
Abstract
Curcumin has been proposed for treatment of various neuroinflammatory and neurodegenerative conditions, including post-traumatic inflammation during acute spinal cord injury (SCI). In this study, we examined whether curcumin anti-inflammation involves regulation of astrocyte reactivation, with special focus on the injury-induced RANTES (regulated on expression normal T-cell expressed and secreted) from astrocytes in acute SCI. Male Sprague-Dawley (SD) rats were subjected to impact injury of the spinal cord followed by treatment with curcumin (40 mg/kg i.p.). RANTES and inducible nitric oxide synthase expression as well as RANTES-positive astrocytes were all induced by injury accompanied by the elevation of lipid peroxidation, and attenuated by the curcumin treatment. In primary cultured rat astrocytes challenged with lipopolysaccharide (LPS) to mimic astrocyte reactivation following SCI, LPS induces robust increase of RANTES expression and the effect was also reduced by 1 μM curcumin treatment. Furthermore, cortical neurons cultured with astrocyte conditioned medium (ACM) conditioned with both LPS and curcumin (LPS-curcumin/ACM), which characteristically exhibited decreased RANTES expression when compared with ACM from astrocytes treated with LPS alone (LPS/ACM), showed higher level of cell viability and lower level of cell death as assessed by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction activity assay and lactate dehydrogenase release assay, respectively. Knockdown of RANTES expression by siRNA (siRANTES) shows reduced RANTES expression and release from LPS-reactivated astrocytes, and ACM obtained from this condition (LPS-siRANTES/ACM) becomes less cytotoxic as compared with the LPS-ACM. Therefore, curcumin reduction of robust RANTES production in reactivated astrocytes both in vitro and in vivo may contribute to its neuroprotection and potential application in SCI.
Introduction
I
The therapeutic target of SCI is the prevention or amelioration of secondary damage to the spinal cord. Curcumin, a lipid soluble antioxidant from curry spice, accentuates radical decay by electron donation from its phenolic hydroxyl group to react with various free radicals, such as lipid peroxyl radicals and superoxide (Priyadarsini, 1997). Curcumin has been proposed as a potential therapeutic for a number of inflammatory and neurodegenerative states, such as Alzheimer's disease, multiple sclerosis, and brain trauma, for its anti-inflammatory, anti-oxidant, and immunomodulatory activities (Menon and Sudheer, 2007). Curcumin antagonizes several mediators of the inflammatory cascade, including activation of nuclear factor-κB, and inducible nitric oxide synthase (iNOS) (Weber et al., 2006). Curcumin is also a superior NO compared to vitamin E (Chan et al., 1998), and functions as a potent neuroprotectant by decreasing lipid peroxidation to attenuate mitochondrial dysfunction in a cerebral ischemia animal model (Wang et al., 2005). Previously, we showed that curcumin inhibited apoptosis and neuronal death, abated astrocyte reactivation, and improved functional outcomes 7 days following SCI (Lin et al., 2011). In that study we observed that curcumin downregulated glial fibrillary acid protein (GFAP) which suggests that curcumin attenuates astrocyte reactivation. The protective effect on neurons may also be attributed to the anti-inflammation effect of curcumin. The role of astrocytes in protecting neurons has been an area of significant research interest and has emerged as a potential therapeutic target in the treatment of SCI.
Chemokines such as RANTES (regulated on expression normal T-cell expressed and secreted) play a double role in neuronal survival in brain injury: a moderate amount of RANTES secreted from astrocytes has been proven protective in Huntington's disease (Chou et al., 2008) and glutamate excitotoxicity (Lin et al., 2010); whereas prolonged and excessive release during neurotrauma by inflammatory stimuli can lead to detrimental outcomes (Pease, 2006). Elevated levels of RANTES are seen in trauma, inflammation, infection, and neurodegenerative diseases (Mennicken et al., 1999; Tripathy et al., 2010). Luo et al. (2002) stimulated cultured mouse astrocytes with RANTES or structurally related chemokines and observed that astrocytes release proinflammatory mediators and reprogrammed their surface molecules, suggesting that RANTES amplifies inflammatory responses within the CNS. Accordingly, any novel treatment that modulates the multifunctional mediator, RANTES, in diminishing neuroinflammation and augmenting neuroprotection would be a valuable therapeutic advancement in SCI therapies.
In the present study, we sought to assess the effects of curcumin on RANTES expression and secretion by astrocytes following SCI. Our results show that curcumin treatment could reduce RANTES in both injured rat spinal cord and lipopolysaccharide (LPS)-stimulated primary cultured astrocytes. Importantly, we found that reducing astrocyte production of RANTES by curcumin and RANTES small interfering RNA (siRNA) could attenuate the neurotoxicity of the LPS-stimulated astrocyte conditioned medium, which suggest that the neuroprotective effect of curcumin in SCI might be attributed to its attenuating effect on the robust RANTES production in injury-activated astrocytes.
Methods
Spinal cord contusion injury in rats
Male Sprague-Dawley (SD) rats (BioLASCO Taiwan, Taipei, Taiwan) weighing 280–330 g were kept two per cage for at least 5 days after their arrival at our laboratory. The rats had access to food and water ad libitum and were housed within a room with a 12:12 h dark–light cycle. This study was performed in accordance with the guidelines provided by the Experimental Animal Laboratory and approved by the Animal Care and Use Committee at Taipei Medical University.
The acute SCI animal model was performed as described previously (Sun et al. 2010). Briefly, the animals were divided into three groups: sham control (n=4), SCI (n=4), and SCI+curcumin (SCI+Cur) (n=4). The rats were anesthetized with isoflurane and placed onto a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA) to position the spinal cord. Laminectomy was performed at the 9th to 10th thoracic vertebrae under a dissecting microscope and the dura mater was left intact. After laminectomy, SCI was performed using a New York University (NYU) impactor with a 10-g weight dropped from the height of 25 mm. Vehicle or curcumin was administered via an intraperitoneal route immediately after the SCI. The fascia and skin were closed with sutures for each layer, and the animals were allowed to recover on a 36.5°C heating pad. Postoperative treatment consisted of saline (1.0 ml subcutaneously) for rehydration, and the animals were returned to preoperative housing conditions. The animals were eating and drinking within 3 h after surgery. Histological assessment of the extent of injured lesion was confirmed 24 h after SCI with or without curcumin treatment. A 5-mm section of the spinal cord with the lesion, or a similar area in the sham control group, was obtained for mRNA extraction and histological analysis.
Treatment of SCI rats
Rats were randomly divided into three groups: vehicle-treated sham-operated control (sham; laminectomy only), vehicle-treated SCI group (SCI), and SCI rats post-treated with curcumin (SCI+curcumin) (each group n=4). Curcumin was purchased from Sigma-Aldrich (St. Louis, MO) and dissolved in 10 % DMSO (Sigma-Aldrich). The SCI groups received intraperitoneal administration of 10% DMSO, and the SCI+curcumin group received intraperitoneal administration of curcumin 40 mg/kg once 30 min after SCI, which dosage used has been shown neuroprotective in our previous study in an SCI rat model (Lin et al., 2011) as well as in the ischemic brain injury rodent model (Wang et al., 2005). The spinal cord tissue were harvested or fixed 24 h after the treatment for mRNA measurement or immunohistochemistry, respectively. The sham group was only subjected to laminectomy without SCI, and administered 10% DMSO i.p. at 1h after laminectomy.
Histology
Animals underwent deep anesthesia with isoflurane on the second day following SCI, and then were perfused with phosphate-buffered saline (PBS), followed by cold 4% paraformaldehyde in 0.15 M sodium phosphate buffer, pH 7.4. The spinal cord was removed immediately, post-fixed for 8 h in the same fixative at 4°C, and cryoprotected for 2–3 days in 15% and 30% sucrose. The spinal cord was frozen in powdered dry ice and stored at −80°C until analyzed. Five-micrometer sections were cut on a cryostat with a sliding microtome at the caudal, central, and rostral parts of spinal cord contusion. Heat-induced antigen retrieval was performed in the sections for further immunostaining as previously described (Chang et al., 2010). Briefly, 5-mm-thick spinal cord tissue sections were cut from paraffin-embedded samples and then de-waxed and rehydrated. The rehydrated sections were transferred to a microwaveable slide container and then heated three times in 500 ml of retrieval solution (0.01 M citrate buffer, pH 6.0) at 750 W power for 7 min each. Sections were allowed to cool at room temperature for 15 min and then washed in 0.05 M Tris buffer saline (TBS), pH 7.0, before antibody application.
Double immunofluorescent staining
Double immunofluorescent staining was performed as previously described by Tomita et al. (2005). Briefly, tissue sections were blocked for nonspecific antibody binding with normal horse (S-2001, Vector Laboratories, Burlingame, CA) or goat (S-1000, Vector Laboratories) sera for 1 h at room temperature. Sections were then consecutively (per slide) incubated with the mixture of antibodies: goat anti-rat RANTES (1:100, Santa Cruz Biotechnology, Inc., Santa Cruz, CA) and mouse anti-rat GFAP (1:1000, Chemicon, Temecula, CA) overnight at 4°C. Once the sections were incubated in the primary antibody, sections were incubated in biotinylated anti-goat IgG secondary antibody. (1:200, Vector Laboratories), rinsed, and incubated with the third antibodies: Texas Red-conjugated Avidin D (1:200, Vector Laboratories) and goat anti-mouse IgG conjugated fluorescein isothiocyanate (FITC) (1:200, Jackson Immunoresearch Laboratories, West Grove, PA) for 2 h at room temperature. Glass cover slips were mounted with fluorescent mounting medium (Vector Laboratories.) For low magnification images of the whole spinal cord section, sections were photographed using an Olympus IX71 fluorescent microscope and DP50 digital camera (Olympus, Tokyo, Japan). For high magnification images, sections were examined by Leica TCS SP5 Confocal Microscope Imaging System (Leica, Solms, Germany). GFAP and RANTES labeled cells at 40 x (low magnification) were photographed and counted. At least five randomly selected fields per section for three sections with the 1-cm epicenter segment were counted. Each section was 5 μm in thickness and the distance between the three selected sections was 10 μm. To study morphological alternation of astrocyte reactivation following SCI, we selected dorsal column white matter at the epicenter as region of interest (ROI) for cell quantification analysis, which has been proven most extensively injured in SCI (Agrawai and Fehlings, 1997). Total GFAP-positive cell numbers in each field were ∼ 30–40 cells in this condition. The percentage population of astrocytes expressing RANTES was calculated by dividing the number of GFAP-RANTES double positive astrocytes by the total number of GFAP+ cells.
Lipid peroxidation
Lipid peroxidation of rat spinal cord tissue was analyzed using malondialdehyde (MDA) assay (Ates et al., 2007; Uchiyama and Mihara, 1978). The whole spinal cord tissue of sham-operated, SCI, and SCI+cucumin rats was removed 24 h after injury. The tissues were homogenized in 10 volumes (w/v) of cold 1.5% KCl. Homogenate at 0.5ml was mixed with 3 ml of 1% H3PO4 and 1 ml 0.6% thiobarbituric acid, then heated in a dry bath for 1 h. The MDA was extracted into 4 ml n-butanol and the absorbance was measured at 535 nm and 520 nm. Tetramethoxypropane was used as the standard. The tissue lipid peroxidate levels were calculated as nanomole MDA/g wet tissue.
Real time reverse-transcription polymerase chain reaction (RT-PCR)
Total RNA was prepared by directly lysing the cultured astrocytes in extraction buffer (Trizol/phenol/chloroform), and reverse transcribing the mRNA into cDNA using oligo-dT and SuperScript II reverse transcriptase (Invitrogen). The cDNAs were subjected to real time PCR to measure the expression of RANTES, iNOS, and the housekeeping gene cyclophilin as an internal control.
For real time PCR, the cDNA samples were analyzed by the SYBR Green gene expression system (ABI PRISM 7300 HT real-time PCR system; Applied Biosystems, Foster City, CA). The minor groove binding dyes and primers for the detection of RANTES, iNOS, and cyclophilin were designed by ABI. The threshold cycle (Ct) (the fractional cycle number at which the amount of amplified target reached a fixed threshold) was determined and Ct value of RANTES and iNOS were normalized by the housekeeping gene cyclophilin, which was used as an internal control. Primers used for real time PCR are shown in Table 1. Each measurement was performed at least in triplicate.
Animals for primary cultures of astrocytes and cortical neurons
Pregnant female SD rats and neonatal 1- to 2-day-old SD rats were used in this study for the primary culture of cortical neurons and astrocytes, respectively. Animals were obtained from BioLasco, Taipei, Taiwan. Rats were killed by an overdose of sevoflurane (Abbott, Osaka, Japan) to minimize pain or discomfort in accordance with the United States National Institute of Health guidelines regarding the care and use of animals for experimental procedures. The procedures for killing pregnant and postnatal rats in this study were reviewed and approved by the Experimental Animal Review Committee at Taipei Medical University.
Primary astrocyte cultures and astrocyte-conditioned medium (ACM)
Astrocyte cultures were prepared as described previously (Lin et al., 2011). Briefly, cerebral cortices were harvested from 1- to 2-day-old SD rats, homogenized by mechanical dissociation, and the cell suspension was diluted in Dulbecco's modified Eagle's medium (DMEM) with F12 media (Invitrogen) supplemented with 10% heat-inactivated fetal bovine serum (FBS). Cells were seeded onto 75-cm2 flasks at an initial density of 2×106 cells/flask. Six to 8 days after seeding, microglia and oligodendrocytes were removed by orbital shaking of the culture flasks at 160 rpm for 24 h at 37°C. The suspended cells were decanted to obtain a pure astrocyte layer adhered at the bottom. The purified astrocytes were subcultured onto 35-mm culture dishes with DMEM/F12 medium containing 10% FBS in a humidified tissue culture incubator with 5% CO2 maintained at 37°C. The characteristic morphology indicated that the astrocyte culture was a minimum ∼ 85% purity. For preparation of ACM, astrocytes grown for 7 days in vitro (DIV) were conditioned with LPS 1 μg/ml or 1 μg/ml LPS +1 μM curcumin for 1 h, re-fed with serum-free DMEM/F12 medium, and incubated for another 23 h. This medium was used to feed primary cortical neurons. The resulting ACM was either used immediately or stored at −80°C.
Curcumin treatment of LPS-induced reactivated astrocytes
Primary cultured astrocytes grown for 7 DIV were challenged with LPS. Astrocytes can be reactivated in response to LPS. This culture model was developed by Tomita et al. (2005) to mimic the conditions of glial reactivation following CNS injury in vivo. To study the effect of curcumin on reactivated astrocytes, primary astrocyte cultures (1 x 106 in a 35-mm dish) grown for 7 DIV were incubated with 0 μg/ml (control group) and 1 μg/ml LPS and 1μg/ml LPS with 1 μM curcumin for 24 h. The mRNA level of RANTES and iNOS was analyzed by real-time RT-PCR. Each experiment was performed at least twice using at least three different astrocyte cultures. LPS from Escherichia coli serotype 055: B5, L-2880 (Sigma Chemical Co., St. Louis, MO) was used.
Immunoblotting of GFAP
Immunoblotting was performed as previously described (Sun et al., 2010). In brief, total protein was extracted from cultured rat astrocytes in a lysis buffer containing 20 mM Tris, pH 8.0, 150 mM NaCl, 1% IGPEAL CA-630, 5% glycerol, and 1x complete protease inhibitor cocktail (Roche, Penzberg, Germany). Forty microgram protein extracts were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis on a 4–12% gradient gel followed by electro- transfer of the electrophoresed proteins from the gel to a nitrocellulose membrane. The membrane was incubated with blocking reagent (Roche, Penzberg, Germany), primary antibodies (rabbit anti-GFAP or mouse anti-GAPDH antibodies [Millipore, Billerica, MA; both were in 1:500 dilution]) overnight at 4°C, and then washed and incubated with horseradish peroxidase (HRP)-conjugated secondary antibody (Jackson ImmunoResearch Laboratories, West Grove, PA) for 1 h at 25°C. Protein was detected by enhanced chemiluminescence protocol (ECL Western blotting detection reagents; Amersham Biosciences, Little Chalfont, U.K.), and the labeled bands were visualized and quantified using the Digital Science Electrophoresis Documentation and Analysis System (Berthold Technologies, Bad Wildbad, Germany).
Primary cultured cortical neurons and cortical neurons cultured with ACM
Primary cortical neuron cultures were prepared from embryonic day 17 rat brains and characterized as previously described by Lin et al. (2008). Cells were re-suspended in the same media and seeded onto a 35-mm culture dish (IWAKI, Tokyo, Japan) coated with poly-
RNA interference and transfection
We used an siRNA specific for the RANTES mRNA to knock down the expression of RANTES in cultured astrocytes as described previously, with some modification (Lin et al., 2008). Astrocytes were transfected with a set of siRNAs specific for RANTES or scrambled RNA (Silencer Pre-designed siRNA, Ambion) using Lipofectamine™ 2000 reagent (Invitrogen). The siRNA sequences for RANTES were 5’- acg ugaagga guauuuuu at -’3 (exon 2) and 5’-CCAUAGAGAUUAUAACUGUtt-’3 (exon 3). Five hours after transfection, the Lipofectamine 2000-containing medium was replaced with astrocyte culture medium to allow cells to recover for another 67 h. Real-time RT-PCR and enzyme-linked immunosorbent assay (ELISA) in ACM were performed to verify the mRNA and protein level of RANTES for the knockdown efficiency.
Neuronal survival analysis
Neuronal survival rate was assessed by using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reduction activity assay (for cell viability), and the lactate dehydrogenase (LDH) release assay (for necrotic cell death). For the MTT assay, cortical neurons cultured in ACM were incubated with MTT for 4 h, followed by addition of 0.3 ml of 0.4 N HCl in isopropanol to the mixture overnight to dissolve the formazan. The dissolved suspension was subjected to an ELISA reader and the absorbance at a wavelength of 600 nm was measured. For the LDH release assay, cortical neurons incubated in ACM were equilibrated with BME medium for 1 h, and 250 μL of culture medium was collected and incubated with 0.1 mg of β-NADH in 1.2 ml of 0.1 mol/L phosphate buffer for 5–15 min at 25°C. The absorbance at a wavelength of 340 nm was measured for 3 min immediately after 0.1 mol/L sodium pyruvate was added to the mixture. The unit activity of LDH was calculated by multiplying the decrease of 340 nm in 1 min by 4000 to obtain the activity in 1 ml of sample.
Detection of RANTES by ELISA
To evaluate the effects of curcumin on RANTES secretion by LPS-induced reactivated astrocytes, ELISA was used to detect RANTES protein released in ACM using Quantikine Mouse RANTES Immunoassay (R&D Systems Inc.). Control or LPS- treated ACM with or without curcumin was subjected to the RANTES immunoassay system according to the manufacturer's protocol. RANTES standards were measured in duplicates, and samples in triplicates, with each group containing four independent samples.
Statistical analysis
Independent two sample t-tests were used to compare data between the control and curcumin groups. If there were more than two groups, one-way analysis of variance (ANOVA) was used to compare continuous variables. When significant differences were apparent, multiple comparisons were performed using the Newman-Keuls method. Data are presented as means±SEM. All statistical assessments were two-sided and evaluated at 0.05 level of significant difference. Statistical analyses were performed using GraphPad Prism software 5.0 (GraphPad Software, Inc; La Jolla, CA).
Results
Curcumin attenuated RANTES and iNOS mRNA expression in vivo 24 h after SCI
SCI was induced with the main impact at T9-T10 in rats. Rats were then treated with either intraperitoneal administration of normal saline or curcumin 40 mg/kg once immediately and continuing for the second day. Real-time PCR was used to examine RANTES and iNOS mRNA expression in the central, rostral, and caudal regions of the spinal cords (Fig. 1A and B). Each experiment was performed three times. Rats subjected to SCI showed significantly upregulated RANTES (p<0.01) and iNOS mRNA expression (p<0.001) at epicenter when compared to control rats. Control rats showed almost no iNOS mRNA expression. Rats with SCI treated with curcumin showed a significant decrease in RANTES and iNOS mRNA expression in the central region of the spinal cord (p<0.05) when compared to untreated rats with SCI. No effect was observed at the spinal segments rostral or caudal to the central region.

Curcumin downregulated RANTES and iNOS expression in vivo in SCI rat model. (
Curcumin-attenuated RANTES expression in vivo 24 h after SCI
After SCI, there was an increase in RANTES immunofluorescent staining in the SCI and SCI+Cur groups at the injured site as compared with the sham group (Fig. 2A–C). Rats with SCI treated with curcumin showed a decrease in protein expression of RANTES in the injured region of the spinal cord when compared to untreated rats with SCI. These morphologic findings were comparable with the inhibitory effects of curcumin on the RANTES mRNA upregulation in rats (Fig.1A).
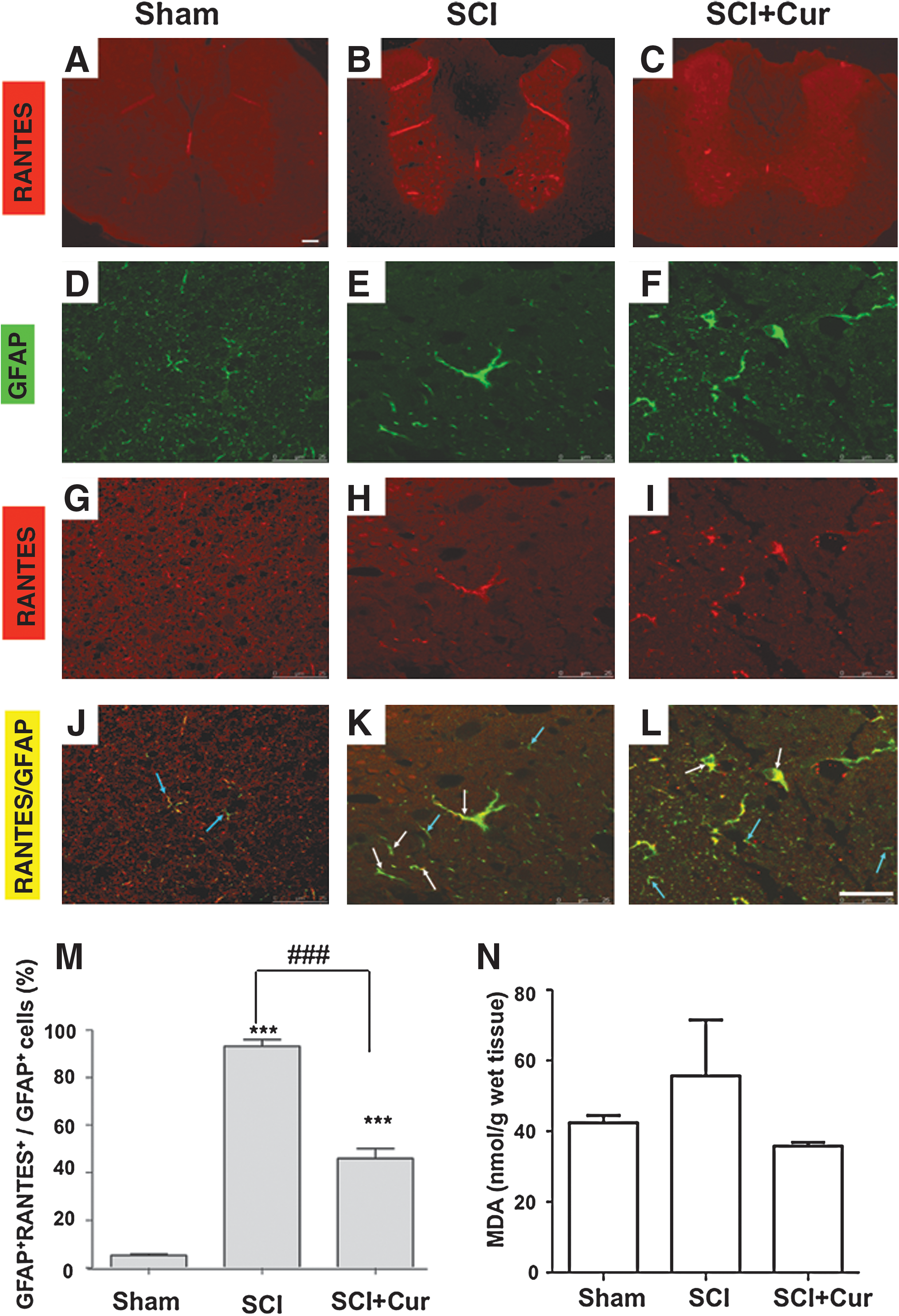
Curcumin reduces RANTES-positive astrocytes and lipid peroxidation in injured spinal cord. Representative photomicrograph of RANTES-stained sections from the spinal cord of (
Double immunofluorescent staining was subsequently performed on spinal cord sections obtained on the second day following SCI to evaluate the effects of curcumin on RANTES level in reactivated astrocytes. Simultaneous visualization of RANTES (Fig. 2 G–I) and reactivated astrocytes labeled with GFAP (Fig. 2D–F) revealed that the RANTES immunoreactivities were increased predominantly on the GFAP-positive astrocytes in SCI and SCI+Cur group (white arrow in Fig. 2 K, L). In comparison, sham control group exhibited sparse RANTES-positive astrocytes (Fig 2J). The inhibitory effect of curcumin treatment on astrocyte-secreted RANTES was observed as a morphologic change (Fig 2H, I) and as a reduction in the number of GFAP+-RANTES+ cells. The injury-increased perinuclear stain of RANTES immunoreactivities on astrocytes could be derived from astrocytes per se or from other cells, and could bind to astrocytes in an autocrine or paracrine manner. Quantitative analysis of the GFAP+-RANTES+ cells in the dorsal column white matter at the epicenter of injured spinal cord, which has been proven most extensively injured in acute SCI (Agrawai and Fehlings, 1997), further shows that there was a markedly increased number of GFAP+-RANTES+ cells in both the SCI and SCI+Cur groups compared to the control group. Whereas both the SCI and SCI+Cur groups exhibited a significantly higher percentage of GFAP+-RANTES+ cells when compared to the control group (p<0.001), rats with SCI treated with curcumin showed a significantly lower percentage when compared to rats with SCI that did not receive treatment (p<0.001) (Fig 2M). The mean percentage of astrocytes expressing RANTES was 5.18±1.12 for the sham group, 93.12±4.92 for the SCI group, and 45.99±7.13 for the SCI+Cur group.
Curcumin reduced lipid peroxidation in injured spinal cord
Curcumin is a lipid radical scavenger. Therefore, we further examined whether curcumin could reduce lipid peroxidation in the spinal cord 24 h after SCI. The result shows that the impact injury we performed in rats moderately increased MDA level, which was reduced by the 40mg/kg i.p. curcumin treatment (Fig. 2N).
Curcumin attenuated RANTES and iNOS expression in primary cultured astrocytes after LPS challenge
Because the RANTES immunoreactivities in injured spinal cord were highly co-localized with GFAP-positive astrocytes and were regulated by the curcumin treatment, we further used primary cultured astrocytes to investigate whether reactivation of astrocytes using inflammatory agent in culture could induce RANTES expression and secretion, and how curcumin could affect this response. Primary cultured astrocytes were challenged with LPS to mimic conditions of astroglial reactivation following CNS injury. To evaluate the effects of curcumin on RANTES and iNOS mRNA expression in reactivated astrocytes, we incubated 7 DIV astrocyte cultures with 1 μg/ml LPS alone, or with 1 μg/ml LPS with 1 μM curcumin (LPS+Cur) for 24 h, and conducted real-time PCR. We found that while both RANTES and iNOS mRNA expression were almost absent in the control group, both were significantly elevated in the LPS-challenged group. LPS-reactivated astrocytes treated with curcumin showed markedly decreased RANTES and iNOS mRNA expression when compared to LPS-reactivated astrocytes without curcumin treatment (p<0.001) as analyzed by quantitative real-time RT-PCR (Fig. 3 A, B). The mean RANTES mRNA level was 1.09±0.55 for the control group, 2777.48±213.39 for the LPS group, and 1512.21±168.24 for the LPS+Cur group. The mean iNOS mRNA level was 1.00±0.13 for control, 48220.75±1837.34 for LPS, and 24834.24±4223.34 for LPS+Cur.
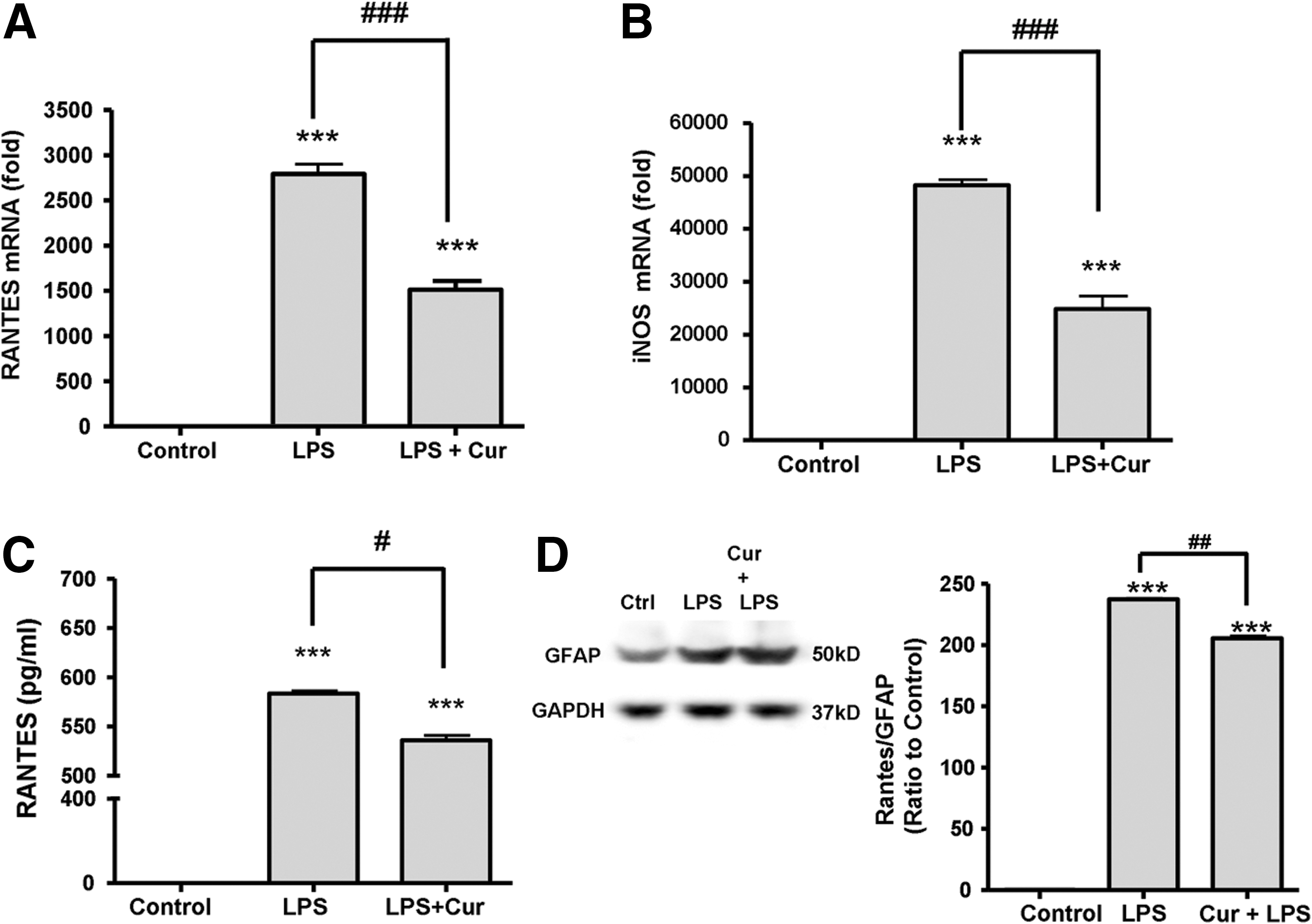
Curcumin downregulated (
Because RANTES is secreted from astrocytes to affect neuronal survival, we performed ELISA to evaluate the effects of curcumin on the protein level of RANTES in the medium of primary cultured astrocytes challenged with LPS. The ACM of the control, LPS, and LPS+Cur group with 1: 200 dilution was obtained for ELISA analysis. Figure 3C shows that RANTES level in the medium of LPS-induced reactivated astrocytes was more abundant than in the control group (p<0.001), and that curcumin treatment resulted in a significant decrease of RANTES protein in the LPS-conditioned astrocyte medium (p<0.05). However, curcumin did not reduce LPS-induced GFAP expression as demonstrated by immunoblotting (Figure 3D, left panel). Further analysis shows that the amount of RANTES in relation to GFAP is significantly lower in the curcumin+LPS group as compared with the LPS group (Figure 3D, right). These in vitro results—that curcumin attenuated robust induction of mRNA and protein expression of RANTES in LPS-reactivated astrocytes without affecting GFAP expression—corroborates the in vivo animal findings.
Curcumin reduced neurotoxicity of reactivated ACM
We further examined whether the conditioned medium obtained from LPS-reactivated astrocytes with or without curcumin treatment have differential effects on neuronal survival. Cortical neurons were incubated with these two ACMs for cell viability and death assays. The results in Figure 4 show that when neurons were cultured in ACM conditioned with LPS 1 μg/ml, LDH release was elevated (Fig. 4A) and MTT was significantly decreased (Fig. 4B) when compared to the ACM-control. Furthermore, cortical neurons incubated with LPS/curcumin-conditioned ACM showed a lower LDH level and a higher MTT level when compared to cortical neurons incubated with LPS-conditioned ACM (p<0.05). Therefore, LPS/curcumin-conditioned ACM is less neurotoxic than the LPS-conditioned ACM.

Curcumin-attenuated neurotoxicity of reactive astrocyte-conditioned medium (ACM). Cultured 10 days-in-vitro, cortical neurons were incubated for 24 h with ACM alone (ACM-control), ACM+curcumin (ACM-Cur), ACM+LPS (ACM-LPS), and ACM+LPS+curcumin (ACM-LPS+Cur). (
Knockdown of RANTES expression in LPS-reactivated astrocytes enhanced neuronal survival
Based on the observation that curcumin can reduce the expression of RANTES in reactivated astrocytes following injury in vivo and in vitro, we further asked whether curcumin reduced the neurotoxicity of LPS-conditioned ACM by its inhibitory effect on RANTES expression in reactivated astrocytes. We performed a loss-of-function study using siRNA to knock down the expression of RANTES in primary cultured astrocytes. As assessed by real-time PCR and ELISA analysis, the knockdown efficiency of RANTES siRNA (siRANTES) in cultured astrocytes was ∼ 85% compared to the scrambled RNA-transfected control group (Fig. 5A). Furthermore, RANTES mRNA expression and protein secretion of LPS-reactivated astrocytes in siRANTES- transfected culture was significantly decreased when compared to the scrambled RNA-transfected group (p<0.01 and p<0.001 respectively) (Fig. 5B). We next cultured 10 DIV cortical neurons with LPS-conditioned ACM. ACMs were prepared from astrocytes with siRANTES or scrambled RNA transfection. We found that cortical neurons incubated in LPS-conditioned ACM showed significantly elevated LDH levels when compared to ACM untreated with LPS (p<0.001) in the scrambled RNA-transfected group (Fig. 5C). In comparison, neurons cultured with ACM from siRANTES-transfected astrocytes treated with LPS showed significant decrease in LDH release compared to neurons treated with ACM from scrambled RNA-transfected astrocytes treated with LPS. These findings indicate that knockdown of RANTES expression in LPS-reactivated astrocytes significantly decreased the neurotoxicity of the ACM to cortical neurons. Therefore, suppression of RANTES release from LPS-reactivated astrocytes by means of either siRNA knockdown or curcumin treatment is effective in rendering the neuronal injury caused by the injury-stimulated astrocytes.
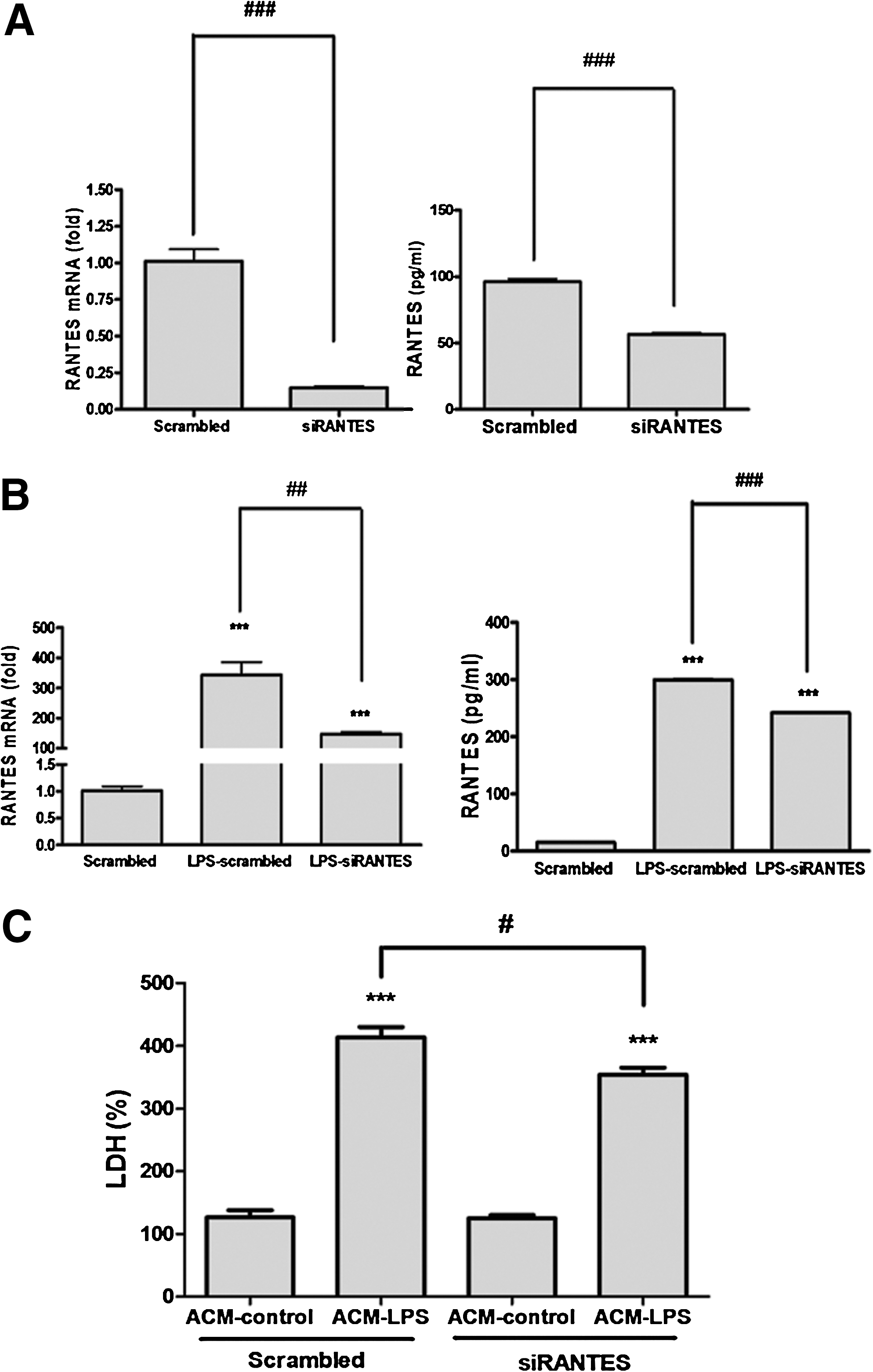
Knockdown of RANTES expression in reactivated astrocytes provided neuroprotection. Primary cultured astrocytes were transfected with siRANTES or scrambled RNA for 5 h in the Lipofectamine-containing medium, and recovered in the culture medium for another 67 h. The mRNA and protein expression of RANTES in astrocyte without (
Discussion
In this study, we demonstrated that curcumin conferred significant benefits in both a rat model of SCI and in a cell culture model of astrocyte reactivation. We showed that both SCI and LPS resulted in upregulated RANTES and iNOS mRNA expression. Our findings indicated that: 1) intraperitoneal administration of curcumin at 40 mg/kg attenuated RANTES and iNOS mRNA expression and decreased the number of RANTES-positive reactivated astrocytes, accompanied by preventing lipid peroxidation in vivo 24 h following SCI; 2) curcumin attenuated RANTES and iNOS expression, RANTES secretion, but not GFAP expression in LPS- reactivated astrocytes; 3) neuronal survival was enhanced in cortical neurons incubated with ACM conditioned with LPS and curcumin; and 4) upregulated RANTES expression in neuroinflammation is detrimental, by evidence that knockdown of RANTES expression in LPS-reactivated astrocytes reduced the ACM-induced neuronal death. These findings corroborate our previous study (Lin et al., 2011), and suggest that curcumin attenuated the degree of secondary inflammation, possibly by virtue of its ability to attenuate protein expression of RANTES in reactivated astrocytes, and its ability to attenuate both RANTES and iNOS mRNA expression at the site of injury.
Chemokines such as RANTES are a family of proinflammatory cytokines involved in leukocyte/lymphocyte trafficking, homeostasis, inflammation, and development of the immune system. Elevated chemokine expression of MCP-1, RANTES/CCL5, and MIP-1b has been characterized in sites of cortical stab wounds in rats and mice (Berman et al., 1996; Glabinski et al., 1996). Originally identified as a T-cell specific gene, RANTES/CCL5 has been demonstrated to be significantly upregulated, and associated with T-lymphocyte activation in SCI (Schall et al., 1988). RANTES/CCL5 mRNA expression remains upregulated for as long as 24 h after SCI in spinal cord microvascular endothelial cells, and precedes or parallels chronic activation of T-lymphocytes. Chronic T-cell activation subsequently contributes to neurodegeneration (Jones et al., 2005) and inhibits repair of injured tissues without directly influencing neuroglial survival (Jones et al., 2002). Therefore, the curcumin modulation of injury-induced RANTES release might attenuate multiple immune responses that contribute to the chronic neurodegeneration following acute injury.
Despite evidence for the neurodegenerative effects of elevated RANTES mRNA expression and T-cell activation in SCI, neuroprotective effects of RANTES have also been described. Wynn (2004), using cDNA microarrays, found that a large number of RANTES-responsive genes in cultured neurons appeared to be involved in neuronal survival and differentiation. More recently, Tripathy et al. (2010) demonstrated that treatment with RANTES in primary cultures of cortical neurons enhanced neuronal survival. The paradoxical nature of chemokines and cytokines has been described as a double-edged sword: they protect and repair, but may also exacerbate chronic or excessive inflammation. With regard to RANTES, it has been suggested that several mechanisms shape the fate of T-cell function and how it is controlled by RANTES and other chemokines. Such mechanisms are probably related to the microenvironment of the injury (e.g., ischemia, apoptosis, demyelination, mitochondrial dysfunction, glutamate excitotoxicity), and the interplay with other chemokines, cytokines, and enzymes. It is likely that overexpression of inflammatory mediator iNOS and the induction of potent chemoattractants such as RANTES are complex and bidirectional. Luo et al. (2002) found that astrocytes respond to RANTES by producing more chemokine and cytokine transcripts, which results in a chemokine cascade and may have the effect of amplifying the inflammatory response.
In CNS injury, as stated previously, astroglial reaction begins 1 h after the injury. Microglial activation accompanies astrogliosis. The post-traumatic inflammatory response is largely characterized by mitochondrial dysfunction, activation of nitric oxygen species (NOS), and reactive oxygen species (ROS). The level of NO is increased in the injured spinal cord (Hamada et al., 1996), and NO produced by iNOS is known to contribute to apoptotic cell death and progression of tissue damage in SCI (Xu et al., 2006). Therefore, the present study showing that curcumin attenuates LPS-induced iNOS expression supports the potential for protecting the injured spinal cord against proinflammatory insults.
Curcumin has already been shown to inhibit IL-1β and TNF-α by inhibiting NF-κB in a number of inflammatory disease states (Kowluru et al., 2007; Moon et al., 2010). The NF-κB transcription factor is a family of transcription factors that regulates not only iNOS gene expression, but also a number of proinflammatory genes (Verma, 2004). It has been demonstrated that the murine RANTES promoter contains a single interferon regulatory factor and three putative NF-κB binding sites, and that TNF-α and interferon γ synergistically activate expression of RANTES (Lee et al., 2000). Both TNF-α and IL-1β have also been shown to exhibit neurotoxicity in cerebral ischemia in the presence of iNOS, whereas, in the absence of iNOS, both cytokines were neuroprotective (Stoll et al., 2000). Therefore, it is likely that curcumin alters the function of RANTES by attenuating NF-κB activation and the expression of proinflammatory mediators such as iNOS, thereby decreasing RANTES expression by astrocytes during neuroinflammation. Future studies are warranted to examine this putative mechanism in SCI.
In conclusion, we demonstrated that curcumin attenuated both RANTES and iNOS mRNA expression in an in vivo model of SCI and in a cell culture model of inflammatory stimuli-induced astrocyte reactivation, and found that curcumin could reduce the neurotoxicity of LPS-conditioned astrocyte medium. Our findings provide additional support of curcumin neuroprotection for the prevention or treatment of secondary inflammatory injury following SCI. Clinical studies are needed to determine the therapeutic potential of curcumin in the treatment of brain and spinal cord injuries.
Footnotes
Acknowledgments
This work was supported by grants from Ministry of Education, Aim for the Top University Plan, 98-wf-phd-02 and 99-wf-phd-02 from Taipei Medical University–Wan Fang Hospital, DOH100-TD-B-111-003 of the Center of Excellence for Clinical Trial and Research in Neuroscience, and NSC 99-2321-B-010−015, NSC99-2314-B-038-044-MY3 from the National Science Council, Taiwan. We thank Dr. Liang-Yo Yang for supporting the use of the NYU impactor, and Chia-Lin Shih for assisting the MDA assay.
Author Disclosure Statement
No competing financial interests exist.