
Abstract
Strategies for nervous system repair arise from knowledge of growth mechanisms via a growth cone. The distinctive process of axon stretch growth is a robust, long-term growth that may reveal new pathways to accelerate nerve repair. Here, a live imaging bioreactor was engineered to closely explore cellular events initiated by applied tension. The stretch growth potential between adult and embryonic dorsal root ganglion (DRG) neurons was investigated, an important difference in nerve repair. Embryonic axons were capable of unidirectional stretch growth rates of 4 mm/d and reliably reached 4 cm in length within 2 weeks. Adult axons could only reach 2 mm/d and took over 3 weeks to reach 4 cm. Utilizing time-lapse imaging, we observed growth cone motility in coordination with stretch growth. Upon initiation of stretching, growth cones retracted. However, within 10 h of continuous stretching, growth cones extended at a rate of 0.2 mm/d opposite the direction of applied tension, contributing to overall axon elongation. We analyzed fast mitochondrial transport under increasing levels of strain to determine the effect of stretch on axonal transport. Transport began to diminish at 24% strain, and was almost completely absent at 39% strain. Surprisingly, axons recovered and were capable of subsequent stretch growth. When tension was completely released (−5% strain), stretch grown axons retracted at rates up to 6.1 μm/sec and slowed as resting tension was restored. This ability to assess the process of axon stretch growth in real time will allow detailed study of how tension can be used to drive axonal growth and retraction.
Introduction
A
Conventional knowledge of axon growth typically considers only the navigation of growth cones and the formation of synaptic connections during early development and regeneration (Dickson, 2002; Yu and Bargmann, 2001). Indeed, developing animals continue to undergo substantial growth and nerves must grow in length to accommodate the increasing separation between the target and the neuronal soma (Bray, 1984; Heidemann et al., 1995; Pfister et al. 2004; Smith et al., 2001). For example, the giraffe's neck increases by ∼2 cm/d at peak growth (Dagg and Foster, 1982), suggesting that axons in the neck are forced to rapidly expand as well. This recently recognized process of axon stretch growth (ASG) likely represents a secondary phase of rapid and long-term axon growth that drives the formation of long nerves. In adults, it is conceivable that the nervous system is forced to grow when tissue expands such as during pregnancy, with extreme fluctuations in weight, or with the use of tissue expanders for plastic surgery (Battiston et al., 1992; De Filippo and Atala, 2002; Del Frari et al., 2004; Elwood et al., 2000; Hu et al., 2001; Iwahira and Maruyama, 1993; Johnson et al., 1993; Kroeber et al., 2001; Malis et al., 1995; Park et al., 2006; Riordan et al., 2003; Swanson and Argenta, 1988; Takei et al., 1998; Tang et al., 2004; Vekris et al., 1999; Whitesides and Meyer, 2004; Wood and McMahon, 1989; Zeng et al., 2003). Our goal is to identify and explore this unique process in an attempt to discover new mechanisms to enhance regeneration of axons following injury.
The application of stretch growth has been explored previously as a tissue engineering method to produce large living nerve constructs. Far exceeding the rate of growth cone extension, it was found that stretch growth could sustain growth rates of 1 cm/d for several days (Pfister et al., 2004, 2006b). These extreme stretch growth conditions stimulated expansion of the central portion of axon cylinders in caliber as well as length, while maintaining a normal cytoskeletal ultrastructure and the ability to generate and convey action potentials (Pfister et al. 2004, 2006a). Indeed, stretch growth likely induces unknown cellular mechanisms to alter protein synthesis and transport to rapidly assemble the axon. The robust growth rates and lengths accommodated by ASG represent an opportunity to discover new mechanisms to stimulate regeneration. Research from other groups also shows that axons can grow under the application of mechanical force, and that this is a distinctively different process from growth cone growth (Heidemann et al., 1995; Lamoureux et al., 2010; O'Toole et al., 2008).
As biomedical engineers, we dedicated our research efforts to develop the tools needed to investigate the processes that accommodate ASG. The original ASG bioreactor was developed to create copious amounts of nervous tissue for transplantation in animal injury models (Smith et al., 2001; Pfister et al., 2004; Pfister et al., 2006b). This system only allowed examination of fixed time points, limiting analysis to absolute comparisons between differing preparations. To effectively study the biomechanical and biological aspects of stretch growth, the system was re-engineered to accommodate real-time imaging and quantification techniques. Here we report new findings from long-term live microscopic monitoring of axons as they are stretch grown into long axon tracts.
Methods
Principles of operation
The ASG bioreactor was engineered to gradually apply tension to axon bundles spanning two separate culture substrates (Loverde et al., 2011; Pfister et al., 2004, 2006b). To accomplish this, a flexible strip of fluoropolymer film (Aclar 33C film; Structure Probe Inc., West Chester, PA) was positioned over the bottom of the culture chamber. This towing substrate underlaid the moving population of cells. The stationary substrate was the bottom of the culture chamber. First, plated neurons were given time to extend axons from the towing substrate onto the stationary substrate via growth cone extension. The towing substrate was then pulled using a micromotion system, applying a gradual stretch to the spanning axons. Starting as a short distance between the soma and growth cones, the axons were stretch-grown into long fasciculated unidirectional axon tracts several centimeters in length.
Bioreactor culture chamber
The bioreactor culture chamber was designed using Pro/ENGINEER software (PTC, Needham, MA) to produce a 3D schematic for fabrication (Supplementary Fig. 1; see online supplementary material at

Live imaging axon stretch growth bioreactor assembled on top of a Nikon TE2000-S inverted microscope. The system accommodates oil immersion objectives and serves as its own incubator. The major components are labeled. Color image is available online at
The culture chamber stretching frame consisted of three independent lanes (elongated wells) each measuring 1×8 cm that allowed for parallel experiments with a controlled variable. A glass cover-slip measuring 4.8×6.5 cm with #1 thickness (#4865-1; Brain Research Labs, Newton, MA) was glued to the bottom of the stretching frame for each experiment. The cover-slip served as the stationary substrate for the attachment of growth cones, and as a window to permit oil immersion microscopy (Fig. 2).

Bioreactor culture chamber principal stretching components and culture substrates. (
Here the design of the towing apparatus has been improved over previous systems. The Aclar towing substrates were glued to arched legs, which extended down and into each culture lane, rigidly holding the substrates. This design eliminated slack in the towing substrates, and ensured that each manipulation of the towing block resulted in the desired stretch of the axons (Fig. 2A, vii). Adjustment screws allowed the legs to be independently lowered to contact the bottom of the cover-slip.
Conducting sustained, live imaging during stretch growth required the bioreactor to double as an incubator. Temperature was maintained using a closed system of heating elements, thermistors, and a temperature controller, as listed in Table 2. A heated lid was fabricated from a polycarbonate frame with an inset indium tin oxide (ITO)-coated glass slide (Fig. 3A). The base of the culture chamber consisted of a stainless steel heat sink with a flexible silicone heating element to deliver uniform heating across the culture lanes. Both heating elements were controlled by a two-channel temperature controller with thermistors for temperature feedback.

Bioreactor culture chamber incubation components. (
Efficient pH buffering of the culture media was maintained by continuous perfusion of premixed air from a tissue culture incubator. Using an aquarium air pump, air was passed from the incubator through 1/8” ID (3.175 mm) PVC tubing to the culture chamber. An inline aquarium regulator valve was used to control flow rate, while a 0.2-μm syringe filter was used to maintain air sterility. Luer-Lok fittings were used for quick attachment of the tubing to the culture chamber. For humidification, air was routed from the inlet through liquid reservoirs built into the hollowed walls of the culture chamber. Air was returned to the incubator through an outlet located opposite the inlet (Figs. 1 and 3C).
Bioreactor preparation
The towing culture substrates were made from 0.002” (50.8 μm) Aclar film (Table 1) cut into thin strips measuring roughly 25×5 mm using a scalpel. To minimize thickness further, the 5-mm edge of the substrate was gently sanded with 1200-grit sandpaper. The cover-slip and Aclar were washed with laboratory detergent, rinsed with deionized water, and sterilized in 70% ethanol. The bioreactor culture chamber was autoclave sterilized separately and dried inside a tissue culture hood. All subsequent assembly was performed inside the hood using aseptic technique. First, the towing substrates were attached starting with adjustment of the legs fully retracted away from the bottom of the chamber. The backs of the towing legs were coated with silicone RTV (Dow Corning #732; McMaster-Carr) using sterile cotton-tipped swabs. The Aclar strips were applied oriented with the sanded edges facing the lid (Fig. 2B). Next, the bottom of the chamber was lightly coated with silicone glue and the cover-slip was attached (Fig. 2C). Using a swab, excess glue and air pockets were removed by pressing against the bottom of the cover-slip. To allow for complete curing of the glue, the culture chamber was dried in the hood for 2 days.
Prior to cell plating, the towing substrates were lowered into the chamber to make contact with the cover-slip. Optimal overlap was determined to be approximately 2–3 mm as measured from the tip of the Aclar. Each lane was coated at the substrate interface with 1 mL of 10 μg/mL high-molecular-weight poly-D-lysine (PDL, #354210; BD Biosciences, Bedford, MA) in serum-free media for 1 h. The lanes were drained and rinsed gently with deionized water and a final rinse with culture medium before plating.
Neuronal culture
Experiments were performed using dorsal root ganglia (DRG) neurons isolated from embryonic day 15 rat pups. Alternatively, adult DRGs were obtained from rats 16–20 weeks old. Then 5–10 embryonic DRG explants or 25,000 dissociated adult DRGs were plated directly onto the towing substrates within ∼500 μm of the edge using a stereo microscope (Fig. 2D). A puddle of approximately 100–500 μL of medium was formed during plating, in which the cells were allowed to adhere for 1–2 h before filling the lanes with growth medium (Neurobasal w/B-27, 0.5 mM L-Glutamine, 1% heat-inactivated FBS, 2.5 g/L D-glucose, 20 ng/mL NGF, and 20 μM FdU+20 μM uridine). Media changes were done prior to stretch growth in order to avoid disturbance of actively stretch-growing axons. All animal protocols were approved by the Rutgers University IACUC.
Axon stretch growth
The bioreactor culture chamber was docked to a computer-controlled micromotion system consisting of a linear motion table and microstepper motor controlled by a programmable motor indexer (Table 2). A Delrin® chassis was fabricated to position the culture chamber and linear motion table on the stage of the microscope such that they did not move during experimentation (Fig. 1). The linear motion table was attached to the chassis using an acrylonitrile butadiene styrene (ABS) plastic mount printed on a rapid prototype 3D printer (SST 1200es; Dimension, Inc., Eden Prairie, MN). The bioreactor was seated within a square cutout section of the chassis, allowing the microscope objective to contact the viewing window. The towing rods of the bioreactor were fastened to the linear motion table by an ABS adapter printed on the 3D printer.
When axons extended onto the cover-slip by at least 1 mm, typically at 5 days, stretch was applied by towing the neuronal soma away from the growth cones by taking a series of short 2-μm steps. The stretching motion was programmed with motion control software (Si Programmer; Applied Motion Products, Watsonville, CA) that set the following parameters: (1) number of motor steps (the size of the stretch steps); (2) the interval between steps (stretch frequency); and (3) the number of iterations (total stretch length). Stretch growth of embryonic axons was initiated at a net 1 mm/d by taking 2-μm steps every 172 sec over 500 iterations (Table 3). Since stretch-induced growth is strain limited, the stretch rate was begun slowly and gradually accelerated to the desired rate (Pfister et al., 2004, 2006b).
All stretch steps are 2 μm in displacement (10 motor steps=2 μm stretch).
Live imaging
A major obstacle to imaging was the floatation of stretch-grown axons above the working distance of high-magnification objectives. Stretch-grown axons were only adhered to substrates at the proximal and distal ends, while the central portions did not form substrate adhesions during stretch growth. Accordingly, we developed a depressor tool to constrain floating axons within the working distance of high-magnification objectives during microscopy (Supplementary Fig. 2; see online supplementary material at
For phase-contrast imaging, the bioreactor system was set up on a Nikon TE2000-S inverted microscope. To collect long-duration time-lapse sequences, a 4× objective was used with a 0.63× camera lens to increase the field of view to ∼3.5 mm. Time-lapse software (Q-Imaging; Q-Capture Pro, Surrey, B.C., CA) was used to create time-lapse sequences, and was set to acquire one image every 7.5 min over 48-h periods.
For fluorescence imaging, high-magnification images were taken to observe intra-axonal transport during and following stretch growth. Mitochondria were stained with MitoTracker Red (M7512; Invitrogen, Carlsbad, CA), and visualized within stretch-grown axons using confocal microscopy. Briefly, the cultures were stained for 1 min in 100 nM MitoTracker Red and rinsed three times with phosphate-buffered saline. After incubation for 60 min, mitochondria were visualized on a Nikon TE2000-E confocal microscope using a 60× oil immersion objective with 561-nm laser excitation. Select planes were imaged for 10–20 min time-lapse sequences at a rate of 0.5 Hz to analyze mitochondrial movement. Laser power was set to just 0.5% in order to avoid photo bleaching and heating damage to the axons over time. Pixel dwell was further reduced to ∼6 μsec in order to limit exposure of the axons to the laser. It was found that 1.5 sec of scanning with a delay of 0.5 sec produced the best results, enabling time-lapse photography of select axon segments at 0.5 Hz for at least 1 h. While these settings did not provide detailed morphology of mitochondria, they were a necessary trade-off in order to record sustained axonal transport (Miller and Sheetz, 2006).
Time-lapse sequences (stacks) were imported into ImageJ software (National Institutes of Health, Bethesda, MD), where bundles were cropped and rotated to span horizontally (transform/rotate with no interpolation). Kymographs were created by reslicing the axon bundle (stacks/reslice, avoid interpolation, 1.0 spacing) to produce a stack of graphs to measure fast axonal transport. Fast transport was identified by faint diagonal lines, while docked or slow transport was identified by vertical lines. To measure fast transport, diagonal lines were traced in order to find the slope (velocity and direction) and length (distance) of moving mitochondria. All data were exported to Excel (Microsoft, Redmond, WA) for subsequent analysis.
Results
Optimization of incubation for live imaging
Conducting prolonged live imaging of axon stretch growth (ASG) required accurate incubation control for optimal results. To control the temperature outside an incubator and to prevent condensation on the lid (which distorted transmitted light), the device was heated from the top and bottom. The media temperature was maintained within 35–37°C by placing the heat block and heated lid on separate controller channels with independent feedback control. To prevent large fluctuations in temperature, the position of the temperature probes for feedback was important. For the heat block, a temperature probe (thermistor #1, Table 2) was optimally positioned within the culture media at the plating area of the culture lanes (within any of the 3 lanes). For the heated lid, a temperature probe (thermistor #2, Table 2) was positioned on top of the insulating lid, adjacent to the heating element and leads (Fig. 1). Final temperature controller settings for the heat block/lid were 33°/30°C, bandwidth of 11/13, and gain of 50/60, respectively.
To maintain the culture media at a physiological pH of 7.4, mixed air containing 6.5% CO2 was pumped into the bioreactor from a tissue culture incubator. Airflow was adjusted until faint bubbling could be seen within the liquid reservoirs. pH was maintained throughout all experiments by inspection of phenol red and occasional measurement. If the airflow was set incorrectly, the culture media either evaporated or turned basic due to excessive or insufficient flow, respectively. Liquid reservoirs typically lasted 1–2 weeks before drying out when airflow was set correctly.
Unidirectional axon stretch growth
Previous ASG experiments separated two populations of interconnected neurons located on the stationary and towing substrates, resulting in bi-directional axon polarity (Fig. 4A). Here axons were stretch grown unidirectionally with active growth cones at one end (Fig. 4B). Although neuronal soma could be plated on either the towing or stationary substrates, optimal imaging of the distal axon and growth cones was achieved by plating soma on the towing substrates (Fig. 5). However, despite precise plating, a small number of cells frequently migrated onto the stationary substrate (Supplementary Fig. 3; see online supplementary material at

Panoramic images of stretch grown axons (TS, towing substrate; SS, stationary substrate). (

Growth cone motility in coordination with axon stretch growth (ASG). (
Axons from embryonic DRG explants were stretch grown unidirectionally for up to 2 weeks at rates up to 6 mm/d. Stretch growth was initiated at 1 mm/d and increased by 1 mm/d every 24 h according to the rate escalation schedule shown in Table 3. It was found that axons could routinely grow without disconnection at stretch rates up to 4 mm/d over a 5-day escalation schedule (pre-tension, 1, 2, 3, and 4 mm/d). Stretch growth could be consistently sustained at 4 mm/d until the system ran out of travel (over 1 week producing>4-cm-long axons). If, however, the stretch rate was increased on day 6 to 5 mm/d, the axons would start to disconnect within 24–48 h. Since stretch rates above 4 mm/d could not be sustained, 4 mm/d was considered to be the maximum growth rate.
Due to the relevance of repairing adult nerves, we investigated stretch growth of dissociated adult DRG neurons with this new bioreactor. Analysis of time-lapse imaging revealed that dissociated adult neurons do not accommodate the same rate escalation schedule as embryonic explants (Table 3). First, dissociated adult neurons required 10 days to extend axons onto the stationary substrate prior to stretch growth (as compared with 5 days for embryonic explants). Second, using the rate escalation schedule established for embryonic explants, dissociated adult DRG neurons would readily disconnect (Fig. 6). To avoid disconnection, a slower rate escalation schedule was adapted, as shown in Table 4. Using this new schedule, we found that the stretch growth rate could be ramped up to 2 mm/d and sustained for over 24 h without axon disconnection. Upon increasing to 2.5 mm/d, however, axons were observed to begin breaking, and ultimately resulted in the disconnection of the majority of axons. Interestingly, in one case, stretch growth of adult axons was achieved at the same 4 mm/d maximum growth rate of embryonic explants. While dissociated undirectional adult axons reached a maximum of 2 mm/d, here we found that incompletely dissociated bidirectional adult axons demonstrate the ability to reach and sustain 4 mm/d growth for 2 days (Fig. 6).

Adult dorsal root ganglion (DRG) rate escalation schedule (X, disconnection of axons). Axons in unidirectional polarity successfully reached 2 mm/d on numerous occasions, and disconnected upon rate increases to 2.5 mm/d. On one occasion during stretch growth of incompletely dissociated and bidirectionally oriented axons, stretch growth reached 4 mm/d without disconnection.
All stretch steps are 2 μm in displacement (10 motor steps=2 μm stretch).
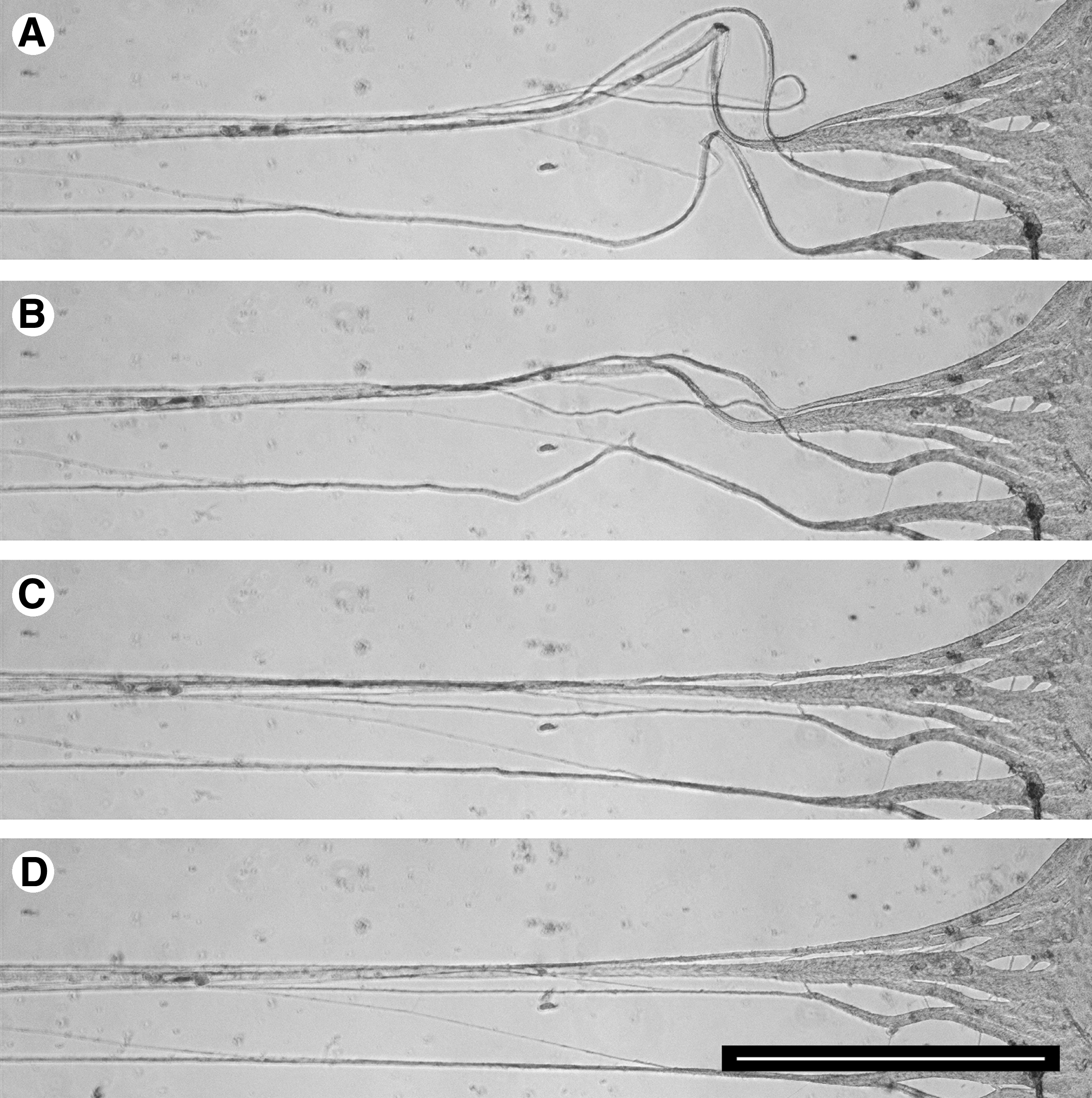
Stretch-growing axons from both embryonic and adult neurons formed bundles as they elongated. Time-lapse imaging showed that axons in close proximity fasciculate by collapsing at the bifurcation point, which proceeds toward the end until axons form a bundle (Fig. 7). However, if the bifurcation of two axons formed a large angle, fasciculation would not occur (Fig. 8A, axon v). The size distributions of axon bundles were analyzed from embryonic and adult neurons stretch-grown over 2 cm in length (when most fasciculation had occurred). Embryonic and adult bundles had similar diameters, ranging from 5–15 μm (Fig. 8).
Fasciculation of axons in coordination with stretch growth. (

Diameter variations in stretch-grown axon bundles. (
Growth cone motility in coordination with stretch growth
Time-lapse imaging of unidirectional stretch growth revealed dynamic growth cone behavior. While embryonic axons were stretch-grown at a rate of 1 mm/d, their growth cones were imaged on the cover-slip stationary substrate. Growth cone activity was characterized by two predominant outcomes. First, a portion of growth cones were found adhered to non-solubilized lysine aggregates on the stationary substrate, where they remained attached for the duration of the experiment (Fig. 5A). Conversely, growth cones that adhered to the solubilized PDL coating were migratory. Time-lapse analysis revealed a dynamic movement of growth cone retraction, branching, and extension, as axons were stretch grown.
Upon the start of stretching, growth cones retracted in the same direction as the applied stretch. Within hours of continuous stretching, the initiation of growth was evident as growth cones transitioned from retraction to branching and extension, which enabled them to maintain their relative positions. In some instances, overall extension exceeded retraction and was evident as growth cones surpassed their original locations. As shown in Figure 5, a motile population of growth cones retracted an average 300 μm over the first 10 h of stretching at 1 mm/d (400 μm stretch), resulting in a retraction rate of 720 μm/d. After 10 h, growth cones no longer retracted, but extended 377 μm over the following 35 h, resulting in a growth cone extension rate of 260 μm/d. Combined elongation of axons and growth cones was 1.2 mm/d, exceeding the stretch rate of 1 mm/d, as calculated from the 10-h time point. Over time and in coordination with stretch rate increases, growth cone motility appeared to diminish and become static.
Fast axonal transport of mitochondria
Samples stained with MitoTracker revealed abundant mitochondria within all stretch-grown axons (Fig. 9A). Due to the thickness of axonal bundles, tracking mitochondria within whole bundles was not possible at the selected imaging rate of 0.5 Hz. Instead, select planes were chosen based on the focus of individual axons that had minimal background noise caused by neighboring axons. Optimal imaging was found by focusing on the bottom of the bundles, where the majority of neighboring axons were outside of the focused plane.
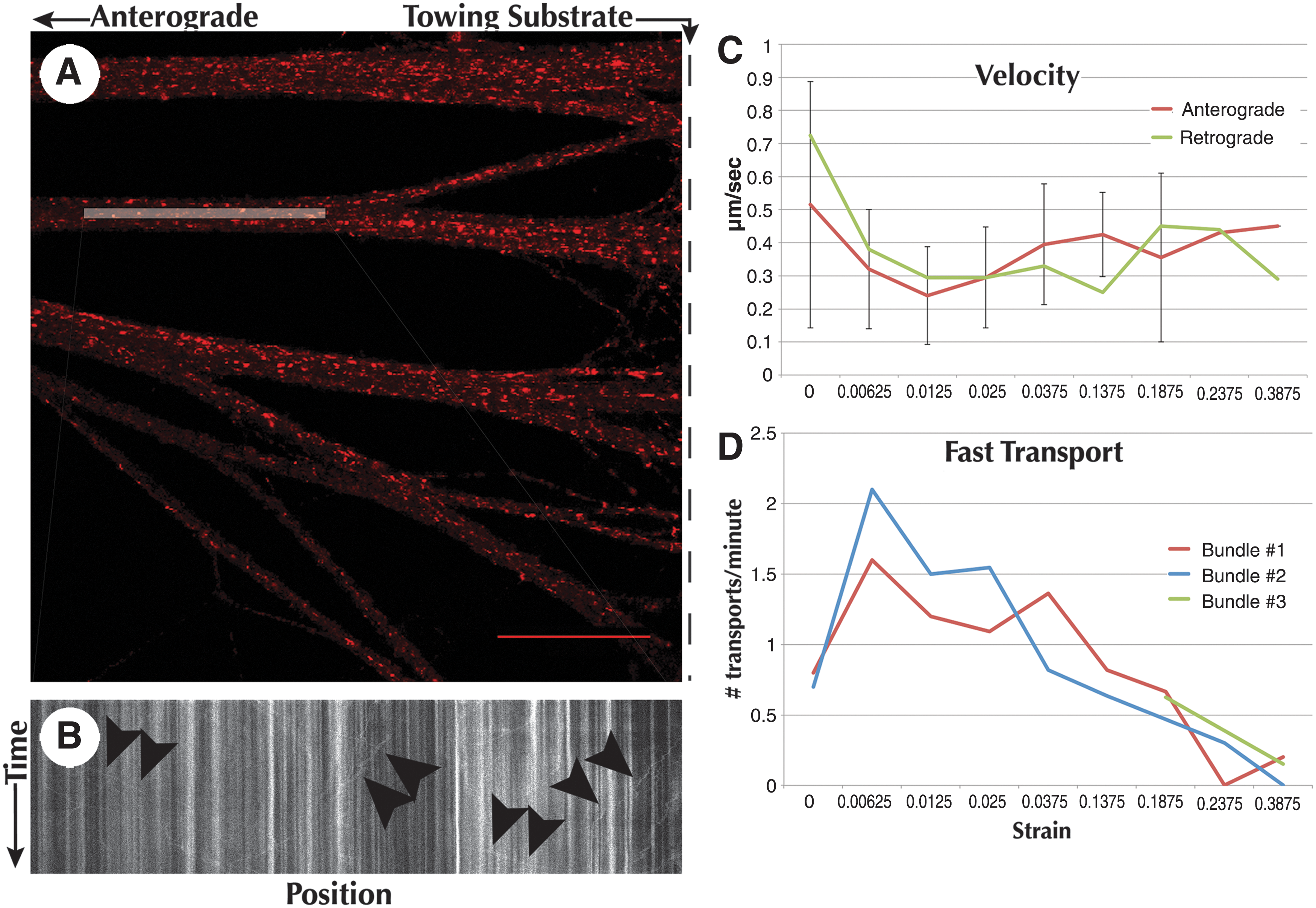
Decrease in fast axonal transport due to strain. Mitochondria were labeled with MitoTracker Red and imaged within the bioreactor on a confocal microscope utilizing a 60× oil immersion objective. (
Mitochondria were tracked at increasing displacement step sizes to determine the effect of strain on fast axonal transport. Embryonic axons stretch-grown to 4 mm in length were given 2 weeks to re-establish basal resting tension. Two parallel axon bundles were imaged simultaneously at the proximal segment within 500 μm of the towing substrate. Sequential 10- to 20-min time-lapse sequences were taken between periods of nearly instantaneous (10 μm/sec) stretch until fast axonal transport was diminished (Fig. 9D).
Averaged data recorded from both axon bundles (Fig. 9D), revealed that anterograde and retrograde fast transport were reduced from an average 0.75 transports/min to 0.57 transports/min at a strain of 19% (750 μm stretch of 4-mm axon). Continued stretching resulted in a further reduction to 0.15 transports/min at 24% strain (950 μm stretch of 4-mm axon), and 0.10 transports/min at 39% strain (1550 μm stretch of 4-mm axon). To confirm this trend, an alternate bundle, which was not previously imaged, was used as verification. Imaging of the alternate bundle revealed 0.63 transports/min at 19% strain, decreasing to 0.15 transports/min at 39% strain, confirming the trend set by the previous bundles. No axons disconnected during testing, which subsequently remained intact for 6 days following experimentation.
The velocity of fast anterograde and retrograde transport remained constant at all applied stretches, producing an average 0.37±0.17 μm/sec and 0.38±0.19 μm/sec, respectively (Fig. 9C). The maximum recorded fast axonal transport rate of 2.57 μm/sec occurred prior to stretching, in both anterograde and retrograde directions. Further, there appeared to be no difference in the average distance traveled by each mitochondrion throughout experimentation, resulting in an average of 17.59±11.49 μm of movement.
Retraction of stretch-grown axons
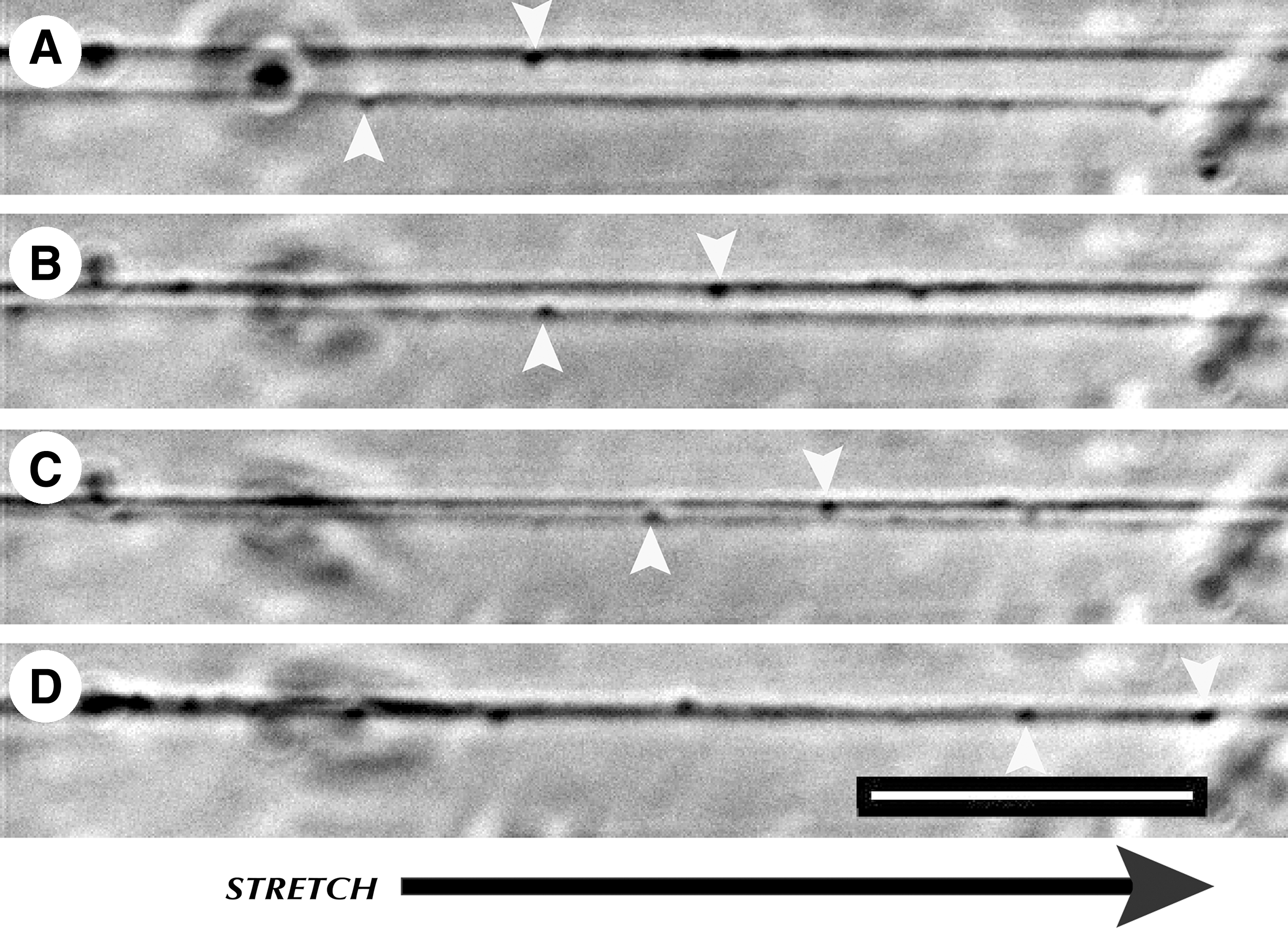
Serendipitously, we found that stretch-grown axons have the ability to retract great lengths to restore tension in the axon fascicles. To further investigate this phenomenon, embryonic axons were stretch grown to 4 cm in length and rested for 1 day to re-establish basal resting tension. Tension was spontaneously relieved by reversing the direction of the towing substrate. A 2-mm retraction (−5% strain) was applied over 6 sec (333 μm/sec), producing undulations in the proximal axon segment nearest the towing substrate (Fig. 10). Using time-lapse imaging set to take images at 0.2 Hz, we monitored the proximal segments of 7 axon bundles from 3 separate preparations as they re-established their original taught appearances. ImageJ was used to analyze time-lapse data by measuring the change in length of undulated axons following slackening. An average initial retraction rate of 6.1 μm/sec was calculated following relief of tension over the first 60 sec. The rate slowed to 3 μm/sec over the following 3–5 min, and further slowed to 0.06 μm/sec over the remaining 3–7 min. Retraction was visually complete within 7–13 min for all axons, which subsequently remained taught and intact for at least 1 week following experimentation. Interestingly, in one trial of adult axons, an average retraction rate of 0.3 μm/sec was calculated over 45 min following initial retraction as axons appeared to re-establish tension. Undulations in both embryonic and adult axons always occurred within the first 5 mm of the proximal segment, while the remaining ∼3.5 cm of the axon was relatively straight.
Retraction of stretch-grown axons. (
Discussion
A new bioreactor system was engineered to accommodate live and prolonged imaging of stretch growth in order to study how nerves grow following growth cone extension, and explore their behavior under mechanical stimulus. Developing this bioreactor was a process of design, construction, and testing. Here we discuss operation of the device, critical improvements in the technique, and new findings that could only be observed from long-term live microscopic monitoring of axons as they are stretch grown into long axon tracts.
To maintain stable temperature of the culture media, independently controlled heating from the top and bottom of the culture chamber was required. Despite our low temperature settings of 33°/30°C, the heating elements exceeded those temperatures by approximately 10°C. On initial attempts, choosing a temperature setting of 37°C caused the heating elements to rapidly exceed 60°C on startup due to the distance between the heating elements and thermistors. Our final settings were chosen by monitoring temperature in several locations, while reducing the gain and bandwidth settings to limit power to the heating elements and provide steady temperature control.
Improvements in substrate attachment and coating
Previous work established that small and frequent stretches are important to the survival of axons undergoing stretch growth, particularly at the start of stretching (Pfister et al., 2004, 2006b). While gradual application of stretch seemed intuitive, axonal disconnection from larger stretches was never observed in real time. A significant finding from time-lapse imaging revealed that while the linear motion table was making the programmed 2-μm steps, movement of the towing substrates was out of sync. The 2-μm steps accumulated and released in jumps of 10 μm or more at the towing substrate. This repeated occurrence caused some growth cones to disconnect from the stationary substrate, and other axons to thin and disconnect at the start of stretching.
Jumping was thought to occur due to stiction (sticking and friction forces) between moving components, including the towing and stationary substrates. The 90° bend of the Aclar towing substrates was thought to be a major site of accumulation of asynchronous steps. We solved these issues by incorporation of rigid towing legs that eliminated flexing of the Aclar towing substrates. The ∼88° arched shape of the legs also helped to provide consistent tension to the tips of the towing substrates where they contact the cover-slip (Fig. 2B). We also reduced friction in the system by increasing tolerances where the towing rods enter the culture chamber. Together, these modifications resulted in precise, synchronous movements of the linear motion table and towing substrates during stretch growth and retraction.
A major improvement was made in neuronal adhesion by using PDL in place of collagen as the substrate coating. Our previous experience with collagen was an unreliable peeling from Aclar surfaces, which would often cause experiments to fail. This peeling was exacerbated on glass. In addition, explants plated on collagen were occasionally dislodged and pulled into the central portion of stretch-grown axons, as seen in Figure 4A. By using high-molecular-weight PDL at high concentration, neuronal cultures maintained greater adhesion to the cover-slip and Aclar substrates, while eliminating movement of explants due to stretching forces.
Unidirectional stretch growth
Plating the neuronal soma on the towing substrate to create unidirectional axon polarity was a major change in our stretch growth protocol. Previously, neuronal soma were plated evenly on both the towing and stationary substrates (Pfister et al., 2004; Fig. 4A). The axons spanning the two populations of cells integrated into the opposing population and visual evidence of axon growth ceased. The resulting stretch-grown bundles had axons polarized in both directions. This arrangement, however, would make it difficult to examine many directionally-dependent processes.
It was previously reported that embryonic explants could be stretch grown up to 10 mm/d (Pfister et al., 2006b). This is indeed true for axons that are stretch grown bidirectionally with soma at each end. In the unidirectional case here, we found that axons could only sustain growth at a rate of 4 mm/d using the same rate escalation schedule established for bidirectionally-polarized axons. We propose two possible explanations for this difference. First, bidirectional axons are growing from both directions, splitting the growth requirements needed to meet the overall stretch. Clearly, the behavior in the middle is unknown. Bidirectional axons stretch grown at high rates may not extend across the entire span of axon bundles. Second, our current experiments involve plating far fewer explants or dissociated cells than the original tissue engineering method, a tradeoff made in order to improve visualization. Plating density and cell-to-cell signaling may play an important role in the growth and survival of stretch-grown axons, and may also factor into the growth rate differences we found between embryonic explants and dissociated adult neurons.
Stretch growth of adult neurons
Work on ASG has been primarily based on DRGs from the embryo. While this adds to our knowledge of nervous system development, application to nerve injury and repair would be better investigated using DRGs from the adult (Pfister et al., 2011). In one instance, it was shown that axons from human adult DRGs could undergo stretch-induced growth (Huang et al., 2008). This study establishes that axons from dissociated adult rat DRG neurons can undergo approximately half the maximum stretch growth rate of embryonic explants. Further, adults were only capable of half the rate escalation schedule as explants (Tables 3 and 4). Under this new rate escalation schedule, our results suggest that adult axons are consistently limited to a rate of 2 mm/d. Interestingly, however, we did have one instance where bidirectionally plated adult axons withstood a rate of 4 mm/d over a 48-h period. While many axons in this experiment failed, the surviving axons support our hypothesis that bidirectionally polarized neurons may not span the entire length of axon bundles, and grow from both ends. However, further study of dissociated embryonic neurons is needed to confirm that the difference in growth limit is in fact due to age, and not due to plating differences.
Stretch, strain, and occlusion
Several studies have shown that axons grow in response to applied tension. If tension is kept above a resting threshold, axons grow. Conversely, if tension is released to below the resting tension level, axons retract (Dennerll et al., 1989; Heidemann and Buxbaum, 1990; Zheng et al., 1991). In this study, we identified growth rate limits by applying predetermined stretch instead of measuring tension. The stretch rate limit was considered to be equivalent to the maximum growth rate, since axons never broke at these stretch-growth rates. In practice, we were only able to sustain stretching of axons equivalent to their resulting growth. Stretch growth is a continuous process of never allowing axons to reach resting tension, maintaining a perpetual state of growth. Axons only tolerated stretch rates beyond their maximum growth rate temporarily (5 mm/d for embryos, 2.5 mm/d for adults), but consistently disconnected if these rates were sustained.
Finding the stretch rate limit was a practice of applying stretch that resulted in growth, and not disconnection. However, it is not sufficient to only use spontaneous disconnection as a measure for stretch rate limit. It is possible that excessive stretching can lead to pathologic occlusion, inhibiting growth of new axon, resulting in delayed disconnection with subsequent stretching. As supported by our observations of delayed growth cone extension following stretching, there is a delay between the stimulus of stretch and the resulting growth. The greatest risk of occlusion is likely to occur during stretch rate changes, when there is a large difference between the stretch rate and the growth rate. Indeed, the risk of disconnection seemed to decrease in longer axons stretched at a constant rate.
We theorized that fast axonal transport would diminish once occlusion occurred. Static axons at resting tension were subjected to increasing strain to determine the relationship between strain and occlusion. Fast axonal transport appeared to diminish at a strain of 39%, but did not result in axon disconnection. Despite apparent occlusion, axons remained intact for 6 days, after which they were capable of subsequent stretching. A possible explanation is that axons may have recovered from temporarily induced occlusion. Axonal tension may be initially localized to the central segment during rapid stretches, but slowly dissipated throughout the proximal and distal segments and ECM over time (O'Toole et al., 2008). A second explanation could be that axonal transport was reduced but not completely diminished. Since our imaging technique was developed to attain long duration sequences quickly (to avoid photobleaching), the sensitivity of our technique is low and may not have detected all mitochondrial transport. Despite our calculation of occlusion strain, applied strain during ASG is relieved continuously due to growth of the axon. Indeed, during ASG, fast transport of mitochondria was observed consistently.
Finally, we note that thicker bundles were more resilient to disconnection than single axons. In experiments exploring the maximum stretch-growth rates of embryonic and adult axons, the thickest bundles were consistently the last to break. Despite this observation, there was no evidence that stretching of long or thick bundles correlated with increased growth rates. It is conceivable that axon-axon adhesions formed during bundling reinforced axonal cytoskeletons, prolonging imminent occlusion. Alternatively, bundling may have simply increased tensile strength.
Conclusion
This new system allows for the simultaneous study of biomechanical and molecular mechanisms. Identification of the processes initiated in response to axonal tension may provide novel targets in the therapeutic treatment and research options in repair of the nervous system. The overall expectation of the live imaging ASG bioreactor is to increase our fundamental knowledge of what occurs during this natural process. In particular, the cellular processes that drive this endogenous nerve growth can be explored with state-of the-art imaging technologies. Our research goal is to decipher the unique underlying biology that promotes sustained and robust growth in order to exploit these processes to promote nerve regeneration. Using this new system, we now have the ability to differentiate the mechanotransduction between different neuronal types and culture conditions.
Though this system has matured greatly, it relies on displacing axons to a preset strain. It is known, however, that axons grow in response to increased tension (Heidemann and Buxbaum, 1990, 1994; Lamoureux et al., 2010; Zheng et al., 1991). A future consideration of this device is incorporating force measurement capabilities. The ability to measure axonal tension will be important for understanding mechanotransduction and maintenance of axon tension.
Footnotes
Acknowledgments
This work was supported by the New Jersey Commission on Brain Injury Research 07-3204-BIR-E-0 and the NSF CAREER CBET-0747615. We thank Mr. John Hoinowski, NJIT Biomedical Engineering Design Studio, for his ingenuity and construction of the device components. Importantly, thanks also go to Douglas H. Smith, M.D. and David F. Meaney, Ph.D. for their continued mentorship and support. We also thank Dr. Kyle Miller for his protocol on labeling and imaging mitochondria in axons.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.