
Abstract
Previously, we reported that every-other-day-fasting (EODF) in Sprague-Dawley rats initiated after cervical spinal cord injury (SCI) effectively promoted functional recovery, reduced lesion size, and enhanced sprouting of the corticospinal tract. More recently, we also showed improved behavioral recovery with EODF after a moderate thoracic contusion injury in rats. In order to make use of transgenic mouse models to study molecular mechanisms of EODF, we tested here whether this intermittent fasting regimen was also beneficial in mice after SCI. Starting after SCI, C57BL/6 mice were fed a standard rodent chow diet either with unrestricted access or feeding every other day. Over a 14-week post-injury period, we assessed hindlimb locomotor function with the Basso Mouse Scale (BMS) open-field test and horizontal ladder, and the spinal cords were evaluated histologically to measure white and grey matter sparing. EODF resulted in an overall caloric restriction of 20% compared to animals fed ad libitum (AL). The EODF-treated animals exhibited a ∼ 14% reduction in body weight compared to AL mice, and never recovered to their pre-operative body weight. In contrast to rats on an intermittent fasting regimen, mice exhibited no increase in blood ketone bodies by the end of the second, third, and fourth day of fasting. EODF had no beneficial effect on tissue sparing and failed to improve behavioral recovery of hindlimb function. Hence this observation stands in stark contrast to our earlier observations in Sprague-Dawley rats. This is likely due to the difference in the metabolic response to intermittent fasting as evidenced by different ketone levels during the first week of the EODF regimen.
Introduction
A
Dietary restriction (DR) regimens like every-other-day-fasting (EODF) have been demonstrated to induce numerous positive effects on animal health and efficacy in various neurological disease models (Anson et al., 2003; Bruce-Keller et al., 1999; Duan and Mattson, 1999; Maswood et al., 2004; Rich et al., 2010; Yu and Mattson, 1999). Recently we reported that EODF initiated after injury improved outcomes from SCI in Sprague-Dawley rats (Plunet, 2010; Plunet et al., 2008, 2011). Rats maintained on an EODF regimen after an incomplete cervical SCI had reduced lesion cavities, more detectable neurons in the vicinity of the injury, and enhanced sprouting of corticospinal axons (Plunet et al., 2008). Importantly, intermittently-fasted rats demonstrated improved functional outcomes, including increased use of their injured forelimb during vertical exploration, more accurate limb placement, and normalization of their walking pattern (Plunet et al., 2008). More recently, we found that this therapeutic regimen of EODF also effectively promoted functional recovery following contusion injury at the thoracic level (Plunet et al., 2011).
Intermittent fasting, in contrast to mere caloric restriction, forces the body to use fat as fuel, thereby increasing circulating ketone bodies like acetone, acetoacetic acid, and β-hydroxybutyrate. These ketone bodies are transported from the liver to other tissues, including the central nervous system, where they are reconverted to acetyl-CoA to produce energy in the form of ATP. Although the molecular mechanisms underlying the beneficial effects of EODF in the context of SCI are poorly understood, a large body of literature emphasizes the beneficial effects of ketones as a clean metabolic energy source and mediators of neuroprotection (Imamura et al., 2006; Kashiwaya et al., 2000; Kweon et al., 2004; Maalouf et al., 2007; Massieu et al., 2003; Mejia-Toiber et al., 2006; Noh et al., 2006; Tieu et al., 2003). In addition, it has been proposed that DR could act through regulation of gene expression, including increased neurotrophic factor levels and heat-shock proteins, as well as through antioxidative, anti-apoptotic, and anti-inflammatory mechanisms (Anson et al., 2003; Dubey et al., 1996; Maalouf et al., 2008; Shelke and Leeuwenburgh, 2003; Sharma and Kaur, 2005; Sohal et al., 1994; Yu and Mattson, 1999). Moreover, DR improves metabolic efficiency and enhances antioxidant defenses (Agarwal et al., 2005; Gong et al., 1997; Rankin et al., 2006; Sreekumar et al., 2002).
In order to exploit transgenic mouse models to study the mechanisms of EODF, the current study was designed to evaluate the efficacy of EODF in spinal cord-injured C57BL/6 mice. Given the effectiveness of a DR regimen in a wide range of mammals, including, mice, rats, dogs, cows, and primates, we expected to see similar effects in mice. However, the results demonstrate that EODF in spinal cord-injured mice has no beneficial effects on functional recovery, which stands in contrast to our initial rat study. It is suggested that this discrepancy is likely due to a different metabolic response to EODF in C57BL/6 mice after injury, as evidenced by attenuation of ketosis following fasting.
Methods
All procedures involving animals were approved by the Animal Care Committee of the University of British Columbia (UBC), in accordance with the guidelines of the Canadian Council for Animal Care and the National Institutes of Health. All efforts possible were made to reduce the number of animals used and to minimize pain.
Animals and general housing conditions
Adult male C57BL/6 mice (20–23 g; n=29) were purchased from Charles River Laboratories and housed in the UBC central animal facility with room temperature controlled at 21°C, and an artificial 12-h:12-h light:dark cycle (lights on at 06:00
Surgical procedure and dietary regimen
All surgeries were performed under general anesthesia using isoflurane gas, and body temperature was monitored throughout surgery and maintained at 36.5°C using a heating plate controlled via a rectal thermometer (Physitemp Instruments, Inc., Clifton, NJ). To alleviate pain, buprenorphine (0.03 mg/kg SC, Temgesic®; Schering-Plough Corporation, Kenilworth, NJ) and lidocaine (20 mg/mL; Bimeda-MTC Animal Health Inc., Cambridge, Ontario, Canada) containing 2% epinephrine (IM in the surgical area) were administered prior to surgery. Postoperatively, the animals were injected with 2 mL of lactated Ringer's solution (Hospira Inc., Montreal, Quebec, Canada) to prevent dehydration. The thoracic crush injury was performed as described previously with some modifications (Plemel et al., 2008). Innervation of the liver and adipose tissue is mainly derived from postganglionic branches of the greater splanchnic nerve, which originate from spinal segments T5–T10 (Buijs et al., 2003; Kreier et al., 2002; Kuo and de Groat, 1985). Because a high thoracic bilateral SCI might interfere with proper liver function, we chose to perform the injury at the T10–T11 level. Briefly, the dorsal aspect of the spinal column was exposed, and laminectomy of vertebrae T10–T11 was performed using a pair of fine rongeurs. A modified Dumont-type forceps with a 0.35-mm spacer was used to laterally compress the spinal cord for 10 sec (Fig. 1). The skin incision was closed with sutures.

Schematic representation of the thoracic spinal cord crush model used. Lateral compression was applied to the spinal cord for 10 sec using a modified Dumont-type forceps with a 0.35-mm spacer.
When the mice regained their ability to move around freely, they were placed back in a fresh recovery cage, and randomized into the control group (n=14), which continued to have access to food AL (28.507% calories from protein, 13.496% from fat, and 57.996% from carbohydrates), or the EODF group (n=15), which received no food on every other day beginning with the first 24-h food deprivation immediately after surgery. This schedule of alternate fasting and feeding days was carried out throughout the remainder of the study. In a pilot study we observed maximum ketone levels sooner when food was taken away at the onset of the dark phase compared to the light phase, and therefore decided to initiate the fasting period 2 h before their active dark period (4:00
In compliance with animal welfare regulations and following instructions from the veterinary staff at our facility, by post-operative day 10 and thereafter both groups received a diet containing 11% fat to overcome dramatic weight loss (19.805% calories from protein, 25.337% from fat, and 54.858% from carbohydrates; LabDiet-5015; Purina Mills). Additionally, the mice were given daily SC injections of lactated Ringer's solution (1 mL; Hospira) for 14 days post-operatively to prevent dehydration. To alleviate post-operative pain, buprenorphine (0.03 mg/kg) was SC injected twice daily for an additional 2 days. The bladders were manually expressed twice daily until the mice regained voluntary micturition.
Feeding and body weight monitoring
Daily food intake and body weights were measured in the mice from both groups for 7 days prior to and up to 5 weeks after surgery. Group-housed mice were provided with a pre-weighed amount of pellets in their food hopper and home cage. Leftover food weight was recorded daily (∼ 03:00
Assessment of functional recovery from spinal cord injury
Hindlimb motor performance was evaluated weekly using two behavioral tests, including the Basso Mouse Scale (BMS) open-field assessment of motor function, and the horizontal ladder. Ladder and catwalk analysis was only done for mice that had reached frequent weight-supported stepping in the open field (BMS score 5).
BMS open-field behavioral assessment
Hindlimb locomotion was scored by the open-field BMS, a rating scale developed specifically for mice (Basso et al., 2006). The behavioral testing was performed on feeding days to allow for the behavioral observers to remain blinded to the treatment groups. The BMS is a 9-point scale (0=complete hindlimb paralysis to 9=normal locomotion), in which mice are scored based on hindlimb movements made in an open field during a 4-min interval while freely exploring the surroundings. Individual hindlimb scores were averaged for each animal at each time point. Once animals showed frequent hindlimb stepping, BMS subscores were collected to reveal more detailed characteristics about locomotor control. The animals were tested prior to injury and once weekly for 14 weeks post-surgery.
Horizontal ladder walking test
We evaluated hindlimb stepping ability while crossing a horizontal ladder with regularly-spaced rungs (Cummings et al., 2007; Metz and Whishaw, 2002). The animals were trained to cross the ladder towards their home cage positioned at the opposite end of the ladder. No further reinforcement was given to motivate the animals to cross the ladder. The horizontal ladder apparatus consists of clear acrylic glass walls and metal rungs elevated 20 cm above the ground. All of the animals had to cross a ladder with equal spacing (1.3 cm) between the rungs. A high-definition camera was positioned to record the animals' performance as they crossed the ladder. Subsequent frame-by-frame video analysis provided scores averaged over three trials per session for the total number of steps, and missed steps were referred to as errors (Cummings et al., 2007). The error rate in stepping of the affected hindlimbs was recorded as a percentage of the number of missed steps divided by the total number of steps.
Measurement of glucose and β-hydroxybutyrate
Before and after spinal cord injury, glucose and β-hydroxybutyrate levels in the blood were determined for all animals using MediSense Precision Xtra blood test strips (Abbott Laboratories, Alameda, CA). Blood samples were collected from animals fed AL, fed EODF, and fasted EODF mice (24 h) at the end of the day (∼ 5:00
Spinal cord tissue processing
All mice were euthanized 15 weeks post-injury. Following intracardiac perfusion with 0.1 M phosphate-buffered saline (PBS; 15 mL) and ice-cold 4% paraformaldehyde in PBS (30 mL), the spinal cord was post-fixed overnight and cryoprotected in graded concentrations of sucrose (12%, 18%, and 24%). The spinal cords were cut into three 4-mm segments (the crush site, and 1 segment rostral and caudal to the crush site) and frozen on dry ice. Coronal serial sections 20 μm thick were cut on a cryostat and mounted onto microscope slides. Sections were stained with eriochrome cyanine or immunostained with an antibody to glial fibrillary acidic protein (GFAP) as described below.
GFAP immunohistology
To assess the lesion area and volume after spinal cord injury, coronal sections 120 μm apart were stained for GFAP. Blocking of non-specific proteins was accomplished by incubating the slides in 10% normal donkey serum (Jackson ImmunoResearch Laboratories, West Grove, PA) in 0.1% PBS-Triton-X 100 for 30 min. The sections were then incubated overnight at room temperature in rabbit polyclonal anti-GFAP antibody (DakoCytomation, Carpinteria, CA) diluted at 1:1000 in 0.1% PBS-Triton-X 100. After washing, the slides were incubated in Cy3-conjugated donkey anti-rabbit (1:1000; Jackson ImmunoResearch Laboratories) for 60 min. Finally the slides were rinsed several times with PBS and cover-slipped with Fluoromount-G (Southern Biotechnology Associates, Birmingham, AL).
Eriochrome cyanine staining
To analyze spared white and grey matter, coronal sections through the thoracic injury site were stained with eriochrome cyanine (EC) to visualize myelinated white matter. Frozen spinal cord sections were rehydrated through graded ethanol solutions (twice in toluene, twice in 100%, once in 95%, and then once in 70% and 50% ethanol). After rehydration in dH2O the slides were stained in EC solution (0.16% EC, 0.4% sulfuric acid, and 0.4% ferric chloride) at room temperature for 10 min, and gently rinsed in dH2O. The slides were differentiated in 0.5% ferric ammonium sulfate at room temperature for 2 min, rinsed in dH2O, dehydrated in graded ethanol solutions (once in 50%, 70%, and 95% ethanol, and two changes in 100% ethanol), cleared in xylene two times, and cover-slipped using mounting medium.
Quantification of tissue sparing and lesion volume
Each slice was digitally photographed using a Zeiss AxioPlan2 imaging microscope (Zeiss Inc., Thornwood, NY) equipped with a Q-Imaging camera (Q-Imaging, Burnaby, B.C., Canada). Tissue sparing and lesion area were manually traced using imaging analysis software (Sigma Scan Pro5; Systat Software Inc., San Jose, CA). The amount of total white tissue sparing was based on positive staining for myelin visualized with eriochrome cyanine staining. Spared grey matter was defined as tissue containing normal gray matter cytoarchitecture based on eriochrome cyanine. The lesion area (μm2) was defined as a central GFAP-negative area of the spinal cord. Total lesion volume (μm3) was calculated by the Cavalieri volume estimation formula ([lesion area * distance between sections] – [0.02 * lesion area epicenter]).
Statistical analysis
All data are presented as mean±standard error of the mean (SEM). The behavioral data and blood ketone and glucose levels were analyzed using an analysis of variance (ANOVA) repeated measures factorial model containing between-subjects variables, and a within-subjects variable (interaction group across time), which allows each mouse to serve as its own control. Pair-wise comparison by week (Student's t-test) was conducted after significant main effects of ANOVA repeated measures between- or within-subject effects. To compare different time points within the same group paired samples t-tests were used. The significance level for all tests was set at p<0.05.
Results
Post-injury mortality was limited to two animals in the first 2 weeks after injury, one from each feeding group. During the initial days after SCI, autonomous bladder voiding was absent, and the bladders had to be expressed manually two times a day. Both AL and EODF animals regained reflex bladder emptying and no longer required manual bladder expression within 5–7 days after injury.
Body weight and food intake
The average food intake and body weight for AL and EODF animals was similar before surgery (Fig. 2). During 5 weeks following spinal cord compression on the days EODF mice had access to food, they ate significantly more than mice fed AL (p≤0.001; Fig. 2C). Caloric intake of EODF mice was significantly increased by ∼60% at weeks 1, 4, and 5 following injury compared to the AL group (p=0.041, p=0.001, and p=0.008, respectively). Thus, even though mice maintained on an alternate day fasting regimen compensated for periods of fasting, their overall food intake was decreased (∼20% calorie restricted).
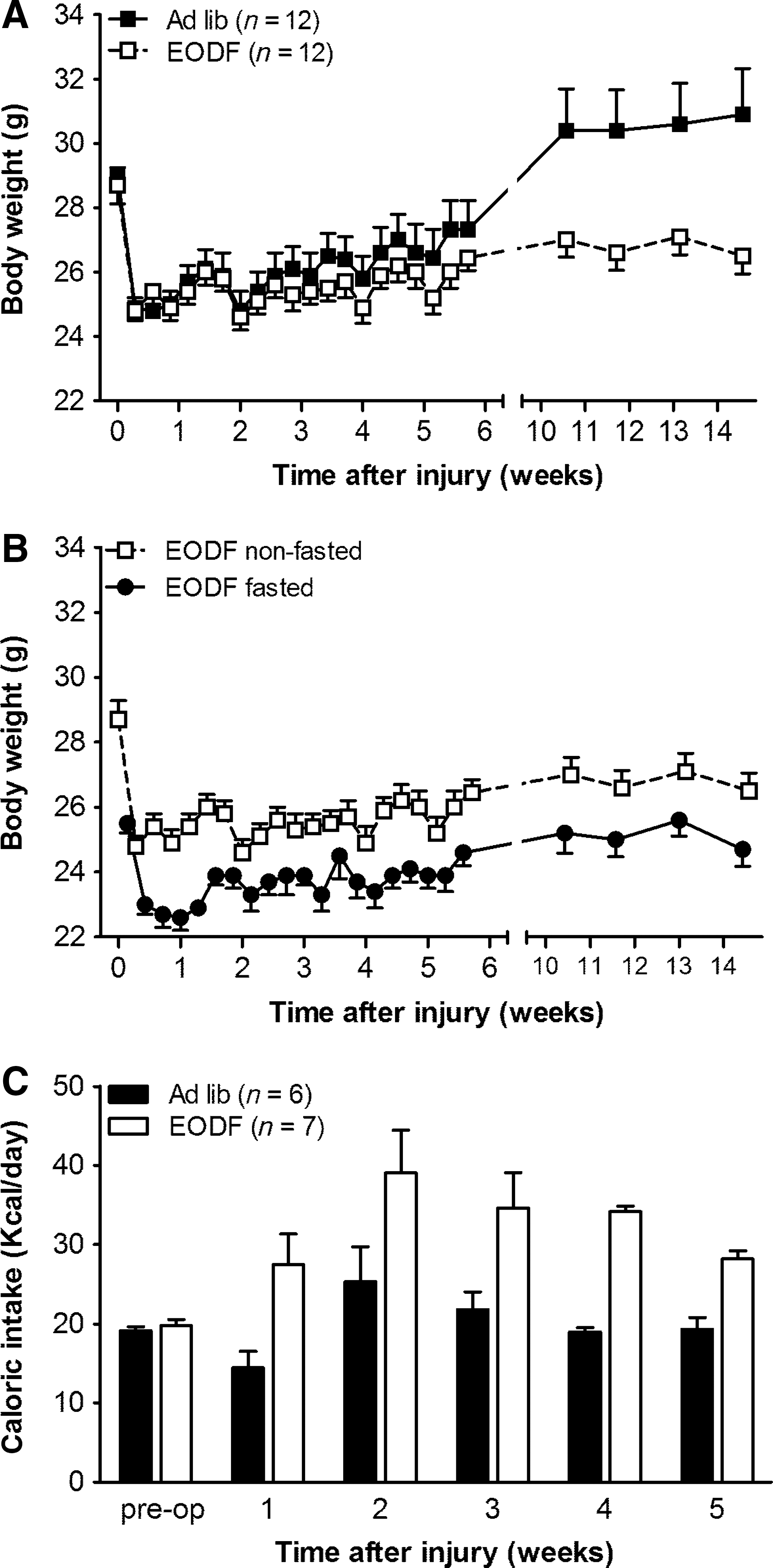
Body weight and caloric intake in every-other-day-fasting (EODF) and ad libitum (AL) mice after thoracic spinal cord injury. (
Independent of dietary regimen, mice showed a body weight drop of ∼4.0 g the first day after injury (Fig. 2A). Over time, the body weight of AL mice gradually increased and returned to above pre-injury levels within 6–10 weeks after crush injury (Fig. 2A).
Although repeated measures ANOVA indicated no significant difference between groups during the 14-week time period (p=0.268), the recovery of body weight was significantly different between AL and EODF mice (p=0.001). Group comparison by week indicated that by 10 weeks after injury until the end of the study, the body weight of EODF mice was significantly reduced compared to AL-fed animals (p≤0.03). At 14 weeks post-injury their body weight still did not reach pre-injury levels and remained ∼14.2% below the values of AL-fed animals (AL: 30.9±1.43 g; EODF: 26.5±0.55 g; p=0.012; Fig. 2A).
Throughout the experiment, a 24-h fast induced an additional ∼1.9 g reduction in body weight compared to eating days (Fig. 2A and B), which resulted in a total body weight drop of ∼20% in EODF mice during the early days after injury compared to AL-fed animals.
Blood ketone and glucose response
Blood β-hydroxybutyrate (β-HB) and glucose levels were measured to investigate the metabolic effect in response to EODF in mice and rats (Fig. 3). In contrast to rats, mice with a thoracic SCI showed a significantly different ketone response upon fasting following 6 weeks of an EODF regimen (p=0.001). Group comparisons per week revealed that at the end of the second, third, and fourth fasting day (days 3, 5, and 7 after injury), ketone levels of mice were significantly reduced compared to rats fasted for 24 h (p=0.001, p=0.0001, and p=0.0001, respectively), and barely differed from non-fasting levels (p=0.858 by paired t-test on day 3, and p=0.138 on day 5; Fig. 3A). Interestingly, this lack of ketosis observed in mice was associated with a substantial drop in blood glucose levels (Fig. 3B). At that time point EODF mice were generally inactive and lethargic. On the following days, mice showed an improvement in their general appearance combined with a recovery of glucose levels and a gradual increase in their ketone response to 24-h fasting, reaching consistently elevated ketone levels as of the seventh fasting day (day 13 after injury).

β-hydroxybutyrate (
Locomotor recovery
Next, we evaluated whether EODF treatment would promote functional recovery after SCI in these mice by scoring their open-field locomotion using the Basso Mouse Scale (BMS; Basso et al., 2006). Figure 4 summarizes the BMS hindlimb motor scores and subscores for AL and EODF mice. Before surgery, a maximal BMS score of 9 was reached by all the mice. For 2 to 4 days after injury, EODF and AL mice were no different in their degree of hindlimb ankle movements, which were slight to extensive (BMS score 0–2; Fig. 4A). BMS open-field scores improved over several days, reaching a stable plateau at 3 weeks post-injury. For the remainder of the experiment, both AL and EODF mice showed frequent to consistent weight-supported hindlimb stepping, with no or some coordination (score of 5). By 14 weeks, a total of seven mice (AL: 2 out of 12; EODF: 5 out of 12) were not capable of frequent or consistent plantar stepping (BMS score ≤4). Statistical analysis of BMS scores indicates that all mice showed some improvement over time (p≤0.001), which was independent of the type of dietary treatment used (p=0.984). During the 14-week time period, there was no group difference between EODF or AL mice in the BMS open-field test (p=0.892), indicating that both groups performed equally well.
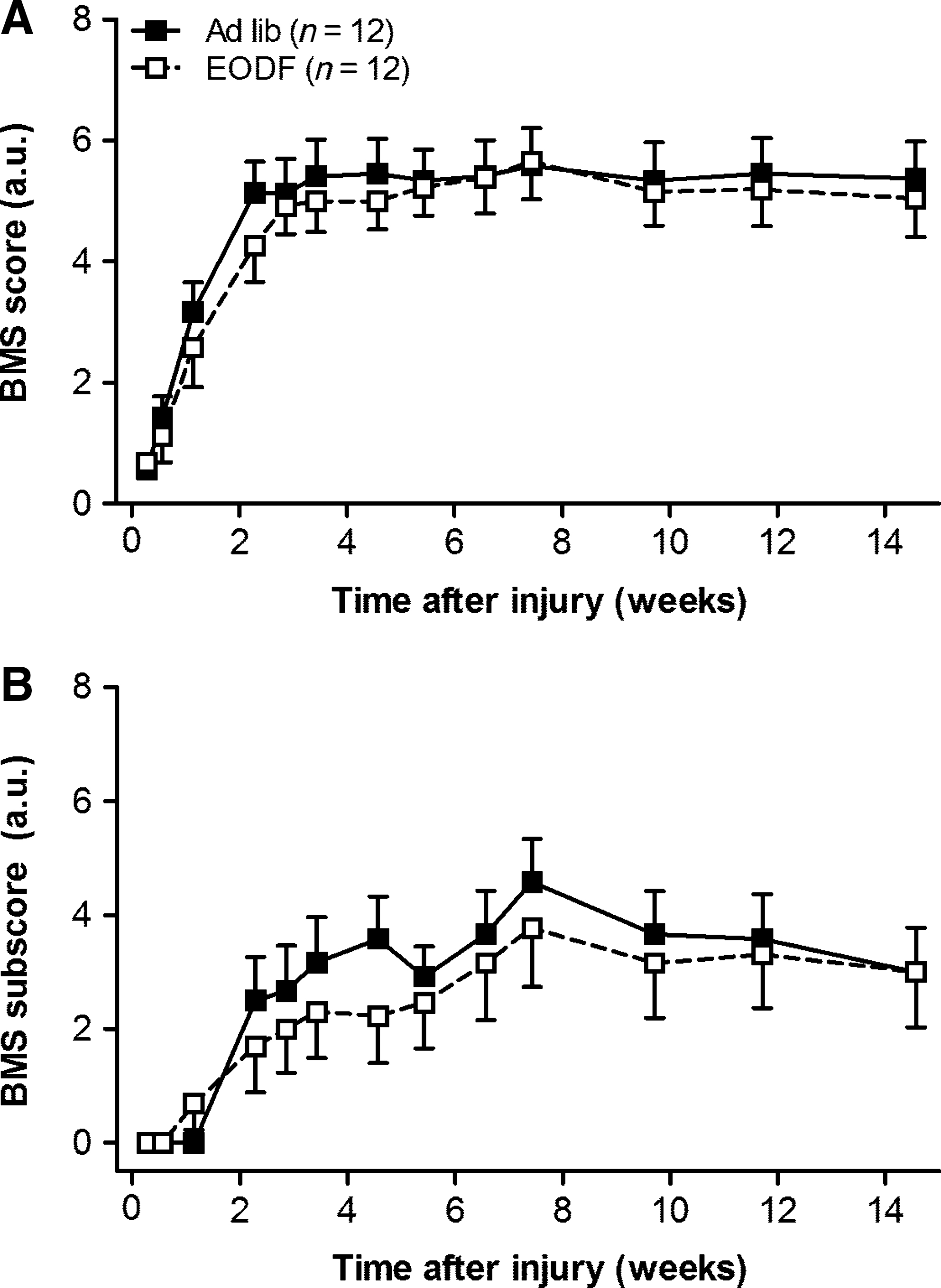
Every-other-day-fasting (EODF) does not improve locomotor recovery in mice as assessed with the Basso Mouse Scale (BMS;
Transient improvement over time within animals was also seen for the BMS subscore (p≤0.001), which was similar for the AL and EODF groups (p=0.955). No difference between groups was observed during the 14-week time period (p=0.770).
To further characterize the recovery of locomotion in our spinal cord-injured mice, we counted the number of times their hindpaws fell between the rungs while the mice were crossing a horizontal ladder (i.e., errors; Fig. 5B). Before injury, both the EODF and AL mice performed the ladder task with only a few hindlimb errors. After SCI the number of hindlimb steps and percentage of errors increased over time for all mice (p≤0.001; Fig. 5). Although not statistically significant, the improvement over time in EODF mice tended to be lower compared to AL animals (p=0.084). This was probably due to more EODF animals dragging on the ladder compared to AL mice (BMS score ≤4; AL: 2 out of 12 animals; EODF: 5 out of 12 animals).
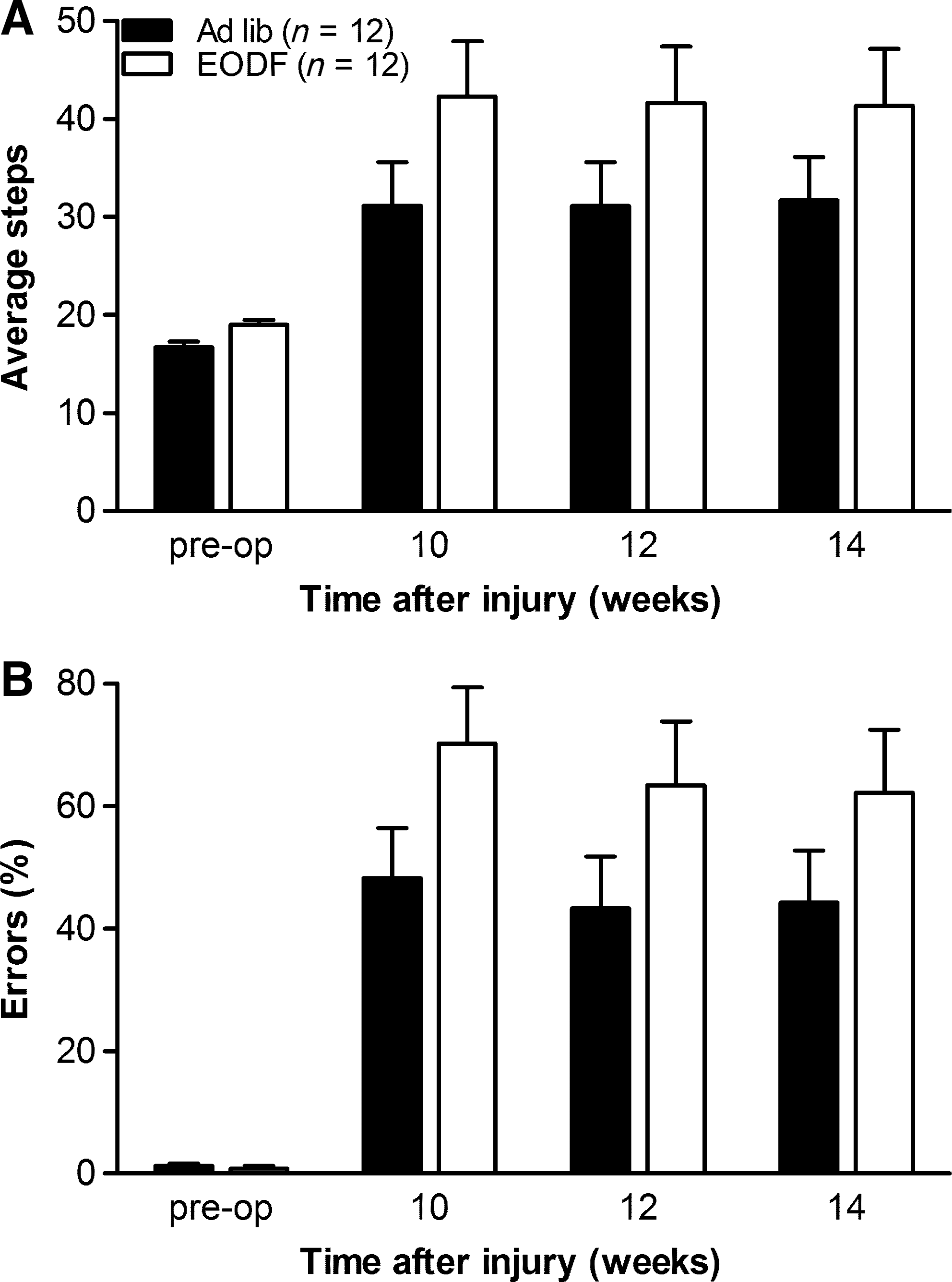
Every-other-day-fasting (EODF) in mice does not improve horizontal ladder performance after thoracic spinal cord injury. (
No significant overall group difference in ladder performance between the AL and EODF mice was observed during the 14-week time period (Fig. 5; p=0.473).
Spared tissue and lesion volume
In our previous study we found significant sparing of the injured cervical spinal cord in rats receiving EODF (Plunet et al., 2008). GFAP staining was used to outline the lesion edge in the injured spinal cords of our mice. At the epicenter, the lesion area encompassed ∼ 45% of the spinal cord for both groups (Fig. 6A), and extended several micrometers (480 μm) in both the rostral and caudal directions from the epicenter (Fig. 6B). Throughout this region, there was no difference in GFAP-negative lesion area (p=0.955; Fig. 6B), or total estimated lesion volume (p=0.923; Fig. 6C) between the AL and EODF groups. Eriochrome cyanine staining was used to visualize myelin preservation and revealed that in both groups the damage at the epicenter included most of the gray matter, and spared a rim of white matter (Fig. 7A–C). No significant differences for gray matter sparing (p=0.459) or white matter sparing (p=0.986) were observed between the treatment groups.

Every-other-day-fasting (EODF) treatment did not reduce the lesion size after thoracic spinal cord injury. (
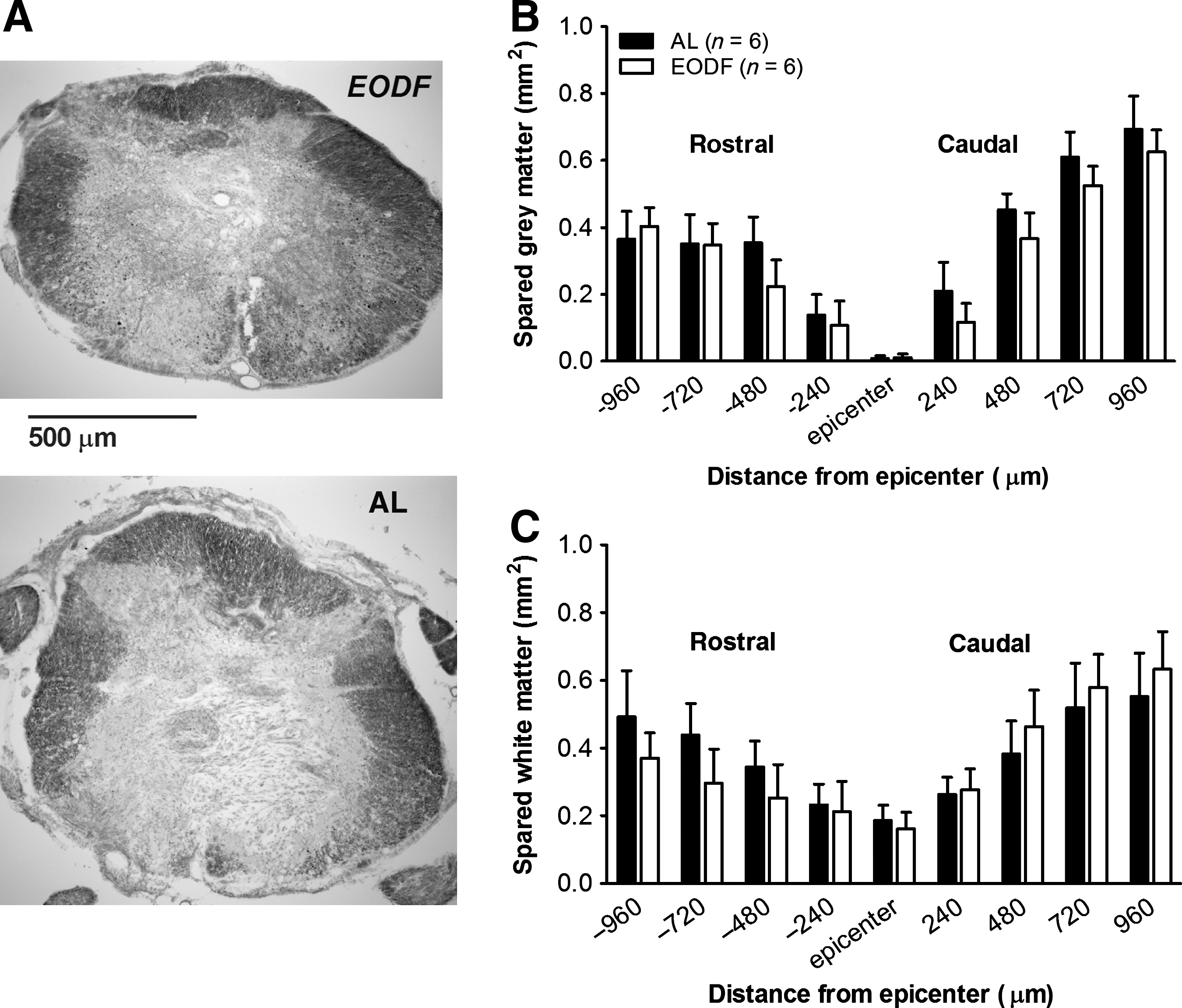
Every-other-day-fasting (EODF) treatment did not affect total spared tissue after spinal cord injury. (
Discussion
The present study demonstrates that C57BL/6 mice fasted every other day did not show improved recovery of locomotor function after thoracic crush injury of the spinal cord. No differences in BMS locomotor scores or hindlimb function during ladder crossing were observed. Furthermore, intermittent fasting in C57BL/6 mice did not have a neuroprotective effect on tissue sparing or lesion size. These findings are unexpected, since we have recently found in Sprague-Dawley rats that intermittent fasting initiated after cervical and thoracic SCI resulted in preserved neuronal survival, reduced lesion volume, increased sprouting, and improved functional recovery (Plunet, 2010; Plunet et al., 2008, 2011).
Multiple studies have shown beneficial effects of dietary restriction in neurodegenerative disorders in different mammalian species including C57BL/6 mice, as used in our study, and transgenic mice on the C57BL/6 background. Dietary-restricted mice exhibit extended longevity (Forster et al., 2003) and improved functional outcome in a mouse model of Alzheimer's disease (Halagappa et al., 2007). Additionally, mice maintained on an intermittent fasting regimen showed increased neuronal resistance to brain injury (Anson et al., 2003). It is important to note that all of these studies initiated dietary treatment before injury, or the onset of age- and disease-related pathologies, and the adaptation to a new diet and hunger did not coincide with the stress of sudden trauma, as seen with our study.
Our observed behavioral deficit in C57BL/6 mice is in agreement with the forceps crush model described by Plemel and colleagues (2008). Using a spacer of 0.35 mm resulted in consistent weight-supported hindlimb stepping with occasional or frequent coordination, which is similar to the behavioral deficit observed after thoracic contusion injury in Sprague-Dawley rats (Plunet, 2011), yet EODF was effective in the rat model. However, the loss of intact tissue that occurred in mice was less severe than that seen in our rat model. Our mice were left with 45% of spared tissue (mostly white matter), whereas after spinal cord contusion of rats, only 15% of tissue sparing was found at the epicenter (Mann et al., 2010; Plunet, 2011). It has been suggested that the neuroprotective effect of fasting correlates negatively with injury severity (i.e., EODF is more beneficial in lighter injuries) (Davis et al., 2008). If this correlation applies to SCI in mice, our model should have revealed a beneficial effect of dietary restriction in mice as well.
A possible explanation for the discrepancy in the response to EODF in Sprague-Dawley rats versus C57BL/6 mice is a difference in whole-body adaptive responses to fasting. In both studies the EODF animals were deprived of food immediately following SCI (i.e., the regimen started with a fasting day). Mice may be more sensitive to deprivation of nutrient supply because their higher metabolic rate per unit body weight leads to faster depletion of their nutrient stores during fasting (Brody, 1945). It has been shown in 129/SV and FVB mice that genes involved in lipid metabolism and ketone-body synthesis most significantly responded after 24–48 h of prolonged fasting (Bauer et al., 2004; Sokolovic et al., 2008), while in male Sprague-Dawley rats this response peaked between 3 and 5 days of fasting (Li et al., 2006). This demonstrates that mice and rats could have a different whole-body adaptive response to fasting, in particular the genes involved in ketone synthesis.
The same principle may apply to the C57BL/6 mouse strain used in our study. Despite an overall similar caloric restriction, EODF in C57BL/6 mice resulted in a 20% weight loss during the first week, compared to only a ∼7% reduction in thoracic spinal-cord-injured Sprague-Dawley rats (Plunet et al., 2011). While several studies using EODF in mice did not report significant weight loss, Anson and colleagues (2003) observed a 14% reduction in body weight in uninjured C57BL/6 mice after 14 weeks of EODF. In the same article they showed that these mice consumed around 1.7–2 times more on the days they had access to food as mice fed AL (Anson et al., 2003). In our study we observed that spinal cord injured mice only partially compensate for periods of fasting by increasing their food intake roughly 1.6 times; however, this declined further over the time period of the experiment. Although we did not include a non-injured EODF group to determine the contribution of EODF versus injury to the pronounced reduction in weight gain in EODF mice with SCI, our data indicate that both EODF as well as SCI contributed to this dramatically reduced weight.
Additionally, we observed a drop in circulating glucose levels during the first 2 fasting days after injury, while our rats showed an increase in serum glucose response levels. This suggests that the C57BL/6 mice depleted their energy stores more rapidly, in contrast to Sprague-Dawley rats. Additionally, EODF-treated rats recovered faster from a transient weight loss after SCI, and regained their original weight within 2 weeks after surgery, and slowly increased body weight thereafter. In contrast, C57BL/6 mice on the intermittent fasting regimen never returned to their pre-injury body weight during the 14 weeks of our experiment, indicating global differences in metabolism between mice and rats when on an EODF regimen. Evidence from Sohal and colleagues (2009) suggests that the beneficial effect of dietary restriction is dependent on a positive imbalance between energy expenditure and energy intake. In our study, EODF-treated Sprague-Dawley rats seem to have a positive energy balance, as indicated by a gain in weight after injury, whereas the energy balance remains relatively neutral if not negative in C57BL/6 mice, since they did not regain body weight after SCI when intermittently fasted. Hence a plausible explanation for the discrepancy between our mice and rats might be a difference in energy balance under the EODF regimen.
The maintenance of the fuel supply during fasting requires an extensive exchange of metabolites between organs. This exchange mainly occurs as glucose, lactate, amino acids, triglycerides, and ketone bodies. A prominent physiological switch occurs in mammals after 18–24 h of fasting, shifting energy supply from glucose to free fatty acids (Sokolovic et al., 2008). Under normal conditions, glucose is the primary energy source of the central nervous system; however, during prolonged fasting ketone bodies are a vital energy source obtained from breakdown of fatty acids in the liver (Maalouf et al., 2007). Ketone bodies can therefore provide up to 60% of energy to the brain when glucose availability is limited (Owen et al., 1976). Recent findings suggest that this shift to ketogenesis could play a direct role in the therapeutic efficacy of EODF, since the exogenous administration of specific ketones has also proven effective in several disease models. β-HB itself can protect neurons in in vivo and in vitro models of Alzheimer's and Parkinson's diseases and glutamate toxicity (Imamura et al., 2006; Kashiwaya et al., 2000; Kweon et al., 2004; Maalouf et al., 2007; Massieu et al., 2003; Mejia-Toiber et al., 2006; Noh et al., 2006; Tieu et al., 2003). In addition, exogenous β-HB administration was neuroprotective when administered after traumatic brain injury in rats (Davis et al., 2008; Prins et al., 2005). Similarly, intravenous infusions of β-HB administered 1 h after transient occlusion of the middle cerebral artery significantly decreased infarct size (Suzuki et al., 2002). Thus the differences between Sprague-Dawley rats and C57BL/6 mice could additionally be due to differences in metabolic adaptation to fasting, especially the differences in the fasting-induced ketone response.
Interestingly, our mice on the EODF regimen showed a ketone response that peaked on the first day of fasting, but surprisingly not on the next 3 fasting days. The ketone response to fasting recovered only gradually on the later fasting days, and did not reach typical levels (∼ 0.6 mmol/L) until 2 weeks after injury and the onset of the EODF regimen. Interestingly, this phenomenon was not observed for Sprague-Dawley rats on EODF. One limitation is that we cannot rule out whether the differences observed in our rats versus mice are due to differences in lesion levels. One would expect that in our mice that were lesioned at the T10–T11 level more sympathetic outflow to the visceral organs was left intact than in the T8-injured rats. Ketogenesis is reduced upon stimulation of α-sympathetic innervation of the liver, although β-hydroxybutyrate itself was not affected in the study of Beuers and colleagues (1986). It is conceivable that relatively more sympathetic outflow in the mice could have contributed to a lesser ketogenic response upon fasting than in rats.
Evidence from DR studies suggests that the relationship between fasting and enhanced cellular stress resistance and repair of damaged cells could be essentially bell-shaped (Bishop and Guarente, 2007; Clancy et al., 2002; Mair and Dillin, 2008; Panowski et al., 2007). Moderate fasting generally protects against a variety of cellular stressors; however, extreme nutrient stress resulting from excessive fasting may actually increase the risk of tissue damage and associated pathologies. Thus dietary restriction may not be beneficial under all circumstances, and an optimal dose range seems to exist. If ketogenesis is one of the contributing mechanisms of EODF, we should be able to see protective effects with EODF pretreatment, since the serum ketone levels will be upregulated at the time of injury. Although the significance of such pretreatment may not be clinically relevant in the setting of SCI, it will facilitate the identification of possible new targets for drug treatment through the use of loss-of-function approaches.
Given the beneficial effects of ketones discussed above it is conceivable that a high-fat, low-carbohydrate diet like the ketogenic diet (KD), which was originally designed to mimic the physiological effects of fasting, may successfully be used in the treatment of SCI. In a pilot study in our lab, we observed that KD in spinal cord-injured C57BL/6 mice increased their serum ketone levels to ∼2 mmol/L immediately after injury. Several reports have indicated a positive correlation between the level of ketosis and KD efficacy (Bough and Eagles, 1999; Huttenlocher, 1976). Hence, if ketones are one of the main mechanisms behind the neuroprotective effects of a KD, the overall effects could be even more pronounced with KD than EODF.
In summary, the present study shows that intermittent fasting in C57BL/6 mice following a moderate to severe thoracic SCI does not improve recovery, which is in contrast to what we observed in injured Sprague-Dawley rats (Plunet et al., 2008, 2011). We suggest that the differential metabolic compensatory response to EODF in spinal cord-injured mice, which includes an attenuated ketone response to fasting, might contribute to this discrepancy. However, a noteworthy difference between our mouse study and the previous rat studies is the switch from the standard rodent to a diet that differed in the relative abundance of fat and protein to overcome the dramatic weight loss in mice. Although it has been suggested that EODF regulates a number of anti-apoptotic and anti-inflammatory mechanisms within hours to days (Anson et al., 2003; Dubey et al., 1996; Maalouf et al., 2008; Sharma and Kaur, 2005; Shelke and Leeuwenburgh, 2003; Sohal et al., 1994; Yu and Mattson, 1999), we cannot exclude the possibility that the difference in diet initiated at 10 days post-SCI might additionally have contributed to the discrepant finding between our rat and mouse study, and more research is necessary.
Footnotes
Acknowledgments
We would like to thank Tony Lim and Susan Sim for helping with the lesion size and spared tissue analysis, Claire Miles, Maxwell Ma, and Alfonso Chen for their help with the behavioral analysis, and our colleagues in the Central Animal Facility for help and advice with animal care.
This research was supported by The Christopher Dana Reeve Foundation (F.S.), and the Canadian Institutes for Health Research (W.T). W.T. holds the Edie Ehlers Chair in Spinal Cord Research funded by the Rick Hansen Foundation.
Author Disclosure Statement
No competing financial interests exist.