
Abstract
There is growing evidence that neck pain is common in adolescence and is a risk factor for the development of chronic neck pain in adulthood. The cervical facet joint and its capsular ligament is a common source of pain in the neck in adults, but its role in adolescent pain remains unknown. The aim of this study was to define the biomechanics, behavioral sensitivity, and indicators of neuronal and glial activation in an adolescent model of mechanical facet joint injury. A bilateral C6–C7 facet joint distraction was imposed in an adolescent rat and biomechanical metrics were measured during injury. Following injury, forepaw mechanical hyperalgesia was measured, and protein kinase C-epsilon (PKCɛ) and metabotropic glutamate receptor-5 (mGluR5) expression in the dorsal root ganglion and markers of spinal glial activation were assessed. Joint distraction induced significant mechanical hyperalgesia during the 7 days post-injury (p < 0.001). Painful injury significantly increased PKCɛ expression in small- and medium-diameter neurons compared to sham (p < 0.05) and naïve tissue (p < 0.001). Similarly, mGluR5 expression was significantly elevated in small-diameter neurons after injury (p < 0.05). Spinal astrocytic activation after injury was also elevated over sham (p < 0.035) and naïve (p < 0.0001) levels; microglial activation was only greater than naïve levels (p < 0.006). Mean strains in the facet capsule during injury were 32.8 ± 12.9%, which were consistent with the strains associated with comparable degrees of hypersensitivity in the adult rat. These results suggest that adolescents may have a lower tissue tolerance to induce pain and associated nociceptive response than do adults.
Introduction
M
An adult rat model of controlled tensile loading to the cervical facet joint has been developed that induces facet capsular ligament stretch that is similar to its loading during painful neck injury (Dong et al., 2008, 2010; Lee and Winkelstein, 2009; Lee et al., 2008). In that model, transient facet joint loading that does not produce ligament rupture is sufficient to induce behavioral hypersensitivity that lasts for up to 4 weeks in the adult rat (Rothman et al., 2008). Using this rat model, vertebral distractions across the C6–C7 joint of 0.7 mm produce strains in the capsular ligament of 24%, as well as sustained spinal glial activation, spinal cytokine upregulation, activation of the neuronal stress response in the dorsal root ganglion (DRG), and modifications in the glutamatergic system in the spinal cord (Dong and Winkelstein, 2010; Dong et al., 2008; Lee et al., 2008; Winkelstein and Santos, 2008). Although previous studies with that model of painful injury in the adult rat suggest a relationship between joint distraction, severity of loading, and behavioral and inflammatory responses, it is not known how that loading paradigm affects behavioral and/or cellular phenomena in an adolescent rat model.
In this study we tested the hypothesis that the mechanical tissue loading imposed in an adult model of painful facet joint injury could be scaled to an adolescent model, in order to investigate age-specific cellular and molecular responses to painful cervical neck injury. Post-natal day 35 rats were used to model a young, early-adolescent human that approximates the epidemiological rise in neck pain seen at around 13–14 years of age (Leboeuf-Yde and Kyvik, 1998). Facet capsule and joint biomechanics were quantified and forepaw mechanical hyperalgesia was measured to assess behavioral outcomes following the scaled facet joint distraction. Metabotropic glutamate receptor-5 (mGluR5) and protein kinase C-epsilon (PKCɛ) activation have been shown to enhance behavioral responses to noxious stimuli and increase the excitability of primary nociceptive afferents in models of pain (Velázquez et al., 2007; Walker et al., 2001a, 2001b). In particular, administration of group I-receptor (mGluR1 and mGluR5) agonists provoke thermal and mechanical hypersensitivity in both mice and rats (Bhave et al., 2001; Dogrul et al., 2000; Hama, 2003; Karim et al., 2001). In addition, mGluR5 has been reported to increase excitability of primary afferents, modulate nociceptive neurotransmission, and be positively related to tissue injury and behavioral sensitivity (Dong and Winkelstein, 2010; Pitcher et al., 2007). Similarly, PKCɛ is found in primary afferent neurons, and has been implicated in several pathological pain states in adult animals (Ahlgren and Levine, 1994; Amadesi et al., 2006; Dina et al., 2000; Ferreira et al., 2005; Khasar et al., 1999; Souza et al., 2002), and can regulate G-protein coupled receptors, such as mGluR5 (Conn and Pin, 1997; Schoepp et al., 1999). Accordingly, in this study, both PKCɛ and mGluR5 expression were evaluated in afferent neurons at the level of injury at day 7 after cervical facet joint distraction. At that same time point, astrocytic and microglial activation in the spinal cord were assessed as markers of glial activation due to injury.
Methods
Surgical procedures
Male Holtzman rats (Harlan Sprague-Dawley, Indianapolis, IN) at post-natal age 35 days (P35), weighing 136–174 g, were housed under U.S. Department of Agriculture (USDA)- and Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC)-compliant conditions with free access to food and water. All experimental procedures were approved by the University of Pennsylvania Institutional Animal Care and Use Committee, and were carried out under the guidelines of the Committee for the Research and Ethical Issue of the International Association for the Study of Pain (Zimmerman, 1983). All efforts were made to minimize the number of animals used and their suffering.
A controlled cervical bilateral facet joint distraction was applied across the C6–C7 cervical facet joint to stretch the capsular ligament, using methods that have been previously described for adult rats (Lee and Winkelstein, 2009; Lee et al., 2008). All surgical procedures were performed under inhalational isoflurane anesthesia (4% for induction, 2.5% for maintenance). Briefly, the rats were placed in a prone position and an incision was made in the skin to expose the C4–T2 spinal levels. The C6 and C7 spinous processes, laminae, and facet capsules were carefully isolated and the ligamentum flavum and interspinous ligament at C5–C6, C6–C7, and C7–T1 were transected to enable rigid attachment of the C6 and C7 spinous processes to a custom loading device via two pairs of microforceps. Capsule stretch injury was imposed as the C6 microforceps were translated rostrally away from the stationary C7 vertebra that was held fixed, imposing tensile distraction across the bilateral C6–C7 joints. As such, using the relative bone and joint sizes of a mature rat and the P35 rat, the 0.7-mm joint distraction used in the adult model (Lee et al., 2004b, 2006, 2008; Lee and Winkelstein, 2009), was scaled to a distraction of 0.4 mm in the P35 rat, with the goal of imposing a comparable magnitude of tissue strain across the facet joint capsule. For these studies, a joint distraction of 0.4 mm was imposed and held for 30 sec, after which the C6 vertebra was unloaded and returned to its initial position (injury group, n = 6). Sham procedures (sham group, n = 4) were performed in a separate group of rats as surgical controls, and involved surgery and attachment to the device, but these animals had no joint distraction. After surgery, the incisions were closed with sutures and surgical staples, and the rats were allowed to recover in room air while being monitored.
Mechanical data were acquired during joint distractions to quantify the vertebral and facet joint loads, relative joint motions, and tissue strains, in order to define the severity of joint injury for each case. The loads and motions imposed across the joint were measured by transducers attached to the customized loading device. Image data also were continuously acquired throughout the distraction period to track the motions of both the bony vertebrae and the facet capsular ligament of the joint. Microspheres (diameter 0.17 ± 0.01 mm; Spherotech, Inc., Libertyville, IL) were placed at the midpoint of the C6 and C7 laminae, and a grid of markers covered the right C6–C7 facet capsule. Marker motions were used to quantify joint displacements and ligament strains (Dong and Winkelstein, 2010; Dong et al., 2008; Lee et al., 2004a, 2004b, 2008). The maximum tensile strain and maximum principal strains in the facet capsular ligament were quantified as measures of ligament injury severity using the marker displacements and LS-DYNA software (Livermore Software Technology Corp., Livermore, CA).
Mechanical hyperalgesia assessment
Behavioral sensitivity was assessed by measuring forepaw mechanical hyperalgesia at post-operative days 1, 3, 5, and 7. Each rat was evaluated for 3 days prior to surgery to define baseline unoperated responses. Mechanical hyperalgesia was assessed using a modified version of Chaplan's up/down method (Chaplan et al., 1994; Decosterd and Woolf, 2000; Hubbard and Winkelstein, 2005; Lee et al., 2008). For each assessment session, the rats were allowed to acclimate for 20 min prior to testing. A single tester performed all testing and was blinded to the surgical procedures. Response thresholds were measured for each of the right and left forepaws by stimulating each forepaw with increasing von Frey filament strengths (Stoelting, Wood Dale, IL), ranging from 0.6–26.0 g (Lee and Winkelstein, 2009; Lee et al., 2008). The lowest-strength filament needed to provoke any response to five stimulations was recorded as the response threshold if the next larger filament strength also elicited a positive response. A positive response was defined as a forceful shaking and/or lifting of the paw, which was sometimes accompanied by licking the affected paw. If the rat was unresponsive to all filaments, the maximum filament strength (26.0 g) was taken as the threshold. Each testing session consisted of three rounds of stimulation with the filaments of increasing strength applied to each forepaw, with at least 10 min between each round to allow for an adequate rest period. The thresholds determined by the three rounds were separately averaged for each forepaw to obtain a single value for that forepaw for each rat in each session. A paired t-test was used to test for symmetry between the left and right forepaw responses for each rat following surgery. A repeated-measures analysis of variance (ANOVA) with Bonferroni post-hoc correction was used to compare injury and sham responses.
Immunohistochemistry procedures
The bilateral DRGs at C6 and C7 were harvested after behavioral testing on day 7 to assess mGluR5 and PKCɛ expression. Matched DRGs were also harvested from naïve unoperated rats (naïve group, n = 2), and were included in all analyses to serve as controls. The rats were deeply anesthetized with sodium pentobarbital (65 mg/kg), and transcardially perfused with phosphate-buffered saline (PBS) and formalin. DRG samples were removed and post-fixed on ice in 4% formalin for 4 h, and then rinsed with PBS three times at 30-min intervals. Samples were embedded in embedding compound and serially sectioned at 10 μm on a cryostat. Every fourth section was thaw-mounted onto slides, with 4–5 sections per slide, and sections spanning the entire DRG tissue from the capsule to the inner DRG and then back to the capsule. The slides were washed three times in Tris-buffered saline (TBS) for 10 min for each wash (8 mM Trizma base, 41.9 mM Tris HCl, and 154 mM NaCl). Antigen retrieval was performed by incubating the slides in sodium citrate buffer (10 mM tri-sodium citrate and 0.05% Tween-20, pH 6.0) for 30 min in an 80°C water bath. The slides were washed three times in TBS for 10 min each, and blocked in TBS + (40 μL normal horse serum + 2 μL Triton X-100/1 mL TBS) for 30 min at room temperature, followed by an overnight incubation in either anti-PKCɛ antibody (1:1000; Santa Cruz Biotechnology, Inc., Santa Cruz, CA) at room temperature, or anti-mGluR5 antibody (1:1000; Millipore, Billerica, MA) at 4°C. The next day, the slides were washed three times with TBS and incubated with Alexa-Fluor 488 donkey anti-rabbit secondary antibody (1:200; Invitrogen, Carlsbad, CA) for 3 h at room temperature in the dark. The slides were then washed three times with TBS for 10 min each, and quick-rinsed with diH2O, and the cover-slipped with Vectashield (Vector Laboratories, Burlingame, CA). Controls included immunostaining DRG sections as above without the primary antibody. Without the primary antibody (PKCɛ or mGluR5) there was no staining in the tissue.
Each DRG section was imaged with a fluorescent microscope (Leica, Wetzlar, Germany) and digital camera (Diagnostic Instrument Spot RT Color, Sterling Heights, MI). The exposure times were standardized for all of the samples. The PKCɛ sections were imaged with a 4-sec exposure, and the mGluR5 sections were imaged with an 8-sec exposure. The samples were analyzed by an experimenter who was blinded to surgical procedure. Three tissue sections from the C6 DRG and three tissue sections from the C7 DRG were analyzed from each rat and for each antibody. The three sections that were analyzed were chosen from the 4–5 tissue sections on each slide as the three sections that were from the inner DRG. In each DRG section all neurons, as identified by cellular shape under phase-contrast microscopy, were measured for diameter by taking an average of the length and width of the cell. Only neurons with a visible nucleus were included, and at least 100 neurons were included from each section. Neurons were classified as small (4–20 μm) or medium (22–40 μm). This resulted in a total count of small- and medium-diameter neurons in each DRG section. Since the DRGs were serially sectioned, any two sequential DRG sections on a given slide represented neurons that were a minimum distance of 40 μm apart. A secondary count of the number of PKCɛ- or mGluR5-immunoreactive small- and medium-diameter cell bodies was performed. A neuron was determined to be labeled for PKCɛ or mGluR5 in a qualitative manner by comparing it to the amount of fluorescence in the surrounding unlabeled neurons and control tissues. The percentage of small- and medium-diameter cell bodies that was positive for PKCɛ or mGluR5 was then quantified from the tissue-specific count of total small- and medium-diameter cells.
While a cell count demonstrates if additional DRG neurons are newly expressing PKCɛ and mGluR5, it does not determine whether there is a change in the amount of PKCɛ and mGluR5 that is being expressed within any given cell. Densitometry provides a semi-quantitative method to determine how much PKCɛ and mGluR5 is being expressed within a certain cell. ImageJ software (National Institutes of Health) was used to quantify the density of PKCɛ and mGluR5 immunoreactivity in each of these different sized neurons; in this way the amount of staining per area for small- and medium-sized PKCɛ- and mGluR5-immunoreactive neurons was quantified. A random sampling of 10 small- and 10 medium-diameter neurons from each section (n = 3 C6 sections and n = 3 C7 sections per rat) was used for the analysis of density. Expression of mGluR5 and PKCɛ between groups was compared using an ANOVA with Bonferroni post-hoc test.
Spinal cord tissue was also analyzed to assess astrocytic and microglial activation at day 7 after injury. Spinal cord samples at the caudal-most region of the C5 spinal segment were post-fixed in 4% formalin, transferred to 30% sucrose, and stored for 3 days at 4°C before being mounted in embedding medium and frozen for axial cryosectioning. Serial spinal cord sections (20 μm) from each rat were prepared for free-floating immunohistochemical staining using glial fibrillary acidic protein (GFAP) for astrocytic reactivity, and OX-42 (CD11b/c) as a marker of microglial reactivity. To stain for astrocytic activity, the slices were blocked for 20 min in goat serum (Vector Laboratories), and incubated in primary antibody containing rabbit anti-GFAP (1:20,000; Dako, Carpinteria, CA) overnight at 4°C. The next day, the sections were incubated in goat anti-rabbit secondary antibody (1:200; Vector Laboratories) for 30 min. For OX-42 staining, the samples were blocked using horse serum (Invitrogen) for 20 min, followed by incubation overnight at 4°C in a primary antibody containing mouse anti-rat CD11b/c (1:500; BD Biosciences, San Jose, CA). The following day, the samples were incubated in horse anti-mouse secondary antibody (1:1200; BD Biosciences) for 30 min.
Both GFAP and OX-42 staining were developed using 3,3-diaminobenzidine (Vector Laboratories). Images were taken of the dorsal horn from each rat at 100 × magnification using a digital camera and stereomicroscope with Axiovision software (Zeiss, Thornwood, NY). Each image was cropped to focus on the superficial laminae of the dorsal horn, and analyzed using densitometry to quantify the percentage of pixels above a threshold determined for expression in naïve un-operated samples. Expression of spinal GFAP and OX-42 were separately compared between groups using an ANOVA with a Bonferroni post-hoc test.
Results
Biomechanics
Based on observation, no obvious differences were seen in head mobility and grooming behavior for any of the rats undergoing surgery. In addition, the rats exhibited weight gains of 6–7 g/d post-operatively. The mean vertebral distraction measured in the injury group was close to the targeted joint distraction magnitude of 0.40 mm (0.42 ± 0.06 mm), with a corresponding average force generated across the joint of 1.10 ± 0.42 N (Table 1). The associated mean capsular ligament distraction was 0.26 ± 0.07 mm, which corresponded to an average maximum tensile strain in that tissue of 25.4 ± 10.5%. The average maximum principal strain sustained in the capsule at the peak distraction was 32.8 ± 12.9%, and was significantly higher than the maximum tensile strain (p < 0.02). There was no evidence of capsular ligament failure or tissue rupture during distraction for any rat undergoing distraction.
Weight = weight of the rat at the time of surgery; Peak force = force measured at peak joint distraction; Reference length = separation of the C6–C7 joint prior to distraction; Vertebral distraction = amount of distraction imposed across the vertebrae of the C6–C7 joint; Average capsule distraction = amount of distraction imposed across the facet capsule; Maximum principal strain = largest strain component measured on the capsule; Maximum tensile strain = largest strain value measured in the direction across the joint.
SD, standard deviation.
Mechanical hyperalgesia
Tactile sensitivity in the forepaw was induced by facet joint distraction in the injury group (Fig. 1). There was no significant difference in the responses measured in the right and left forepaws, so measurements from both forepaws were averaged for each rat. Sham procedures did not alter sensitivity from the baseline thresholds at any time point after surgery (Fig. 1). However, injury did produce a reduction in threshold as early as day 1, that was sustained at each post-operative time point (Fig. 1). This increase in sensitivity after injury was significant compared to both sham and matched baseline uninjured responses (p < 0.001).
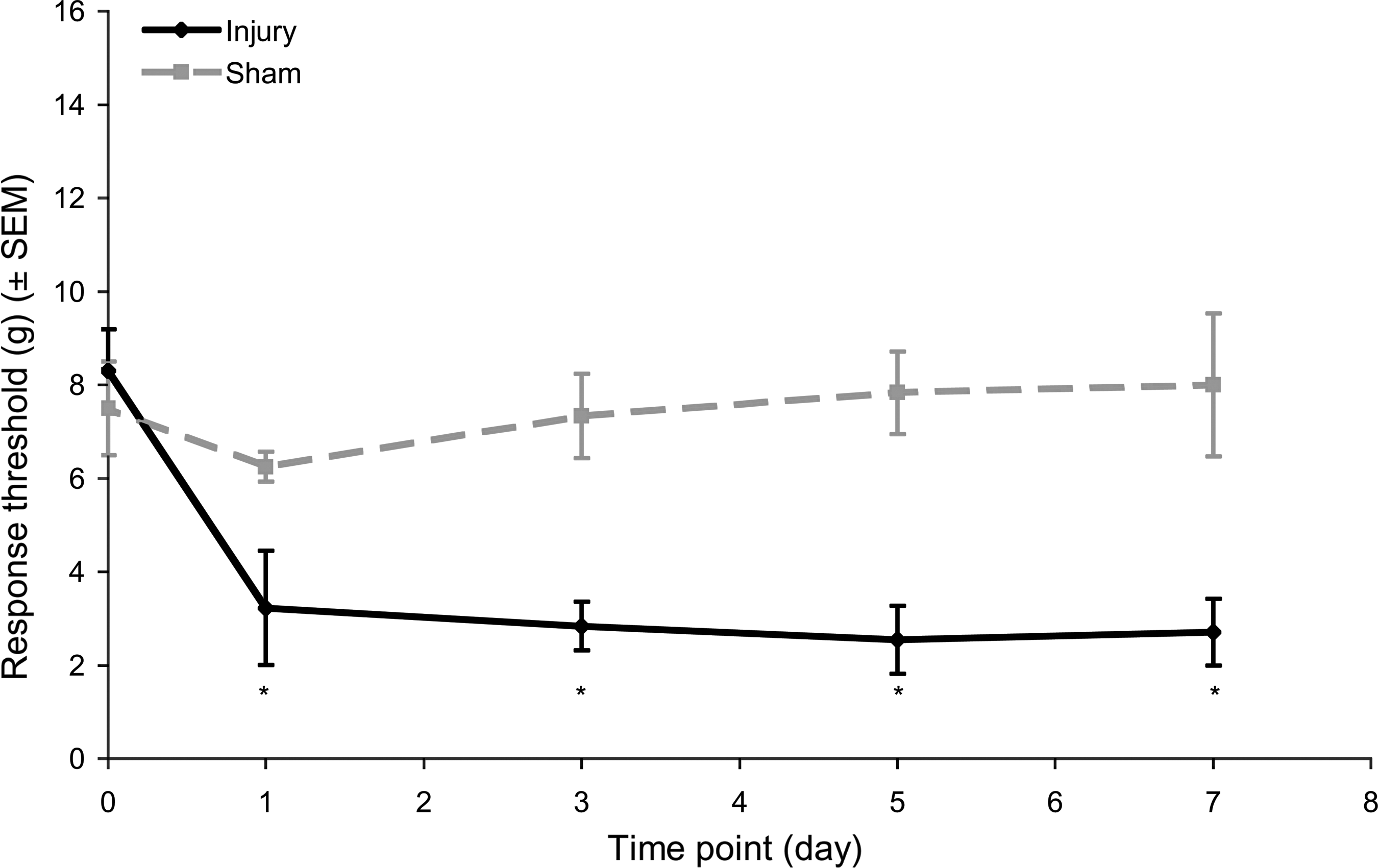
Mechanical hyperalgesia in the forepaw as measured by the average threshold in response to von Frey filament stimulation. Increased sensitivity corresponds to a reduced response threshold. Injury significantly reduced thresholds below those of sham animals for all post-surgical testing days (*p < 0.001). Data shown are average ± standard error of the mean.
DRG PKCɛ and mGluR5 expression
PKCɛ expression in DRG neurons at day 7 after injury paralleled the behavioral responses (Fig. 2). The percentage of small-diameter neurons immunoreactive for PKCɛ was significantly higher after injury than in sham (p < 0.05) and naïve (p < 0.001) animals (Fig. 2). Similarly, the density of PKCɛ immunoreactivity within small-diameter neurons was also significantly higher after injury compared to sham (p < 0.05) and naïve (p < 0.001) animals. In contrast, there was no significant difference in the number of medium-diameter neurons expressing PKCɛ following sham or injury. However, there was an increase in the density of PKCɛ immunoreactivity in medium-diameter neurons following injury compared to sham (p < 0.05) and naïve (p < 0.001) animals. Sham procedures did increase the number of small-diameter neurons immunoreactive for PKCɛ, and the density of PKCɛ immunoreactivity in small-diameter neurons compared to naïve animals (p < 0.01; Fig. 2).

Representative images and quantification of PKCɛ in the DRG at day 7. Injury (I) significantly increased the percentage of positive cells and density of PKCɛ in both small- and medium-diameter neurons over sham (S) (φp < 0.05) and naïve (N) (***p < 0.001) animals. Sham procedures also increased the percentage of positive cells and density of PKCɛ in small-diameter neurons over naïve animals (**p < 0.001). Data shown are average ± standard error of the mean (PKCɛ, protein kinase C-epsilon; DRG, dorsal root ganglion).
Expression of mGluR5 at day 7 in the DRG was less robustly modified by injury than was PKCɛ expression (Fig. 3). There were no differences between sham and naïve animals for the percentage of DRG neurons expressing mGluR5, or the density of mGluR5 immunoreactivity. However, there was a significant increase in the number of small-diameter neurons immunoreactive for mGluR5 compared to sham (p < 0.05) and naïve (p < 0.001) animals (Fig. 3). There was no difference in the percentage of medium-diameter neurons immunoreactive for mGluR5, or in the density of mGluR5 immunoreactivity, in small- or medium-diameter neurons following injury compared to sham or naïve animals.

Representative images and quantification of mGluR5 in the DRG at day 7. Injury (I) significantly increased the percentage of cells positive for mGluR5 in small-diameter neurons over sham (S) (φp < 0.05) and naïve (N) animals (***p < 0.001). Data shown are average ± standard error of the mean (mGluR5, metabotropic glutamate receptor-5; DRG, dorsal root ganglion).
Spinal glial activation
Spinal astrocytic and microglial activation in the dorsal horn at day 7 increased following injury (Figs 4 and 5). GFAP expression following injury was significantly elevated over both sham (p < 0.035) and naïve (p < 0.0001) levels (Fig. 4). Yet OX-42 expression after injury was only significantly elevated over naïve levels (p < 0.006; Fig. 5). There were no differences in expression of either marker between sham and naïve animals.
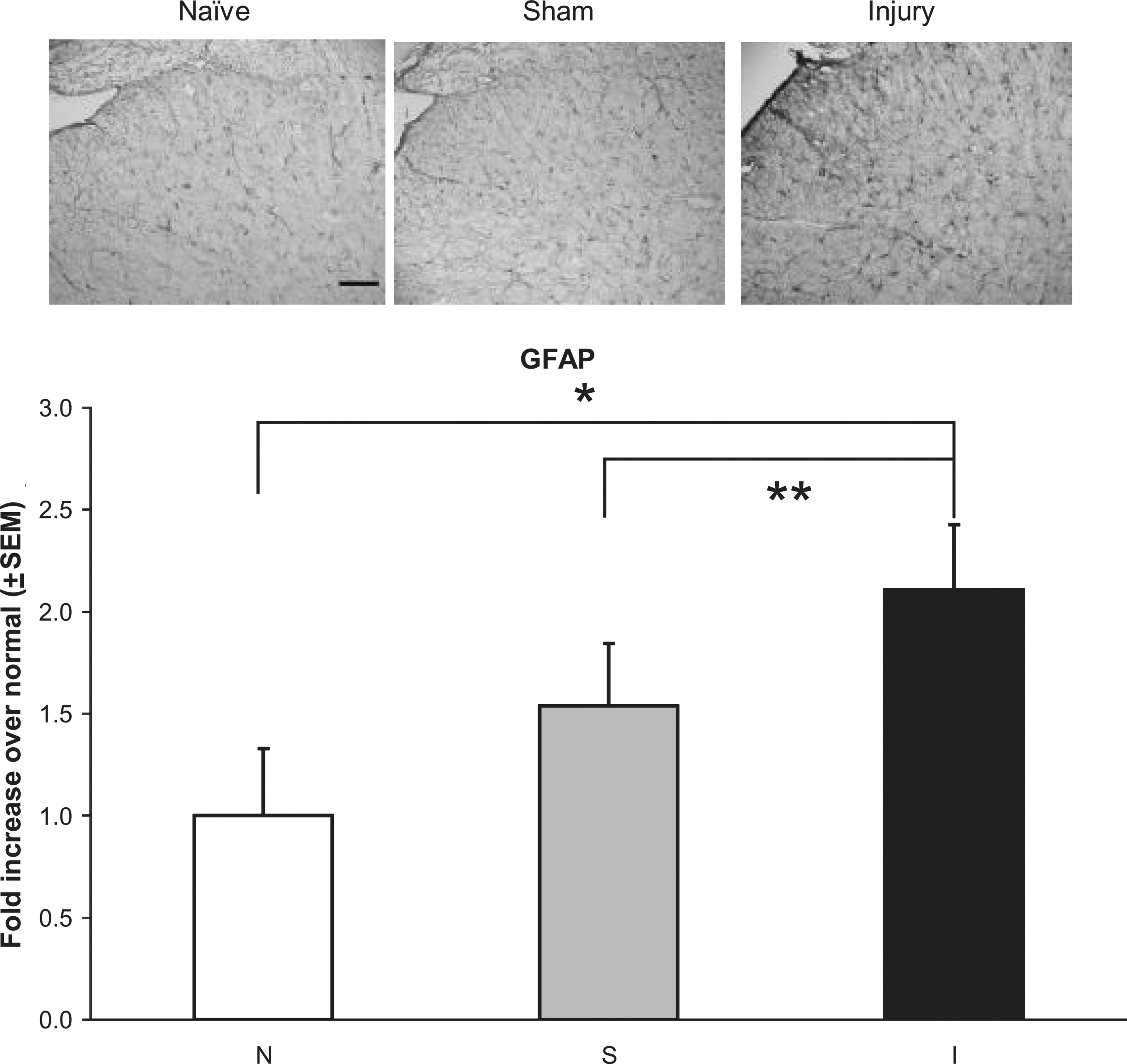
Representative images and quantification of GFAP immunoreactivity in the dorsal horn of the C5 spinal cord at day 7. Injury (I) significantly increased GFAP expression over sham (S) (**p < 0.035) and naïve (N) animals (*p < 0.0001). Data shown are average ± standard error of the mean (scale bar = 100 μm and applies to all panels; GFAP, glial fibrillary acidic protein).
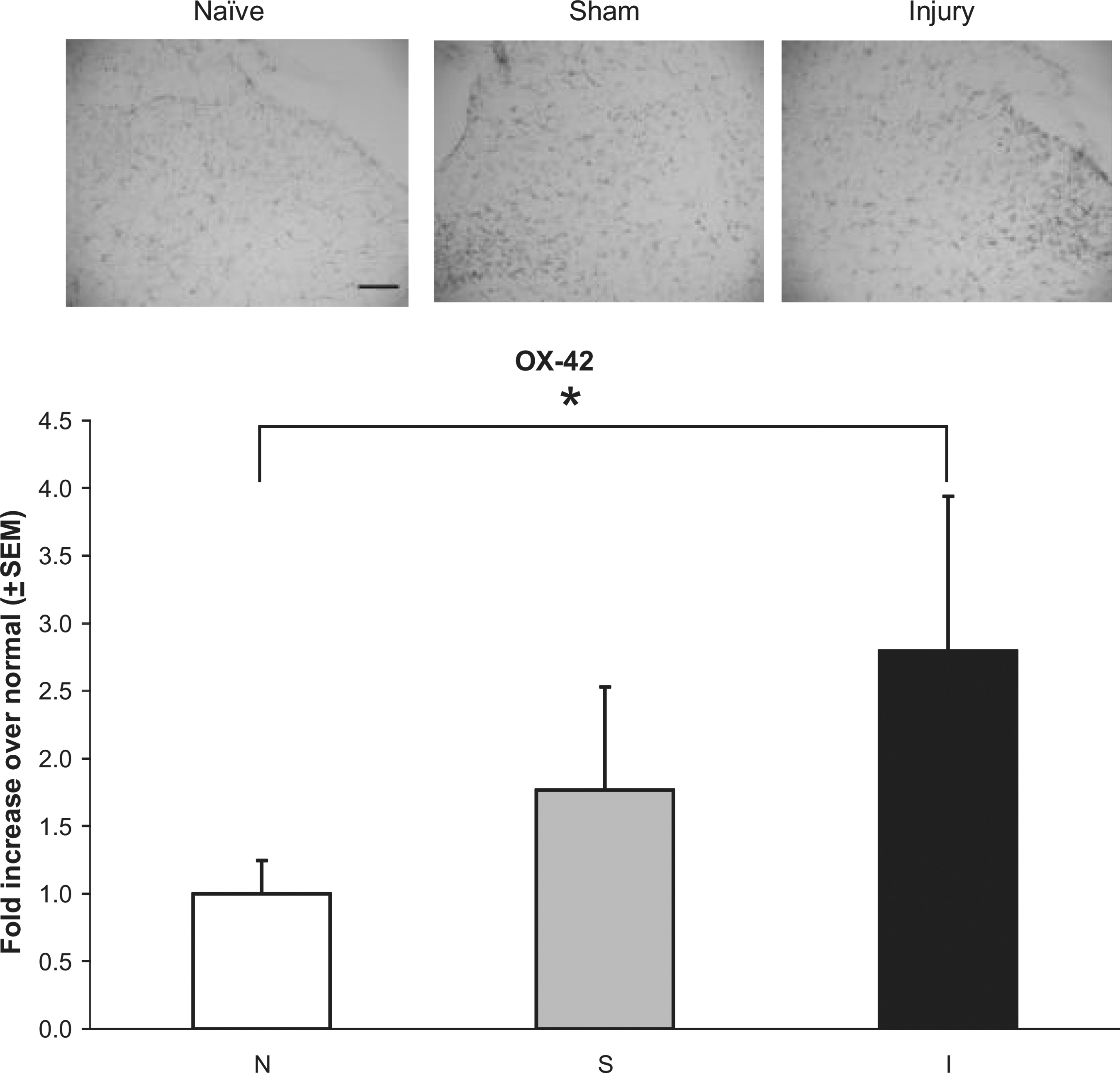
Representative images and quantification of OX-42 immunoreactivity in the dorsal horn of the C5 spinal cord at day 7. Injury (I) significantly increased OX-42 immunoreactivity over naïve (N) animals (*p < 0.006). Data shown are average ± standard error of the mean (scale bar = 100 μm and applies to all panels).
Discussion
Although neck pain is more common in adolescents than in adults, understanding the unique neurophysiological mechanism of pain from cervical facet joint–mediated injuries and how they may contribute to an increased risk of neck pain in adolescents is lacking. To our knowledge, this is the first study to model a cervical facet joint injury in an adolescent rodent model. Furthermore, such a sub-failure mechanical joint injury in the spine not only induced sustained hypersensitivity, but also increased expression of several nociceptive mediators (PKCɛ and mGluR5) in primary afferent neurons, and increased glial activation in the spinal cord (Figs. 2 –5). As such, this model system may be particularly useful for comparing the underlying pain pathologies in the adolescent and adult populations for mechanical injuries to the cervical facet joint. Such a platform would provide a scientific foundation for improved medical treatment or the prevention of neck pain in the adolescent population.
Age-related pain and biomechanics
The mechanisms underlying an adolescent's susceptibility to neck injury and pain are largely understudied. To our knowledge, no models exist that can define the cellular and molecular differences that underlie painful neck injury across age groups in any animal model system. There is limited clinical evidence for differences in these populations as they relate to recurrent neck pain and headaches (Weber Hellstenius, 2009); neck pain in adults often is initiated by a traumatic or repetitive injury. The resulting neck pain is often associated with upper cervical joint dysfunction, reduced cervical rotation, and focal areas of tension in the cervical paraspinal muscles (Hall and Robinson, 2004; Jull et al., 2007; Sjaastad et al., 2003). In contrast to adults, pre-adolescents with head and/or neck pain often do not have a diagnosable injury, and do not exhibit focal areas of tension in the cervical paraspinal muscles. Instead, adolescents often present with a combination of focal tension of the sub-occipital muscles, forward head posture, and right lower cervical joint dysfunction, without evidence of any overt injury. One potential source of these subacute painful injuries has been postulated to result from backpack use (Sheir-Neiss et al., 2003), and younger students actually have greater changes in craniovertebral angles, spinal curvature, and posture with backpack loads (Grimmer et al., 1999; Korovessis et al., 2004). Furthermore, backpack-induced changes in the craniovertebral angle correlate with disability as a result of neck pain in adolescents (Cheung et al., 2009).
Previous studies have quantified the mechanical tissue tolerance limits in terms of vertebral distraction (0.7 ± 0.1 mm and 0.72 ± 0.01 mm, respectively, in Lee et al., 2008 and Lee and Winkelstein, 2009), and associated capsular ligament strains (23.1 ± 9.3%, 27.7 ± 11.9%, 33.6 ± 3.1%, 21.5 ± 12.3%, and 24 ± 10%, respectively, in Lee et al., 2004a, 2004b, 2006, Dong et al., 2008, and Dong and Winkelstein, 2010), that are required to elicit hypersensitivity in an adult rat. But these threshold values were previously undetermined in the adolescent, in whom anatomical differences exist and mechanistic relationships may be different. In the current study, a distraction of 0.4 mm produced maximum principal strains of 32.8 ± 12.9% in the capsule, and induced sustained hyperalgesia in the forepaw (Fig. 1). In the context of the biomechanical data from the adult model, it appears that the anatomic scaling of the joint distraction applied in this study was successful in generating similar capsule strains as in the adult. Of note, despite a consistently applied vertebral distraction magnitude for the rats in this study, the capsule strains were quite variable (Table 1). Although the degree of variability is consistent with studies using adult rats and other studies of tissue biomechanics, it may contribute to the variability seen in the behavioral and nociceptive outcomes among rats. Despite a smaller distraction magnitude being imposed across the joint, the capsule strains in this study were similar to those seen in this same pain model in the adult rat (Lee and Winkelstein, 2009; Lee et al., 2004b, 2006). These strains are sufficient to activate both group III and IV afferents, by capsular strains above 10.7 ± 3.3% and 10.0 ± 4.6%, respectively, in a similar goat model of joint distraction (Lu et al., 2005a, 2005b). Furthermore, the maximum forces (1.10 ± 0.42 N) measured across the joint in the adolescent rat were lower than those seen in the adult rat (4.00 ± 1.81 N; Lee and Winkelstein, 2009) for comparable ligament strains. Taken together, these findings suggest that adolescents may have a decreased mechanical tolerance to pain compared to adults, given the lower load-thresholds needed to induce similar strains and behavioral modifications (Table 1 and Fig. 1; Lee and Winkelstein, 2009). Although it appears that similar strain thresholds may exist across age ranges for this model, in this study we did not directly measure the strain threshold for inducing pain and/or the cellular responses observed here. Additional work is needed to ascertain if the strain threshold for painful facet joint injury varies between the adolescent and the adult. Further, it is not known how these relationships apply for joint injuries in even younger age groups. Additional studies are needed with these and other age groups to more clearly define the relationships between mechanical loading, pain, and age.
Nociceptive mechanisms
In the current study, injury increased the number of small-diameter DRG neurons that express PKCɛ, as well as the density of PKCɛ immunoreactivity within both small- and medium-diameter DRG neurons (Fig. 2). Multiple protein kinase C family members have been implicated in intracellular signaling at multiple levels of the neuro-axis, and across numerous neurotransmitter and neuromodulatory peptide systems (Velázquez et al., 2007). PKCɛ is found in primary afferent neurons and has been implicated in mediating acute nociceptive pain (Amadesi et al., 2006; Ferreira et al., 2005; Khasar et al., 1999; Souza et al., 2002), inflammatory pain (Khasar et al., 1999), and neuropathic pain (Ahlgren and Levine, 1994; Dina et al., 2000) in adult animals. We have previously reported developmental regulation in the expression of PKCɛ in primary afferent DRG neurons, such that at 1 week after birth only 41% of mainly small- and medium-diameter DRG neurons are positive for PKCɛ, which rapidly increases to 60% of small-, medium-, and large-diameter DRG neurons at 3 weeks of age (Sweitzer et al., 2003, 2004a, 2004b). By adulthood, 90% of DRG neurons express PKCɛ (Khasar et al., 1999). Interestingly, inhibition of PKCɛ translocation is anti-nociceptive in models of formalin-induced nociception (Sweitzer et al., 2004a), morphine withdrawal-associated hyperalgesia (Sweitzer et al., 2003, 2004b), and ethanol-withdrawal hyperalgesia (Shumilla et al., 2005), even in very young animals. This suggests a functional role for PKCɛ in modulating pain throughout development. The current study extends these findings into adolescent animals, and suggests that PKCɛ may also modulate cervical facet pain in adolescent animals.
PKCɛ is well suited within the intracellular signaling cascade to integrate signals across numerous nociceptive receptor pathways into a final effector that increases excitatory signaling and decreases inhibitory signaling, resulting in persistent pain. At the level of the primary afferent neuron, PKCɛ has been shown to be activated downstream of bradykinin receptors, endothelin receptors, prostaglandin receptors, the proinflammatory cytokine tumor necrosis factor, galanin, the purinergic receptor, and insulin (Velázquez et al., 2007). Upon activation, PKCɛ translocates from the cytoplasm to the membrane, where it phosphorylates and increases cation flow through a host of ion channels, including TRPV1, acid-sensing ion channels, and NaV1.9, which directly modulate neuronal hyperexcitability (Velázquez et al., 2007). In fact, inhibition of PKCɛ translocation can attenuate capsaicin-induced release of glutamate and calcitonin gene-related peptide from isolated spinal cords (Sweitzer et al., 2004a).
Similarly, PKCɛ can regulate G-protein coupled receptors such as mGluR5 via increased release of calcium (Conn and Pin, 1997; Schoepp et al., 1999). In the current study, the number of small-diameter DRG neurons expressing mGluR5 increased concomitantly with an increase in the number of small-diameter DRG neurons expressing PKCɛ, suggesting a potential role of PKCɛ-dependent mGluR5 signaling in hyperalgesia following painful cervical facet joint injury in adolescent animals. In juvenile rats, the induction of long-term potentiation in trigeminal primary afferent neurons requires mGluR5-mediated activation of PKC (Liang et al., 2005). In trigeminal ganglia, mechanical hypersensitivity is mediated by peripheral mGluR5 in a PKCɛ-dependent manner (Lee and Ro, 2007). Activation of PKC by mGluR5 agonists induces a rapid exocytosis of TRPV1 to the cell membrane in DRG neurons (Morenilla-Palao et al., 2004). Previous studies have shown that mGluR5 is upregulated in small-diameter A-fibers in the uninjured DRG following L5 spinal nerve ligation. Furthermore, following nerve injury, mGluR5 on small-diameter A-fibers was co-localized with newly expressed TRPV1, and was hypothesized to modulate thermal hyperalgesia (Hudson et al., 2002). TRPV1 and mGluR5 have also been shown to be functionally coupled on presynaptic terminals of nociceptive neurons (Kim et al., 2009). Future studies should examine the co-localization of mGluR5, TRPV1, and PKCɛ in DRG neurons, as well as test the effectiveness of mGluR5 and PKC inhibitors on hyperalgesia in this adolescent facet joint injury model, to better define their effects on the onset and maintenance of pain.
Increased mGluR5 expression is known to be associated with activation of astrocytes as early as day 7 after spinal cord injury (Gwak and Hulsebosch, 2005). Astrocytic activity after injury can lead to a host of inflammatory responses, including the release of glutamate, nitric oxide, prostaglandins, and cytokines, all of which can maintain behavioral hypersensitivity (Colburn et al., 1999; DeLeo and Yezierski, 2001; Newman, 2003; Rutkowski and DeLeo 2002; Watkins et al., 2001). Increased glutamate released from astrocytes can then activate metabotropic glutamate receptors (Mills et al., 2000; Osikowicz et al., 2009). After injury, astroglial regulation becomes imbalanced, resulting in an abundance of glutamate receptor activity (Osikowicz et al., 2009; Zonta et al., 2003). In the current study, cervical facet joint distraction injury increased microglial and astrocytic activation in the spinal cord, which paralleled the elevated levels of mGluR5 and PKCɛ in the small-diameter neurons in the DRG (Figs. 2 and 3). Quantification of the neuronal expression in the DRG may represent a biased estimate of the actual number of neurons, since a stereological approach was not used in the current study. However, the total percentages of small- (46.0 ± 6.6%), medium- (50.8 ± 5.7%), and large-diameter neurons (3.0 ± 1.7%) identified in this study are consistent with those proportions identified in lumbar and cervical DRGs (Bergman and Ulfhake, 1998; Carlton and Hargett, 2007). Certainly a stereological approach would provide a more rigorous analysis of these responses. Future work should examine the temporal patterns of glial activation in relation to mGluR5 and PKCɛ activity, to determine if age-related differences exist. Additionally, co-localization studies of mGluR5, PKCɛ, and glial activation in the spinal cord would offer insight into centrally-mediated inflammatory responses.
This study is the first to investigate behavioral and cellular changes in the DRG and spinal cord following a cervical facet joint distraction injury in an adolescent rat model of spinal injury. Behavioral and tissue outcomes demonstrate similar responses, with increased hyperalgesia, and PKCɛ, mGluR5, and glial activation, indicating that these processes may work together to potentiate pain in facet-mediated pain. Furthermore, this study shows that despite a lower magnitude of applied facet joint distraction in the adolescent-scaled age model than in the adult, behavioral and cellular responses were elevated after injury. However, in the context of injury strain, adolescents may have a different mechanical tissue loading tolerance for pain, and/or a host of nociceptive cascades throughout the nervous system. This work provides insight into the mechanical, behavioral, and cellular mechanisms driving cervical facet joint pain in an adolescent rat model, and could provide better understanding to develop potential therapeutic interventions.
Footnotes
Acknowledgments
This work was funded by the Catharine D. Sharpe Foundation and the National Science Foundation (NSF) Center for Child Injury Prevention Studies (CChIPS) at the Children's Hospital of Philadelphia. The authors would like to acknowledge the Industry Advisory Board (IAB) members of CChIPS for their support, input, and advice. The views presented are those of the authors and are not necessarily the views of CChIPS, the NSF, or the IAB members.
Author Disclosure Statement
No competing financial interests exist.