
Abstract
We report the results of controlled cortical impact (CCI) centered on the caudal forelimb area (CFA) of rat motor cortex to determine the feasibility of examining cortical plasticity in a spared cortical motor area (rostral forelimb area, RFA). We compared the effects of three CCI parameter sets (groups CCI-1, CCI-2, and CCI-3) that differed in impactor surface shape, size, and location, on behavioral recovery and RFA structural and functional integrity. Forelimb deficits in the limb contralateral to the injury were evident in all three CCI groups assessed by skilled reach and footfault tasks that persisted throughout the 35-day post-CCI assessment period. Nissl-stained coronal sections revealed that the RFA was structurally intact. Intracortical microstimulation experiments conducted at 7 weeks post-CCI demonstrated that RFA was functionally viable. However, the size of the forelimb representation decreased significantly in CCI-1 compared to the control group. Subdivided into component movement categories, there was a significant group effect for proximal forelimb movements. The RFA area reduction and reorganization are discussed in relation to possible diaschisis, and to compensatory functional behavior, respectively. Also, an inverse correlation between the anterior extent of the lesion and the size of the RFA was identified and is discussed in relation to corticocortical connectivity. The results suggest that CCI can be applied to rat CFA while sparing RFA. This CCI model can contribute to our understanding of neural plasticity in premotor cortex as a substrate for functional motor recovery.
Introduction
T
Non-human primate studies in premotor cortex following ischemic damage in primary motor cortex (M1) are especially relevant to this issue of plasticity in related areas within the motor cortex hierarchical network (Dancause et al., 2005; Liu and Rouiller, 1999). The hand representation in the ventral premotor area (PMv) expands after an ischemic lesion in the M1 hand area (Dancause et al., 2006b; Frost et al., 2003). In addition, corticocortical axons from the spared PMv hand area sprout and form novel connections with parietal somatosensory hand areas (Dancause et al., 2005).
It is less clear whether similar changes occur in rat motor cortex. Homologies between primate and rodent motor areas are not straightforward. For example, there are at least seven separate hand representations in the primate motor cortex, whereas only two have been identified in rodents. It is thought, however, that the caudal forelimb area (CFA) and the rostral forelimb area (RFA) of rodents are equivalent to the M1 hand area and the premotor area of primates, respectively (Nudo and Frost, 2006). Also, rat cortical motor areas exhibit not only intrinsic and intracortical connections comparable to those of non-human primates, but also similar structural relations among cortical and subcortical motor areas (Fang et al., 2005; Keller, 1993; Neafsey et al., 1986; Rouiller et al., 1993; Stepniewska et al., 2006). Taken together, it is reasonable to use rodent models of plasticity in spared cortical motor regions to investigate mechanisms of functional motor recovery after cortical injury.
A substantial body of evidence supports both structural and functional plasticity in spared motor structures after cortical lesions. However, the type of injury induction is critically important in the subsequent neural reorganization (Gonzalez and Kolb, 2003; Voorhies and Jones, 2002). For example, in a study by Gonzalez and Kolb comparing three models of permanent ischemia as well as aspiration lesions, behavioral effects were similar. But examination of dendritic branching and spine density demonstrated atrophy in some lesion models and hypertrophy in others (Gonzalez and Kolb, 2003). Neurophysiological changes in spared cortical areas also have been documented at least since the pioneering work of Glees and Cole, who used surface stimulation techniques in monkeys to show that the thumb representation re-appeared in the adjacent cortical location after focal damage performed by undercutting (Glees and Cole, 1949). Similar results have been demonstrated after focal ischemic lesions, though post-injury use of the impaired hand was necessary for the neurophysiological changes to occur (Nudo et al., 1996b). Expansion of premotor hand representations after ischemic lesions in the primary motor cortex hand area has also been shown (Frost et al., 2003).
After traumatic brain injury (TBI), functional plasticity, especially in the injured hemisphere, may be more limited due to the potential for more widespread disruption of axonal and dendritic processes, especially in corticocortical networks. However, neurophysiological studies of plasticity in spared areas after TBI have to date been relatively rare. In one notable study, Boyeson and associates used ICMS techniques in rats, similar to those used in the present study, after undercutting, suction ablation, or contusion injuries of limb representations, and found no evidence for re-emergence of the damaged representation (Boyeson et al., 1991). In fact, after 30–290 days post-contusion, hindlimb movements could not be evoked at all, suggesting that contusion injuries result in a slowly-evolving lesion, and that the substrates for recovery are located elsewhere in the brain.
The present study was designed to determine the feasibility of examining plasticity in spared cortical motor structures in the same hemisphere after a controlled cortical impact (CCI) in adult rats. Our primary goal was to develop a set of CCI parameters that results in restricted damage to the CFA, sparing neurons in the RFA, but producing chronic motor deficits. Since the RFA, or any other remote motor representation, was not specifically examined in the earlier work by Boyeson and associates (1991), this represents a unique test of the hypothesis that spared motor regions in the same hemisphere are still intact, and may functionally reorganize after contusion injuries. Determining the relationship between behavioral recovery and neurophysiological reorganization in spared motor structures in this model may provide insight into the underlying neural mechanisms for recovered functions. To this end, three sets of CCI parameters comparing the effects of subtle differences in impactor tip surface, size, and stereotaxic position, were analyzed. Specifically, we assessed the behavioral performance before and for 35 days after CCI, using contact placing (reflex) tests, the skilled reach task (single pellet retrieval), and footfault tests, and subsequently examined movement representations within RFA using ICMS.
The study confirmed chronic deficits in forelimb skilled behavior using each of the three CCI parameters and the functional integrity of the RFA. No obvious structural damage within the RFA was evident. The resulting size of the RFA representation area varied according to lesion proximity to the RFA.
Methods
Subjects and group assignments
Long-Evans hooded rats (n = 24; 300–400 g; Charles River Laboratory, Raleigh, NC) were procured at 4 months of age. All animal use was in accordance with National Institutes of Health regulations, and approved by the Institutional Animal Care and Use Committee of the University of Kansas Medical Center. During the first month after arrival, the rats were handled daily to acclimate them to human touch, and to habituate them to the new cage environment. Each rat was singly housed in a transparent cage and provided with food and water ad libitum. The room was kept on a 12-h:12-h light:dark cycle, and ambient temperature was maintained at 22°C.
Skilled reach (single pellet retrieval) task
A pellet-retrieval task (Whishaw et al., 1991) was used to assess the effects of CCI and subsequent recovery on forelimb motor behavior. The testing chamber was constructed of acrylic glass and measured 25 cm in length × 25 cm in width × 35 cm in height. A 1-cm-wide reaching slot, extending from the bottom of the chamber to 20 cm in height, was centered at the front of the chamber. A food shelf (4 cm deep × 25 cm wide) was secured to the outside of the chamber 3 cm from the bottom. A pair of indentations was made in the food shelf 2 cm from the front and 0.5 cm to the left and right side of the reaching slot, to hold 45-mg food pellets (Bioserve, Frenchtown, NJ). The rats were restricted to 15–20 g of rat chow at the end of each shaping or training day to increase their motivation to retrieve the pellets (Bury and Jones, 2002; Hsu and Jones, 2006). All sessions were videotaped with a camcorder.
Behavioral shaping
Shaping consisted of having rats reach out for single food pellets placed centrally (between the indentations), 1 cm from the front of the chamber. Thus the rats could retrieve pellets using either forelimb. They usually become proficient within 5 days. Forelimb dominance was then evaluated during 20 reaches in each of two consecutive daily 20-min sessions (Hsu and Jones, 2006). The dominant forelimb was defined in each rat as the limb used on more than 70% of the trials (Maldonado et al., 2008). A total of 18 rats displayed left dominance, while 6 displayed right dominance.
Skilled reach training
Once forelimb dominance was established, we began baseline training by compartmentalizing the chamber with a removable wall in order to constrain reaches to the dominant forelimb only. Each training session consisted of a maximum of 60 trials or 20 min per day (whichever came first). Training sessions were conducted 5 days per week for 2 weeks. A trial was counted as successful when the rat grasped and transported the pellet to the mouth. A failure was tallied when (1) a pellet was successfully grasped, but dropped before reaching the mouth, (2) a pellet was dislodged from the food shelf, or (3) the rat failed to contact the pellet after five reaches (e.g., the rat grasping air without contact with the pellet).
Pre-CCI baseline probe sessions
After training was completed, five probe sessions (two in the first week and three in the second week) were conducted to define baseline skilled reach performance. Lasting no longer than 15 min, the probe sessions were identical to the training sessions except only 20 trials were administered. The scores were reported as the number of successfully retrieved pellets.
Post-CCI probe sessions
After CCI, performance was assessed during probe sessions conducted weekly beginning on post-CCI day 7 and continuing through week 5.
Footfault task
The footfault task was used to measure locomotor ability separately for the forelimb and the hindlimb. The apparatus consisted of a plastic platform grid (57 cm in length × 44 cm in width) elevated 30 cm from the laboratory table. The grid had openings of 3.5 cm × 3.5 cm, and the grid rung diameter was 2 mm. The task required no prior training. In a probe session, each rat was placed on the platform and was videotaped from below for a minimum of 2 min (Gilmour et al., 2005) as it traversed the platform grid. Video recordings of forelimb and hindlimb footfaults were analyzed in slow motion. Footfaults were defined as the placement of the limb through a grid opening. We tallied: (1) the number of footfaults by each limb and (2) the total number of forelimb or hindlimb steps. A step was defined as the sequential movement of both the left and right limbs. We analyzed the dominant limb only (i.e., the impaired limb after CCI). Forelimb performance was quantified by the percentage of forelimb footfaults, the percentage of footfaults by the dominant forelimb divided by the total number of forelimb steps (Hsu and Jones, 2006). Hindlimb performance was calculated in the same manner. Baseline performance was defined during four pre-surgery probe sessions (two in the first week and two in the second week). Post-CCI performance was assessed during weekly probe sessions beginning on post-CCI day 7 and continuing through week 5.
Contact placing
A contact placing (reflex) test was administered only during days 1–3 post-CCI. The rat was held in a suspended position facing the corner of a flat surface. Then it was gently moved closer to the corner from the lateral side until either the vibrissae (vibrissae-forelimb placing) or the forelimb (forelimb-forelimb placing) made contact with the surface (Whishaw et al., 2004). For normal performance (lifting the forelimb ipsilateral to the side being stimulated before forced contact), a score of “1” was recorded. When the lifting response was not elicited within 3 sec, an unsuccessful score of “0” was recorded. The test was repeated 10 times with a maximum score of 10 for each side on each test day.
Controlled cortical impact (CCI) procedure
After the completion of baseline behavioral assessment, the rats were randomized to one of three CCI groups (CCI-1, CCI-2, and CCI-3), differing in impactor tip configuration, and a control group, consisting of rats of similar size and age that were not subjected to the surgical procedures (Table 1). Each CCI rat was anesthetized with an initial dose of 4% isoflurane and maintained with 3.5% isoflurane/30% oxygen through a mouthpiece, remaining under aseptic conditions until the end of the procedure. The animal was placed prone with its head position fixed in a stereotaxic frame. The skin over the skull was incised and a small hole (3–4 mm diameter) was made in the skull using a dental drill with a trephine bit over the cortex contralateral to the dominant forelimb. A total of 14 animals received CCI in the right hemisphere and 4 on the left.
L, lateral to midline; P, posterior to bregma.
The CCI device consisted of a linear motor, the impactor, (P01-23 × 80; LinMot, Inc., Zurich, Switzerland), and an electronic servo controller (E100-MT; LinMot, Inc.) as previously described (Bilgen, 2005; Onyszchuk et al., 2007, 2008). A stainless steel impactor tip was attached to the end of the linear motor. A three-dimensional motion manipulator (Kopf Instruments, Tujunga, CA) was used to precisely align the impactor tip with the stereotaxic coordinates corresponding to the CFA (Table 1). When using the larger impactor tips (CCI-2 and CCI-3), the stereotaxic coordinates were shifted posteriorly to avoid injury to the RFA, the focus of the electrophysiological experiments. The area of the craniotomy was constantly irrigated with sterile, physiological, room-temperature saline, while the underlying dura was kept intact. The impactor tip was slowly lowered through the craniotomy hole perpendicular to the cortex until it was in contact with the dura surface, as examined under light microscopy. The motion profile of the impactor tip was programmed to withdraw from the surface a distance of 20 mm, and then deliver a downward stroke of 22 mm (i.e., indentation depth of 2.0 mm), with a preset velocity of 1.5 m/sec, and indentation duration of 85 msec. The skin was pulled over the intact dura, then sutured back to complete the procedure and the anesthesia was terminated. Buprenorphine (0.6 mg/kg SC) was given as a post-surgical analgesic at least 1 h after the rat regained consciousness.
Intracortical microstimulation (ICMS) procedure
Two weeks following the final post-CCI behavioral assessment (i.e., 7 weeks post-CCI), an electrophysiological mapping procedure was performed in the frontal cortex ipsilateral to the CCI. For the purposes of this procedure, the animal was anesthetized with an initial dose of ketamine hydrochloride (70 mg/kg IP) and xylazine (5.0 mg/kg IP). Additional doses of ketamine (20 mg/kg IP) were administered as necessary to maintain a stable anesthetic level. The animal was placed prone with its head stabilized in a stereotaxic frame. After a craniotomy over the frontal cortex corresponding to the RFA and removal of the dura, a highly-magnified digital image of the cortex vasculature was taken to guide the points of microelectrode penetration. Using a computer graphics program (Canvas; ACDSee, Victoria, British Columbia, Canada), grid lines (250 μm apart) were overlaid on the image. Using a hydraulic microdrive, a NaCl-filled glass micropipette (tapered to 15–20 μm outer diameter tip; impedence = 500–700 kΩ) was lowered perpendicularly to the cortical surface at each grid intersection point until the tip reached the depths of cortical layer V (1700–1800 μm). A stimulus isolation unit (BAK Electronics, Mount Airy, MD) was used to deliver 200-μsec monophasic, cathodal constant-current pulses at the rate of 350 Hz for 40 msec (∼ 13 pulses). Joint and muscle movements elicited by the stimulation were inspected visually, and were considered reliable when observed in at least 50% of the ICMS trains. The minimum current required to elicit movement was recorded. The absence of visually detectable movements at the maximum current level of 80 μA was recorded as “no response.” Using custom and commercial software, the cortical surface areas representing each movement category (digit, wrist, elbow, shoulder, neck, and face) were calculated. Average current levels required to evoke each of the movement categories were also determined. Procedures identical to the above have been used in a number of experiments from our laboratory on both rodents and non-human primates (Kleim et al., 1998, 2002; Nudo et al., 1990; Nudo and Milliken, 1996).
Histology
Following the completion of the ICMS procedure, each animal was deeply sedated, then perfused via the left ventricle with 10% formalin solution. The brain was removed and stored in 10% formalin until it was ready for histology. To verify the lesion volume and location, the brain was first cryoprotected in 30% sucrose solution at 3°C for over 48 h. Serial 30-μm-thick frozen coronal sections of the lesion vicinity, from approximately 3.7 mm anterior to the bregma to 3.2 mm posterior to the bregma, were then collected using a cryostat (Paxinos and Watson, 2007). The sections were mounted on microscope slides and Nissl stained.
The caudal and rostral extent of the lesion was determined. Reconstruction of the lesion was done indirectly by tracing the perimeter of the cortices using the Cavalieri method in Stereo Investigator (Microbrightfield, Inc., Williston, VT); the lesion volume was estimated by the difference of the cortical volume in the injured hemisphere subtracted from the cortical volume in the intact hemisphere (Tennant and Jones, 2009).
Statistical analysis
SPSS version 17.0 (SPSS Inc., Chicago, IL) for Windows software was used for statistical analysis. One-way analyses of variance (ANOVAs) were used to analyze the histological and ICMS data, followed by Fisher's least significant difference post-hoc test when appropriate. A non-parametric test (chi-square test) was used to test the distribution of CCI severity with respect to callosal damage across groups. Due to a lack of normality as well as unequal variance, a non-parametric test (Kruskal-Wallis H test) was used to analyze the behavioral data for an overall group effect (two-tailed). In the case of significant group effects, Mann-Whitney U tests were used for pair-wise comparisons (two-tailed). A simple linear regression ANOVA was performed on ICMS results (RFA area) to determine if there was a significant relationship between the rostral and caudal extent of the impactor tip and the subsequent RFA area. Results for parametric tests are provided as mean ± SEM, and non-parametric tests in median ± 95% confidence interval. The minimum value for statistical significance was p ≤ 0.05.
Results
Histology
All animals were sacrificed approximately 7 weeks following CCI. Figure 1 illustrates a dorsal view of the lesion and coronal sections at the levels of the RFA (Fig. 1B), and the CFA (Fig. 1C–E). Histological inspection at the level of the CFA showed that in seven cases (CCI-1: n = 1; CCI-2: n = 3; CCI-3: n = 3) all cortical layers were destroyed, with the corpus callosum partially or completely intact (Fig. 1C). In five cases (CCI-1: n = 2; CCI-2: n = 1; CCI-3: n = 2), the deepest part of layer six was spared (Fig. 1D), and in another five cases (CCI-1: n = 2; CCI-2: n = 2; CCI-3: n = 1), the corpus callosum appeared severed due to direct impact (Fig. 1E). The presence or absence of damage in the corpus callosum was not related to CCI group (chi-square = 1.88, p = 0.758). At the level of the RFA, no cortical damage was evident under light microscopy in any of the cases. There was no statistically significant difference in lesion volume among the three impact groups (F
2,10 = 0.64, p = 0.54; Fig. 2A). However, while not examined quantitatively, there appeared to be a difference in lesion shape based on impactor tip shape. The flat tip (Fig. 1C) tended to produce more consistent columnar injuries compared than the rounded tips (Fig. 1D and Supplementary Fig. 1; Supplementary Data are available online at

(

Histological results. (
Because the rostrocaudal location of the impactor tip differed slightly between groups (CCI-1 was 0.5 mm more rostral, but its tip diameter was 0.25 mm smaller), the rostral and caudal extents of the histological damage relative to the bregma were also estimated (Fig. 2B). The rostral extent of the lesion did not differ across groups (F 2,10 = 1.24, p = 0.33). However, the caudal extent differed significantly (F 2,10 = 4.20, p = 0.047), measuring more anterior in group CCI-1 compared to groups CCI-2 and CCI-3. Although neurophysiological maps of the CFA were not derived, in all cases the lesion location corresponded to typical stereotactic coordinates for the CFA (Neafsey et al., 1986; Nudo et al., 1990).
Behavioral performance
Skilled reach task
To eliminate outliers from the skilled reach task analysis, the rats were required to have a baseline (pre-injury) skilled reach performance score of at least 10 successful trials out of 20. As a result, one rat from the control group and one rat from group CCI-1 were omitted from the skilled reach analysis, were but retained for all remaining analyses. As illustrated in Figure 3A, no group differences were detected in the mean number of successfully retrieved pellets on any of the pre-lesion assessment days. A significant group effect in forelimb function was evident on post-lesion days 7 (H = 11.4, p = 0.009), 14 (H = 10.0, p = 0.018), 21 (H = 11.3, p = 0.01), and 35 (H = 8.1, p = 0.044), but not on post-lesion day 28 (H = 6.9, p = 0.072). On post-lesion days 7, 14, 21, and 35, each of the three CCI groups displayed a lower number of successful retrievals than the non-lesioned control group. There were no significant differences among the three CCI groups on any given assessment day.
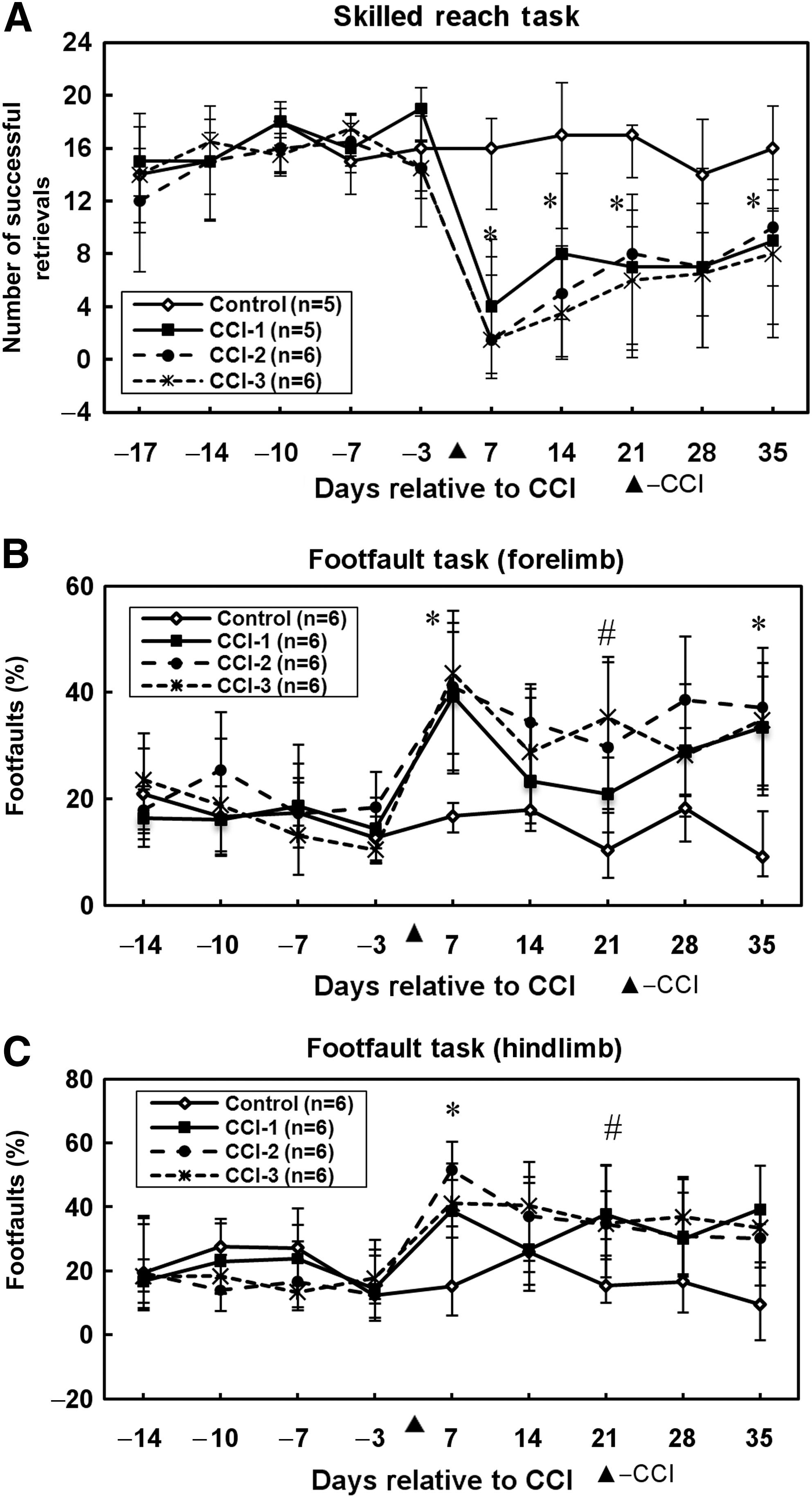
Behavioral results. (
Footfault task
In baseline sessions, all groups maintained equivalent forelimb footfault performance levels while traversing the elevated grid (Fig. 3B). Following CCI, a significant group difference in performance was found on post-lesion days 7 (H = 11.6, p = 0.009), 21 (H = 12.8, p = 0.005), and 35 (H = 11.6, p = 0.009), but not on post-lesion days 14 (H = 7.4, p = 0.061) or 28 (H = 4.5, p = 0.215). On post-lesion days 7 and 35, post-hoc tests revealed that the percent footfaults in each CCI group was higher than that of the control group. On post-lesion day 21, the percent footfaults of CCI-2 was higher than that of the control group, whereas the percent footfaults of CCI-3 was higher than that of both the control and CCI-1 groups.
As was described for forelimb footfaults, in the baseline sessions all groups maintained equivalent hindlimb footfault performance. After CCI, a significant group effect in hindlimb function was found on post-lesion days 7 (H = 9.9, p = 0.019) and 21 (H = 8.4, p = 0.038), but not on days 14 (H = 2.8, p = 0.422), 28 (H = 6.2, p = 0.10), or 35 (H = 5.7, p = 0.126; Fig. 3C). Post-hoc analysis revealed that on post-lesion day 7, each of the CCI groups made more footfault errors than the control group. On post-lesion day 21, groups CCI-2 and CCI-3 displayed higher percentages of footfaults than the control group, but group CCI-1 showed no significant difference compared to the control group. On this assessment day, however, the three CCI groups did not differ significantly from one another.
Contact placing test
The vibrissae-forelimb/forelimb-forelimb test was used to assess sensorimotor reflex function through the first 3 days post-lesion, prior to motor performance assessment, which began on post-lesion day 7. Only the non-lesioned control animals maintained contact placing with both limbs. The animals in all CCI groups showed a complete loss of contact placing for the limb contralateral to the cortical injury, scoring 0/10, and intact placing for the limb ipsilateral to the cortical injury, scoring 10/10 on each assessment day (data not shown).
Microstimulation mapping in the RFA
ICMS procedures were conducted in the intact RFA of the injured hemisphere in all animals 7 weeks following CCI. Digit and wrist movements evoked by ICMS were classified as “distal forelimb,” and elbow and shoulder movements as “proximal forelimb,” for both topographical area and movement threshold analyses. Topographic and threshold results were successfully obtained in 18 of 21 rats (5/5 control, 5/6 CCI-1, 4/4 CCI-2, and 4/6 CCI-3). Three rats died during the ICMS procedure. In the remaining 3 rats (one rat from group CCI-1 and two rats from group CCI-3), no evoked movements were observed from ICMS stimulation in the RFA. Such outcomes are not uncommon in ICMS experiments, and are typically attributable to improper anesthetic depth that cannot be corrected during the course of the procedure. As a result, quantitative neurophysiological analyses were based on a total of 18 rats. Representative topographic maps of the distal and proximal forelimb representations are shown in Figure 4A.
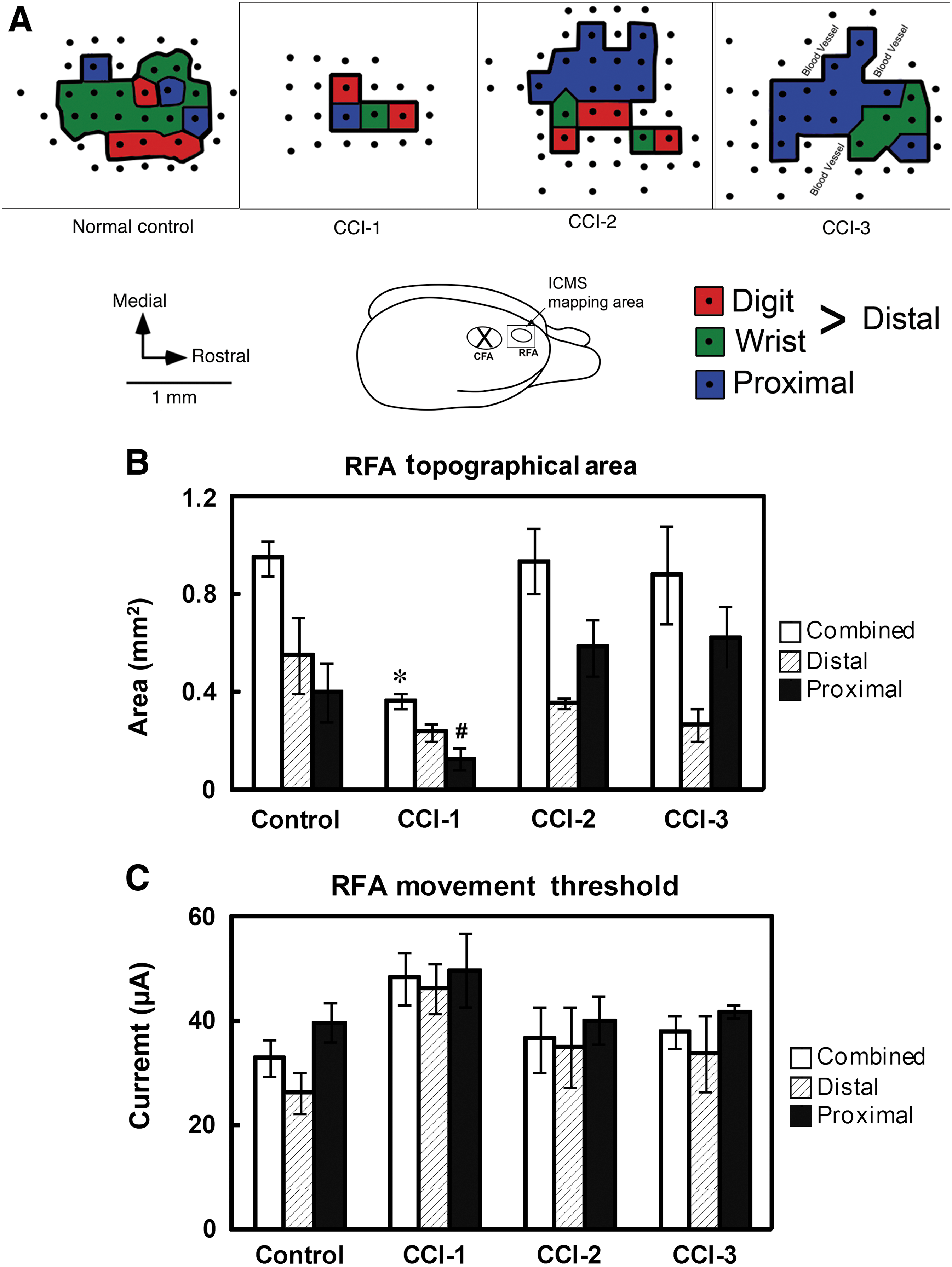
Neurophysiological results. (
The total RFA (combined distal and proximal RFA topographical area) was significantly different among groups (F 3,14 = 6.72, p = 0.005). Post-hoc comparisons revealed that the RFA was significantly smaller in group CCI-1 compared to each of the other groups (Fig 4B). The RFA size of neither group CCI-2 nor CCI-3 was different from that of the control group or from each other. When the total RFA was subdivided into distal and proximal areas, the proximal forelimb area showed significant differences across groups (F 3,14 = 4.62, p = 0.019). Group CCI-1 displayed a smaller proximal forelimb area than groups CCI-2 and CCI-3, but none of the CCI groups was different from the control group. However, while the mean distal area was smaller in each of the experimental groups compared with the control group, there was no significant group effect for the distal forelimb area (F 3,14 = 2.297, p = 0.122). It should be noted that this result may have been attributable to a single outlier. In one of the control rats, no distal forelimb movements were evoked, an outcome that is rare in RFA maps. When this single outlier was excluded from the analysis, the ANOVA showed a significant group effect (F 3,13 = 10.69, p = 0.0008), and pair-wise analysis showed a significant difference between the control group and each of the experimental groups.
No significant differences were found in the threshold currents to evoke forelimb movements for the combined (distal + proximal) RFA (F3,14 = 2.25, p = 0.128), distal forelimb (F3,13 = 2.07, p = 0.154), or proximal forelimb (F3,13 = 1.02, p = 0.414) movements. On average, however, group CCI-1 required 20 μA, 11 μA, and 13 μA higher currents than the control, CCI-2, and CCI-3 groups, respectively (Fig. 5C).
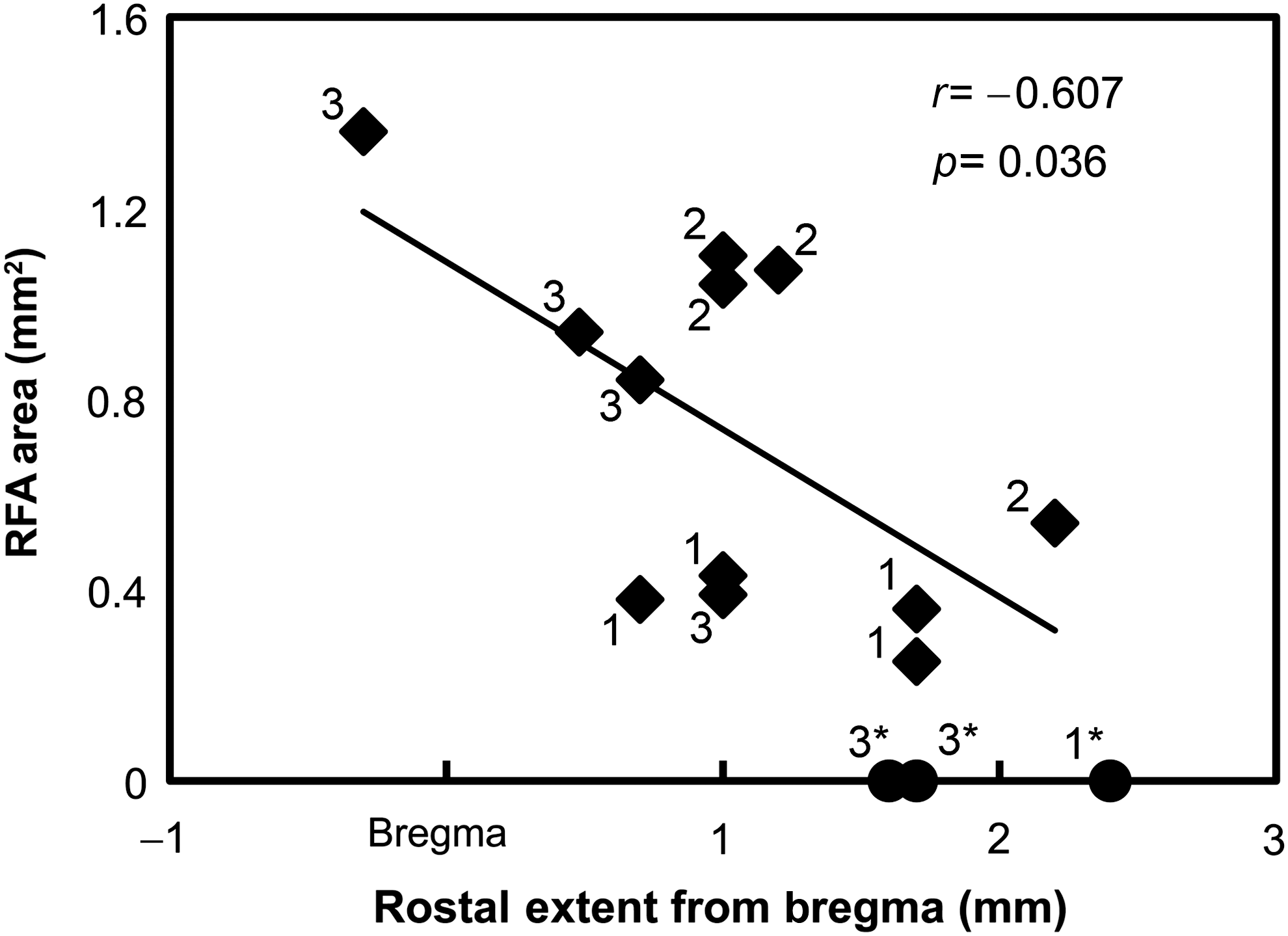
Neurophysiologically-based RFA area as a function of rostral lesion extent. The more rostral the lesion extended, that is, the closer the lesion was to the RFA, the smaller the RFA area obtained during the post-CCI mapping procedure (diamonds). Three cases in which we could not evoke movements (and therefore were excluded from the statistical analysis) are also illustrated (circle). Numbers indicate CCI groups (CCI, controlled cortical impact; RFA, rostral forelimb area).
A simple linear regression ANOVA demonstrated a significant negative relationship between the rostral lesion extent (based on histology) and the size of the RFA (F = 5.828, p = 0.036, r2 = 0.368; Fig. 5). In other words, the closer the lesion was to the RFA, the smaller the RFA topographical area. There was no relationship between the rostral lesion extent and the RFA threshold (F = 0.012, r2 = 0.001, p = 0.914).
Discussion
We aimed to develop a set of CCI parameters in rats that would result in long-term functional deficits in forelimb use while sparing the premotor forelimb area. We examined three sets of CCI parameters that differed in impactor tip surface shape, size, and location, targeting the caudal forelimb area (CFA). The cortical injury created by the CCI device was consistently reproduced across rats, resulting in a tissue cavity in the CFA and adjacent cortical areas, though some variability was observed in cortical depth and white-matter involvement. We focused our neurophysiological studies on the rostral forelimb area (RFA), since it is a cortical region similar in many respects to the premotor cortex in primates (Nudo and Frost, 2006). The main findings were that (1) all three CCI groups displayed motor deficits throughout the 5-week post-CCI test period, (2) the RFA was structurally intact in every rat, (3) the RFA was functionally intact in most rats, though there were significant changes in movement representations, and (4) the functional responsiveness of the RFA to cortical microstimulation was related to the anterior extent of the injury.
Effect of CCI parameters on lesion volume and behavioral deficits
The three sets of impactor tip parameters were statistically indistinguishable in lesion cavity volume and motor deficits. This result is not surprising in light of a previous study that found no group differences in lesion volume or behavioral deficits after contusion injuries, using an even wider range of impactor tip sizes, but with an impact depth similar to that used in our study (Whishaw et al., 2004). In both rat and mouse CCI models, tissue damage and severity of behavioral deficits are generally related to impact depth (Dixon et al., 1991; Feeney et al., 1982; Mao et al., 2010; Saatman et al., 2006), rather than tip diameter. However, impactor tip shape is also thought to be an important factor (Mao et al., 2010). In the present data, while there was no difference in injury volume, the shape of the damage was somewhat non-uniform with the rounded tip, and a more consistent column of damage was seen with the flat tip.
Behavioral deficits after CCI in the rat motor cortex
Among the widely implemented assessments in rat TBI studies are tasks that measure postural and locomotor behaviors, such as beam walking and balancing (Dixon et al., 1991; Feeney et al., 1982; Goldstein, 1993; Soblosky et al., 1996). In the present study, we primarily assessed forelimb motor behavior, since we were interested in characterizing deficits resulting from a focal impact injury in the CFA, the rodent equivalent of the primary motor hand area in primates. The skilled reach task is one of the most sensitive indicators of forelimb motor deficits, even during the chronic post-injury period (Adkins and Jones, 2005). In addition, the footfault test is a sensitive measure of both forelimb and hindlimb locomotor function after CCI, though typically only forelimb footfaults are examined (Grossman and Stein, 2000).
All animals sustained skilled reach (pellet retrieval) and forelimb footfault impairments for the entire 35 days of the post-CCI assessment time period. A more transient hindlimb footfault deficit was observed through post-lesion day 21. This differential effect on forelimb function is probably due to the fact that the CCI was centered on the CFA, though it is possible that some damage to the hindlimb motor representation occurred as well, since the hindlimb representation is located in close proximity to the CFA. While the impairments were significant, skilled forelimb behavior was not completely abolished. Even in the early stages after CCI, the rats could still achieve 20–40% success on this task, and were able to traverse the grid. Even though the initial loss in forelimb performance steadily corrected, the CCI animals never reached baseline performance, indicating persistent and perhaps chronic forelimb deficits. Our results are similar to those of a CCI study by Whishaw and associates, who also found chronic deficits in forelimb use (Whishaw et al., 2004). Based on their CCI parameters, the contusion likely included not only the CFA, but also the RFA (see Fig. 1 in Whishaw et al., 2004). The similarity in outcome indicates that sparing the RFA did not preclude the chronic deficits, and that a more focal CCI that spares the RFA can result in a similar consequence for skilled forelimb use.
There is a comparatively larger number of studies examining the effects of ischemic lesions. While a direct comparison between ischemic and contusive lesions has not been made, it is likely that deficits following CCI would be somewhat more severe. A recent study from our laboratory using endothelin-1 injections (Fang et al., 2010) to produce focal ischemic infarcts in approximately the same cortical territory of the CFA, resulted in a skilled reach success rate of about 50% 1 week after the lesion (lesion-only control group), compared with about 20% success in the present study. Whether this difference in severity is related to greater neuronal death with CCI, axonal shearing of corticocortical fibers, or subcortical trauma has yet to be identified.
Reorganization of motor maps in the RFA after CCI in the rat motor cortex
The present results demonstrate that CCI injuries can be confined to the CFA while leaving the RFA structurally and functionally intact. No obvious structural damage was evident in any of the CCI rats, at least when examined at the light microscopic level using Nissl stains. Forelimb movements in the RFA were evoked in response to ICMS with normal threshold currents that were comparable to previous ICMS studies in intact rats (Kleim et al., 1998; Neafsey et al., 1986; Nudo et al., 1990). These results at 7 weeks post-CCI parallel those of Boyeson and associates (Boyeson et al., 1991), who demonstrated return of ICMS thresholds to normal values in the adjacent, intact cortex within 15 days. The present results suggest that this model can be effective in examining post-injury reorganization in the more remote premotor cortex following focal CCI.
The size of the RFA representation, as defined by ICMS, was consistent across normal intact control rats, and similar to the size of the RFA reported in previous ICMS articles (e.g., Kleim et al., 1998). However, group CCI-1 showed a reduction in total (combined distal and proximal) forelimb area of more than 60%. This reduction occurred in both proximal (69% reduction) and distal (57% reduction, though not statistically significant) representations. In contrast, the combined forelimb representation was unaffected in groups CCI-2 and CCI-3. However, when a rare outlier was eliminated from the control group, groups CCI-2 and CCI-3 showed reduced distal representations. Thus an overall reduction in forelimb representation was evident in group CCI-1, while a redistribution of distal and proximal representations may have occurred in groups CCI-2 and CCI-3.
The smaller size of the RFA in CCI-1, and possibly higher currents required to evoke movement, were likely due to the difference in CCI parameters. The impactor tip in group CCI-1 was placed 0.5 mm more rostral. Although the tip diameter was 0.25 mm smaller, more rostral lesions may have introduced more direct or indirect damage to the RFA.
Redistribution of forelimb movement representations in spared motor areas: A case for behavioral compensation?
An important question for understanding mechanisms of recovery after CCI is whether altered movement representations in spared motor regions underlie functional recovery. Following a cortical injury, animals adjust the kinematics of forelimb movements to compensate for deficits in the affected musculature, often resulting in both proximal forelimb and postural compensation (Whishaw et al., 2004). Compensatory use of the proximal musculature is also commonly observed in humans after stroke (Cirstea and Levin, 2000). Functional outcomes improve over time, but true recovery may be masked (Whishaw, 2000; Whishaw et al., 1991), or even hindered (Alaverdashvili et al., 2007, 2008), by the use of alternative movement strategies (Levin et al., 2009).
In the rat CFA, as well as in the monkey primary motor cortex, motor skill training induces an expansion of distal forelimb representations at the expense of proximal representations (Kleim et al., 1998; Nudo et al., 1996a). In the present study, after CCI in the CFA, it is reasonable to hypothesize that improved motor behavior on the skilled reach task was related to functional changes in the RFA. While there was substantial variability in the component movement representations in the ICMS forelimb maps, the results suggest a redistribution of movements in groups CCI-2 and CCI-3 from distal to proximal. Therefore, if RFA plasticity formed the basis for motor recovery after CCI in the present study, and if motor skill acquisition drives the topography of motor maps, RFA plasticity likely supported compensatory motor strategies, rather than recovery of the original movement patterns. A more detailed behavioral analysis of pellet-retrieval strategies would be required to resolve this issue.
Structural and functional reorganization is not limited to spared regions of the injured hemisphere, but may occur in homotopic regions of the intact hemisphere as well (Jones and Schallert, 1992). However, Jones and colleagues have provided substantial evidence that structural changes in both homotopic and heterotopic areas of the intact, contralateral cortex are related to hyper-reliance on the intact limb, rather than recovery of the impaired limb (Allred et al., 2008; Bury and Jones, 2002; Chu and Jones, 2000). Human neuroimaging studies have also repeatedly shown bihemispheric changes in activation patterns after stroke. However, the functional significance of increased activity in the intact hemisphere is still subject to intense debate. It is not yet clear whether changes in fMRI patterns represent an adaptive, maladaptive, or ephiphenomenal effect (Nowak et al., 2009).
Role of corticocortical circuitry in remote effects after CCI
Diaschisis, classically defined, is reduced function in an area remote from, but connected to, an injured brain area (Feeney and Baron, 1986; von Monakow, 1914). To ascertain whether changes in RFA topography could be the result, at least in part, of diaschisis, it is important to understand normal corticocortical connectivity patterns in rats. Similarly to the relationship between the PMv and M1 in primates (Dancause et al., 2006a), the RFA and CFA are reciprocally interconnected (Rouiller et al., 1993), and more importantly, CFA is the principal target of corticocortical fibers from the RFA. Our preliminary anatomical studies suggest that projections are not equally distributed across the CFA, as the projection from the RFA to the more rostral portion of the CFA is particularly dense (Bury and Nudo, unpublished data). Thus, a focal CCI in the CFA should damage a significant proportion of corticocortical axons originating in the RFA (and reciprocal connections), but more rostrally-located lesions would result in differentially more damage.
In the present study, pooling the CCI cases demonstrated a significant inverse relationship between the rostral extent of the injury and the resulting size of the RFA map. As discussed earlier, the RFA maps were smallest in group CCI-1, the group with a more rostral impactor tip. In 3 of 6 cases (in all groups) in which the rostral extent of the injury was between 1.5 and 2.5 mm relative to the bregma, no movements could be evoked using ICMS. These physiological results suggest that while the RFA remained structurally and functionally intact in general, its integrity was increasingly compromised as lesions became more proximal. While a quantitative assessment of cell loss in the RFA after CCI in the CFA has not yet been done, it is reasonable to assume that as lesions edge closer to the rostral forelimb representation, some proportion of RFA neurons underwent retrograde degenerative changes and may not have survived.
Previous studies assessing axonal (Hall et al., 2008; Yaghmai and Povlishock, 1992), and dendritic (Posmantur et al., 1996), cytoskeletal structures after CCI found that the acute degenerative effect of CCI spread beyond the contusion's immediate surroundings in a rostral-caudal direction within the injured hemisphere. Hall and associates documented a continuous increase in the volume of degenerating axons that emanated at least 3 mm rostrally and caudally from the contusion epicenter, between 24 h and 7 days post-CCI (Hall et al., 2008). Morphological observations showed that a number of swollen, bulbous, and disconnected axons became widespread in the injured hemisphere on day 7 post-CCI (Chen et al., 2003). In addition, Posmantur and colleagues found MAP2 immunoreactivity losses in the ipsilateral cortex, indicating widespread sublethal responses resulting in disassembly of apical dendrites from pyramidal layers, which affected the function of dendrites leading to synaptic transmission dysfunction (Posmantur et al., 1996).
Spared motor areas as targets for therapy after TBI
Sprouting of axons in the peri-lesional cortex has been documented during early post-CCI (Harris et al., 2010) and post-ischemic (Carmichael et al., 2005) periods. In addition, it has been suggested that spared motor areas participate in recovery (Dancause, 2006; Liu and Rouiller, 1999). At least in non-human primates, corticocortical axons sprouting from spared premotor areas after M1 ischemic lesions form novel connections with parietal somatosensory hand areas (Dancause et al., 2005). While the CFA and RFA play different roles in movement control in intact rats (Barth et al., 1990), it is possible that the RFA is particularly plastic in its structure and function following CFA injury. After electrolytic lesions in the CFA, RFA plasticity has been demonstrated with a form of ICMS that uses long-duration trains of stimulation to elicit complex movements (Ramanathan et al., 2006). In rats that were not rehabilitated, the RFA map of complex movements was similar to that of intact rats. Meanwhile, in rats that received rehabilitative training on a pellet-retrieval task, the RFA map significantly expanded. Combined with the present results, it would appear that the RFA may be an important substrate for restorative therapies after TBI in rats.
Footnotes
Acknowledgments
We thank Dr. Yong-yue He for performing the CCI surgical procedures, and Dr. Pei-chun Beard for her technical assistance in histology. The work was supported by Department of Defense-Army Research Grant W81XWH-08-1-0168 (to R.J.N.), National Institutes of Health Grant NS030853 (to R.J.N.), and the University of Kansas Summer Research Fellowship (to M.N.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.