
Abstract
Sports concussions affect thousands of individuals every year and are a major public health concern. Still, little is known about the long-term and cumulative effects of concussions on brain neurophysiology. The principal objective of this study was to investigate the long-lasting effects of multiple sports concussions on sensorimotor integration and somatosensory processing in a sample of 12 concussed athletes and 14 non-concussed athletes of similar age (mean, 23 years) and education (mean, 16 years). Right median nerve stimulation was paired with transcranial magnetic stimulation (TMS) of the left primary motor cortex to investigate sensorimotor integration with short latency afferent inhibition (SAI) and long latency afferent inhibition (LAI) at five interstimulus intervals (18, 20, 22, 100, 200 msec). Somatosensory evoked potentials (SEP) were recorded from the left centro-parietal region. We also investigated primary motor cortex inhibitory mechanisms with three TMS protocols: cortical silent period, long interval intracortical inhibition, and short interval intracortical inhibition. Motor evoked potentials were recorded from the right abductor pollicis brevis muscle. No differences were observed between groups for SAI, LAI, and SEP. However, cortical silent period duration was prolonged and long interval intracortical inhibition was enhanced in the concussed group. These findings suggest that multiple sports concussions lead to specific, long-term neurophysiological dysfunctions of intracortical inhibitory mechanisms in primary motor cortex while somatosensory processing and sensorimotor integration are spared. This study provides additional evidence for the presence of specific and stable alterations of GABA
Introduction
T
Post-concussion symptoms usually consist of headache, dizziness, visual difficulties, memory disturbance, and concentration problems (Cantu, 1996). It is generally agreed that post-concussive symptoms typically disappear between 2 to 10 days after the incident (McCrory et al., 2005), a time window that closely coincides with the resolution of the neurometabolic cascade of concussion (Giza and Hovda, 2001). In addition to the transient effects on cognition, motor function alterations in the form of gait stability and balance control have been documented in the acute post-concussion phase (Cavanaugh et al., 2005; Guskiewicz et al., 2001; Parker et al., 2006). In the 48 hours following brain injury, athletes display changes in postural control that appear to be linked to deficits in sensory interactions between the visual, somatosensory, and vestibular systems (Guskiewicz et al., 2001). Furthermore, Catena and collaborators (2007a, 2007b) have shown that concussed individuals show increased medial/lateral motion in gait stability tasks.
Despite the apparent ephemeral nature of post-concussive symptomatology and the uncommon presence of tissue damage using routine imaging techniques such as CT scan (Kibby and Long, 1996), recent studies have suggested detrimental long-term effects of concussions, which might eventually evolve into devastating neurological conditions with aging. Neuropathological analysis of brain tissue has revealed chronic traumatic encephalopathy, a progressive tauopathy, in athletes who suffered multiple concussive injuries (McKee et al., 2009), and diffusion tensor imaging (DTI) has shown numerous white matter alterations after mild TBI (Maller et al., 2010). Studies have also shown that approximately 17% of retired professional boxers will eventually develop dementia pugilistica, a disorder characterized by motor and cognitive symptoms resembling those of Parkinson's disease (Rabadi and Jordan, 2001). The exact mechanism by which concussions contribute to the development of severe neurological conditions remains largely unknown; only a handful of studies have investigated the neurophysiological impact of brain injury in otherwise healthy individuals beyond the acute phase.
Despite the fact that concussed athletes typically stop complaining about motor symptoms 10 days after concussion, a recent study suggests long-lasting gait stability abnormalities whereby athletes showed altered postural stability in a dual task more than 28 days following concussion (Parker et al., 2006). Supporting these findings, neurophysiological studies of mTBI using transcranial magnetic stimulation (TMS) have recently shown primary motor cortex (M1) dysfunctions in the acute phase (Chistyakov et al., 2001) that seem to be long-lasting (De Beaumont et al., 2007). In a series of studies, Chistyakov and collaborators (1998, 1999, 2001) used TMS to investigate the presence of M1 neurophysiological alterations after traumatic brain injuries of different severity. These investigators reported altered motor cortex excitability in minor-to-severe TBI (Chistyakov et al., 1998, 1999, 2001), as well as prolonged cortical silent period (CSP) duration in mild-to-moderate TBI (Chistyakov et al., 2001), which is indicative of dysfunctional intracortical inhibitory systems. While these results documented immediate neurophysiological dysfunctions, the possible persistence of those alterations was unknown. In fact, it is only recently that researchers have investigated the possible long-term effects of sport concussions on M1 neurophysiology. Using TMS, De Beaumont and collaborators (2007a) have reported the presence of long-lasting intracortical inhibitory system abnormalities within the primary motor cortex of university football athletes who sustained multiple concussions. In line with the results of Chistyakov and colleagues (2001), they observed a lengthening of the CSP, which was linked to concussion severity but was independent of the amount of time elapsed since the last concussion. This suggests that sports concussions can produce neurophysiological alterations that persist well beyond the acute phase. Although the neurophysiological underpinnings of the CSP are still debated, the majority of pharmacological studies have attributed CSP lengthening to alterations of GABA
In addition to specific alterations of GABAB receptor activity, there is evidence for an alteration of cholinergic systems in severe traumatic brain injury resulting in diffuse axonal injury (Fujiki et al., 2006), where short latency afferent inhibition (SAI), a marker of cholinergic activity involved in sensorimotor integration, was reduced (Tokimura et al., 2000). Alterations in cholinergic activity and motor inhibitory circuits have been found in diverse pathologies such as Gilles de La Tourette syndrome (reduced SAI, Orth et al., 2005), asymptomatic Parkin mutation carriers (reduced SAI, Bäumer et al., 2007), Alzheimer's disease (reduced SAI, Di Lazarro et al., 2002, 2004, 2005), and Parkinson's disease (reduced long latency afferent inhibition [LAI], Sailer et al., 2003). Knowing that the aforementioned pathologies all share motor/memory dysfunctions similar to those associated with post-concussion syndrome (Jotwani and Harmon, 2010) and that SAI/LAI interacts with concussion-vulnerable inhibitory circuits of the primary motor cortex for sensorimotor integration (Chen, 2004), assessing the integrity of this system in concussed athletes is of particular clinical interest.
In contrast with the growing body of evidence for M1 inhibitory mechanism alterations (De Beaumont, 2007a, 2009), few concussion studies have investigated the presence of neurophysiological abnormalities in non-motor areas. Somatosensory evoked potential (SEP) testing represents a useful technique to assess the integrity of somatosensory cortex, as well as afferent conduction, and it is routinely used as a clinical tool in severe TBI to predict functional recovery (Chistyakov et al., 1999; Lew et al., 2003). Abnormalities in the SEP N20 component were found in comatose, diffuse axonal injury patients and in moderate brain injury victims (Chistyakov et al., 1999), and altered N20 latency seems to be related to clinical disability in severe TBI (Rappaport et al., 1990). Abnormal N60 latencies lasting up to three months were also found with concussion in a sample of consecutive patients presenting to the emergency unit (Zumsteg et al., 2006). Prolonged central sensory conduction time (CSCT) has also been reported in the acute coma phase after brain injury (Chistyakov et al., 1999) and abnormalities in central motor conduction time (CMCT) were found after brain injury, mostly in patients who sustained axonal damage (Chistyakov et al., 1999).
The principal objective of this study was to investigate the specificity of the previously reported long-lasting inhibitory dysfunction in primary motor cortex of concussed athletes. To this end, a comprehensive neurophysiological evaluation of sensorimotor function was performed in concussed athletes to better circumscribe areas of dysfunction in the aim of developing objective markers of concussion to facilitate diagnosis, gather prognostic insights, and facilitate return-to-play decisions. This is especially relevant in the case of asymptomatic athletes, in whom neuropsychological testing, neurological examination, and symptom checklist often fail to reveal any lingering dysfunction. A sample of symptom-free concussed athletes who sustained their last concussion on average two years prior to testing were assessed on the following measures: short latency afferent inhibition, long latency afferent inhibition, and somatosensory evoked potentials. Furthermore, the integrity of ascending and descending pathways was evaluated with sensorimotor conduction times. Finally, M1 GABA-mediated intracortical inhibition was evaluated with measures of short interval intracortical inhibition, long interval intracortical inhibition, and cortical silent period.
Methods
Participants
Data were obtained from 26 participants who were active football players from Canadian university football and were recruited through the team physician. The following exclusion criteria were used to determine participation in the study: no history of psychiatric illness, learning disability, alcohol or drug abuse, neurological condition (i.e., seizures, brain tumor), or TBI unrelated to sports or medical conditions requiring daily medication. The inclusion criteria for concussed participants were two or more concussions, last concussion more than 12 months prior to testing, absence of symptoms, and active university-level football player. Participants were all right handed. The study was approved by the local ethics committee and all participants provided written informed consent prior to testing. Subjects received a financial compensation of Can$60 for their participation.
The study included two experimental groups. The first group consisted of 14 athletes with no history of sports concussion with a mean age of 23 years (mean, 22.36; SD=1.69) and a mean level of education of 16 years (mean, 15.93; SD=1.33). The second group consisted of 12 athletes with a history of two or more sports concussions (mean, 3.25; SD=0.97) that occurred more than one year prior to testing (mean, 23.17 months; SD=5.92). Concussion history was based on medical records for accidents that occurred throughout the athletes' university years while previous concussion history was self-reported. At the time of testing, concussed athletes were asymptomatic, reporting very few, if any, symptoms on the post-concussion symptoms scale (PCS, mean, 2.15; SD=2.08) (Maroon et al., 2000). Concussion severity ratings were provided by the team physician and were graded according to the American Academy of Neurology parameters (1997), from grade 1 (confusion for less than 15 min without amnesia or loss of consciousness) to grade 3 (loss of consciousness, duration either brief [seconds] or prolonged [minutes], with a mean grade of severity of 2 (mean, 2.00; SD=0.67). All concussions were rated as mild (score of 13 to 15) on the Glasgow Coma Scale.
Procedure
The experiment consisted of a single 90-min testing session during the football off-season. This session included the administration of a concussion history questionnaire, a general health questionnaire, the PCS (refer to De Beaumont et al., 2007b, for details on questionnaires), and the acquisition of TMS and SEP recordings.
TMS recordings
TMS was delivered through an 8-cm figure-of-eight coil connected to a MagPro transcranial magnetic stimulator (Medtronic, Minneapolis, MN). The stimulating coil was placed flat on the skull with the handle pointing backwards and 45° away from the midline. The induced current flow was biphasic with a posterior-anterior direction. Pulses were delivered over the optimal position to elicit a maximal electromyographic (EMG) response of the controlateral abductor pollicis brevis (APB) muscle to conform to afferent inhibition paradigms (Kessler et al., 2005). The EMG signal was amplified using a Powerlab 4/30 system (ADInstruments, Colorado Springs, CO), filtered with a band pass 20–1000 Hz and digitized at a sampling rate of 4 KHz. Motor evoked potentials (MEPs) were recorded using Scope v4.0 software (ADInstruments) and stored offline for analysis. A Brainsight frameless stereotaxic system (Rogue Research, Montreal, Canada) was used to ensure stable coil positioning over the stimulation site.
Intracortical inhibition
The resting motor threshold (rMT) was first established as the minimum stimulation intensity necessary to evoke MEPs of 50 μV in 50% of 10 consecutive trials while the targeted hand was at rest. According to the method described by Kujirai and collaborators (1993), short interval intracortical inhibition (SICI) was elicited by applying a sub-threshold conditioning stimulus (80% of the resting motor threshold) 2 msec before a supra-threshold test stimulus (TS) adjusted to reliably induce MEPs of approximately 1 mV peak-to-peak amplitude. Ten MEPs were recorded for this SICI paradigm. A single pulse TS condition of 15 consecutive trials was used as baseline. To evoke long interval intracortical inhibition (LICI), two pulses set at an intensity that produced a TS MEP between 0.20 and 1.50 mV were administered with an interstimulus interval of 100 msec. Ten pairs of such MEPs were collected. To induce a cortical silent period (CSP), single-pulse stimulations set at TS intensity (1 mV peak-to-peak amplitude) were applied over the left primary motor cortex while the participant maintained a voluntary isometric contraction of the right APB muscle at approximately 10% of maximal strength. Ten MEPs were recorded for this condition. TMS paradigms were delivered in a pseudo-randomized order.
LICI was expressed as the ratio of the test stimulus relative to the conditioning stimulus, whereas SICI was measured by comparing MEP amplitude evoked by the TS when preceded by the conditioning stimulus with that elicited in the unconditioned condition (TS alone). The length of the CSP was assessed manually and was defined as the period from the onset of EMG suppression until the resumption of sustained post-stimulus EMG activity.
Spinal and central motor conduction times
To obtain spinal conduction time, TMS stimulations were applied directly on the surface of the C7-C8 dorsal root at an intensity of 60% of maximal stimulator output. Spinal conduction time was defined as the period between stimulation and onset of EMG response. Corticospinal conduction time was the mean latency of the MEPs recorded after TMS stimulation over M1. To obtain central motor conduction time, the mean latency of the spinal stimulation was subtracted from the mean corticospinal stimulation latency. Ten MEPs were recorded from the APB muscle for each condition.
Afferent inhibition by somatosensory input from the hand
Afferent inhibition was elicited by applying a median nerve electrical conditioning stimulus followed by a TMS test stimulus at different time intervals. To elicit short latency and long latency afferent inhibition, a Grass S88 stimulator (Grass Co., Quincy, MA) was used to stimulate the median nerve at the level of the right wrist. Standard bipolar electrodes were used with the cathode positioned proximally. The electrical stimulation consisted of a square wave pulse of 0.2 msec duration. Conditioning electrical stimulation (CS) intensity was adjusted slightly over the threshold to evoke a small muscle twitch at the thumb. The TS intensity applied over the left motor cortex was adjusted to evoke an MEP in the resting APB of approximately 1 mV peak-to-peak amplitude. In keeping with previous studies, interstimulus intervals (ISI) of 18, 20, and 22 msec between the CS and TS were used to produce short latency afferent inhibition (Tokimura et al., 2000) while ISIs of 100 and 200 msec were used to elicit long latency afferent inhibition (Chen et al., 1999; Nakamura et al., 1997). A TS-alone control condition was also performed to subsequently compute ratios of the mean amplitude of the MEPs recorded for each ISI with respect to the mean amplitude of the control, unconditioned response (SAI and LAI conditions/TS alone). Due to technical difficulties, data for SAI and LAI were not collected in four participants (two in each group).
Somatosensory evoked potentials recordings and data analysis
In a subset of 21 participants (9 concussed, 12 controls), somatosensory evoked potentials were recorded. Electrical stimulation was applied with a Grass S88 stimulator to stimulate the median nerve at the level of the right wrist through standard bipolar electrodes, with the cathode positioned proximally. A square wave pulse of 0.2msec duration was used. Stimulation intensity was adjusted to evoke a small muscle twitch at the thumb. For SEP recordings, a 32-channel acquisition system (Neuroscan Labs, El Paso, TX) was used. Continuous EEG signals were recorded from the seventh cervical vertebra (Cv7), and four stainless steel electrodes were placed on the following sites according to the 10–20 International system: C3′, C4′, Oz, and Fpz (Cooper et al., 1980). The left and right mastoids were used as references and the electrode Fpz as the ground. Skin-electrode impedance was kept under 5000 ohms. The session consisted of 500 electrical stimulations at 3 Hz. The timing was controlled by PsyScope X software running on a MacBook Pro computer (Apple, Cupertino, CA).
BrainVision Analyser software (Brain Products, Munich, Germany) was used for data analysis. The duration of a single epoch was 100 msec with a pre-stimulus period of 30 msec. A semi-automatic artifact exclusion was performed where epochs including blinking or ocular movements and cardiac artifacts exceeding 150 uV in peak-to-peak amplitude were excluded. Averaged N20 component amplitude recorded at C3′ electrode was measured from peak-to-peak amplitude with P27, which is the first positive component following the N20 peak. N20 and N13 latencies were also measured. The central sensory conduction time (CSCT) was defined as the interpeak latency of the cervical N13 and cortical N20.
Statistical analysis
All values are expressed as means plus/minus standard deviations. Intracortical inhibition measures, SAI/LAI, SEP data, and central conduction time data were subjected to standard descriptive statistics and ANOVAs. Independent sample t tests were performed to assess differences between the two groups.
Results
Intracortical inhibition of M1
In agreement with previous findings (De Beaumont et al., 2007a, 2009), increased intracortical inhibition was found at baseline in athletes with a history of multiple concussions. Compared to their unconcussed counterparts, LICI was significantly enhanced (t 24=2.11; p=0.05) and CSP duration was significantly prolonged (t 24=2.35; p=0.03). Consistent with recent findings, SICI did not differ between groups (t 24=1.67; p=0.11; Table 1).
Short and long latency afferent inhibition
SAI
A one-way ANOVA was first performed on raw MEP amplitude to verify that the CS modulated TS response. There was a main effect of ISI (F3, 23=13.11; p=0.0001), which was caused by inhibition of the conditioned TS compared to TS alone at all intervals. Using the ratio of the conditioned stimulus over the test stimulus alone, a mixed ANOVA (Group X ISI) revealed no main effect of ISI (F2, 23=2.88; p=0.08), no main effect of group (F2, 23=0.02; p=0.88), and no interaction (F2, 23=0.15; p=0.80; Fig. 1A).
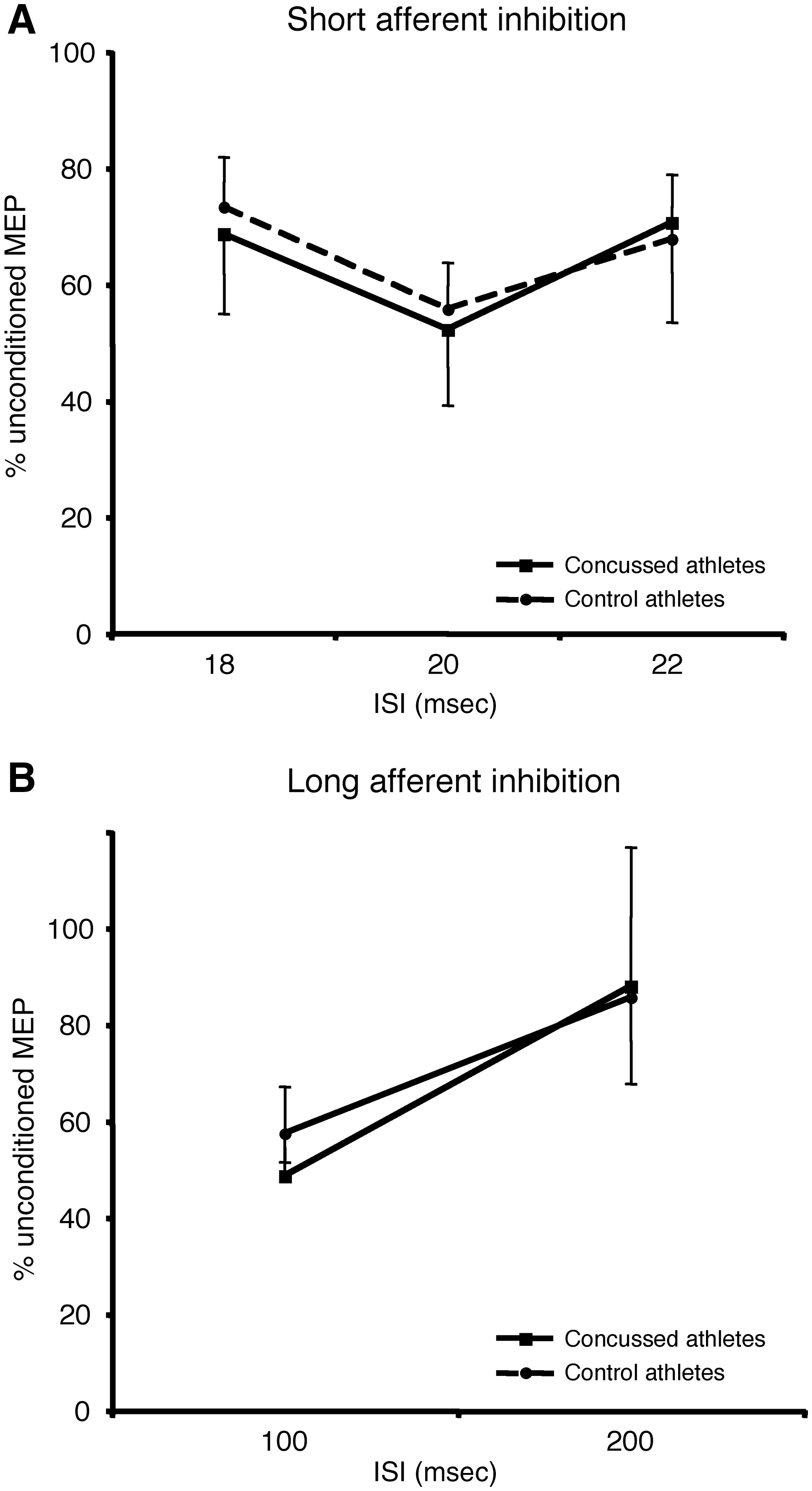
(
LAI
A one-way ANOVA was first performed on raw MEP amplitude to verify that the CS modulated TS response. There was a main effect of ISI (F2, 23=16.78; p=0.0001), which was caused by inhibition of the conditioned TS compared to TS alone at all intervals. Using the ratio of the conditioned stimulus over the test stimulus, mixed ANOVA (Group X ISI) revealed a significant main effect of ISI (F2, 23=9.80; p=0.005), no main effect of group (F2, 23=0.02; p=0.88), and no interaction (F2, 23=0.15; p=0.80). The main effect of ISI was explained by increased inhibition at the 100 msec interval (Fig. 1B).
Somatosensory evoked potentials
Between group comparisons for each SEP component are shown in Table 2. Both N20 and N60 components were found in each participant for electrode C3′. Student t tests revealed no significant difference between the two groups for N20 latency (t 19=1.77; p=0.10) and N20 amplitude (t 19=0.60; p=0.56). Student t tests also revealed no significant difference between the two groups for N60 latency (t 19=1.16; p=0.26) and N60 amplitude (t 19=0.66; p=0.53). A N13 component was found in every participant for electrode Cv7. Student t tests revealed no significant difference between groups for N13 amplitude (t 19=0.50; p=0.63) and N13 latency (t 19=0.04; p=0.97). One participant in the concussed group displayed a large N20 component that considerably increased variability. Removing this participant from analysis did not lead to a different between-group outcome and N20 amplitudes were below 3 SD.
Conduction time
Mean conduction times and standard deviations for both groups are shown in Table 3. For central sensory conduction time (CSCT), Student t test revealed no statistical difference between groups (t 17=1.19; p=0.25). Similarly, TMS assessment of central motor conduction time (CMCT) (t 23=0.26; p=0.80), spinal conduction time (t 23=0.53; p=0.58), and corticospinal conduction time (t 23=0.55; p=0.21) did not reveal any significant difference between groups.
Discussion
The major finding of the present study is the specificity of long-term intracortical inhibitory dysfunction in primary motor cortex of concussed athletes that is presumably GABAB mediated. Compared to unconcussed athletes, CSP and LICI measures were abnormal, suggesting increased GABAB inhibition. By contrast, GABAA-mediated inhibition (SICI), sensorimotor integration assessed with cholinergic-dependent SAI, and basic somatosensory processing (SEPs) were similar in the concussed and unconcussed groups. Finally, both afferent and efferent sensorimotor conduction times were of similar duration in the two groups.
Cholinergic abnormalities have been reported in patients with severe TBI associated with diffuse axonal injury, where short latency afferent inhibition was significantly reduced (Fujiki et al., 2006). Administration of a single dose of an acetylcholinesterase inhibitor was sufficient to restore normal SAI in this population, in line with previous studies showing similar effects in Alzheimer's disease (e.g., Di Lazzaro et al., 2002, 2004). The present findings suggest that cholinergic circuits in sensorimotor areas are unaffected by sports concussions two years after the last concussive event. The absence of significant differences between the two groups suggests that sensory signals originating from the median nerve interact normally with primary motor cortex circuits in the concussed brain. SAI appears to be partly mediated by GABAA receptors at the motor cortex level (Di Lazzaro et al., 2007). Data from the present and other (De Beaumont et al., 2007, 2009) studies have shown spared GABAA-mediated SICI in athletes tested 1–2 years after their last concussion. Although SAI and SICI appear to involve different subtypes of GABAA receptors (Di Lazzaro et al., 2007), it has been shown that they are reciprocally connected (Alle et al., 2009). Normal LAI was also found in the present population of formerly concussed athletes, and it is believed that LAI and SAI are mediated through different sensory-motor circuits. Although M1-S1 cortico-cortical interactions appear to underlie LAI, the exact nature of this inhibitory phenomenon is unclear (Pirio et al., 2009). Nevertheless, the present data point to the absence of long-term effects of sports concussions on the interaction between sensory input and corticospinal excitability. Whether this represents a state of recovery from short-term alterations is an open issue that warrants further investigation.
It is important to note that in contrast to previous studies from our group (De Beaumont et al., 2007b, 2009), MEPs were recorded from the APB rather than from the first dorsal interosseus muscle (FDI). It has been shown that excitatory and inhibitory patterns induced with TMS differ between muscle groups. For example, MEP area for muscles at rest is greater and onset latency longer for small hand muscles compared to forearm muscles, while CSP durations are longer for small hand muscles (Wu et al., 2002). Differential effects have also been reported between proximal and distal muscles, where increased intracortical inhibition and decreased intracortical facilitation are present in proximal compared to distal muscles (Abbruzzese et al., 1999). More relevant to the present study, it has been suggested that function may explain differences in inhibition between muscles, with intrinsic hand muscles playing an active role in fine motor acts requiring enhanced inhibitory control (Abbruzzese et al., 1999). As such, it is unlikely that the selection of one hand muscle over the other significantly affected the present data, but it is an open question whether similar group effects would be observed in proximal muscle groups.
The absence of sensorimotor integration dysfunction was matched by a lack of short/middle latency SEP abnormalities in concussed athletes. Concussions have been shown to increase middle-latency SEPs (N60) in the acute phase, an effect that tended to normalize to pre-injury levels three months after the concussive event (Zumsteg et al., 2006). The same study reported no significant latency differences in the early SEP components between concussed individuals and healthy controls (Zumsteg et al., 2006). The present data are in agreement with these findings, revealing no differences between concussed and control athletes on measures of SEP amplitude and latency. As such, if an increase in N60 latency in the acute and post-acute (3 months) phases was present in our group of concussed athletes, a post-injury period of more than one year is sufficient to restore basic somatosensory processing to normal levels. It should be noted, however, that participants in the study by Zumsteg and colleagues (2006) had lost consciousness following concussion, were older (35.4 years), spanned a much wider range of ages (22 to 62 years), and included patients with varied mTBI causes. This contrasts with the current sample, which was younger and much more homogeneous. As such, it is difficult to ascertain whether the lack of SEP abnormalities in the present sample of concussed athletes reflects recuperation from a dysfunctional state or merely the absence of SEP effects in the acute phase. This is an important issue in light of the fact that short-latency SEPs have been repeatedly shown to be good predictors of outcome in patients with severe TBI and are one of the best predictors of coma outcome (Carter and Butt, 2005). If SEP abnormalities are present acutely following sports concussions, it would be of great clinical interest to determine whether they can predict the severity of long-lasting impairments that have recently been discovered in the cognitive and motor domains.
In keeping with the lack of SEP abnormalities and sensorimotor integration dysfunction, the present data suggest that ascending and descending peripheral and central pathways are not affected by the presence of multiple sports concussions. In addition to normal SEP latencies, central somatosensory and motor conduction times were similar between groups. Numerous studies have used diffusion tensor imaging (DTI) to investigate the integrity of white matter fibers in mTBI (for review, see Maller et al., 2010). Of particular relevance to the present findings, reduced fractional anisotropy (FA) has been reported in the corticospinal tract and internal capsule of mTBI individuals (e.g., Bendlin et al., 2008). Indeed, reduced FA in the internal capsule of mTBI patients has been shown numerous times (Bendlin et al., 2008; Lipton et al., 2008; Miles et al., 2008) and has been observed up to 6 years post-injury (Inglese et al., 2005). Similarly, reduced white matter integrity of the corticospinal tract has been reported both in the short-term (2 months post-injury) (Bendlin et al., 2008) and long-term setting (107 months post-injury) (Kraus et al., 2007). Our physiological data show that if structural damage is present in white matter fibers of the corticospinal tract, it is not sufficient to modify ascending and descending conduction times. It should be noted that DTI studies of mTBI sometimes included patients with visible structural brain damage, prolonged unconsciousness, variable age groups, and presence of cognitive impairments, which contrasts with the homogeneous nature of the present sample. Indeed, every participant in the current study reported no overt symptom, was young and active, and had identical educational profiles. In this specific population, integrity of the corticospinal tract may be spared altogether.
The lack of significant somatosensory, sensorimotor, and conduction abnormalities found in the present study thus points to the specificity of long-term intracortical inhibitory dysfunction in concussed athletes. Previous findings of increased CSP duration in concussed athletes were replicated and additional evidence for the involvement of GABAB receptor activity was provided in the form of increased LICI. In healthy subjects, administration of selective GABAB agonist Baclofen increases LICI, possibly through facilitation of inhibitory post-synaptic potentials (McDonnell et al., 2006). As such, increased CSP and LICI measures suggest that both magnitude and duration of GABAB-mediated intracortical inhibition (McDonnell et al., 2006) is affected in sports concussion. It is important, however, to consider that TMS is an indirect measure of GABAergic activity. Evidence for an association between activity of specific receptor subtypes and TMS measures of intracortical inhibition comes primarily from pharmacological studies where GABAA or GABAB agonists or antagonists modulate M1 responses to TMS (Ziemann, 2004). Direct confirmation of the involvement of specific GABA receptors in the pathophysiology of sports concussions are needed to establish this fact with certainty.
A possible mechanism explaining presumed increased GABA transmission in concussion may be related to protective effects against glutamate excitotoxicity. Excessive glutamate stimulation is a core feature of brain response to TBI (Baker et al., 1993; Faden et al., 1989) leading to “glutamatergic excitotoxicity” and neurodegeneration (Rothman and Olney, 1986). Increased levels of glutamate are associated with NMDA receptors, and beneficial effects of NMDA receptor antagonists in TBI patients have been reported (e.g., Yurkewicz et al., 2005). In addition to modulation of glutamate levels, there is evidence for the involvement of GABA in the response to TBI, which could be secondary to the increase in glutamate and represent an attempt at minimizing glutamatergic excitotoxicity. Although the effects of TBI on GABA transmission in humans have been poorly studied, it has been reported that GABA concentration in ventricular CSF is greatly elevated in patients with severe brain injury (Palmer et al., 1994). There is significant data suggesting increased GABA levels following TBI in rat models of injury. For example, impact injury of rats has been shown to induce long-lasting working memory (WM) deficits that are associated with increased GABA levels for as long as 1 month post-TBI (Kobori and Dash, 2006). Reduction of GABA levels in those rats by administration of GABA antagonists restored memory function, suggesting that TBI is associated with “excess GABA-mediated inhibition” (Kobori and Dash, 2006). Magnetic resonance spectroscopy has shown a similar pattern of response following TBI in rats, where GABA concentration is elevated after injury (Pascual et al., 2007). The idea that long-term GABA increases in concusssed athletes is a response to glutamatergic excitotoxicity is obviously highly speculative. Indeed, it has recently been shown with MR spectroscopy that glutamate levels are decreased in the motor cortex of concussed athletes 3–4 days post-injury (Henry et al., 2010). However, since animal models have shown an immediate release of glutamate following mTBI (Katayama et al., 1990), complex interactions between excitatory and inhibitory neurotransmitters may significantly modulate the initial response in the days following concussive injury.
It is important to take into account the fact that the neurophysiological profile that is reported here comes from an homogeneously small sample made up of young, otherwise healthy high-level athletes. This has some important advantages, such as the presence of a comparable control group and reduction in the prevalence of co-morbid conditions. It should therefore be emphasized that the present results may not easily generalize to a broader population of individuals with TBI, where etiology, age, general health, and co-morbidity are important variables. For example, it remains to be seen whether individuals with more severe TBI also display abnormal M1 intracortical inhibition. With respect to concussed athletes, however, we can safely assume that our results are generalizeable despite the small sample size since similar dysfunction of inhibition in M1 have been found in older former athletes (DeBeaumont et al., 2009) and in another sample of young, active athletes (DeBeaumont et al., 2007).
In conclusion, the current data show that intracortical GABA dysfunction in the motor cortex of concussed athletes shows a surprising degree of specificity. The reported alteration appears to be long lasting and stable, as abnormal CSP durations have also been found more than 30 years after the last injury (De Beaumont et al., 2009). The specificity and duration of M1 dysfunctions make it plausible that TMS measures of cortical excitability may in the long run provide diagnostic and prognostic cues in TBI. In light of the fact that depression and post-traumatic stress disorder (PTSD) are often present in TBI patients (Kim et al., 2007; Rogers and Read, 2007), objective measures of M1 neurophysiology may find useful clinical utility. This is particularly relevant since both depression and PTSD have specific cortical excitability dysfunction profiles that are different from that of TBI. In PTSD, hemispheric-specific reductions in SICI and SAI (Rossi et al., 2009) have been reported, whereas depressive patients show a consistent pattern of right-left hemisphere motor threshold differences (Maeda et al., 2000) and reduced CSP durations (Bajbouj et al., 2006). Further studies will be necessary to determine the value of TMS measures in the differential diagnosis of these pathologies. Additionally, at present, return-to-play decisions following sports concussion are highly dependent on the athlete being physically and cognitively asymptomatic, as assessed by medical and neuropsychological evaluations (Reddy and Collins, 2009). The present data show that when these symptoms subside, highly specific dysfunctions in primary motor cortex may linger for years in some athletes. The development of objective, neurophysiological measures of brain dysfunction following concussion may therefore provide valuable return-to-play information in the future. However, whether the presence of intracortical inhibitory dysfunction can be a predictor of future concussive events, for example, is an open issue that will need to be addressed directly in future studies.
Footnotes
Acknowledgments
This work was supported by grants from the Canadian Institutes of Health Research and the Fonds de la Recherche en Santé du Québec.
Author Disclosure Statement
No competing financial interests exist.