
Abstract
In contrast to mammals, lampreys are capable of recovering apparently normal locomotion after complete spinal cord transection, and the spinal axons regenerate selectively in their correct paths. Descending serotonergic projections to the spinal cord play a role in the modulation of locomotion at spinal levels in both mammals and lampreys. In this study, we used combined immunofluorescence and tract-tracing techniques to show that in the sea lamprey, serotonergic descending neurons of the caudal rhombencephalon (vagal nucleus) regenerate their axons across the lesion site after complete spinal cord transection. The spinal cord of mature larval sea lampreys was transected at the level of the fifth gill, then after a recovery period of 5 months, the spinal cord was exposed again, 1 mm caudal to the injury site, and the tracer Neurobiotin™ was applied. Double-labeled cells were observed in the caudal portion of the serotonin-immunoreactive vagal nucleus of the caudal rhombencephalon. In order to investigate whether the reinnervation was due to sprouting from axons above the injury site or to regeneration of axotomized axons, the experiments were performed again, but the tracer Fluoro-Gold™ was applied at the time of transection. Triple-labeled cells were observed in the vagal nucleus, indicating that at least part of the reinnervation corresponds to true regeneration. This study provides a new and interesting model for investigating the intrinsic molecular mechanisms involved in regeneration of the serotonergic descending axons in vertebrates. Use of this model may provide valuable information for proposing new therapies for patients with spinal cord injury.
Introduction
A
The presence of serotonergic cells with projections to the SC has been reported in the rhombencephalon (raphe nuclei) of different jawed vertebrates, such as zebrafish (McLean and Fetcho, 2004a,2004b), amphibians (van Mier et al., 1986; Woolston et al., 1994), and rats (Aitken and Törk, 1988). Descending serotonergic projections to the spinal cord play a role in the modulation of locomotion at spinal levels in both mammals (Jordan et al., 2008) and lampreys (Grillner et al., 2008). Moreover, the development and anatomical organization of the serotonergic SC-projecting neurons in the brainstem is highly conserved between lampreys and mammals (Barreiro-Iglesias et al., 2008). In mammals, paralysis caused by SCI results from a loss of supraspinal tracts that underlie voluntary initiation of locomotion (e.g., corticospinal or reticulospinal glutamatergic tracts; Rekling et al., 2000; Murray et al., 2010). Furthermore, the paralysis that occurs following SCI also occurs because of loss of descending brainstem tracts that provide spinal neurons with their major source of neuromodulators, such as serotonin (5-HT) (Murray et al., 2010). In mammals, a loss of brainstem-derived 5-HT after SCI acutely reduces motor neuron excitability and accordingly depresses all motor functions (Murray et al., 2010). Recent studies have also shown that, in rodents, autonomic dysreflexia after SCI is associated with the lack of serotonergic axonal reinnervation (Cormier et al., 2010). Regeneration of the serotonergic descending projections will therefore probably benefit therapies designed to facilitate recovery of function after SCI.
Spontaneous regeneration of the serotonergic descending projections after SCI has not been experimentally demonstrated in any vertebrate species. Immunofluorescence experiments carried out at spinal levels in the sea lamprey have shown that serotonergic processes reinnervate the SC below the lesion site after a complete SC transection (Cohen et al., 2005). However, regeneration of the severed serotonergic projections from brainstem neurons was not conclusively demonstrated in the latter study. Thus, recovery of serotonergic innervation below the site of injury may be due to axonal sprouting from intrinsic spinal neurons. In the present study, we show, by means of combined immunofluorescence and tract-tracing methods, that descending serotonergic neurons of the caudal rhombencephalon regenerate their axons across the lesion site after complete SC transection in the sea lamprey. This study offers a new and interesting model for investigating the intrinsic molecular mechanisms that favor regeneration of the descending serotonergic axons in vertebrates, which may provide valuable information for developing new therapies for patients with SCI.
Methods
Mature larval sea lampreys (between 100 and 140 mm in body length; 5–7 years old) were caught in the river Ulla (Galicia, northwest Spain), and maintained in freshwater with a bed of river sediment and appropriate ventilation and temperature conditions until use. Before the experiments, all animals were deeply anesthetized with 0.05% benzocaine (Sigma-Aldrich, St. Louis, MO) in freshwater. The experiments were approved by the Bioethics Committee at the University of Santiago de Compostela, and conformed to the European Union (86/609/EEC) and Spanish (Royal Decree 223/1998) regulations for the care and handling of animals in research.
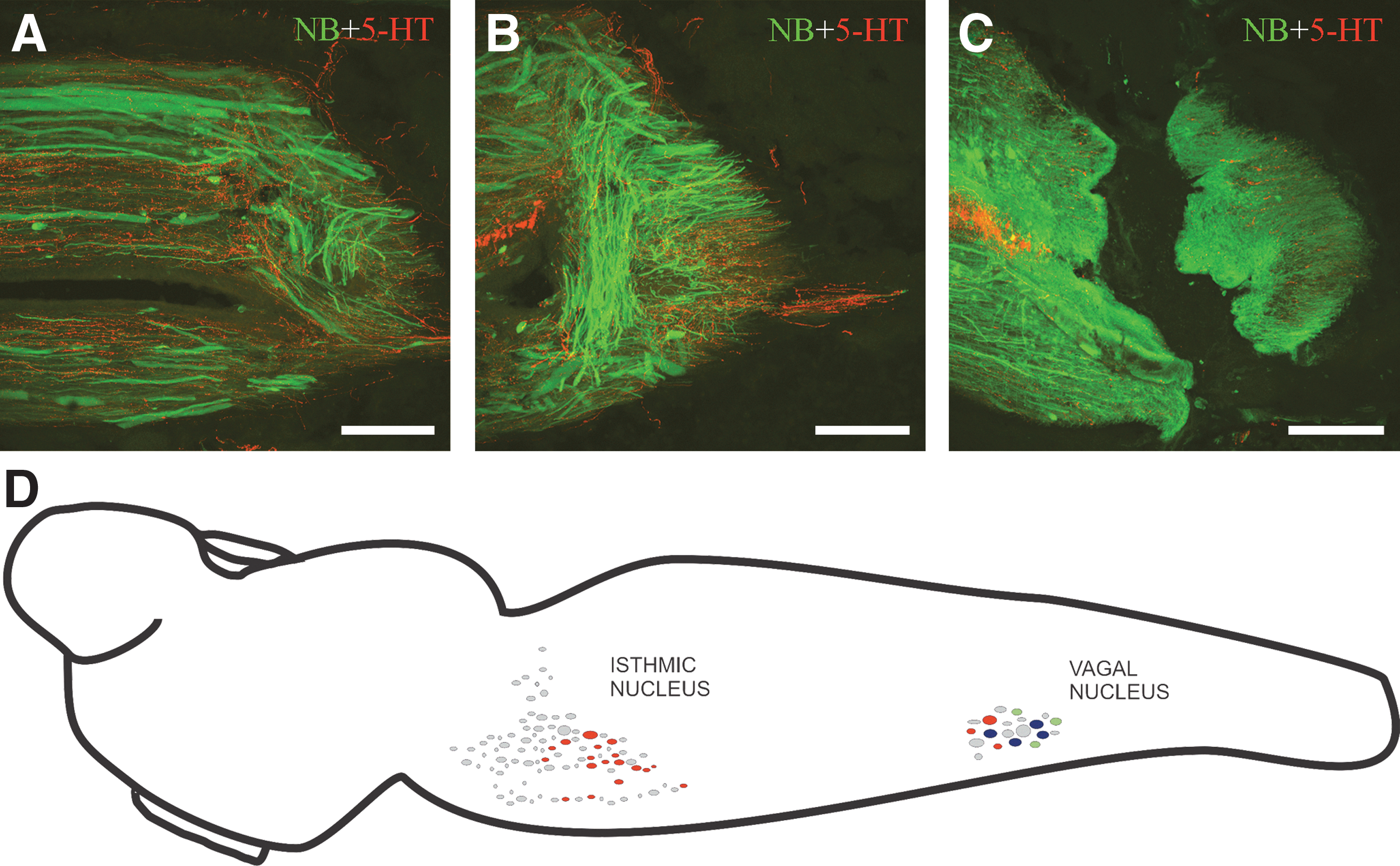
In experimental animals (n=5), the SC was exposed from the dorsal midline at the level of the fifth gill, complete SC transection was performed with a scalpel, and the SC cut ends were visualized under a microscope. The incision was closed with Histoacryl® tissue glue (B. Braun Surgical, Tuttlingen, Germany). Each transected animal was examined 24 h after surgery to confirm that there was no movement caudal to the lesion site. Transection was tentatively considered complete if on stimulation of the head, the animal could move only its head and body rostral to the lesion site. The animals were allowed to recover in freshwater tanks at 19–20°C, since lampreys are known to be able to recover full functionality at this temperature (Cohen et al., 1999). After a recovery period of 5 months, the SC was again exposed and crystals of the tracer Neurobiotin™ (NB; 322.8 Da molecular weight; Vector Laboratories, Burlingame, CA) were applied with the aid of a minute pin (00) 1 mm caudal to the injury site at the level of the sixth gill (Fig. 1A–C). The incision was again closed with Histoacryl, and the animals were maintained at 4°C with appropriate aeration conditions in lamprey Ringer solution of the following composition: 137 mM NaCl, 2.9 mM KCl, 2.1 mM CaCl2, and 2 mM HEPES, for 5 days to allow transport of the tracer. After the incubation period, the larvae were deeply anesthetized, and the heads (from the snout to the seventh gill) were fixed by immersion in 4% paraformaldehyde in 0.4 M Tris buffer (pH 7.4; TBS) for 6 h. The samples were rinsed in TBS, and then cryoprotected with 30% sucrose in TBS overnight, embedded in Tissue-Tek® (Sakura, Torrance, CA), frozen in nitrogen-cooled isopentane, and cut serially on a cryostat (20 μm thick) in the transverse (from the snout to the third gill), and horizontal (from the third gill to the seventh gill) planes. The sections were mounted on subbed glass slides. The sections were then incubated with a rabbit polyclonal anti-5-HT antibody (dilution 1:2500; Incstar, Still Water, MN) at 4°C for 72 h. For indirect immunofluorescence detection of 5-HT, Cy3-conjugated goat anti-rabbit antibody (dilution 1:200; Millipore, Temecula, CA) was used (incubation time: 1 h at room temperature). All antibodies were diluted in TBS containing 15% normal goat serum and 0.2% Triton X-100 as detergent. For indirect detection of the tracer, the sections were incubated at room temperature for 4 h with fluorescein isothiocyanate (FITC)-labeled avidin D (Vector Laboratories) previously diluted to 1:1000 with TBS containing 0.3% Triton X-100. The sections were rinsed in distilled water and then cover-slipped with mounting medium for fluorescence (Vectashield; Vector Laboratories). Photomicrographs were taken with a spectral confocal laser microscope (Leica TCS-SP2), and adjusted for brightness and contrast with Adobe Photoshop 7 or CS2 software. Cell number and sizes were counted and measured in confocal stacks with the aid of Leica confocal software (LITE). To avoid counting cells twice, a series of photographs were aligned with LITE, and the positions of double-labeled cells in pairs of consecutive sections were compared. This unbiased approach made it unnecessary to introduce any correction factor for possible overestimation of cell numbers. The total number of serotonergic descending cells was counted in each animal, and the minor axes of these cells were measured. Statistical comparisons were performed by a Student's t-test, with IBM SPSS 18.0 software. Data are reported as means±standard deviations.
(
Control larvae (n=3) were processed as above for NB labeling at the level of the sixth gill and 5-HT immunofluorescence, but without prior transection of the SC at the level of the fifth gill. In order to confirm that the neurons labeled with NB in injured animals actually correspond to neurons that reinnervated levels below the lesion site, some larvae (n=4) were processed in the same way as the experimental larvae, but the tracer NB was applied 1 week post-injury, when severed axons are suffering retraction but regrowth has not started (Shifman et al., 2007). In order to determine whether the serotonergic neurons labeled with NB after the SC transection actually correspond to neurons with regenerated axons and not to non-injured neurons that had regrown their axons from above, some larvae (n=3) were processed in the same way as the experimental group, but the tracer Fluoro-Gold™ (FG, molecular weight 532.6 Da; Fluorochrome LLC, Denver, CO) was applied to the SC at the level of the fifth gill at the time of SC transection.
Results
In the sea lamprey, 5-HT-immunoreactive (-ir) fibers of the SC have two origins: intrinsic fibers arising from 5-HT-ir cells of the ventral margin of the SC (Abalo et al., 2007; Barreiro-Iglesias et al., 2009), and extrinsic 5-HT-ir fibers of supraspinal origin (Barreiro-Iglesias et al., 2008). We have previously reported the development and organization of the descending serotonergic projections to the SC of the sea lamprey as assessed by combined tract-tracing and immunofluorescence techniques (Barreiro-Iglesias et al., 2008). In mature larval sea lampreys, two brainstem nuclei bear 5-HT-ir cells that project to the rostral SC (at the level of the second/third gill, which is close to the brainstem-SC transition), and the isthmic and vagal serotonergic nuclei (Fig. 1D; Barreiro-Iglesias et al., 2008). Here we performed control experiments in which the tracer NB was applied at the level of the sixth gill of non-injured animals. In these animals, serotonergic descending cells were only observed in the caudal portion of the vagal nucleus of the caudal rhombencephalon (Fig. 1D). The number of double-labeled cells in the vagal nucleus was 18.7±4.2 (n=3), and the size of these descending 5-HT-ir neurons was 11.3±2.6 μm in the minor axis (Fig. 2A–D). The experiments in control larvae revealed that the serotonergic descending cells of the isthmic nucleus probably only project to the rostral SC (at the level of the second/third gill), whereas some serotonergic descending cells of the vagal nucleus project to more caudal SC levels (at the level of the sixth gill).

Photomicrographs of transverse sections of the serotonergic reticulospinal group of the vagal region of mature larval sea lampreys as revealed by combined retrograde tract-tracing using Neurobiotin (NB; green channel), Fluoro-Gold (FG; blue channel), and serotonin (5-HT) immunofluorescence (red channel). (
In experimental larvae, complete SC transection was done at the level of the fifth gill to ensure that all serotonergic descending neurons projecting to the SC at the level of the sixth gill were axotomized. Five months post-injury and after application of the tracer NB at the level of the sixth gill (1 mm caudal to the injury site), double-labeled 5-HT-ir cells were also observed in the vagal nucleus (Fig. 2E–H), and not in the isthmic nucleus. As in normal animals, 5-HT-ir descending neurons of the vagal nucleus with regenerated axons were observed in the caudal portion of the nucleus. The number of 5-HT-ir neurons labeled by the tracer NB 5 months after complete SC transection in the vagal nucleus was 7±1.4 (n=5), and the size of these neurons was 8.7±1.8 μm in the minor axis. Statistical analysis revealed a significant reduction in the size of neurons between control and experimental larvae (p<0.05). When the tracer NB was applied at the level of the sixth gill to larvae 1 week post-injury (n=4), no double-labeled cells were observed in the vagal nucleus, or in any other brain region. In larvae in which the tracer FG was applied at the level of the fifth gill at the time of SC transection, and NB was applied at the level of the sixth gill 5 months later, between 1 and 3 5-HT-ir neurons labeled by both FG and NB were observed in the vagal nucleus (n=3; Fig. 2I). In addition, 5-HT-ir cells only labeled by NB (6±4.3), or only labeled with FG (9.5±3.5), were also observed (data not shown).
Discussion
Previous studies in invertebrates have shown the spontaneous regeneration of axotomized serotonergic neurons (Chiasson et al., 1994; Koert et al., 2001; Murphy et al., 1985), but this phenomenon has not previously been described in any vertebrate species. This is therefore the first experimental study reporting the spontaneous regeneration of at least part of the serotonergic descending projection after complete SC transection in a vertebrate species. A previous study in the sea lamprey reported the recovery of 5-HT expression below the site of injury after complete SC transection in the mid-body region (Cohen et al., 2005). However, in the previous study regeneration of the serotonergic descending projection was not experimentally assessed. Since the lamprey SC possesses serotonergic neurons (Abalo et al., 2007; Barreiro-Iglesias et al., 2009; Harris-Warrick et al., 1985; Iglesias et al., 2009), it was not known if reinnervation of the SC below the site of injury was due to actual regeneration of the severed descending axons or to compensatory reinnervation from intraspinal serotonergic cells. Here we experimentally demonstrated that, after complete SC transection, at least some of the serotonergic descending neurons of the caudal rhombencephalon regenerate to levels caudal to the lesion site.
The number of serotonergic descending cells in the vagal nucleus revealed by application of FG at the level of the fifth gill was approximately half the number revealed by application of NB at the sixth gill level. This strongly suggests that FG was metabolized or not transported in about 50% of the neurons projecting to the level of the fifth gill. With this in mind, comparison of the results obtained in control and experimental animals revealed some interesting findings: (1) a population of serotonergic cells of about 40% of the original population of descending serotonergic neurons in the vagal nucleus reinnervated levels caudal to the lesion site after complete SC transection; (2) the size of the descending serotonergic cells after SCI was significantly smaller than the descending serotonergic cells in control larvae; and (3) at least 20% of the descending serotonergic neurons regenerate their axons after axotomy (this may be as much as 40% if FG is metabolized or not well transported). In the sea lamprey, the regenerative abilities differ greatly between identified reticulospinal neurons and neuronal groups (Jacobs et al., 1997). In previous studies by other groups, neurons regenerating 20% of the times or less have been considered as “bad regenerating” reticulospinal neurons (Shifman and Selzer, 2000; Shifman et al., 2009). In the present study, we report the regenerative ability of a neuronal population defined by its neurochemistry and not only by its anatomical location. In addition, the present results indicate that sprouting of non-axotomized neurons from above may help to compensate for the lost descending serotonergic innervation. This is consistent with recent data in mammals, which show that serotonergic neurons have a better sprouting ability than other neurons of the central nervous system (Hawthorne et al., 2011). In any case, the serotonergic descending projection is not completely restored after SC transection in mature larval sea lampreys. This indicates that partial recovery of the serotonergic descending innervation below the SC lesion site may be sufficient to allow functional recovery of locomotion.
It is not known why complete restoration of the serotonergic descending projection does not occur. A recent study in the sea lamprey has shown that the axons of some identifiable reticulospinal neurons do not regenerate after complete SC transection, because they undergo a process of delayed cell death and eventually disappear (Shifman et al., 2008). Thus, the absence of regeneration of the entire serotonergic descending projection after axotomy may also be due to activation of cell death or atrophy. Shrinkage of the cell bodies of the serotonergic neurons with regenerated axons (present results) may support the idea that because of the SCI event, axotomy affects the viability of the serotonergic neurons, and that only those neurons that “escape” from the cell death process would regenerate their axons. Another possible explanation for the smaller size of the regenerated cells is that the serotonergic cells with small cell somas are better regenerators than the serotonergic cells with larger somas. In the present study, only 25% of the descending serotonergic cells had small somas (around 8.3 μm) in control animals, whereas about 40% of the descending serotonergic cells regenerated the axons, and almost all of them had small somas, after complete SC transection. The difference in the percentage of cells with small somas in all descending cells of the vagal group between control and experimental groups supports the idea that the somas of serotonergic neurons shrink after axotomy.
The molecular processes guiding the growth of the regenerating serotonergic axons are not known. In invertebrates, it has been shown that 5-HT released from the regenerating axon may autoregulate this process by fine-tuning axon guidance, and branching by inducing local collapse responses in the extending neurite (Koert et al., 2001). The sea lamprey may serve as a vertebrate model in which to study the possible role of this neurotransmitter in the autoregulation of axonal regrowth of the severed axon.
The serotonergic descending projection to the SC modulates locomotor activity and appears to promote the optimal function of the central pattern generator networks (CPGs) in vertebrates, from lampreys (Grillner et al., 2008) to mammals (Jordan et al., 2008). The regeneration of this system after SCI probably contributes to restoring the locomotor CPGs. Previous studies in rats have shown that, after complete SC transection, transplantation of embryonic raphe cells enhances recovery of motor functions, and that this enhancement is 5-HT-related (Majczyński et al., 2005). In addition, other studies have reported that combinations of Schwann cell bridges, olfactory-ensheathing glia transplantation, and chondroitinase ABC treatment, promote the regeneration of descending serotonergic fibers and recovery of locomotion after SCI in rats (Fouad et al., 2005; Vavrek et al., 2007). However, it should be noted that all forms of therapy that involve cell transplantation require labor-intensive in vitro propagation and manipulation, followed by transplantation and establishment of the cells at appropriate sites in injured patients (Barreiro-Iglesias, 2010). Finding a non-invasive way of activating the regeneration of the severed serotonergic descending processes in vivo would be a good way of avoiding cell transplantation. Future studies in the sea lamprey model should identify cellular mechanisms underlying the spontaneous regeneration of serotonergic descending axons after SCI, allowing potential application of these mechanisms to enhance spontaneous sprouting, or to induce regeneration of severed serotonergic axons in mammals. This direction of research will be important for generating therapies for humans with SCI.
Footnotes
Acknowledgments
This work was supported by a grant from the Xunta de Galicia (INCITE08PXIB200063PR).
Author Disclosure Statement
No competing financial interests exist.