted in no significant change in axon diameter or G ratios at this relatively early time point after injury (PT no NIRWARP10 versus Unoperated no NIRWARP10; p > 0.05; Table 2, Fig. 3). Similarly, 10 days after partial ON transection, the numbers of olig-2-immunopositive oligodendrocytes in the ventral ON vulnerable to secondary degeneration were not significantly changed (PT no NIRWARP10 versus Unoperated no NIRWARP10; p > 0.05), nor were they affected by NIR light treatment using the WARP 10 array (PT + NIRWARP10 versus PT no NIRWARP10; p > 0.05; Table 2). The pattern of MBP immunoreactivity was similarly unaffected (results not shown).
Axon area, G-ratios of myelination, and numbers of olig-2-immunopositive oligodendrocytes in ON vulnerable to secondary degeneration were not changed 10 days following partial ON transection, with (PT + NIRWARP10) or without (PT no NIRWARP10) NIR light treatment with the WARP 10 LED array for 30 min per day for 6 consecutive days (p > 0.05). Results are expressed as mean ± standard error of the mean, and compared to unoperated control animals (Unoperated no NIRWARP10). Axon area and G-ratio values are the result of electron microscopic assessment of at least 150 axons per animal in a defined area of the central ON. G ratios were calculated by dividing the minimum diameter of each axon by the minimum diameter of the fiber, including axon and myelin. The numbers of olig-2-immunopositive oligodendrocytes were assessed in the ventral half of the transverse section of ON at the injury site or equivalent.
NIR, near infrared; PT, partial transection; ON, optic nerve; LED light-emitting diode; olig-2, oligodendrocyte.
NIR light treatment with the WARP 10 LED array restored visual function after partial ON injury
At 8 days following partial ON transection, the numbers of smooth-pursuit and fast-reset phases of the optokinetic nystagmus were reduced compared to unoperated controls (PT no NIRWARP10 versus Unoperated no NIRWARP10; p ≤ 0.05; Fig. 4A). However, following NIR light treatment using the WARP 10 array (30 min/day for 6 days), the numbers of smooth-pursuit and fast-reset phases at 8 days were not significantly different from unoperated controls (p > 0.05; Fig. 4A). Similarly, partial ON transection significantly reduced a rat's ability to make correct choices in the Y-maze pattern discrimination task (PT no NIRWARP10 versus Unoperated no NIRWARP10; p ≤ 0.05) at 10 days after injury. However, NIR light treatment using the WARP 10 array restored Y-maze discrimination to control values (Fig. 4B).
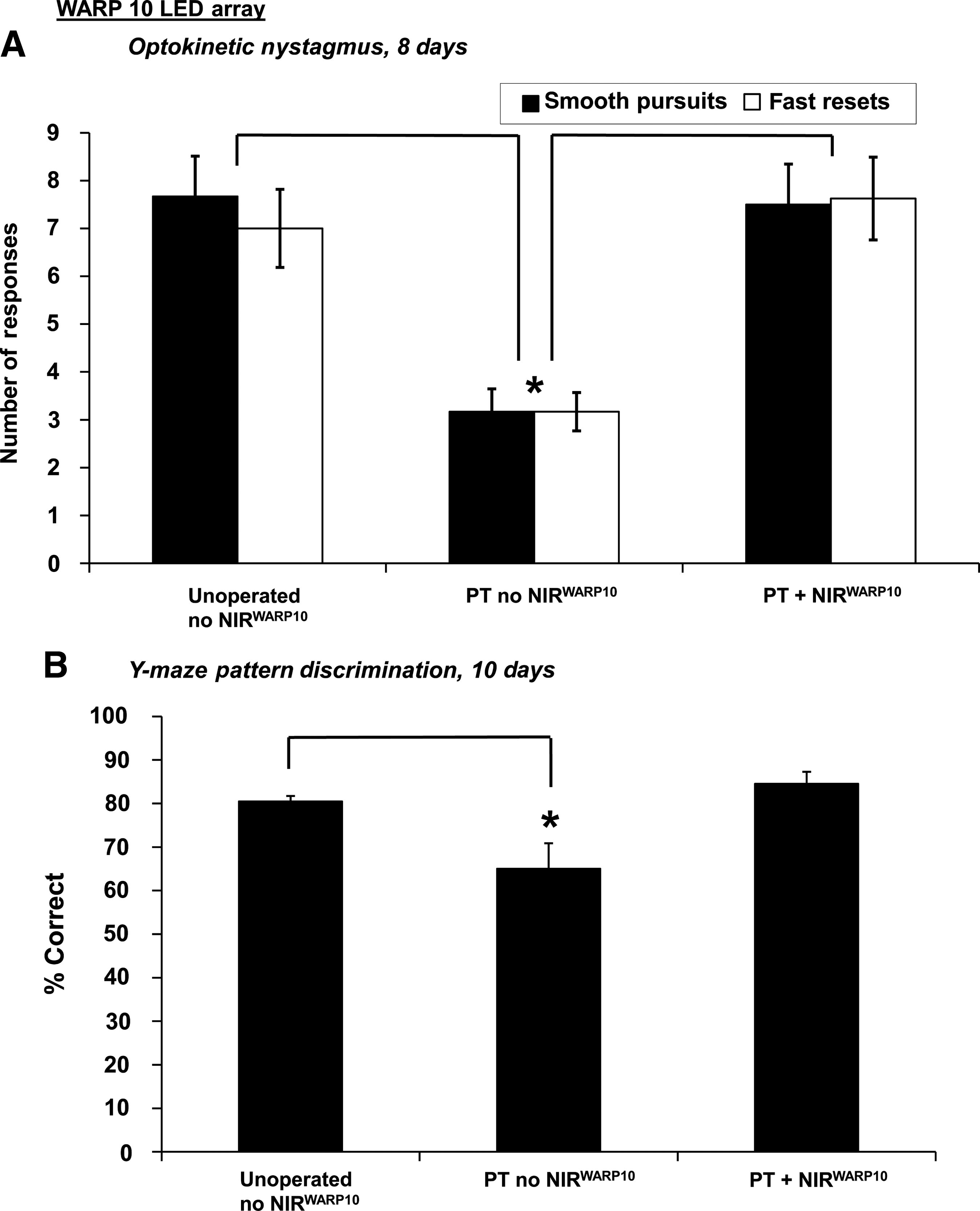
Optokinetic nystagmus and Y-maze pattern discrimination tests of visual function following partial ON transection and NIR light treatment using the WARP 10 LED array (30 min/day for 6 days). Optokinetic nystagmus
An alternate LED array delivering a more intense exposure (VET 75; 3 min/day for 6 days) was used in an effort to reduce treatment times and avoid repeated anesthesia, as well as to assess if approximately 75% of the light dose per treatment time was equally effective at restoring vision. At 8 days after partial ON transection, animals treated with the VET 75 array also had numbers of smooth-pursuit and fast-reset phases that were not significantly different from unoperated controls (PT + IRVET75 versus Unoperated no NIRVET75; p > 0.05). However, there were no significant improvements in the responses by animals treated with the VET 75 array, compared to untreated animals with partial ON transection, for either smooth pursuits or fast resets (PT + NIRVET75 versus PT no NIRVET75; p > 0.05; Fig. 5A). Furthermore, at 28 days the effect of treatment with the VET 75 array was not sustained, and compared to controls the numbers of smooth-pursuit and fast-reset phases were significantly lower with or without treatment using the VET 75 array (p ≤ 0.05; Fig. 5B). Due to the lack of a sustained behavioral effect of NIR light treatment with the VET 75 array, immunohistochemical analysis of ONs from these treatment groups was not conducted.

Optokinetic nystagmus tests of visual function following partial ON transection and NIR light treatment using the VET 75 LED array (3 min/day for 6 days). Optokinetic nystagmus was assessed at day 8
Discussion
In this partial injury study, we have demonstrated that treatment with NIR light (WARP 10 LED array, 670 nm) reduced oxidative stress in areas of ON vulnerable to secondary degeneration. NIR light treatment also prevented the secondary increases in NG-2-immunopositive OPCs that occurred as a result of partial ON transection. Importantly, visual function was restored to normal by NIR light treatment using the WARP 10 LED array, as assessed using optokinetic nystagmus and the Y-maze pattern discrimination task. To our knowledge, this is the first demonstration that 670-nm NIR light can reduce oxidative stress and improve function in the CNS after traumatic injury in vivo.
Improvements in visual function following partial ON transection were achieved with daily 30-min exposures to the WARP 10 array, for the 6 days immediately following injury. It is possible that the improvements in performance on the Y-maze pattern discrimination task were partially due to non-specific improvements in working memory resulting from NIR light treatment (Michalikova et al., 2008). Similarly, improvements in optokinetic nystagmus may be influenced by neuroprotective mechanisms not directly related to protection of the injured ON, perhaps involving transfer of visual responses to motor commands (Delgado-Garcia, 2000; Haustead et al., 2008). However, electroretinogram responses in completely normal rats were not altered with exposure to 670-nm NIR light LED arrays (Eells et al., 2003), indicating that our observed improvements in visual function following traumatic injury are likely to be a result of direct effects on the partially injured ON. Furthermore, our demonstration that 670-nm NIR light penetrates to the injured ON as well as through the retina to the back of the eye supports the premise of a direct effect of NIR light on the ON. Beneficial outcomes may also be due to direct effects of 670-nm NIR light on RGC somata in the retina, although we did not observe improvements in RGC axonal profiles.
Traumatic injury to the CNS results in glutamate excitotoxicity and altered Ca2+ flux, which leads to oxidative stress and reduced function of neurons and glia (Park et al., 2004; Tezel, 2006). We have recently demonstrated that oxidative stress spreads rapidly through ON vulnerable to secondary degeneration, through the linked astrocytic network (Fitzgerald et al., 2010). In this study, we demonstrated that NIR light treatment using the WARP 10 array reduced this early oxidative stress in hypertrophic astrocytes of ventral ON vulnerable to secondary degeneration in vivo, and also reduced the later accumulation of MnSOD-immunoreactive aggregates within cellular debris (Fitzgerald et al., 2009a), 10 days after injury. Our observed effects of NIR light treatment in reducing secondary oxidative stress after traumatic injury are likely to be due to increases in cytochrome c oxidase activity, reduced reactive oxygen species, and improvements in mitochondrial function, such as those described in a number of in vitro and in vivo studies (Liang et al., 2006; Rojas et al., 2008; Wong-Riley et al., 2005).
Reductions in oxidative stress in astrocytes may also protect oligodendrocytes in ventral ON vulnerable to secondary degeneration, with resultant beneficial effects on the integrity of myelin and visual function (Fitzgerald et al., 2009a; Nave, 2010; Park et al., 2004; Williams et al., 2007). However, due to the short-term nature of the current study, myelin was not yet sufficiently disrupted to enable detection of improvements in myelination following treatment with NIR light. Nevertheless, the increases in NG-2-immunopositive OPCs that occur in ON vulnerable to secondary degeneration were prevented with NIR light treatment using the WARP 10 LED array. Disruption of myelin causes increases in NG-2-immunopositive cells (Franklin and Ffrench-Constant, 2008). We therefore hypothesize that NIR light treatment may reduce oxidative stress in astrocytes, leading to protection of oligodendrocytes and myelin, thereby preventing increased numbers of NG-2-immunopositive OPCs in nerves vulnerable to secondary degeneration. Improvements in visual function would result from the preservation of myelin sheaths surrounding axons in areas of the ON vulnerable to secondary degeneration. NIR light treatment did not prevent reductions in β-III tubulin-immunopositive axonal profiles, or increases in resident or activated microglia/macrophages in the ventral ON vulnerable to secondary degeneration. However, visual function can be maintained with a small percentage of intact retinal ganglion cells (Sabel, 1999), and it may be that preservation of the myelin sheath is more important for preservation of function than large numbers of axons. Nevertheless, it is likely that combinatorial treatment strategies that protect axons as well as reduce oxidative stress in glia, such as NIR light combined with the CNS-specific neuroprotective calcium channel blocker lomerizine, will be required for even more effective prevention of secondary degeneration (Fitzgerald et al., 2009a, 2009b).
The relatively long duration of each treatment using the WARP 10 array necessitated repeated anesthesia of experimental animals, which can be problematic. Therefore assessing longer-term outcomes using this successful treatment regimen was beyond the scope of the current study. Treatment using a commercially-available LED array delivering greater irradiance (the VET 75 array), which provided a larger treatment area and potentially more thorough exposure to NIR light in a shorter time period, resulted in only minimal improvements that were not sustained. It is possible that the use of LED arrays with higher energy densities may enable reductions in treatment times.
Light in the red to NIR region of the spectrum has positive effects on a range of neural cells both in vitro and in vivo, when delivered by low-energy laser or LED (Eells et al., 2004; Liang et al., 2008; Rochkind, 2009; Rochkind et al., 1990, 2009; Wong-Riley et al., 2005). LED delivery of NIR light avoids some of the potential pitfalls of low-energy laser delivery, with flat arrays providing wider treatment areas and minimal heat production (Eells et al., 2004). It has been calculated that light emitted by LED arrays can penetrate tissue to a depth of 23 cm (Beauvoit et al., 1994; Whelan et al., 2001), and we have demonstrated penetration of 670-nm light over a distance of a few centimeters through skull, brain, and associated tissue around the ON. The use of these arrays has been deemed a non-significant risk for use in humans by the FDA (Desmet et al., 2006). Our study indicates that 670-nm NIR light offers a potential treatment modality for prevention of secondary degeneration following partial traumatic injury to the CNS, which may be particularly useful in combination with other neuroprotective agents. The use of NIR light LED arrays therefore represents a feasible alternative for the treatment of secondary degeneration following traumatic injury to the CNS.
Footnotes
Acknowledgments
This work was supported by the Neurotrauma Research Program (Western Australia), the Raine Foundation, Western Australia, and the NH&MRC (grant ID 572550). Prof. Dunlop is a Professorial Fellow (Research) and Senior Fellow of the NH&MRC, Australia (grant ID 254670). The WARP 10 LED array was a kind gift from Janis T. Eells. We thank Jervynna Teh and Michael Archer for assistance with technical procedures.
Author Disclosure Statement
No competing financial interests exist.