
Abstract
Neuropathic pain and motor deficits are detrimental consequences of injury to the spinal cord. In experimental settings, numerous neuroprotective agents are being explored for their therapeutic benefits. Vascular endothelial growth factor (VEGF) is an interesting candidate molecule in this respect since it is not only associated with angiogenesis, but also with neuroprotection and neurite growth. Other investigators have reported improved motor outcomes following intraparenchymal VEGF treatment. Here we demonstrate the therapeutic effects of daily intrathecal treatment of the contused thoracic rat spinal cord with the 165-isoform of VEGF during the first week after injury. We show that VEGF treatment resulted in a statistically significant attenuation of mechanical, but not thermal, hypersensitivity of the hindpaws, while motor deficits remained unaffected. Tissue sparing was also unchanged by VEGF treatment. Microglial responses at the lumbar spinal cord, which have been linked with spinal cord injury–induced hypersensitivity, were found to be unaffected by VEGF treatment. We conclude that repetitive intrathecal VEGF delivery has limited therapeutic effects on spinal cord injury outcome.
Introduction
Vascular endothelial growth factor (VEGF) is a key factor in the process of angiogenesis, but has also been associated with neuroprotection and neurite outgrowth (Rosenstein and Krum 2004; Sondell et al., 1999). Increased levels of VEGF are detected around spinal injury sites (Bartholdi et al., 1997; Herrera et al., 2009; Skold et al., 2000; Vaquero et al., 1999). Treatment of the spinal cord parenchyma with anti-VEGF antibody decreased the number of surviving neurons around the epicenter of a contusion injury, indicating a neuroprotective role of endogenous VEGF (Herrera et al., 2009). Delivery of the 165-isoform of VEGF (VEGF165) via a single intraparenchymal bolus injection at the time of spinal cord contusion was found to promote tissue preservation (Widenfalk et al., 2003). In a more recent investigation, elevated levels of VEGF were maintained for at least 6 weeks following clip compression of the rat spinal cord by intraparenchymal injection of a viral vector containing a transcription factor that induces VEGF expression (Liu et al., 2010). This treatment resulted in enhanced tissue sparing within a range of a millimeter distance from the lesion epicenter, and a mild improvement of motor function (Liu et al., 2010), as measured by the Basso-Beattie-Bresnahan locomotor scale.
In the present study, we have investigated the effect of repetitive VEGF treatment on SCI-induced hypersensitivity. In addition, motor outcome was measured using an objective and sensitive gait analysis system. We show that daily intrathecal injections of VEGF165 for 1 week following SCI resulted in a slight attenuation of mechanical but not thermal hypersensitivity, while motor deficits were unchanged. The extent of morphological preservation of spinal cord tissue was not affected by the treatment. Microglial responses associated with SCI-induced hypersensitivity were observed to a similar degree in both VEGF165-treated and vehicle-treated animals.
Methods
All experimental procedures were performed according to the recommendations of the European Commission (European Communities Council Directive of 24 November 1986; 86/609/EEC), and protocols were approved by the committee on animal research of Maastricht University (DEC 2009–030). Twenty-one female Sprague-Dawley rats (Charles River, Wilmington, MA) were used in this study, weighing about 200 g at the start of the experiment. Throughout the study the animals were individually housed in a temperature- and humidity-conditioned room (19–24°C and 55 ± 5%), with a normal 12-h:12-h light-dark cycle and background music to reduce stress. In the 2-week CatWalk-training period (see below) preceding surgery, and on each post-surgery day before recording of CatWalk runs, the animals were kept on a food-restriction protocol (12–15 g chow per day). Otherwise, the animals received standard rat chow ad libitum. The animals had continuous free access to water for the entire period of the study.
Surgery and experimental groups
The animals received a subcutaneous injection of buprenorphine (0.1 mg/kg body weight) at about 30–60 min before surgery. An Inventor 400® injection system vaporizer (Zevenaar, The Netherlands) with an open mask system was used for anesthesia. For induction and maintenance of anesthesia 5% and 1–2% isoflurane (in air) was used, respectively. First, a polyethylene catheter was implanted via an incision in the atlanto-occipital membrane. The tip of the catheter was gently maneuvered to the eighth thoracic (T8) spinal cord level. Then, the T9–T10 spinal cord was exposed by a laminectomy of the T8 and T9 vertebrae. A moderate spinal cord contusion was inflicted using the MASCIS impactor with a 10-g rod dropped from a distance of 12.5 mm onto the exposed dorsal surface of the spinal cord. Then the animals were randomly assigned to two treatment groups: animals either received an intrathecal bolus injection of 0.5 μg VEGF165 in 13 μL (n = 9), or 13 μL vehicle solution (n = 9), followed by a 17-μL saline flush over a period of about 20 sec. Human recombinant VEGF165 was produced by forced expression in HEK293 cells (Mineur et al., 2007), and was purified from the conditioned medium by immuno-affinity using anti-VEGF antibodies (Avastin; Genentech, South San Francisco, CA), coupled to Affi-Gel Hydrazide gel (Bio-Rad, Hercules, CA). VEGF concentration was measured using a DuoSet ELISA (R&D Systems, Minneapolis, MN). Immediately after surgery, the bladders were manually voided and the animals received 5 mL subcutaneous saline solution for rehydration. Subcutaneous antibiotic treatment with enrofloxacin (5 mg/kg body weight) was given once daily until normal bladder emptying returned (typically within the first week after SCI) to prevent urinary tract infection. At 1–7 days following surgery, the animals received the same daily treatment (VEGF165 or vehicle) as was given directly after surgery. As such, VEGF-treated animals received a total of 4 μg VEGF165 over the first week following surgery. This amount of VEGF has previously been shown to limit spinal tissue degeneration in a rat model of severe spinal cord contusion injury (Widenfalk et al., 2003). After the last intrathecal bolus injection, the catheter was carefully removed while the animal was anesthetized as described above. A total of three naïve animals served as controls in this experiment.
Behavioral testing
Motor function was assessed using CatWalk gait analysis (Hamers et al., 2001), and the Basso-Beattie-Bresnahan (BBB) locomotor rating scale (Basso et al., 1995). For gait analysis, the animals were trained on the CatWalk runway during the first 2 weeks preceding surgery according to previously documented protocols (Deumens et al., 2007a). During this training period the rats were positioned at one end of the runway and sweet food pellet rewards (Noyes Precision pellets PJPPP-0045; Sandown Chemical Ltd., Hempaton, U.K.) were located at the other end of the runway. The rats were allowed to explore the runway three times per day. Motivation of the animal was reinforced by use of a daily 12–15 g chow diet. After 2 weeks of training, the animals consistently displayed uninterrupted runs and such runs were recorded before surgery. Then CatWalk runs were recorded weekly starting at the fourth week after surgery. Only runs with runway-crossing times of 1–2 sec were used for analysis. This criterion was used to warrant a stable velocity of gait, which is of crucial importance because velocity of gait can largely influence gait parameters (Koopmans et al., 2007). For each time point at least three runs per animal were analyzed using CatWalk XT v8.1 software (Noldus Information Technology, Wageningen). Here we focused on four CatWalk parameters that have previously been reported to be affected in rat models of SCI. First, stride length (i.e., the distance between two subsequent positioning of the same hindpaw; the values of both hindpaws were averaged); second, base-of-support (BOS) (i.e., the average distance between the center of the right and left hindpaws during the gait); third, print area (i.e., the total area that makes up the print of the hindpaws during gait, with the values of both hindpaws averaged); and fourth, mean intensity (i.e., the mean grey value of the pixels corresponding to the hindpaw print at maximum contact during a step cycle, with values of both hindpaws averaged). BBB scores were obtained before surgery and at 1, 3, 5, and 7 days after surgery, and thereafter once a week until the end of the experiment (8 weeks after surgery). Because reliable assessment of coordination using the BBB scale is limited, CatWalk-based coordination data were integrated into BBB scores at 4, 6, and 8 weeks following SCI, as previously reported (Koopmans et al., 2005).
A dynamic plantar aesthesiometer (UgoBasile, Collegeville, PA) was used to assess sensitivity to mechanical stimuli. The animals were habituated to the testing environment for about 30 min until exploration and major grooming ceased. This testing environment consisted of transparent plastic boxes positioned on top of a plastic mesh floor. A metal probe was applied to the mid-plantar surface of the rat hindpaw, and this probe exerted a gradually increasing force (50 g in 10 sec) until a withdrawal reflex was elicited. The threshold evoking a withdrawal reflex was determined for each hindpaw (mean of three measurements, referred to as paw withdrawal threshold [PWT]). PWTs were determined before surgery and then weekly starting at the second week after surgery. To assess mechanical sensitivity, the area under the curve (AUC) of the PWT/time plot was calculated for each animal over the 8 weeks following SCI. Thermal sensitivity was determined using a thermal plantar analgesia instrument (UgoBasile) as previously described (Hargreaves et al., 1988). Following an habituation period of about 30 min, an infrared laser beam was positioned at the mid-plantar surface of the rat hindpaw. Before surgery, the intensity of the laser was fixed at a degree that elicited a withdrawal reflex of the hindpaws with a latency of 10–15 sec; this intensity was maintained throughout the experiment. Three withdrawal latencies were determined per hindpaw and averaged for each animal to render the paw withdrawal latency (PWL). PWLs were determined before surgery and then weekly starting at the second week after surgery. In addition, AUCs were determined as described above.
Histology
After the final behavioral tests at 8 weeks after surgery, the animals were terminally anesthetized using an intraperitoneal bolus injection of pentobarbital (180 mg/kg body weight). Subsequently they were transcardially perfused with ice-cold saline solution followed by ice-cold fixative (4% paraformaldehyde in 0.1 M phosphate buffer [PB] with 15% picric acid, pH 7.4). The spinal cord was then isolated and immersion fixed at 4°C overnight. Cryoprotection was commenced by an overnight incubation in 10% sucrose (in PB), followed by an incubation in 25% sucrose (in PB) for at least 72 h (all incubations were at 4°C). Then each spinal cord was dissected into two tissue samples: a sample with a length of about 1.5 cm centered at the lesion epicenter (lesion samples), and a sample with a length of about 1.5 cm containing the lumbar region L3–L6 (lumbar samples). Each sample was frozen using dry ice powder and stored at −80°C until further use for histological processing. Cryosectioning was performed on a Leica CM3050S cryostat (Leica Microsystems, Bannockburn, IL). The samples were mounted in Tissue-Tek Optimal Cutting Temperature solution (O.C.T.; Sakura FineTek Europe, Zoeterwoude, The Netherlands). Transverse sections 30 μm thick were obtained over the range of the spinal lesion site and from the L4 and L5 spinal cord; all cryosectioning was performed at a temperature of −30°C. All samples were serially cut by taking every 12th section on the same (gelatin-coated) glass slide.
Every 12th section of the lesion site samples was stained with Luxol fast blue (LFB) to identify the injury epicenter. A Confocal Spinning Disk (SI-SD) system (MBF Bioscience, Williston, VT), consisting of a BX51 microscope (Olympus, San Jose, CA), a customized spinning disk unit (DSU; Olympus), and controlling software (StereoInvestigator; MBF Bioscience) was used for the analysis. For each lesion sample, the total area of spinal tissue in an uninjured transverse section was measured. This value was then taken as a reference value for that particular lesion sample. Subsequently, the percentage of tissue sparing was determined as the area of preserved spinal tissue in a transverse section compared to the reference value of that lesion sample. The section with the lowest percentage of spared tissue was assigned as the lesion epicenter. Transverse sections with an interval of 360 μm rostral and caudal to this lesion epicenter were analyzed for percentage tissue sparing up to a distance of 2520 μm away from the lesion epicenter.
Every 12th section of the lesion sample was stained for glial fibrillary acidic protein (GFAP) to study the scar tissue surrounding the central cavity at the spinal lesion site. Samples of each experimental group were investigated for astroglial scar tissue in a blinded manner by two independent observers.
A stain for ionized calcium-binding adapter molecule-1 (Iba-1) was used to study microglia in the L4–L5 spinal cord. Spinal sections were first incubated with blocking solution (Tris-buffered saline with 0.3% Triton-X100 [TBS-T] containing 5% normal donkey serum) for 1 h at 4°C; then overnight incubation was performed with primary antibody at 4°C (rabbit α-Iba-1; Wako, Osaka, Japan) 1:1000 in TBS-T containing 1% normal donkey serum. Staining with rabbit α-GFAP-antibody (1:1000 in TBS-T; Dako, Carpenteria, CA) was used to study astroglia in the L4–L5 spinal cord. A donkey α-rabbit-Alexa-488 antibody (1:100 in TBS-T; Invitrogen, Carlsbad, CA) was used as a secondary antibody. The stained sections were embedded in 80% glycerol/TBS and cover-slipped. Photomicrographs were taken with identical exposure times using a microscope for detection of fluorescence, and an Olympus AX-70 grayscale F-view cooled CCD camera (Paes, Zoeterwoude, The Netherlands). For quantitative analysis, cell profiler software (CellP©; Olympus) was used, and the dorsal horns were delineated as regions of interest (ROI) by drawing a line perpendicular to the central canal and then following the shape of the dorsal horn. After background signal subtraction, the average grey value (intensity) of the staining was measured in the ROI as described elsewhere (Deumens et al., 2009). A total of 10 right and 10 left dorsal horns at L4–L5 were analyzed per animal and averaged.
Statistical analysis
All data were analyzed using SPSS 15.0 statistical software. Repeated-measures analysis of variance (ANOVA) followed by Bonferroni post-hoc corrections was used to test for behavioral differences over time. Subsequently, VEGF treatment effects were tested using the independent Student's t-test. Effects of treatment on signal intensities of immunohistochemical stainings were tested using a one-way ANOVA with Bonferroni post-hoc corrections.
Results
Two out of the 18 vehicle-treated SCI animals were excluded from the study. One animal developed ascites during the first week after surgery, and the other animal developed a urinary tract infection despite enrofloxacin treatment; it lost more than 20% of its pre-surgery body weight and was then euthanized for ethical reasons.
VEGF treatment did not affect chronic motor function
SCI induced a large decrease in BBB score, which gradually recovered over the first week (Fig. 1A and B). CatWalk gait analysis was begun when weight-supported stepping of hindpaws returned (typically during the second week after injury). Between 4 and 8 weeks after injury, CatWalk-based coordination (regularity index) measures were obtained and integrated into the BBB score. This resulted in scores around 18 (Fig. 1B), reflecting nearly perfect coordination between fore- and hindpaws. In comparison with the control group, VEGF-treated animals did not show statistically significantly different scores on the BBB test at any time point of the investigation. CatWalk gait analysis showed only limited deficits in motor skills of the rats while crossing the glass runway. No statistically significant decrease in stride length (Fig. 1C), or the coordination measure “regularity index” (data not shown), could be observed from 4 weeks post-surgery onwards. The BOS (i.e., the distance between the centers of both hindpaws) was found to be significantly increased in vehicle-treated animals after injury compared to baseline (F 3,18 = 9.8; p < 0.01), which was not the case for VEGF-treated animals (Fig. 1D). A reduction in print area of the hindpaws after injury was observed for both vehicle-treated (F 3,18 = 10.7; p < 0.01) and VEGF-treated animals (F 3,24 = 8.5; p < 0.01; Fig. 1E). Moreover, a reduction in mean intensity of the hindpaws after injury was observed for both vehicle treated (F 3,18 = 8.9; p < 0.01) and VEGF-treated animals (F 3,24 = 6.4; p < 0.01; Fig. 1F). Although reductions of print area and mean intensity were significantly lower in vehicle-treated animals than in VEGF-treated animals at 6 weeks after injury (p < 0.05), there was no group difference at 8 weeks after injury.
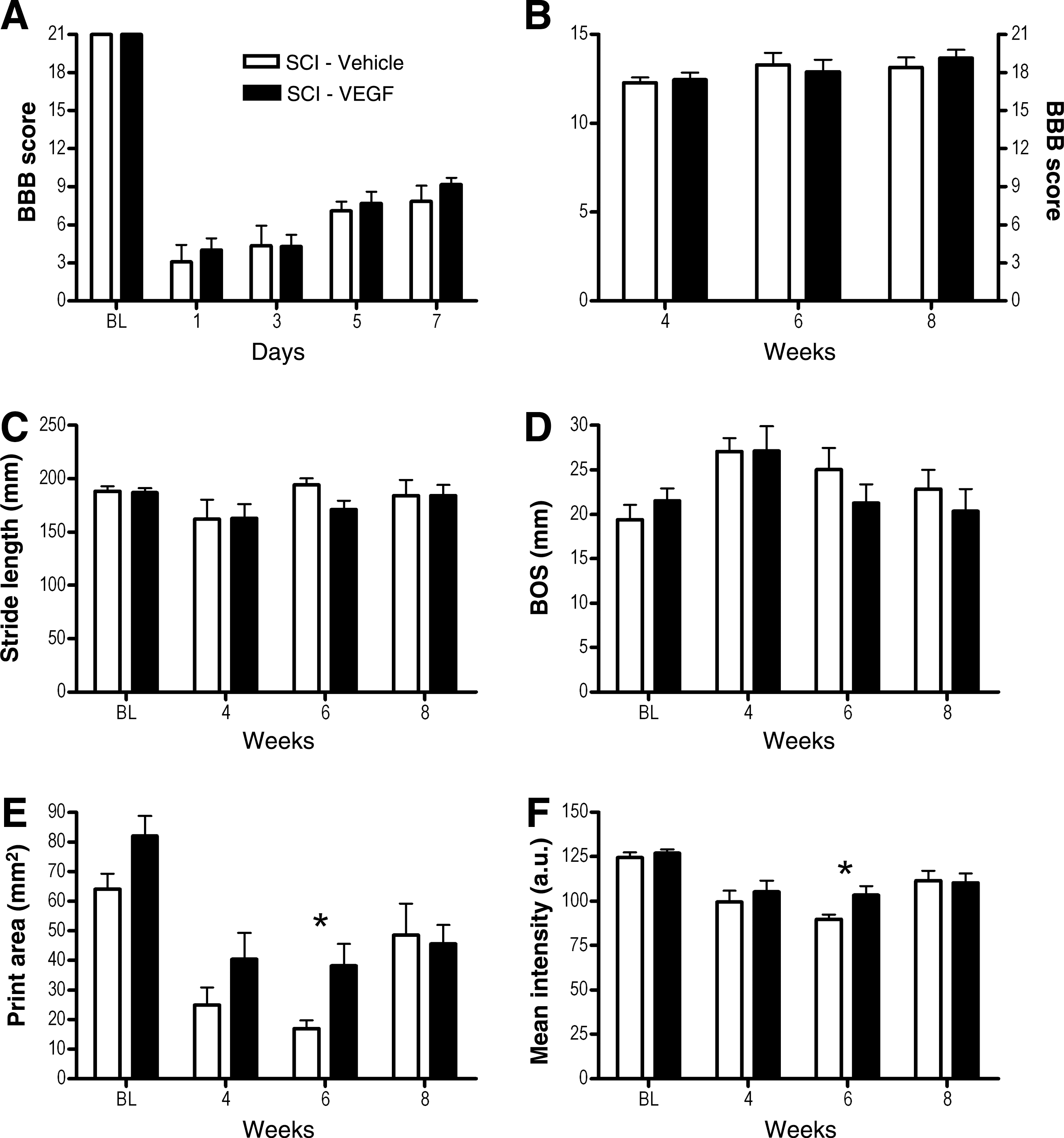
Unaffected chronic motor outcome following vascular endothelial growth factor (VEGF) treatment of the contused spinal cord. Basso-Beattie-Bresnahan (BBB) testing showed strong decreases in scores within the first weeks
VEGF treatment slightly attenuated mechanical but not thermal hypersensitivity following SCI
Spinal cord contusion injury resulted in a significant drop of PWT to mechanical stimulation of the hindpaws in both vehicle-treated (F 7,42 = 7.2; p < 0.01) and VEGF-treated animals (F 7,56 = 11.6; p < 0.01; Fig. 2A). At baseline, PWTs were elicited at about 33 and 36 g in vehicle-treated and VEGF-treated animals, while at 8 weeks after SCI PWTs were reduced to 22 and 26 g, respectively. VEGF had a minor therapeutic effect on mechanical hypersensitivity. During 7 weeks following contusion injury, the PWT dropped continuously in control animals, whereas the VEGF-treated animals reached a plateau at a lower level of hypersensitivity (Fig. 2A). Using a repeated-measures ANOVA for time, an overall difference between VEGF-treated and vehicle-treated animals was detected (F 1,14 = 5.8; p < 0.05). Student's t-tests showed an approximate 15% reduction in the PWT of VEGF-treated animals compared to vehicle-treated animals at 7 and 8 weeks post-injury (p < 0.05). This quantitatively resulted in an 8% reduction of the AUC for VEGF-treated versus vehicle-treated animals (Fig. 2B, p < 0.05). The attenuation of mechanical hypersensitivity by VEGF was significant at 7 and 8 weeks after SCI (Fig. 2A). The PWL to thermal stimulation of the hindpaws was also strongly reduced by SCI (F 7,42 = 4.5; p < 0.01 for vehicle-treated animals, and F 7,56 = 8.0; p < 0.01 for VEGF-treated animals; Fig. 2C). While PWLs of about 13 sec were observed at baseline, group average values of 8 sec were observed at 8 weeks after SCI. There was no significant difference between the two groups of animals (Fig. 2C and D).
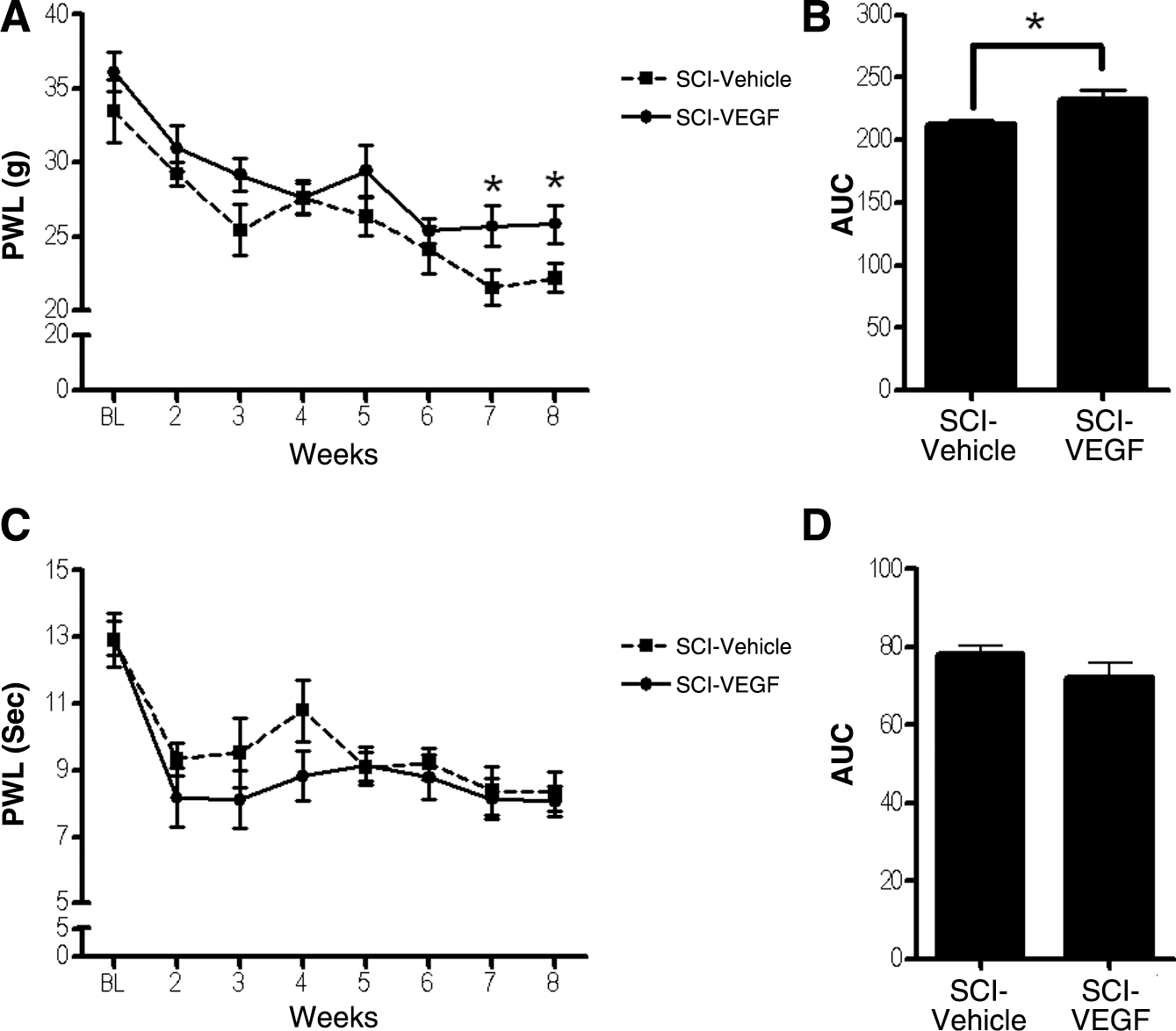
Mechanical but not thermal hypersensitivity were slightly attenuated by vascular endothelial growth factor (VEGF) treatment.
Astroglial scarring occurred around the spinal cord contusion site
GFAP staining of the injured spinal cord showed clear and dense astroglial scar tissue around the cystic cavities at 8 weeks following injury (Fig. 3A and B). The GFAP staining was too dense to clearly discriminate individual astroglial cells, although occasional hypertrophied cell profiles could be detected (arrows in Fig. 3C and D). GFAP staining of vehicle-treated and VEGF-treated animals did not show any apparent differences.
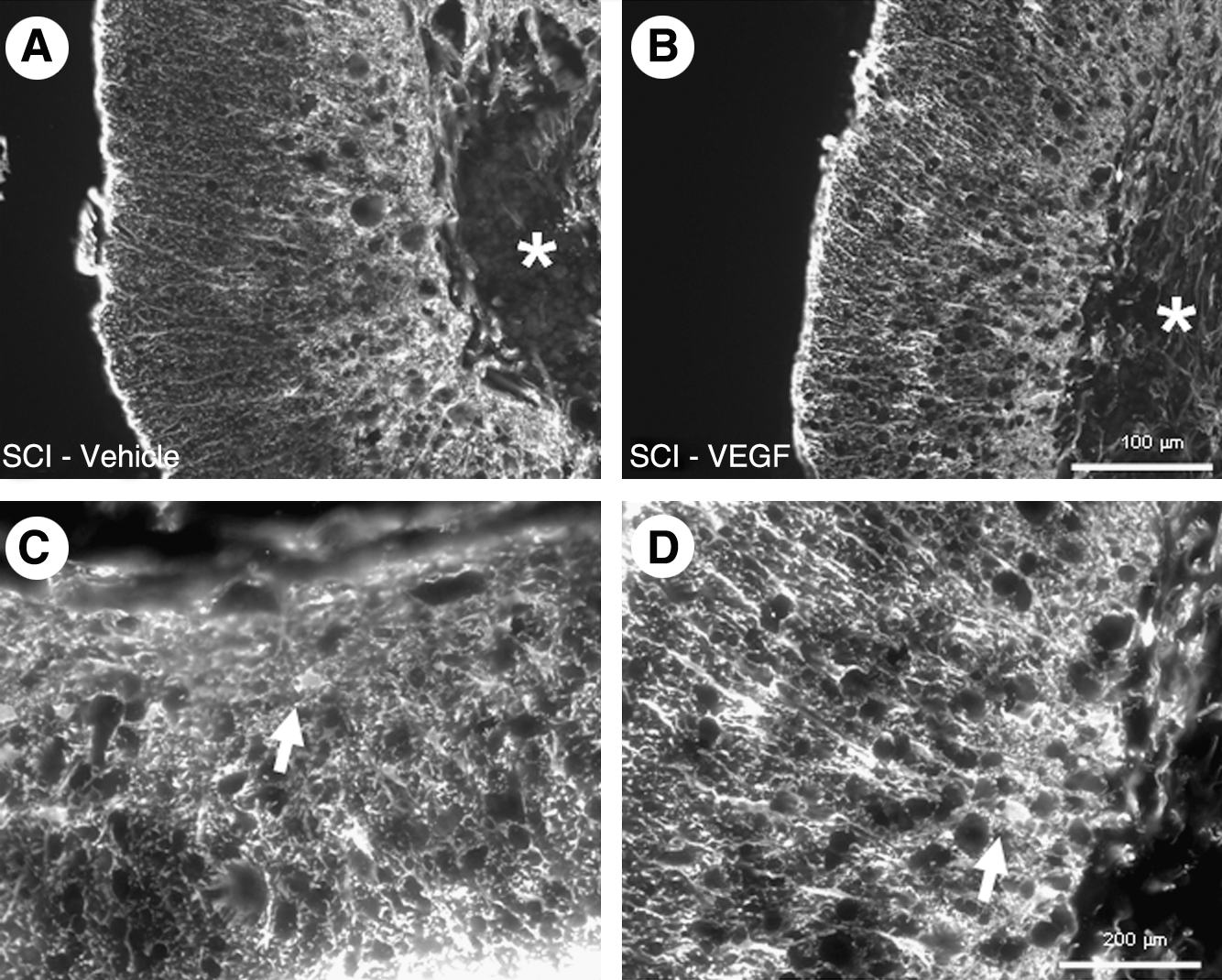
Astroglial scarring at the lesion site.
Spinal tissue sparing was unaffected by VEGF treatment
In order to assess the neuroprotective effects of VEGF treatment, morphometric analysis was performed on serial transverse spinal cord sections stained with Luxol fast blue. Only residual tissue without inflammatory cell infiltrates such as phagocytic macrophages was regarded as spared spinal tissue, and sections containing the lowest amount of spared spinal tissue were defined as the lesion epicenter. At the epicenter only a rim of white matter remained intact and devoid of inflammatory cell infiltrates at 8 weeks after SCI (Fig. 4A–D). The amount of spared tissue was no different between VEGF-treated and vehicle-treated animals (Fig. 4E).
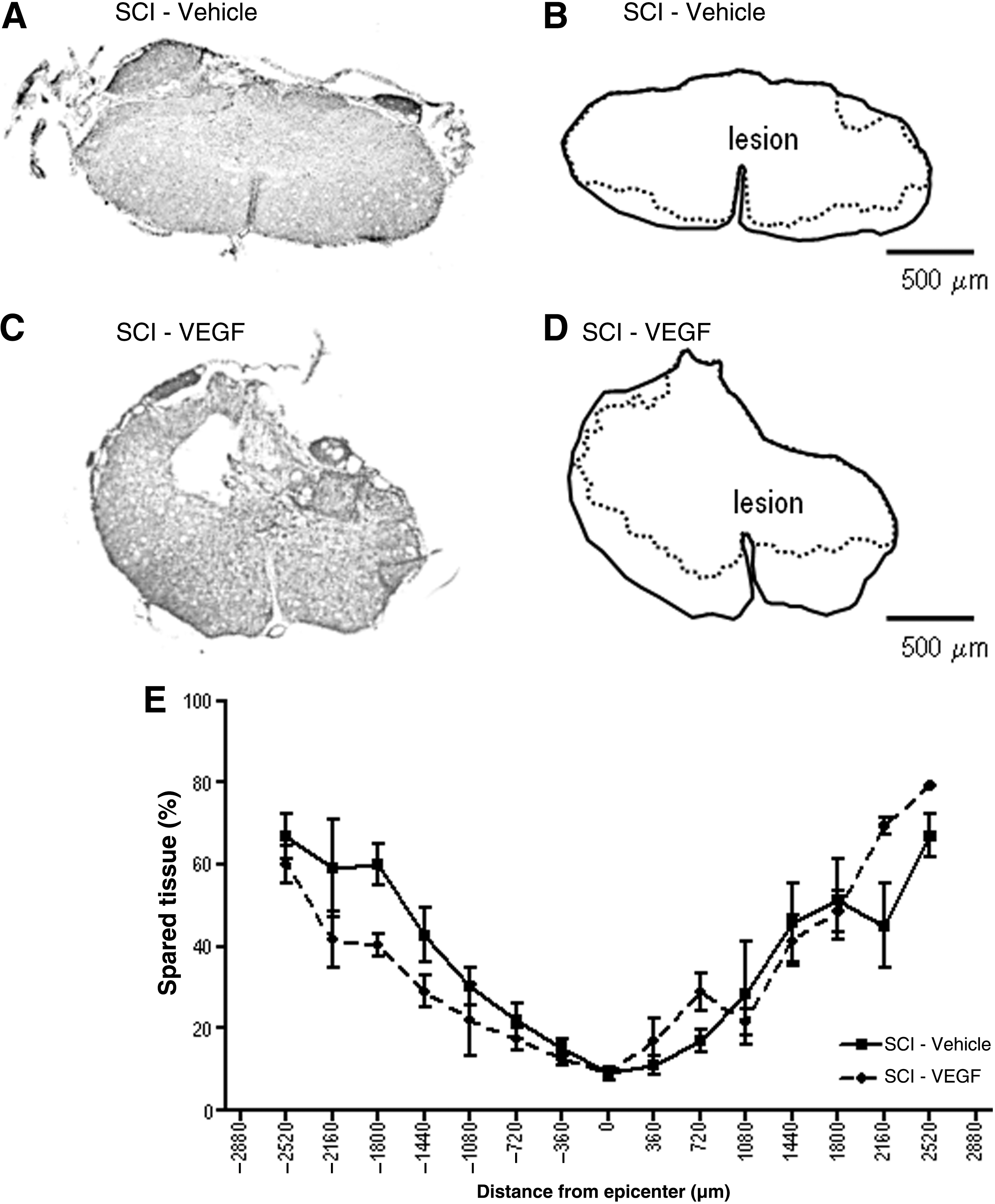
Tissue sparing was not affected by vascular endothelial growth factor (VEGF) treatment.
The microglial response to SCI remained unaffected by VEGF
Moderate spinal cord contusion of the thoracic spinal cord resulted in a strong upregulation of microglial Iba-1 staining (Fig. 5A and B). VEGF treatment did not show any overall change in staining compared to vehicle treatment (Fig. 5C). At higher magnification, intact naïve animals showed microglial cells with highly ramified morphologies (Fig. 5D), while SCI induced hypertrophy of the cell bodies and an increase in the density of cells in both vehicle-treated and VEGF-treated animals (Fig. 5E). Quantitative evaluation showed a statistically significant upregulation in both vehicle-treated and VEGF-treated animals compared to naïve control animals (Fig. 5G). GFAP staining was performed to assess astroglial responses at 8 weeks after SCI, but it was unchanged in SCI animals (both vehicle-treated and VEGF-treated) compared to naïve control animals (Fig. 5H–N).

Lumbar glial responses to spinal cord injury (SCI) in vehicle-treated and VEGF-treated animals. Iba-1
Discussion
In the present study the effect of repetitive intrathecal delivery of VEGF165 during the first week after thoracic spinal cord contusion injury was assessed. We found that VEGF treatment had only limited therapeutic effects, which involved a slight but significantly attenuated mechanical hypersensitivity of the hindpaws. The extent of tissue loss and astroglial scar tissue were unaffected by the treatment, and no lasting improvement in functional recovery could be obtained. Microglial responses in the lumbar spinal cord were present in VEGF165-treated and vehicle-treated animals, while astroglial responses (i.e., GFAP upregulation) were absent at 2 months following injury.
Neuropathic pain is one of the most debilitating consequences of injury to the spinal cord (Felix et al., 2007; Nepomuceno et al., 1979; Siddall et al., 2003). In this study, VEGF treatment resulted in a slight but statistically significant attenuation of mechanical, but not thermal, hypersensitivity of the hindpaws. Microglial responses have previously been shown to play a vital role in mechanical and thermal hypersensitivity at about 1 month after SCI (Hains and Waxman, 2006). Such microglial responses, classically characterized by morphological transformations such as hypertrophy and retraction of microglial processes, but also by proliferation, do not only occur at the level of the spinal injury, but can extend over several spinal segments (Detloff et al., 2008), and can even be detected at supraspinal locations (Zhao et al., 2007b). In the present study, upregulation of the microglial protein Iba-1 was observed in the dorsal horn of the lumbar spinal cord in both vehicle-treated and VEGF-treated animals. On the basis of these data, it is rather unlikely that microglial changes are involved in the minor reduction in mechanical hypersensitivity seen following VEGF treatment. It has to be stated that morphological changes are an oversimplified way to look at microglia, as such changes do not provide any information about alterations in cellular function. Indeed, microglia are thought to have many different functional states, many if not all of which are characterized by morphological changes (Ransohoff and Perry, 2009). With respect to functional microglial states, in particular the expression of the eicosanoid prostaglandin E2 (PGE2) has been linked to microglial-mediated hypersensitivity following SCI (Hains et al., 2001; Zhao et al., 2007a). However, since VEGF treatment did not change thermal hypersensitivity of the hindpaws in the present study, and such hypersensitivity depends on PGE2 (Zhao et al., 2007a), it is unlikely that the VEGF-induced reduction of mechanical hypersensitivity was due to modified PGE2 signaling. Surprisingly, we did not find any indications of astroglial responses in the lumbar spinal cord at 8 weeks after SCI, while astroglial responses are often suggested to be involved in symptoms of neuropathic pain (Gwak and Hulsebosch, 2009; Ji et al., 2006; Nesic et al., 2005; Tanga et al., 2006). Nevertheless, the link between such astroglial responses and neuropathic pain symptoms is typically seen within the first 4–5 weeks, while later time points are not frequently investigated. Interestingly, astroglial responses in the lumbar dorsal horn at 3 months after sciatic nerve injury have been found to be inversely related to mechanical hypersensitivity (Deumens et al., 2009).
The VEGF-induced attenuation of mechanical hypersensitivity of the hindpaws was only minor, and therefore its physiological relevance remains uncertain. A subset of investigations has nevertheless claimed therapeutic effects of VEGF treatment in animal models of SCI (Facchiano et al., 2002; Liu et al., 2010; Widenfalk et al., 2003), while others have found no effects or even negative effects, including an increase in the permeability of the blood–spinal cord barrier (Benton and Whittemore, 2003; Vaquero et al., 1999). Therapeutic effects have been related to enhanced plasticity in neuronal structures, which has also been frequently associated with mechanical hypersensitivity (Christensen and Hulsebosch, 1997; Deumens et al., 2007b, 2008; Hofstetter et al., 2005). Injured corticospinal axons that showed particularly weak spontaneous regenerative responses following SCI (Deumens et al., 2005) were found to re-grow following VEGF165 treatment (Facchiano et al., 2002). In addition, VEGF has been associated with neuroprotective effects. A single 4-μg bolus injection of VEGF165 into the damaged spinal parenchyma at the time of spinal contusion injury was found to result in enhanced spared spinal tissue (Widenfalk et al., 2003). In the present study, no such neuroprotective effects were observed. The amount of tissue loss at the lesion epicenter reached up to 90% in both vehicle-treated and VEGF-treated animals.
The lack of neuroprotective effects of VEGF treatment found in the present study may be attributable to our treatment design. It was previously shown that intraparenchymal injection of 4 μg, but not 1 or 20 μg of VEGF165 resulted in motor improvements according to the BBB scale (Widenfalk et al., 2003). In the present study, a total of 4 μg of VEGF165 was given intrathecally, equally distributed over a total of eight daily bolus injections within the first week following injury. Tissue penetration of VEGF165 may be less extensive following intrathecal administration compared to intraparenchymal administration. Although astroglia also express VEGF receptors (Krum and Rosenstein, 1998; Krum et al., 2002), astroglial scar tissue around the spinal lesion site did not appear to be affected by VEGF treatment. An alternative explanation for the lack of neuroprotective effects of VEGF seen in the present study may involve the duration of treatment. In this study treatment was restricted to the first week, because tissue sparing after VEGF165 treatment has previously been reported during the first week following SCI (Widenfalk et al., 2003). Recent findings show that experimentally-induced upregulation of VEGF at the site of spinal cord compression injury for a total of at least 6 weeks substantially reduced tissue loss around the lesion epicenter (Liu et al., 2010). Moreover, BBB scores were significantly improved following prolonged elevations in VEGF levels (Liu et al., 2010); however, in our study BBB scores were unaffected by VEGF treatment. Of note, CatWalk-based BBB scores were found to be particularly high (around 18) from the fourth week onwards, indicating the general well-being of the animals in this study. Gait analysis using the CatWalk system further showed that the motor impairments seen following moderate spinal cord contusion were relatively mild. Gait analysis can be reliably used to assess motor outcome after SCI in the rat (Deumens et al., 2006b, 2006c; Hamers et al., 2006; Koopmans et al., 2007, 2009). Previously, the base-of-support of the hindpaws was found to be increased by spinal cord contusion in female Wistar rats, and this was thought to reflect compensatory behavior for an injury-induced reduction in the stability of gait (Koopmans et al., 2006). In the present study, the base-of-support was only increased during the first 4 weeks after injury, after which pre-operative levels were reached. Stride length of the hindpaw (i.e., the distance between two subsequent placements of the same hindpaw), which was previously found to be affected following dorsal hemisection injury of the spinal cord (Deumens et al., 2006a), remained unaffected after spinal cord contusion. Chronic deficits in motor function were unaffected following VEGF treatment in the present study, a finding in agreement with recent results that showed no lasting motor improvement induced by VEGF treatment (Patel et al., 2009).
On the basis of the present study it may be concluded that VEGF has only limited therapeutic benefits for the treatment of SCI. It is clear that suboptimal conditions related to the treatment design (including dosing, timing, and drug delivery route) may partly explain the relatively poor therapeutic outcome of VEGF treatment seen in the present study, with only mild reductions in mechanical hypersensitivity and transient motor improvements. Nevertheless, VEGF has a rather narrow therapeutic range of action with respect to neuroprotection following SCI (Widenfalk et al., 2003). Thus alternative drug candidates need to be explored in the search for neuroprotective therapies to improve SCI outcomes. Anti-inflammatory therapies may be of interest in this respect, as such therapies may prevent inflammation-induced exacerbation of tissue damage, and some have been reported to attenuate SCI-induced hypersensitivity and motor deficits (Gris et al., 2004; Koopmans et al., 2009; Park et al., 2007; Plunkett et al., 2001). Nevertheless, anti-inflammatory therapies may also interfere with beneficial effects associated with the inflammatory processes seen following SCI (Faulkner et al., 2004; Moalem et al., 1999; Schwartz et al., 1999; Sofroniew, 2005). Other neuroprotective approaches that selectively interfere with apoptosis may therefore be more suitable for the treatment of SCI (Citron et al., 2008; Li et al., 2000).
In conclusion, repetitive intrathecal delivery of VEGF165 over the first week after rat spinal cord contusion injury showed only minor therapeutic effects, which included slightly attenuated mechanical but not thermal hypersensitivity and unaffected motor outcome. Microglial responses, normally linked to SCI-induced hypersensitivity, were present in the lumbar dorsal horn after spinal cord contusion following both vehicle treatment and VEGF treatment. Tissue sparing at the lesion site and astroglial scarring were unaffected by VEGF treatment. As such, VEGF has only limited beneficial effects on SCI outcomes.
Acknowledgments
The authors thank Dr. Pierre Mineur for his efforts regarding the production of VEGF165. In addition, the authors wish to thank Ms. Marijke Lemmens for her assistance in the use of the confocal SI-SD system. The authors are grateful to the Royal Dutch Academy of Arts and Sciences (KNAW) for financial support of this work via a KNAW-Hendrik Casimir Karl Ziegler Research Stipend (to R.D.), and a KNAW-van Leersum Fonds grant (to R.D.). Special thanks also go to the Dutch Society for Anesthesiology (NVA) for their financial support of this study via the Young Investigator Grant (to R.D.). The SI-SD system was funded by grant no. 911-06-003 from the Medical Section of the Dutch Scientific Organization (NWO). This work was supported by an FP6-Marie Curie Early Stage Training Site, grant number MEST-CT-2005-020013 (Euron consortium) to J.M.
Author Disclosure Statement
No competing financial interests exist.