
Abstract
In adult mammals, restoration of function after peripheral nerve injury is often poor and effective therapies are not available. Previously we have shown in mice that a peptide which functionally mimics the human natural killer cell (HNK)-1 trisaccharide epitope significantly improves the outcome of femoral nerve injury. Here we evaluated the translational potential of this treatment using primates. We applied a linear HNK-1 mimetic or a functionally inactive control peptide in silicone cuffs used to reconstruct the cut femoral nerves of adult cynomolgus monkeys (Macaca fascicularis). Functional recovery was evaluated using video-based gait analysis over a 160-day observation period. The final outcome was further assessed using force measurements, H-reflex recordings, nerve histology, and ELISA to assess immunoreactivity to HNK-1 in the treated monkeys. Gait deficits were significantly reduced in HNK-1 mimetic-treated compared with control peptide-treated animals between 60 and 160 days after injury. Better outcome at 160 days after surgery in treated versus control animals was also confirmed by improved quadriceps muscle force, enhanced H-reflex amplitude, decreased H-reflex latency, and larger diameters of regenerated axons. No adverse reactions to the mimetic, in particular immune responses resulting in antibodies against the HNK-1 mimetic or immune cell infiltration into the damaged nerve, were observed. These results indicate the potential of the HNK-1 mimetic as an efficient, feasible, and safe adjunct treatment for nerve injuries requiring surgical repair in clinical settings.
Introduction
T
We have recently introduced a novel approach for treatment of nerve injuries in mice using mimetics of glycans, carbohydrate moieties on protein or lipid molecules. Glycans are essential for the functional properties of the carrier proteins and increasing evidence indicates that these carbohydrates are beneficial for neural repair (Eberhardt et al., 2006; Gravvanis et al., 2007; Kleene and Schachner, 2004; Papastefanaki et al., 2007). The human natural killer (HNK) cell glycan (3′ sulfoglucuronyl β1,3 galactoside) known as HNK-1 epitope and expressed on different cell adhesion molecules (ffrench-Constant et al., 1986; Kruse et al., 1984, 1985; Löw et al., 1994; Schachner and Martini, 1995), has been associated with proper targeting of regenerating motor axons (Brushart, 1993; Martini et al., 1992). We have shown that application of a peptide that mimics the functional properties of the HNK-1 epitope (Simon-Haldi et al., 2002) to the injured femoral nerve of adult mice promotes functional recovery (Simova et al., 2006). Another glycan involved in nerve regeneration is α2,8 polysialic acid (PSA). This carbohydrate is carried by the neural cell adhesion molecule (NCAM; Rutishauser, 2008), and neuropilin-2 (Curreli et al., 2007), and is upregulated in axons and skeletal muscle fibers after nerve injury in adult rodents (Covault et al., 1986). Recently, we showed that the application of a PSA-mimic peptide leads to improved gait and better axonal remyelination after femoral nerve injury in mice (Mehanna et al., 2009).
Although encouraging, the beneficial effects of glycomimetics observed in mice do not necessarily provide a safe basis for clinical translation. We considered that the use of primates might provide stronger evidence for the efficacy and safety of a cyclic HNK-1 mimetic treatment after nerve lesion. The larger body size of primates and superior development of the motor and sensory cortices compared with mice makes them an experimental model that more closely resembles the pathology and time-course of recovery after nerve injury in humans (Kaas and Collins, 2003; Krarup et al., 2002). Also, since the HNK-1 epitope is the target for IgM autoantibodies in patients with demyelinating neuropathy (van den Berg et al., 1990), we considered the possibility that the HNK-1 mimetic could be immunogenic in primates, and thus compromise rather than enhance regeneration. Finally, the effects of a glycan mimic may differ among species because of the different structure and binding properties of endogenous carrier molecules and receptors, and species differences in carbohydrate expression (Kleene and Schachner, 2004). Our data provide evidence for the efficacy and safety of the HNK-1 mimetic in primates.
Methods
Mimic peptides
We used the cyclic hexapeptide c-(RTLPFS) derived from the peptide sequence TFQLSTRTLPFS, which has been discovered in phage display studies as a functional mimetic of the HNK-1 oligosaccharide (Bächle et al., 2006; Simon-Haldi et al., 2002). This hexapeptide shows a higher binding affinity to the HNK-1-specific antibody 412 than the original linear peptide, and is also a functional HNK-1 mimetic (Bächle et al., 2006; Simova et al., 2006). The cyclic peptide c-(RDTLPFS), which contains D-threonine instead of L-threonine, was used as control. Due to the different turn geometry of the peptide this substitution results in a loss of function. In vitro experiments have shown that this control peptide had no effect on motoneuron outgrowth or survival in vitro and nerve regeneration in vivo (Bächle et al., 2006; Simova et al., 2006).
Animals and experimental design
The experiments were performed at the Yunnan Laboratory Primate Inc. (former Yunnan National Laboratory, Primate Centre of China), Jinghong, Xishuangbanna, People's Republic of China, in accordance with the local regulations and the European Community and National Institutes of Health guidelines for care and use of laboratory animals. Data from functional assays, nerve histology, and ELISA assays were obtained at the Zentrum für Molekulare Neurobiologie Hamburg. Ten male cynomolgus monkeys (Macaca fascicularis, 6 years of age, body weight 4.2–5.0 kg) were used in experiments involving nerve surgery. Two of these animals were used in a pilot experiment to test and optimize the surgical reconstruction of the femoral nerve and establish an approach for quantitative assessment of locomotor deficits induced by the femoral nerve palsy, procedures that, to the best of our knowledge, have not been previously reported in primates. The main experimental group consisted of 8 macaques. After initial video recording, the animals were subjected to transection and surgical repair of the femoral nerve (Fig. 1B–D). Immediately after the surgical reconstruction, the HNK-1-glycomimetic peptide was applied into the silicone cuff used as a nerve conduit in 4 animals; the other 4 animals received control peptide (Fig. 1D). Following repeated video recordings over a period of 160 days, force measurements and electromyographic (EMG) recordings were performed, and after the animals were euthanized using an overdose of anesthetic, tissue samples were collected (Fig. 1A). In addition to these experimental animals, 4 age-matched male macaques not subjected to any previous treatment were used to obtain normal EMG recordings and blood samples. All 10 animals used for surgery were kept in one large cage (6×4×3 m) and received a standard diet including vegetables, fruits, and standard laboratory pellets. Health monitoring and anesthesia were performed by an experienced veterinarian. Excessive grooming, scratching, or biting of the denervated limb, and signs of paresthesias or neuropathic pain were not seen in any of the operated macaques during the observation time period.
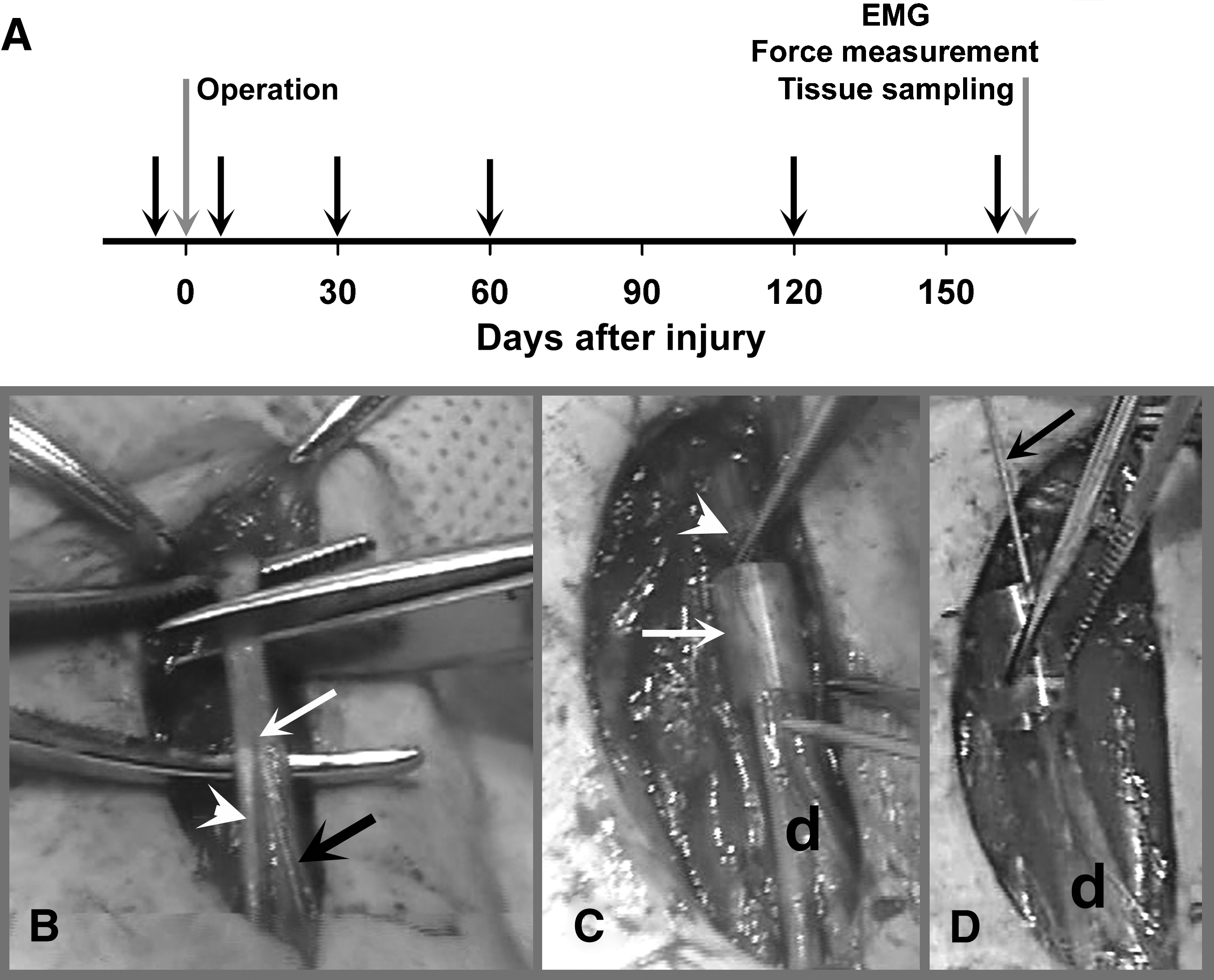
Experimental flow and femoral nerve surgery. Eight adult male cynomolgus macaques (Macaca fascicularis) were subjected to transection and repair of the left femoral nerve (gray arrow at 0 days in
Femoral nerve surgery
The animals were anesthetized by intramuscular injection of ketamine hydrochloride (25 mg/kg body weight; Jiangsu Hengrui Medicine Co., Ltd., Lianyungang, China). The left femoral nerve was exposed and nerve transection performed at a distance of about 10 mm proximal to the bifurcation of the nerve (Fig. 1B). The cut ends of the nerve were inserted into a silicone tube (6 mm long, 3.35/4.65 mm inner/outer diameter; AMT Medizintechnik, Düsseldorf, Germany; Fig. 1C) and fixed with two epineural 9-0 nylon stitches (Ethicon, Norderstedt, Germany) so that a 2-mm gap was present between the proximal and distal stumps. The tube was filled with phosphate-buffered saline (PBS), pH 7.3, containing scaffold peptide that forms a gel matrix support (0.25% PuraMatrix Peptide Hydrogel; 3DMatrix, Cambridge, MA) supplemented with either control peptide or HNK-1 mimetic peptide (200 μg/mL). After peptide application, the skin wound was closed with 4-0 sutures (Ethicon). All experiments and data analyses were performed blindly with regard to type of peptide application. Following the femoral nerve surgery, the sartorius nerve was resected in all animals by excising a 4-cm segment in the thigh. The proximal nerve stump was ligated and sutured to the proximal musculature using 4-0 sutures to prevent regeneration. This procedure was performed in order to eliminate the possibility of functional compensation of the femoral nerve injury by use of the sartorius muscle as a knee extensor.
Analysis of locomotion
The animals were customized, in 2–3 short sessions on two successive days, to guided out-of-cage walking using a collar and handles. Thereafter, left- and right-side views of 4–6 walking trials were video recorded at 25 frames per second using a Panasonic VDR-D250 camera (Panasonic Deutschland, Hamburg, Germany) prior to operation (0 days), and 7, 30, 60, 120, and 160 days after nerve injury. The video sequences were examined with VirtualDub software, a video capture/processing utility written by Avery Lee (free software available at
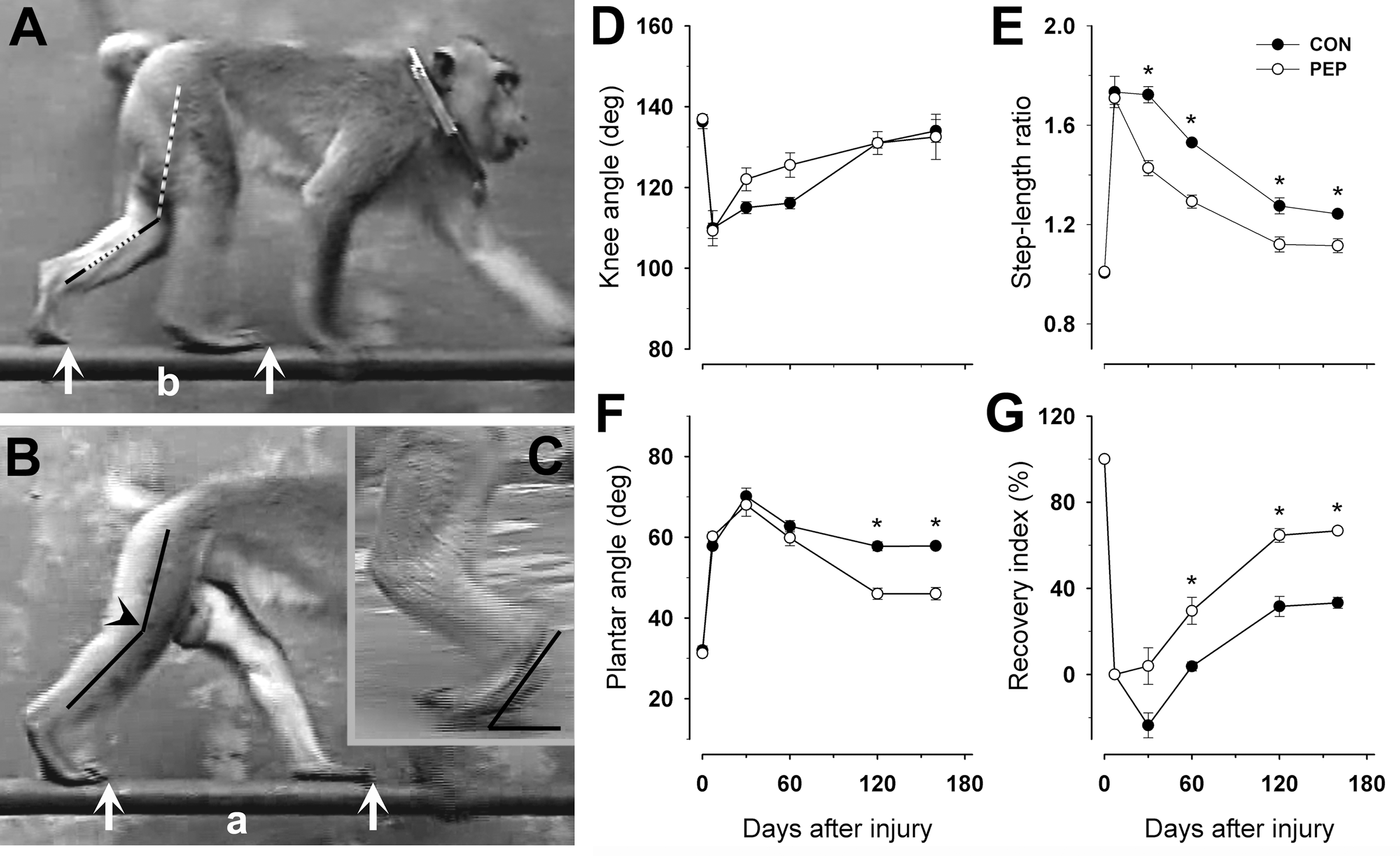
The human natural killer cell-1 (HNK-1) glycomimetic peptide improves gait after nerve repair. (
Knee angle at contralateral foot placement, step-length ratio, and plantar angle at mid-swing were measured using single video frames from sequences in which the animals walked in a straight line at constant speed. The knee angle (Fig. 2B) was measured when the contralateral foot was placed on the ground (beginning of the stance phase). The step-length ratio was calculated as ratio of the lengths of two successive steps (Fig. 2A and B). The plantar angle was defined as the angle between the plantar (sole) surface and the horizontal plane, and was measured in video frames showing the leg in mid-swing (Fig. 2C). For each parameter, 4–8 measurements were made per animal and time point, and the average was used to calculate mean group values. No differences were noticed between results based on videos of gait inside and outside of the cage.
As a measure of functional recovery at different time points after injury, we calculated the overall recovery index (RI), which is a mean of the RI for 2 or 3 parameters as indicated in the text. The index for each parameter is calculated in percent as:
where Ypre, Yden, and Yreinn are values prior to operation, during the state of denervation (7 days after injury), and at given time point thereafter, respectively (Irintchev et al., 2005).
Force measurements
The deeply anaesthetized animals were placed seated on a table and the trunk was kept in an upright position by an animal caretaker. The knees were placed slightly beyond the table's edge so that the lower legs hung free. A weight of 375 g was attached to one ankle using an adhesive band (Fig. 3A and B). An investigator applied supramaximal tetanic (50 Hz) stimuli transcutaneously to the proximal femoral nerve using a pair of brass electrodes (110 mm diameter; Fig. 3A and B) until the leg was maximally stretched. During stimulation, lifting of the knee was prevented by the investigator by fixing it with two fingers placed lateral to the patella (Fig. 3A and B). For each leg, the stimulations were repeated six times and recorded using a Panasonic VDR-D250 camera. The video camera was positioned at a distance of approximately 2 m from the leg at the level of the knee and oriented perpendicularly to the leg axis. Note that the photographs in Figure 3A and B aim to show the experimental set-up, and were taken using a position of the camera different from that used during video recording. For each leg, the angle between the lower leg (knee-ankle) axis and the vertical line was measured before each stimulation (Fig. 3A), and upon maximal contraction during that stimulation (Fig. 3B), using single video frames. Force was estimated as the difference between the two angles (in degrees). The three best results (largest difference between the angles) were used to calculate the mean value per leg, and the mean value for the left (injured) leg was normalized to the value for the contralateral uninjured leg.
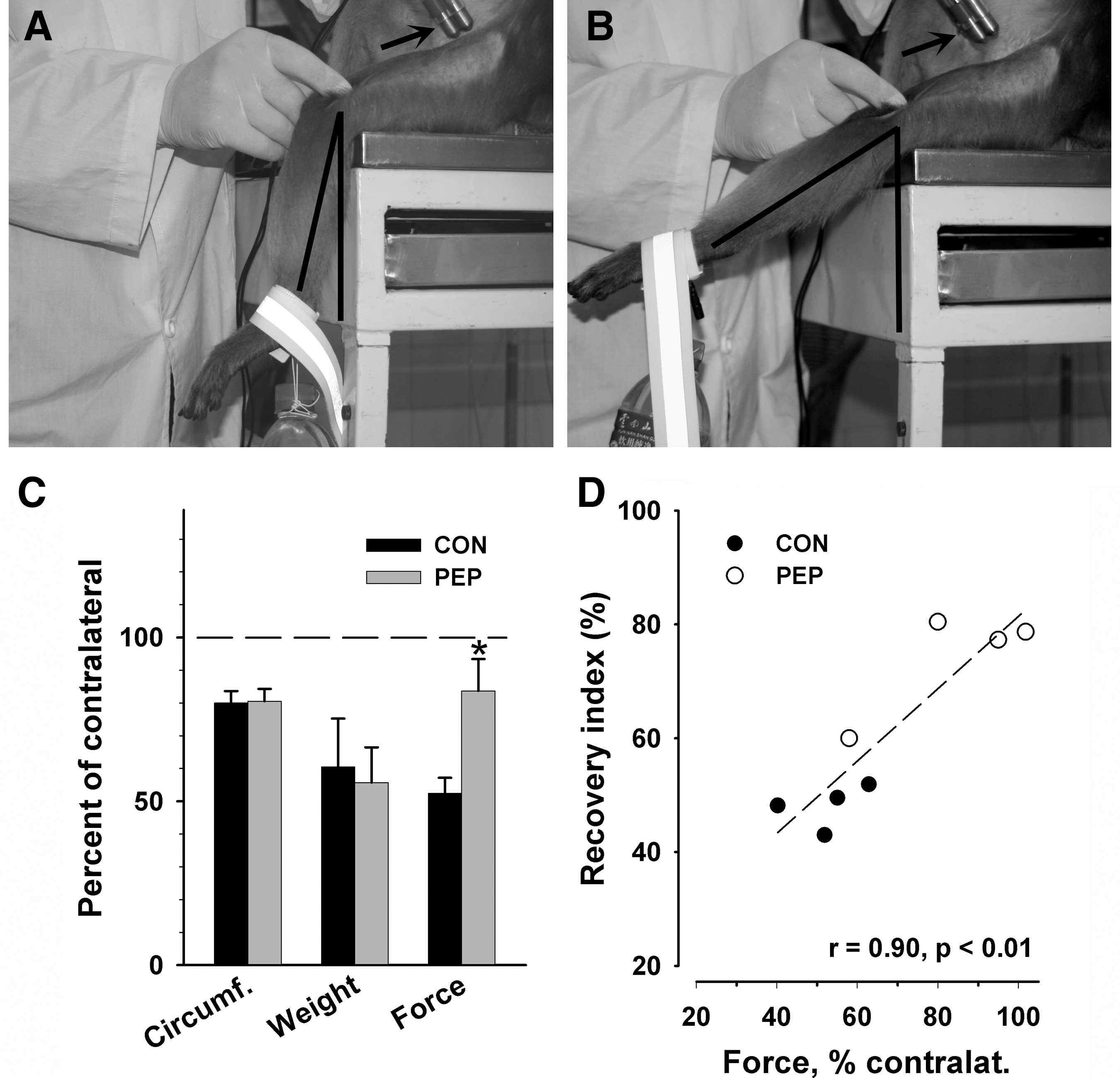
Human natural killer cell-1 (HNK-1) glycomimetic treatment leads to an increase of muscle force. (
Electromyography
The animals were anesthetized using one-half of the dose required for surgery and fixed in a standard chair for restraining primates. A ground surface electrode (brass, diameter 110 mm) was fixed to the wrist using adhesive tape. The femoral nerve was stimulated proximal to the injury site using a bipolar electrode (Fig. 3A and B). The point of stimulation was defined as the point closest to the nerve conduit at which reproducible reflex responses could be elicited. The stimulation was performed using an insulated stimulator (Model 2100 Isolated Pulse Stimulator; A-M Systems, Calsborg, WA) and bipolar electrical pulses of 0.2 msec duration. The active recording surface electrode (Ag/AgCl, EL208WS; WPI, Sarasota, FL) was attached with an adhesive disk (ADD208; WPI) to the skin covering the rectus femoris muscle, at the border between the mid- and distal third of the muscle head. The reference electrode was fixed to the skin of the contralateral leg. The signal from the recording electrode was transmitted to a differential amplifier (P55 General Purpose AC Preamplifier; Grass Technologies, West Warwick, RI), magnified (100×), and filtered (300-Hz high-pass and 1-kHz low-pass filters). The signals were digitized using an A/D converter (ADC42; Pico Technology, Cambridgeshire, U.K.) and PicoScope 5 data acquisition software (Pico Technology). To measure maximum reflex responses, electrical pulses of different stimulus intensity were delivered at a frequency of 0.1 Hz to elicit maximum M- and H-waves (M-max and H-max, respectively). To estimate rate depression of the H-reflex, stimulus intensity was increased until maximal and stable H-responses were elicited. Thereafter, stimulation continued at the defined level at frequencies of 0.1, 0.3, 0.5, 1, 2, 5, and 10 Hz. Six consecutive responses were recorded at each frequency. The amplitudes of M- and H-waves were measured as peak-to-peak values, averaged (excluding the first response at each frequency), and used to calculate H:M ratios. The latencies of the responses were measured as the time elapsed between the trigger and the peak of each waveform.
Measurement of muscle circumference and weight
Muscle mass and circumference were measured during the terminal experiment 163–165 days after injury. The lateral borders of the quadriceps muscle in the middle of the thigh length axis were palpated, and the circumference of the enclosed thigh segment was measured using a measuring tape. After dissection, the wet weight of the rectus femoris muscle was measured using a laboratory balance.
Morphometric analyses of regenerated nerve branches
Femoral nerves were dissected, fixed in 4% formaldehyde, and post-fixed in 1% osmium tetroxide in 0.1 M sodium cacodylate buffer, pH 7.3, for 1 h at room temperature. Then they were dehydrated and embedded in resin according to standard protocols. Transverse 1-μm-thick sections from the motor nerve branch were cut at a distance of approximately 3 cm distal to the bifurcation and stained with 1% toluidine blue/1% borax in distilled water. Using random sampling, digitized images were obtained on a bright field microscope using a 100×oil objective (Axioskop; Zeiss, Jena, Germany). Fiber and axon diameters were measured and fiber density was estimated using AxioVision 4.8 software (Zeiss), and the g-ratio, defined as the ratio of axon to fiber diameter, was calculated for each nerve fiber.
Determination of anti-HNK-1 antibody titer in sera by ELISA
Concentrations of IgGs and IgMs and of total proteins in the sera from untreated non-injured monkeys, and from injured and control peptide- or glycomimetic-treated monkeys were determined using the BCA test (Perbio; Thermo Fisher Scientific, Bonn, Germany), and anti-IgG (Fc specific; Sigma-Aldrich, Taufkirchen, Germany), and anti-IgM (Jackson ImmunoResearch, Dianova, Hamburg, Germany) antibodies. All sera contained comparable amounts of proteins and overall concentrations of antibodies (both IgG and IgM). To determine the anti-HNK-1 antibody titer in sera from control peptide- and HNK-1 glycomimetic peptide-treated monkeys, 384-well PolySorp plates with high binding surface were coated with sera (100 and 500 μg total serum protein/well), BSA (100 μg/well and 500 μg/well), or, for control, anti-HNK-1 monoclonal antibody 412 (1 μg/well; Kruse et al., 1984) or anti-PSA monoclonal antibody 735 (1 μg/well; Frosch et al., 1985) in PBS overnight. The coated plates were then washed once with PBS, blocked for 3 h with blocking buffer (1% BSA in PBS; 50 μL per well) at room temperature, and washed 2×with PBS. Biotinylated HNK-1 glycomimetic peptide or biotinylated PSA glycomimetic peptide (2.5 μg/well; in PBS) were added and incubated for 2 h at room temperature. After five washing steps with PBS, horseradish peroxidase (HRP)-coupled Neutravidin (Perbio; 1:2500 in PBS; 25 μL/well) was added to detect the captured glycomimetics, and the wells were incubated for 1 h at room temperature. Next, the wells were washed again 5×with PBS and orthophenylene diamine (OPD; 0.5 mg/mL) was added as substrate for the HRP. The reaction was stopped after 10 min by adding 25 μL 2.4 M sulfuric acid in each well and the absorbance was measured at 490 nm in a microplate reader (μQuant; BioTek, Winooski, VT). The absorbance in wells coated with anti-HNK-1 antibody 412 served as positive control, whereas wells coated with anti-PSA antibody 735 or BSA were used to define background reactivity. All probes were tested in triplicate.
Statistical analysis
All numerical data are presented as group mean values with standard errors of the mean (SEM). Comparisons between groups were performed with the SPSS 15 software package (SPSS, Chicago, IL) as indicated in the text and figure legends. In all cases in which parametric tests were used, the data conformed to the requirements for normality and equal variance. Sigma Plot 11 software (SPSS) was used for regression analyses. The threshold value for acceptance of differences was 5%.
Results
Femoral nerve injury causes long-lasting gait deficits which are alleviated by HNK-1 glycomimetic application
We performed transection and surgical repair of the left femoral nerve of adult male macaques (Fig. 1A–C), and applied HNK-1 glycomimetic peptide (n=4) or control peptide (n=4) into the silicone cuff used as a nerve conduit (Fig. 1D). Video recordings of walking trials were performed prior to surgery and 7–160 days thereafter (Fig. 1A). Immediately after femoral nerve injury and recovery from the anesthesia, the animals experienced a severe gait disturbance due to loss of function of the quadriceps muscle, the main extensor in the knee joint. As soon as the injured leg had to support the body weight (stance phase), the knee joint collapsed resulting in a near fall when walking on the side-wall bars in the cage. Surprisingly, as soon as 1 day after injury, the animals overcame this disability to a degree that made their walking in the cage appear normal to the eye. Analysis of video recordings revealed that the macaques compensated for the quadriceps muscle dysfunction using “trick movements” similar to humans with nerve lesions (Parry, 1970). The stability of the knee joint was increased by internal rotation of the leg (not shown) and the weight-bearing (stance) phase was shortened by asymmetric steps, a large one with the injured leg and a short one with the intact leg (Fig. 2A and B). Despite these gait adaptations, measurements of the knee angle 7 days after injury revealed knee instability (Fig. 2A and B). Compared with pre-operative values (0 days), the angle was decreased from about 140° to 110° (−20%, Fig. 2D). More pronounced was the alteration in the step lengths. While prior to operation the left-to-right step-length ratio was 1, it increased to 1.7 at 7 days (+70%, Fig. 2E). A third measurable change in gait after injury was the increase of the plantar angle (the angle between the plantar surface and the ground plane), from about 30° to 60° at 7 days (+100%, Fig. 2C and F). This alteration was a likely consequence of the increased length of the step with the injured leg, and thus the need to lift the knee joint higher than normal during the swing phase. The gait disturbances, as estimated by the three parameters, were similar in animals treated with control peptide and HNK-1 mimetic 7 days after injury (Fig. 2D–F). Between 7 and 160 days, the knee angle gradually recovered to pre-operative values, and no difference between the groups was found at any time point (Fig. 2D). The other two parameters, step-length ratio and plantar angle, did not reach control values even 160 days after injury, but recovery was better in mimetic-treated than in control peptide-treated animals (Fig. 2E and F). Thus, it appeared that knee instability, as measured by the knee angle, was compensated by longer steps with the injured leg and more pronounced lifting of the knee during swing in control compared with mimetic-treated animals. We also calculated individual recovery indices (degree of return of function, 100% indicating complete recovery) for each of the 3 parameters, and using them we calculated the mean of 2 or 3 of these recovery indices per animal, designated overall recovery indices. When overall recovery was estimated using the two parameters that showed differences between the groups, the step-length ratio and the plantar angle (Fig. 2E and F), a better recovery in mimetic- versus control peptide-treated animals was found at 60 days after injury, and maximum recovery at 160 days reached 67% with mimetic application, compared with only 33% in the control group (Fig. 2G). Also, overall recovery indices calculated for all three parameters used for gait analysis (Fig. 2D–F) showed a higher degree of recovery in mimetic- than in control peptide-treated animals at 160 days after injury (74±5% versus 48±2%, respectively, p<0.05 by t-test). The distribution of individual values of these recovery indices is seen in Figs. 3D and 4E and F.

Glycomimetic application enhances the H-reflex. (
Force of reinnervated muscles is improved by mimetic application
The conclusion that recovery of muscle function was deficient in both experimental groups was supported by the finding of muscle atrophy at the end of the observation time period. Compared with the intact contralateral muscles, in both experimental groups the outer circumference of the quadriceps muscle was reduced by 20%, and the wet weight of the rectus femoris muscle was about 40% lower (Fig. 3C). The finding of similar degrees of atrophy in the two groups, despite differences in gait, is explainable by the low accuracy of the measurements (circumference), and large inter-individual variability in muscle weight (note the large error bars for this parameter in Fig. 3C). The weight of the rectus femoris muscle might not be representative of the whole mass of the quadriceps, which is comprised of four heads: rectus femoris, vastus lateralis, medialis, and intermedius. It is conceivable that the mass of each of these heads is prone to inter-individual variability even in intact animals, and after nerve injury, the degree of reinnervation, and thus alleviation of muscle atrophy, might not be identical in the 4 heads of one muscle.
Despite similar weight of the rectus femoris muscle in the two experimental groups, estimates of muscle force of the whole quadriceps muscle (Fig. 3A and B) revealed a significantly better function in mimetic- than in control peptide-treated animals (Fig. 3C). Upon supramaximal tetanic stimulation of the proximal stump of the femoral nerve, control animals could lift a weight fixed to their ankle to only about 50% of the height reached by the contralateral intact leg. The performance of the injured leg was significantly better in mimetic- than in control peptide-treated animals (more than 80% of contralateral). Thus, in agreement with the results of the gait analysis, force measurements showed a positive effect of the mimetic treatment. Moreover, regression analysis revealed a significant covariation between recovery of gait (the overall recovery index calculated for the three parameters shown in Fig. 2D–F) and muscle force (Fig. 3D), suggesting that variability in muscle strength underlies the inter-individual variability in gait.
Enhanced H-reflex responses in mimetic-treated animals
At the end of the observation time period, we analyzed the Hoffmann (H-) reflex to assess alterations in the excitability of the homonymous stretch reflex. In uninjured age-matched male monkeys, electrical stimulation of the femoral nerve produced typical EMG responses at baseline stimulation frequency (0.1 Hz), consisting of a short-latency M- (muscle) wave and a long-latency H-wave elicited by stimulation of motor axons and Ia afferents, respectively (data not shown). Responses of similar amplitudes and latencies were observed in the intact contralateral legs of animals with nerve lesions (Fig. 4A). Compared with intact contralateral nerves, the amplitudes of the H-waves after stimulation of regenerated nerves proximal to the injury site in control peptide-treated animals were reduced by 70%, while the M-wave amplitudes were normal (Fig. 4B and D). Thus, the H:M ratio, a measure of reflex excitability, was reduced to less than one-third of normal (Fig. 4D). In HNK-1 mimetic-treated animals, the M-wave amplitude elicited upon stimulation of the injured nerve was also no different from normal (Fig. 4D). However, the H-wave responses were strongly enhanced, by more than twofold, compared with injured nerves of control animals, and the H:M ratio was proportionally increased (Fig. 4C and D). These results indicate that the mimetic treatment causes changes in the reflex pathway that favor motoneuron recruitment. As indicated by a positive linear correlation between individual H-wave, but not M-wave amplitudes on the one hand, and recovery indices on the other hand (Fig. 4E and F), reflex excitability appears to be a factor influencing the degree of locomotor recovery. Upon incremental increases of the stimulation frequency on the injured side from baseline (0.1 Hz) to a maximum of 10 Hz, the H-wave amplitude declined insignificantly in both groups of animals, indicating low rate depression of the H-reflex. However, at all frequencies used, the H:M ratio was significantly higher in mimetic- than in control peptide-treated animals (data not shown). In addition to the H-wave amplitude, the glycomimetic treatment showed an effect on the latency of the H-reflex. In control animals, the peak of the H-wave on the operated side appeared with a delay of 1 msec after the stimulation compared with the intact contralateral side (8.6±0.15 versus 7.6±0.41 msec, p<0.05 by t-test). In mimetic-treated animals, the latency of the H-reflex elicited through the injured femoral nerve (7.7±0.14 msec) was significantly reduced compared with control regenerated nerves, and was thus similar to the contralateral normal value (7.6±0.43 msec, p>0.05 by t-test).
Superior axonal regeneration after mimetic treatment
Analysis of sections from the quadriceps nerve branches distal to the site of injury at 165 days (Fig. 5A and B) revealed no differences in fiber densities between the experimental groups (3527±1194 and 4182±1565 fibers per mm2 in control peptide- and glycomimetic peptide-treated monkeys, respectively), suggesting no difference in the total number of myelinated nerve fibers. However, regenerated axons had significantly larger diameters in mimetic- than in control peptide-treated animals (+26%, Fig. 5C). A similar difference between the groups was found for fiber diameters (axon plus myelin sheath, Fig. 5C). Accordingly, the g-ratios (axon to fiber diameters) were similar in the two groups (Fig. 5C). These results indicate that the mimetic treatment leads to regeneration of thicker axons without affecting the degree of axonal remyelination. Since axon potential propagation is proportional to axonal thickness, the finding of larger axonal diameters likely explains the significant reduction, as compared with control animals, of the H-wave latency in mimetic-treated macaques. This notion is supported by the finding of a significant negative correlation between individual latency values and mean axonal diameters in the same animals (r=−71, p=0.047 by ANOVA for regression). A positive correlation was found between individual recovery indices and mean axonal diameters (r=0.64, p=0.08 by ANOVA for regression). These correlations suggest links between axonal thickness on the one hand, and degree of locomotor recovery and reflex latency on the other hand.
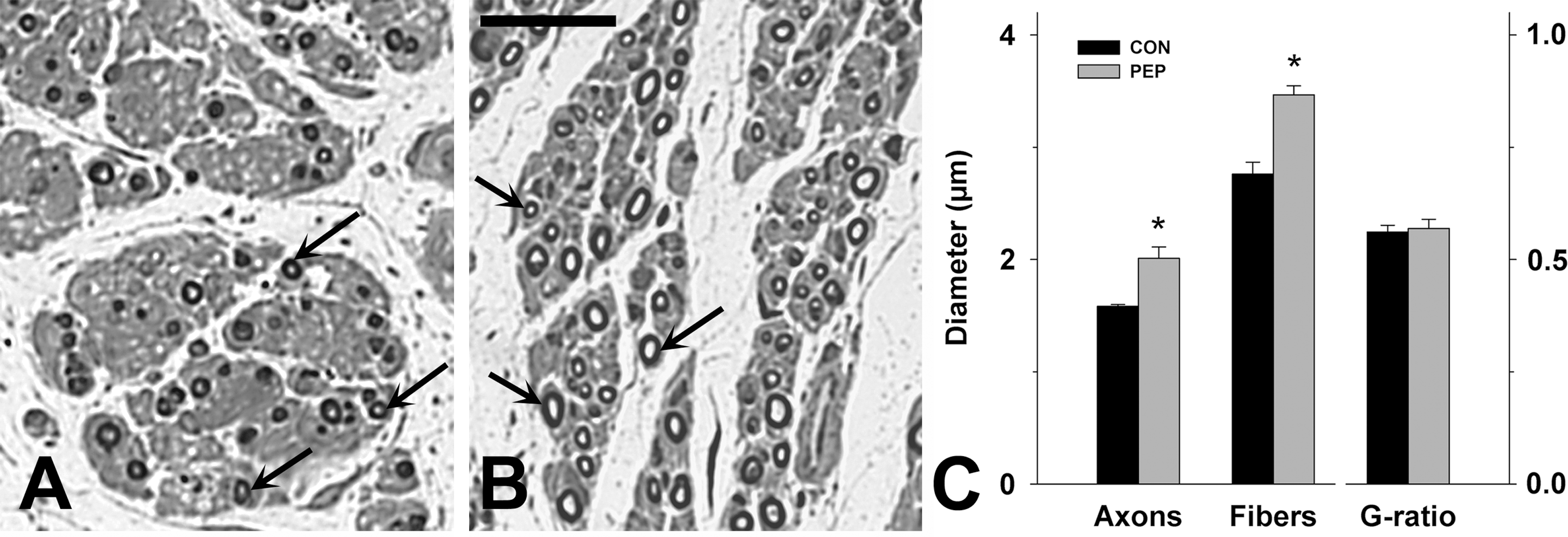
Treatment improves diameters of regenerated axons. (
The HNK-1 mimetic is not immunogenic
Analysis by ELISA revealed that sera from mimetic- and control peptide-treated animals collected 163–165 days after injury and from untreated age-matched macaques react at similar levels with the HNK-1 mimetic peptide (Fig. 6). These levels were comparable to background reactivity estimated by binding of a PSA-mimetic peptide, which none of the animals received. For control, substrate-coated HNK-1 and PSA-specific antibodies reacted well with the appropriate mimic peptide. These findings indicate that the HNK-1 mimetic does not induce an antibody response in treated animals. In addition, cellular infiltrates and necrosis were not observed in nerve sections from mimetic-treated, as well as control peptide-treated animals (Fig. 5A and B), indicating that cellular immune responses were also not induced.
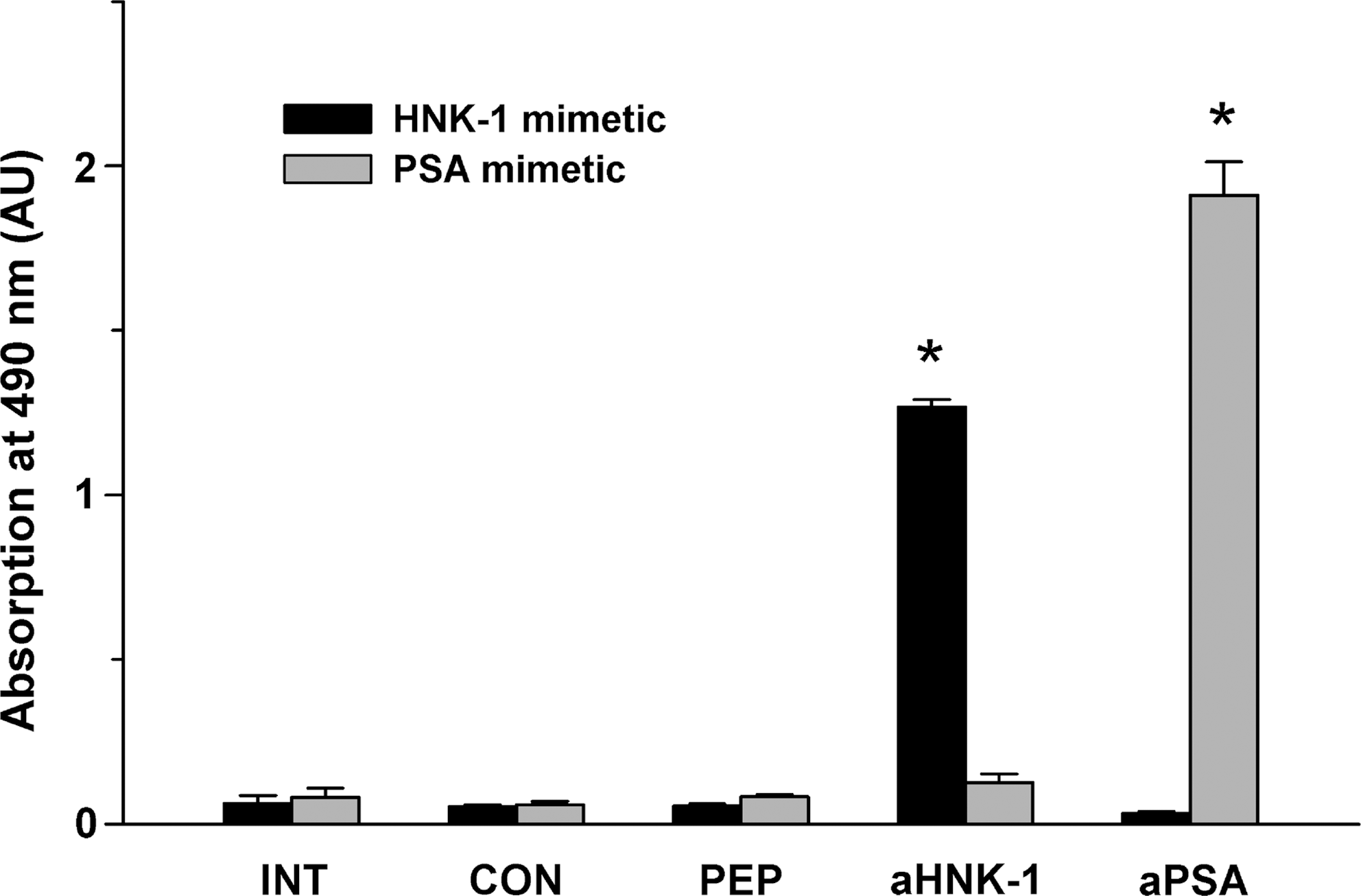
Circulating antibodies against the human natural killer cell-1 (HNK-1) mimetic are not detectable in treated animals. Binding of HNK-1 mimetic peptide (HNK-1 mimetic) or α2,8 polysialic acid (PSA)-mimetic peptide (PSA mimetic) to substrate-coated serum proteins was assessed by ELISA using sera from control peptide-treated (CON) and mimetic-treated (PEP) animals collected 163–165 days after injury, and from untreated (INT) age- and sex-matched control macaques. The monoclonal antibody 412 against the HNK-1 epitope (aHNK-1) served as a positive control. The anti-PSA antibody 735 was used to assess background reactivity. Shown are mean values+standard error of the mean of triplicate measurements. Asterisks indicate significant differences compared with values of untreated (INT) animals (p<0.05 by one-way analysis of variance with Dunnett's post-hoc test; AU, arbitrary units).
Discussion
This study provides evidence for the beneficial effects of an HNK-1 glycomimetic peptide on the outcome of femoral nerve injury in adult primates. More than 5 months after application, we observed improved gait, increased muscle force, enhanced H-reflex responses, reduced H-reflex latency, and increased axonal diameters compared with control treatment.
In this study we used, to the best of our knowledge, for the first time the femoral nerve injury model in a non-human primate. Therefore, to substantiate our claims of treatment efficiency, here we have to critically consider the reliability and adequacy of our outcome measures, in particular the gait parameters that have not been described before. To assess gait deficits after femoral nerve injury in mice, we have devised a quantitative video-based approach, the single-frame motion analysis (Irintchev et al., 2005), which has proven its reproducibility and usefulness in a series of studies (Ahlborn et al., 2007; Eberhardt et al., 2006; Guseva et al., 2009; Malin et al., 2009). The parameters used for mice were, however, inadequate for the macaque model. We defined new parameters which were sensitive to detect alterations in gait after injury (see Fig. 2A–F). The overall measure of recovery, the recovery index calculated from the three most characteristic parameters, was approximately 50% in control peptide-treated animals 160 days after injury, indicating the persistence of a significant functional deficit (maximum recovery is 100%). This estimate of 50% functional disability was in good correspondence with deficits in muscle force and muscle weight, which were by 50% and 40% lower, respectively, in the injured compared to the contralateral, non-injured side of control peptide-treated macaques at 160 days after injury. We therefore believe that our functional analysis, used to follow the time course of recovery and to estimate the final outcome, is adequate and precise.
In mimetic-treated animals, we found a significant, compared with control peptide-treated macaques, improvement of gait parameters at different time points after injury, and the improved gait correlated with increased muscle force at 160 days. Although muscle weight was no different in the two groups, which we attribute to large inter-individual variability, we can conclude that in conjunction with the observation of increased axonal diameters, the treatment with the glycomimetic peptide leads to a better outcome of femoral nerve repair in macaques. However, we would like to emphasize that the mimetic treatment improves the outcome of nerve repair, but does not lead to complete restoration of function.
Our data on the M-wave (known also as compound muscle action potential, CMAP) seem to contradict the results of gait and force analyses in two ways. First, recovery appears to be complete in control peptide-treated animals (90% compared to contralateral, non-denervated muscles) and second, no effect of treatment with the glycomimetic peptide was found. The CMAP is a measure of muscle reinnervation and is useful in following the time course of reinnervation in clinical and experimental studies, as shown also for macaques (Krarup et al., 2002). It cannot be considered, however, to represent a precise measure of muscle function at late time points after nerve injury. For example, in macaques the absolute CMAP amplitudes reach normal values (90–93% of intact animals) 2 years after transection and repair of the median nerve by direct suture or a 5-mm nerve guide, surgical reconstructions which rarely if ever result in complete restoration of function (Krarup et al., 2002; Lundborg, 2003). Therefore, the finding that M-wave amplitudes were normal, while functional deficits were present in both mimetic- and control peptide-treated animals, can readily be explained.
In contrast to the M-waves, we found (1) that the amplitudes of the H-waves on the operated side of control peptide-treated monkeys were abnormally low when compared to the contralateral non-operated side, and (2) that the glycomimetic peptide treatment resulted in a significant increase of the H-wave. We interpret this effect of the glycomimetic as evidence for improved functionality of the spinal reflex circuitry. Studies in rats have led to the conclusion that the H:M ratios negatively correlate with the functional outcome of peripheral nerve injuries (Valero-Cabré and Navarro, 2001). This notion is, however, based on relative values (H:M ratios normalized to control H:M ratios), and it is not clear how changes in absolute H-wave values relate to restoration of function in the rat. It is also not clear to what degree conclusions reached using one experimental model are relevant for other models, considering the large differences in the normal H-reflex responses (e.g., stimulus-response characteristics, amplitudes, H:M ratios, and rate of depression) in different species, such as rats and humans, in different muscles of the same animals, for example plantar and gastrocnemius muscles of the rat, and obtained under different recording conditions (Cliffer et al., 1998; Gozariu et al., 1998; Knikou, 2008; Valero-Cabré and Navarro, 2001). Therefore, our interpretation of the treatment-induced alteration in the H-reflex is not necessarily in contradiction with previous results.
Considering previous observations in mice after glycomimetic application (Mehanna et al., 2009; Simova et al., 2006), it is not surprising that a single intra-operative application of a peptide can produce late-appearing and long-lasting effects despite the fact that its local availability is most likely limited to hours or days because of diffusion from the cuff and degradation by peptidases originating from the damaged tissues and infiltrating macrophages. Our current explanation of such long-lasting effects is that the mimetic causes priming of the injured femoral nerve, both at and around the lesion site, in the sense that early cellular and molecular responses to injury are modulated so that the subsequent regeneration process is favorably influenced over weeks. For the HNK-1-mimicking peptide (Simova et al., 2006), we proposed that priming is achieved by activation of the receptor for advanced glycation end-products (RAGE) signaling pathway. In the case of a PSA mimetic (Mehanna et al., 2009), we found evidence for an NCAM/fibroblast growth factor receptor-mediated mechanism of action. Priming of the response to injury is apparently not restricted to glycomimetics. A long-lasting effect is observed after a brief low-frequency electrical stimulation (1 h, 20 Hz) of the proximal nerve stump of the femoral nerve immediately after nerve transection. This treatment significantly shortens the period of asynchronous axonal regrowth staggered over weeks after femoral nerve lesion in rats (Al-Majed et al., 2000b; Brushart et al., 2002), and accelerates functional recovery after femoral nerve lesion in mice (Ahlborn et al., 2007). These effects are associated with an accelerated and enhanced upregulation of the expression of brain-derived neurotrophic factor (BDNF) and its tyrosine kinase B (TrkB) receptor in motoneurons induced by the depolarization of motoneuron cell bodies during the 1-h stimulation period (Al-Majed et al., 2000a).
Here we found a remarkable increase of axon diameters, but no effect on the degree of remyelination. In contrast, in the mouse remyelination is enhanced by the HNK-1 mimetic, while axon diameters are not affected (Simova et al., 2006). These differential effects in the two species are encouraging: despite differences in tissue responses, glycomimetic treatment significantly reduced injury-induced gait deficits in both species. Likewise encouraging in the present study is that effects of the treatment could be detected at the functional and structural levels, despite of the use of a limited number of animals, and despite in contrast to inbred mice, the genetic variability of the primates.
In conclusion, the results of this study, in conjunction with previous observations in the mouse, provide evidence for the clinical potential of the HNK-1 glycomimetic treatment, and encourage independent replication studies and clinical trials.
Footnotes
Acknowledgments
This work was supported by the Deutsche Forschungsgemeinschaft (SCHA 185/54-1 to M.S. and A.I.), and the New Jersey Commission for Spinal Cord Research (M.S.), Natural Science Foundations of China (30872829 and 30571998 to S.Y.), and the Army Research Foundations of Health and Medicine (06MA148 and 01MB039 to H.Z.). We are grateful to Emanuela Szpotowicz for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.