
Abstract
A major obstacle for the transplantation of neural stem cells (NSCs) into the lesioned spinal cord is their predominant astrocytic differentiation after transplantation. We took advantage of this predominant astrocytic differentiation of NSCs and expressed the paradigmatic beneficial neural cell adhesion molecule L1 in radial glial cells and reactive and nonreactive astrocytes as novel cellular vehicles to express L1 under the control of the promoter for the human glial fibrillary acidic protein (GFAP-L1 NSCs). Behavioral analysis and electrophysiological H-reflex recordings revealed that mice transplanted with GFAP-L1 NSCs showed enhanced locomotor recovery in comparison to mice injected with wild type (WT) NSCs or control mice injected with phosphate-buffered saline (PBS). This functional recovery was further accelerated in mice transplanted with L1-expressing radial glial cells that had been immunoisolated from GFAP-L1 NSCs (GFAP-L1-i cells). Morphological analysis revealed that mice grafted with GFAP-L1 NSCs exhibited increased neuronal differentiation and migration of transplanted cells, as well as increased soma size and cholinergic synaptic coverage of host motoneurons and increased numbers of endogenous catecholaminergic nerve fibers caudal to the lesion site. These findings show that L1-expressing astrocytes and radial glial cells isolated from GFAP-L1 NSC cultures represent a novel strategy for improving functional recovery after spinal cord injury, encouraging the use of the human GFAP promoter to target beneficial transgene expression in transplanted stem cells.
Introduction
In this study, we examined a novel approach to apply L1, an adhesion molecule beneficial to regeneration in the nervous system, by taking advantage of the predominant astrocytic differentiation of NSCs to express L1 under the control of the human glial fibrillary acidic protein (GFAP) promoter, to test whether radial glia and astrocytes that normally do not express L1 could serve as cellular vehicles to improve regeneration, as the L1-positive Schwann cells are capable of (Lavdas et al., 2010). The hypothesis that L1-expressing NSC-derived astrocytes would promote regeneration is driven by a previous study in which we demonstrated that adeno-associated virus-driven ectopic expression of L1 in endogenous spinal cord astrocytes had beneficial effects on regeneration after spinal cord injury (Chen et al., 2007). Our novel approach was designed to provide new insights into how the human GFAP promoter may serve as a driving force in NSCs and their progeny for expression of regeneration-conducive molecules in general.
The human GFAP promoter has been shown to be active not only in reactive and non-reactive astrocytes but also in radial glial cells in the developing mouse brain (Liu et al., 2006; Platel et al., 2009) and has successfully been used to isolate radial glial cells from the developing rodent brain (Malatesta et al., 2000). The neural cell adhesion molecule L1 promotes neurite outgrowth, neuronal migration, and neuronal survival (Maness et al., 2007). It has successfully been applied to enhance neuronal differentiation and survival of grafted embryonic stem cells after transplantation into the central nervous system of mice (Bernreuther et al., 2006; Chen et al., 2005). Furthermore, transplanted L1 overexpressing embryonic stem cells rescued host dopaminergic neurons in a mouse model of Parkinson's disease (Cui et al., 2010). Because radial glial cells have been shown to exert beneficial effects after transplantation into the rodent spinal cord (Chang et al., 2009; Hasegawa et al., 2005), are stable with regard to their lineage in the intact animal and considered to be less, if at all, prone to form tumors, we purified L1- expressing radial glial cells comprising ventricular cells with expression of L1 under the control of the human GFAP promoter prior to transplantation into the injured spinal cord to monitor the effects on functional recovery, opening a novel approach to using L1- expressing cells when differentiated into astrocytes to express a regeneration-conducive molecule, thus overcoming the purported negative effects of astrocytes as a molecular barrier for regeneration in the hostile environment of the adult mammalian nervous system and allowing these cells to express a beneficial molecule known to be regeneration conducive when expressed or overexpressed by Schwann cells (Lavdas et al., 2010).
Methods
Animals
Female C57BL/6J mice were used at the age of 3–4 months. Mice ectopically expressing L1 under the control of the human GFAP promoter were generated as described (Diedrich et al., 2008). These animals develop normally, and have a normal life span and no obvious behavioral or morphological phenotype compared to their wild-type littermates, although a detailed analysis has not been performed. To visualize donor cells for transplantation, these mice were cross bred with mice ubiquitously expressing enhanced green fluorescent protein (GFP) under the control of the chicken β-actin promoter (Okabe et al., 1997). The resulting double transgenic mice were viable, without a conspicuous phenotype, with the exception that all cells were green under fluorescent light. All mice used in this study were backcrossed to C57BL/6J for at least 10 generations to avoid the need for immune suppression when NSCs derived from these mice were transplanted syngeneically into a recipient C57BL/6J mouse.
Culture of neural stem/progenitor cells
Stem cells were isolated from 14-day-old embryos (E14) as described (Dihne et al., 2003). Briefly, lateral and medial ganglionic eminences were removed from every fetus, mechanically dissociated using a pipette, and grown in a defined medium composed of a 1:1 mixture of Dulbecco's Modified Eagle's Medium and F-12 supplemented with glucose (0.6%, Sigma-Aldrich, Deisenhofen, Germany), sodium bicarbonate (3 mM, Invitrogen, Karlsruhe, Germany), B27 (2%, Invitrogen), glutamine (2 mM; Invitrogen), and HEPES buffer (5 mM, Sigma-Aldrich). The medium was supplemented with epidermal growth factor (EGF; 20 ng/mL; PreproTech, Rocky Hill, NY), and basic fibroblast growth factor (FGF-2; 20 ng/mL; PreproTech). The cells grew in neurospheres that were passaged by mechanical dissociation ∼ every 5th day and reseeded with a density of 50,000 cells/mL. Vital cells were determined by 0.5% Trypan blue dye (Invitrogen) exclusion. Experiments were performed with neurospheres between passages 3 and 6.
Immunocytochemistry and immunohistochemistry
For immunocytochemical analysis of fixed cells, cultured cells were washed in phosphate-buffered saline (PBS), pH 7.3, and fixed in 4% paraformaldehyde. For immunohistochemical analysis of the spinal cord, mice were anesthetized and transcardially perfused with fixative consisting of 4% formaldehyde and 0.1% CaCl2 in 0.1 M cacodylate buffer (Sigma-Aldrich), pH 7.3, for 15 min at room temperature (RT). Spinal cords were removed following fixation, incubated in 4% formaldehyde for 24 h followed by incubation in 15% sucrose in PBS, pH 7.3, for ∼48 h at 4°C. Serial longitudinal sections were cut in a cryostat. Serial longitudinal sections 25 μm thick were cut in a cryostat and collected on SuperFrost Plus glass slides (Roth, Karlsruhe, Germany). Antigen retrieval was performed in 0.01 M sodium citrate solution, pH 9.0, for 30 min at 80°C prior to the application of primary antibodies. Before application of primary antibodies, coverslips or sections were incubated in PBS containing 0.2% v/v Triton X-100, 0.02% w/v sodium azide (Sigma-Aldrich) and 5% v/v appropriate normal serum (Jackson Immunoresearch Laboratories, West Grove, PA) for 30 min. Primary antibodies were applied overnight at 4°C. After washing in PBS, appropriate secondary antibodies were applied for 1 h at RT. Finally, coverslips or sections were washed with PBS. To determine total cell numbers in vitro, cells were counterstained with DAPI (Sigma-Aldrich) for 2 min. For 5-bromo-2-deoxyuridine (BrdU) staining, DNA was denatured with 2 M HCl for 30 min at 37°C. Monoclonal mouse antibody to BrdU (1:100; Developmental Studies Hybridoma Bank, Iowa City, IA) was administered overnight at 4°C. For immunocytochemistry of living cells, fixation was omitted, and cells were washed and incubated with primary antibody on ice for 20 min, followed by washing and incubation with secondary antibody for 15 min on ice. Subsequently, cells were fixed in 4% paraformaldehyde in PBS and mounted on objectives with Fluoromount G medium (Polysciences, Warrington, PA). For negative controls, primary antibody was omitted. Specimens were examined with a fluorescence (Axioplan 2; Carl Zeiss Microimaging, Thornwood, NY) or confocal laser-scanning microscope (Leica SP2; Leica Microsystem, Heidelberg, Germany).
Primary antibodies used were polyclonal goat antibodies against doublecortin (DCX) (1:200; Santa Cruz Biotechnology, Santa Cruz, CA) and choline acetyltransferase antibody (ChAT) (1:100; Millipore, Hofheim, Germany), polyclonal rabbit antibodies against RC2 (1:10; Developmental Studies Hybridoma Bank), tyrosine hydroxylase (TH) (1:800; Millipore), GFAP (1:1000; Dako, Glostrup, Denmark), neuronal class III ß-tubulin (Tuj1) (1:2000; Covance, Berkeley, CA), L1 (1:250) (Rathjen et al., 1984), Ki-67 (1:500; Abcam, Cambridge, UK), rat monoclonal antibody against 5-hydroxytryptophan (5-HT) (1:100, Millipore), and mouse monoclonal antibodies against CaMKII (1:1000; Sigma), L1 (1:100; Chemicon), nestin (1:50; Developmental Studies Hybridoma Bank), and 2′,3′-cyclic nucleotide 3′ phosphodiesterase (CNPase; 1:1,000, Sigma-Aldrich). Appropriate cyanine 2 (Cy2)-, Cy3-, and Cy5-conjugated secondary antibodies (Jackson Immunoresearch) were used to detect primary antibodies.
Western blot analysis
Protein samples were heated at 95°C for 5 min in SDS sample buffer. An SDS-PAGE gel was run at constant voltage of 60 V for 20 min and then at 100 V until the Bromophenol Blue line had migrated to the bottom of the gel. Proteins were transferred at constant voltage of 80 V for 150 min at 4°C from the SDS-polyacrylamide gel onto a nitrocellulose membrane (Protran, Nitrocellulose BA 85, Schleicher & Schüll, Dassel, Germany) using a Mini Transblot apparatus (Bio-Rad, Munich, Germany) as described in the manufacturer's instructions. Immunoreactivity was visualized by the enhanced chemiluminescence detection system (ECL; Pierce, Rockford, IL). Antibodies used were rabbit antibody against L1 (1:1000) (Rathjen et al., 1984) and mouse antibody against glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (1:1000; Millipore).
Immunoisolation of L1-positive cells
Immunoisolation of L1 overexpressing NSCs was performed as described (Jungling et al., 2003) with modifications. Briefly, plastic Petri dishes were coated with a goat anti-rabbit IgG antibody (80 μl antibody, 1.3 mg/mL; Jackson Immunoresearch) in 10 ml Tris/HCl (100 mM, pH 9.5) for 24 h at 4°C. After washing with PBS, the dishes were incubated with bovine serum albumin (0.2% in PBS, pH 7.3) for 24 h at 4°C. Thereafter, a polyclonal rabbit antibody against L1 (Rathjen et al., 1984) was added and incubated overnight at 4°C (diluted in 0.2% bovine serum albumin [BSA] in PBS, final antibody concentration 5 μg/mL). Dissociated neurospheres were added to the L1-coated Petri dish and incubated for 1.5 h at RT. Then, nonadherent cells were removed by washing once with PBS. Adherent cells were harvested and cultured as neurospheres for further experiments.
Proliferation and differentiation of neural precursor cells in vitro
For measurement of overall proliferation of neural precursor cells, dissociated precursor cells were plated on coverslips coated with 0.01% poly-
Neurosphere migration assay
The neurosphere migration assay was performed as described (Nguyen-Ba-Charvet et al., 2004) with modifications. Briefly, neurospheres of similar diameters (100–300 μm) were picked up individually from suspension cultures using a 10-μL pipette tip and seeded in 5-μL droplets on PLL-substrated coverslips for 20 min. After 20 min of adhesion, the medium was adjusted to 500 μL per well with NSC culture medium without EGF and FGF-2. Migrating cells were monitored for 24 h. The longest distance between the edge of a sphere and the leading NSCs was measured in each quadrant. At least 40 spheres were analyzed, and the experiment was performed in triplicate.
Spinal cord injury surgical and cell transplantation procedures
Spinal cord injury was performed as described (Apostolova et al., 2006). Briefly, mice were anesthetized by intraperitoneal injections of ketamine and xylazine (100 mg of Ketanest [Parke-Davis/Pfizer, Karlsruhe, Germany] and 5 mg of Rompun [Bayer, Leverkusen, Germany] per kilogram of body weight). Laminectomy was performed at the T7–T9 level with mouse laminectomy forceps (Fine Science Tools, Heidelberg, Germany). A mouse spinal cord compression device was used to elicit compression injury (Curtis et al., 1993). The spinal cord was maximally compressed (100% in accordance with the operational definition of Curtis and colleagues [Curtis et al., 1993]) for 1 sec by a time-controlled 12-V (maximum voltage) current flow through an electromagnetic device. For cell transplantation, NSCs were dissociated and resuspended in PBS at a density of 100,000 viable cells per μL and 1 μL was injected into the spinal cord 0.5 mm both rostral and caudal to the lesion site by slightly off-midline penetration using a 1-μL Hamilton syringe without needle bevel fixed on a stereotaxic frame. The injections were performed slowly (1 μL over 5 min) and the needle of the Hamilton syringe was retracted slowly after the cell transplantation to avoid leakage of the cell suspension. Viability (>90%) was determined by Trypan blue exclusion prior to transplantation. After surgery mice were allowed to recover from anaesthesia at 35°C and were provided water and food ad libitum. For the duration of our follow-up period (2 months) we did not observe any apparent signs of increased pain sensitivity or the presence of chronic pain, and there were no self mutilations in any experimental group. This is important to note for the animals transplanted with L1-overexpressing cells, because a previous report suggested a role for L1 in sprouting of pain fibers and hyperalgesia after spinal cord injury (Hoschouer et al., 2009). All experiments were performed in accordance with the German and European Community laws on protection of experimental animals, and the animal surveillance and the surgical procedures used were approved by the responsible Animal Care and Use Committee of the University and State of Hamburg.
Graft volume and density
Unbiased estimates of the total number of grafted cells and graft volume per animal 4 and 8 weeks after transplantation were calculated according to the optical dissector and Cavalieri methods (Gundersen, 1986). An Axioskop microscope (Carl Zeiss) and a Neurolucida software-controlled computer system were used for quantitative analysis (MicroBrightField Europe, Magdeburg, Germany). Graft volume and cell density of the graft were determined measuring every tenth section. Transplanted cells were identified by their GFP signal. Graft areas were outlined on digitized images to calculate volumes considering section thickness and frequency. Using random sampling in the graft core and in the periphery of the graft, cell counts were performed at a magnification of×40.
Measurement of the longitudinal and horizontal extent of the graft
The lesion site was delineated at low magnification and the longest distance between GFP-positive cells and the center of the lesion in horizontal and longitudinal direction was measured.
Determination of differentiation, proliferation, and neurite outgrowth of transplanted cells
To determine total numbers of donor cells in vivo, GFP-positive cells were counted and the ratio of cell type-specific marker-positive cells of all GFP-positive cells was calculated. To measure neurite outgrowth, graft-derived (GFP-positive) β-tubulin III-positive neurons were identified and the length of the longest neurite of each β-tubulin III-positive cell was determined.
Quantification of motoneuron soma size and perisomatic terminals
Estimations of soma size of and perisomatic terminals on endogenous motoneurons were performed as described (Apostolova et al., 2006; Jakovcevski et al., 2007). Longitudinal spinal cord sections stained with antibodies against ChAT were analyzed distal to the lesion site. Stacks of 1-μm-thick images were obtained on a Leica SP2 confocal laser-scanning microscope; using a 63x, 1.5 oil-immersion objective and digital resolution of 512×512 pixel. Four adjacent stacks (frame size, 153×153 μm) were obtained consecutively in a rostrocaudal direction; motoneurons located both close to and remote from the lesion scar were analyzed. The area of the soma of ChAT-positive neurons and the linear density of ChAT-positive puncta were determined by measuring one image per cell at the level of the largest cell body cross-sectional area. Areas and perimeters were measured using the Image Tool 2.0 software program (University of Texas, San Antonio, TX). Linear density was calculated as number of perisomatic terminals per unit length.
Monoaminergic reinnervation of the distal spinal cord
TH- and 5-HT-positive axons were quantified as described (Jakovcevski et al., 2007). Briefly, numbers of axons projecting beyond an arbitrarily selected border 250 μm distal to the lesion site in spaced serial parasagittal sections were counted 8 weeks after spinal cord injury.
Estimation of the regrowth of the corticospinal tract (CST)
For estimation of the regrowth of the CST, animals from each group were killed and consecutive parasagittal spinal cord sections were collected. Spinal cord sections were then immunostained with antibodies against CaMKII. This antibody stains specifically descending corticospinal axons and no ascending axons in dorsal columns of the rodent spinal cord (Terashima et al., 1994). To quantify corticospinal axon regrowth, the rostral border of the lesion site demarcated by GFAP immunostaining was chosen as the reference point for measurements. We chose this border as it was used in our previous studies (Chen et al., 2005, 2007), and our data indicate that there was no difference among experimental groups in lesion scar volume and extent of GFAP immunoreactivity. The distance between the rostral border and the furthest detectable axon tips of the CST axons in consecutive sections was measured.
Estimation of lesion scar volume
Spaced serial 25-μm-thick sections 250 μm apart stained with cresyl violet were used for estimations of the scar volume using the Cavalieri principle. Areas of the scar required for volume estimation were measured directly under the microscope using the Neurolucida software (MicroBrightField Europe).
Measurement of astrogliosis
To evaluate the astrogliosis, astrocytes were stained with antibodies against GFAP and the immunofluorenscence intensity was quantified. Briefly, confocal images were acquired using a Leica confocal microscope (Leica SP2, Leica Microsystems). Quantification of fluorescence intensity was performed. The confocal settings for image acquisition were maintained for all sections. Image stacks were merged into a single image by a maximum projection and fluorescence intensity was analyzed with ImageJ software (
Analysis of motor function
Recovery of ground locomotion was evaluated using the Basso Mouse Scale (BMS) score (Basso et al., 2006). Moreover, motor functional recovery was analyzed using single-frame motion analysis for numerical assessment of motor behavior (Apostolova et al., 2006; Chen et al., 2007). This method included evaluation of three parameters in two different tests: beam walking (foot-stepping angle, rump-height index) and voluntary movements without body-weight support (extension–flexion ratio). Video recordings were taken before and at 1, 3, and 6 weeks after spinal cord injury with a high-speed video camera and analyzed with the affiliated software (SIMI Motion, SIMI Reality Motion Systems GmbH, Unterschleissheim, Germany). Values for the left and right extremities were averaged. Recovery indices were used as a measure of functional recovery on an individual animal level as described (Apostolova et al., 2006). The recovery index (RI) was calculated (percentage) as follows: RI=[(X 7+n – X 7)/(X 0 – X 7)] x 100, where X 0, X 7, and X 7+n are values before operation, 7 days after injury, and a time point n days after spinal cord injury, respectively. In simpler terms, this measure estimates gain of function (X 7+n – X 7) as a fraction of the functional loss (X 0 – X 7) induced by the operation (Apostolova et al., 2006).
Rate depression of the H-reflex electromyography (EMG) recordings
The EMG recordings were performed as described (Lee et al., 2009). Briefly, mice were anesthetized by intraperitoneal injections of ketamine and xylazine as described previously. The sciatic nerve was stimulated using bipolar electrical pulses of 0.2 ms duration to elicit reflex responses with electronic pulses at 6 weeks after spinal cord injury (Model 2100 Isolated Pulse Stimulator, A-M systems, Calsborg, WA). The signals of M- and H-responses were measured by a recording electrode placed in the plantar muscles between the second and third cuneiform bones during repetitive stimulation of the sciatic nerve and transmitted to a differential amplifier (P55 General Purpose AC Preamplifier, Grass Technologies, West Warwick, RI), magnified (x1000) and filtered (300 Hz high pass and 1 kHz low pass). The latencies of the responses were measured as time elapsed between trigger and peak of each waveform. Stimulus intensity was gradually increased until both M- and H-waves had latencies of ∼2 and 5 ms, respectively. After the threshold measurement, stimulus intensity was further increased until maximal and stable H-responses were elicited. Thereafter, stimulation continued at the defined suprathreshold level at frequencies of 0.1, 0.3, 0.5, 1, 2, 10, and 20 Hz. The amplified analog signal was delivered to an A/D converter (ADC42, Pico Technology, Cambridgeshire, U.K.) and the data were analyzed using the Picoscope data acquisition software (PicoScope 5). The amplitudes of M and H waves were measured as peak-to-peak values and averaged to calculate H/M ratios.
Results
Generation and immunoisolation of NSCs ectopically expressing the cell adhesion molecule L1 under the control of the human GFAP promoter
NSCs were isolated from the ganglionic eminence of 14-day-old mouse embryos that ectopically express L1 under the control of the human GFAP promoter (GFAP-L1 NSCs). Because L1 expression is driven by the human GFAP promoter, L1 is predicted to be expressed by radial glial cells, immature and mature astrocytes. Immunocytochemical analysis of undifferentiated GFAP-L1 NSC cultures under the influence of FGF-2 and EGF revealed that L1-expressing cells accounted for ∼5% of GFAP-L1 NSCs only (Fig. 1A). L1 was co-expressed in a subset of cells with RC2 (Fig. 1B) and nestin (Fig. 1C), markers commonly expressed in radial glial cells. Few GFAP-L1 NSCs co-expressed L1 and the astrocytic marker GFAP (Fig. 1D). In wild type (WT) NSCs, L1 expression was observed in <0.1% of all cells. Seven days after induction of differentiation by withdrawal of FGF-2 and EGF, astrocytes differentiated from GFAP-L1 NSC cultures showed positive staining with antibodies against L1 (Fig. 1E). No L1 expression was detected in astrocytes derived from WT NSCs (not shown). Western blot analysis before and after induction of differentiation showed that L1 expression was negligible in WT NSCs before and 4 days after induction of differentiation, but was increased 7 days after induction of differentiation because of neuronal differentiation of a subset of NSCs. In GFAP-L1 NSCs, L1 expression was strongly enhanced at all time points with highest L1 expression 4 days after induction of differentiation (Fig. 1F).
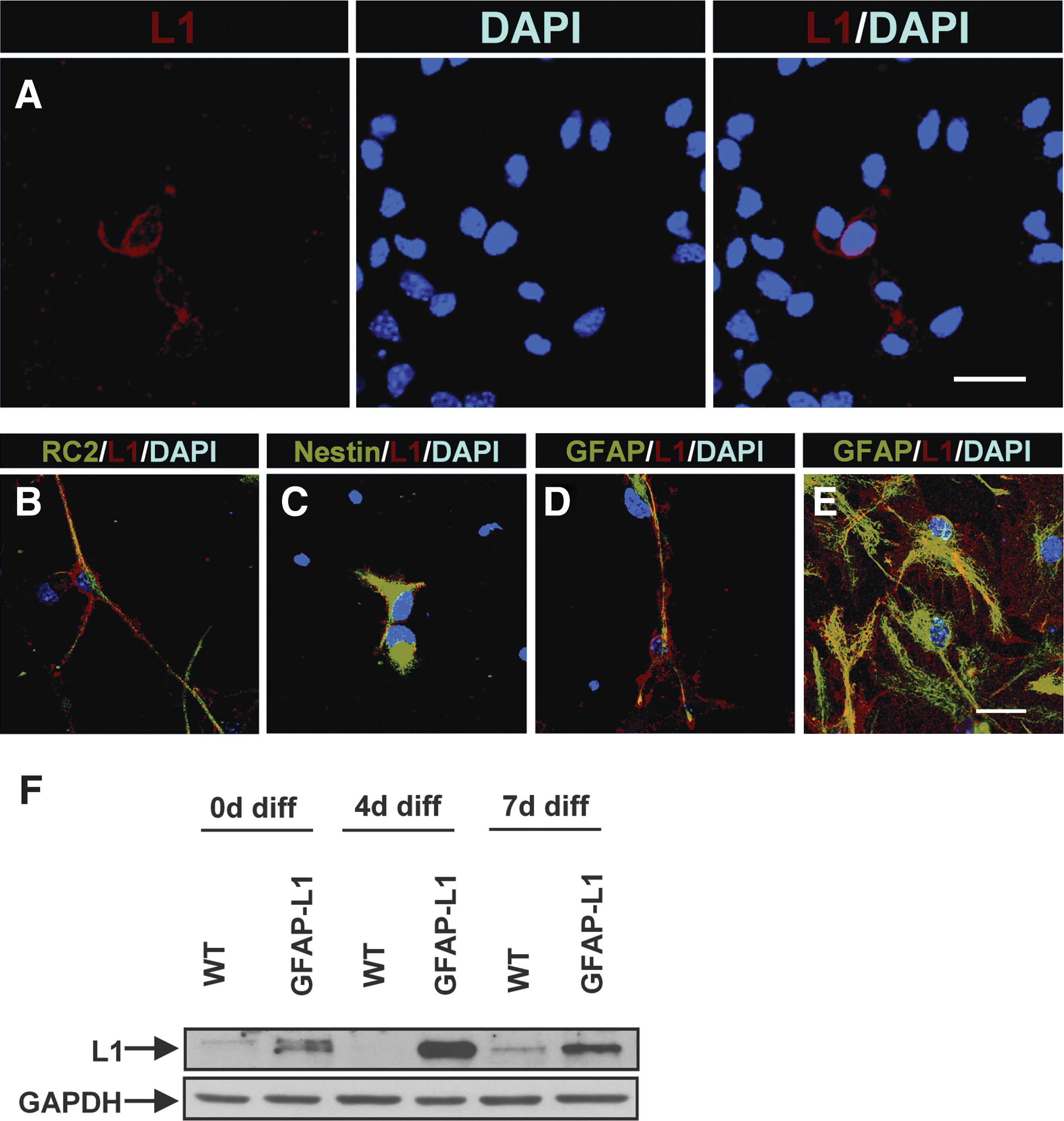
Cell adhesion molecule L1 expressed under the control of the human glial fibrillary acidic protein (GFAP) promoter is ectopically expressed on neural stem cell-derived radial glial cells and astrocytes in vitro.
To enhance the percentage of L1-expressing cells, NSCs that had been derived from mice cross-bred with mice ubiquitously expressing GFP under the control of the chicken actin promoter to allow for easy detection in subsequent transplantation experiments were subjected to immunoisolation using L1 polyclonal antibodies (Fig. 2A) to purify L1-expressing cells from GFAP-L1 NSCs. Whereas the L1-positive cells isolated from GFAP-L1 NSCs (GFAP-L1-i cells) could be further cultured as multipotent proliferating neurospheres in medium containing EGF and FGF-2, no proliferating L1-positive cells could be isolated from WT NSCs under the same conditions. Immunocytochemical live staining of cells using a monoclonal antibody against L1 revealed that L1 expression remained stable in GFAP-L1 cells after immunoisolation, with the majority of isolated cells showing L1 expression at the cell surface 6 days after isolation (Fig. 2B). Furthermore, most of the immunoisolated cells expressed the radial glia markers nestin and RC2, whereas marker proteins of differentiated neurons and astrocytes were rarely observed (Fig. 2B).
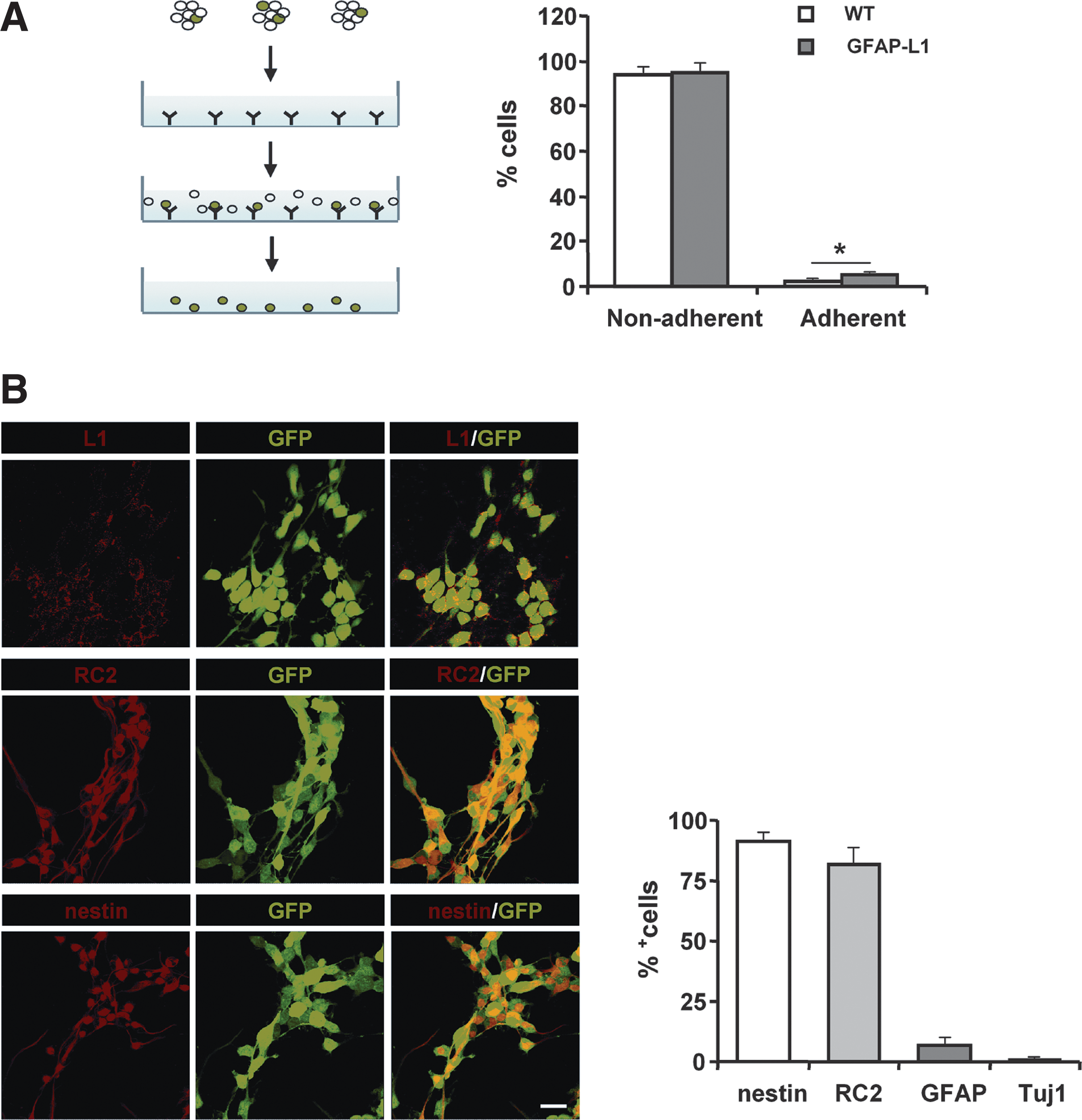
Immunoisolation and immunocytochemical characterization of L1 expressing cells derived from glial fibrillary acidic protein (GFAP)-L1 neural stem cells (NSCs) or wild type (WT) NSCs expressing green fluorescent protein.
Ectopic expression of L1 in GFAP-L1 NSCs and GFAP-L1-i cells reduces proliferation, enhances migration and neuronal differentiation, and decreases astrocytic differentiation in vitro
Adherent cultures of GFAP-L1 NSCs, GFAP-L1-i cells, and WT NSCs maintained in the presence of FGF-2 and EGF were labeled with BrdU. Immunohistochemical analysis revealed that GFAP-L1 NSCs and GFAP-L1-i cells showed a significant reduction of BrdU-positive cells, when compared to WT NSCs (Fig. 3A).
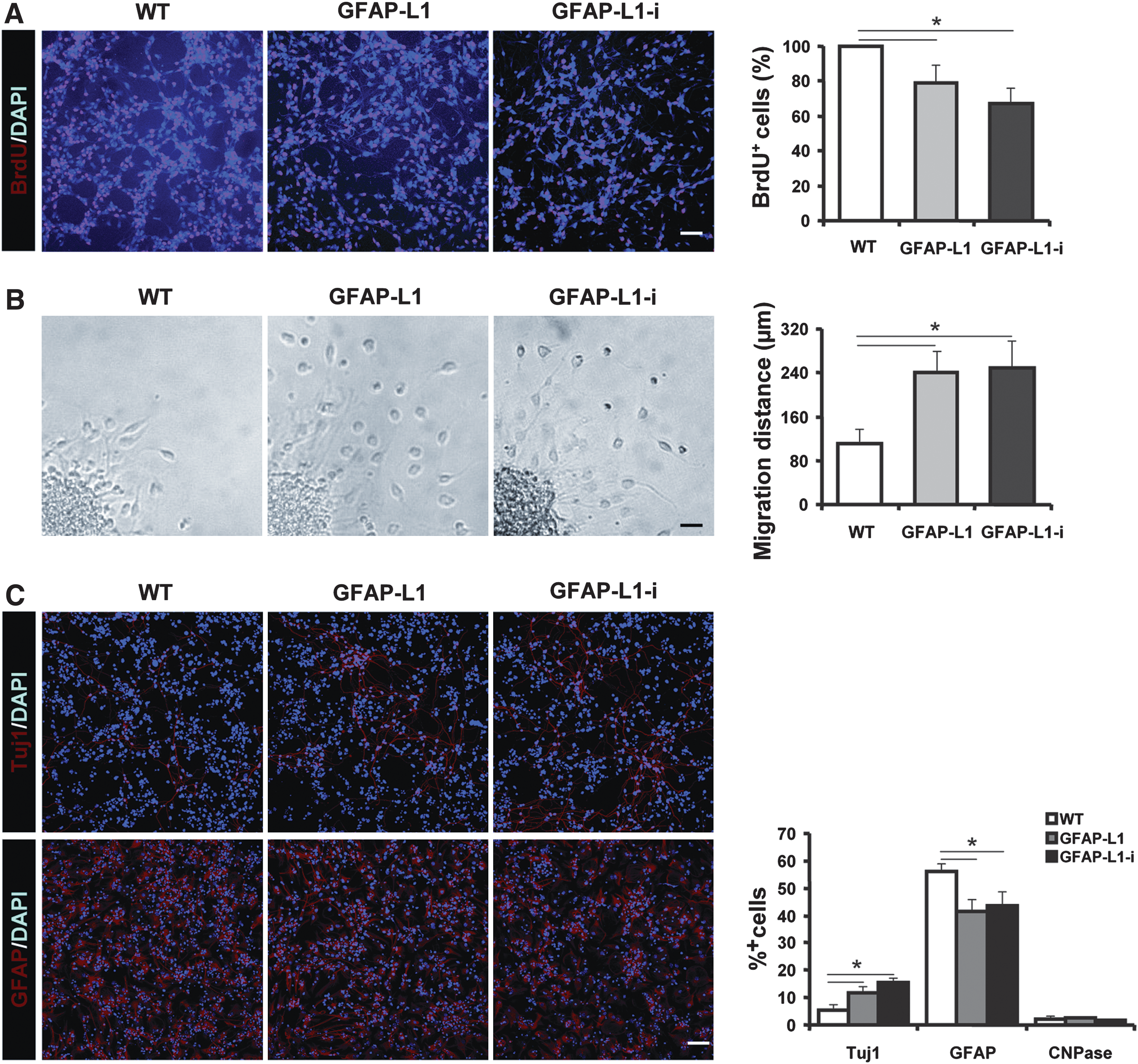
Ectopic expression of L1 in glial fibrillary acidic protein (GFAP)-L1 neural stem cells (NSCs) and GFAP-L1-i cells inhibits proliferation, enhances migration and neuronal differentiation, and decreases astrocytic differentiation in vitro.
To investigate whether ectopic expression of L1 in NSCs affected migration of cells, neurospheres were plated on a PLL-coated substrate. Twenty-four hours after plating, GFAP-L1 NSCs and GFAP-L1-i cells were found to migrate further from the neurosphere edge than WT NSCs (Fig. 3B).
Furthermore, immunohistochemical analysis with antibodies against the neuronal marker protein β-tubulin III revealed an increased neuronal differentiation in L1 overexpressing cells 7 days after withdrawal of FGF-2 and EGF in the GFAP-L1 and GFAP-L1-i groups as compared to WT NSCs, whereas the percentages of GFAP-positive astrocytes were decreased in GFAP-L1 NSCs and GFAP-L1-i cells as compared WT NSCs (Fig. 3C). In addition, increased neuronal differentiation, decreased astrocytic differentiation, and enhanced neurite outgrowth was observed in WT NSCs co-cultured on a monolayer of GFAP-L1 NSC-derived astrocytes, when compared to WT NSCs co-cultured on WT NSCs (Supplemental Fig. 1). Differentiation into oligodendrocytes was negligible in all groups (not shown).
Ectopic expression of L1 under the human GFAP promoter enhances survival and migration of grafted cells after transplantation into the injured spinal cord
To investigate whether GFAP-L1 NSCs and GFAP-L1-i cells had a beneficial effect on functional recovery after transplantation into the injured spinal cord, the spinal cord of young adult C57BL/6J mice was compression lesioned. Four days after spinal cord injury, dissociated GFAP-L1, GFAP-L1-i, and WT NSCs in PBS or PBS alone was injected 0.5 mm rostral and caudal to the lesion site (Fig. 4A). All transplanted NSCs were derived from mice ubiquitously expressing GFP and could therefore be identified by their green fluorescence in vivo (Fig. 4B).
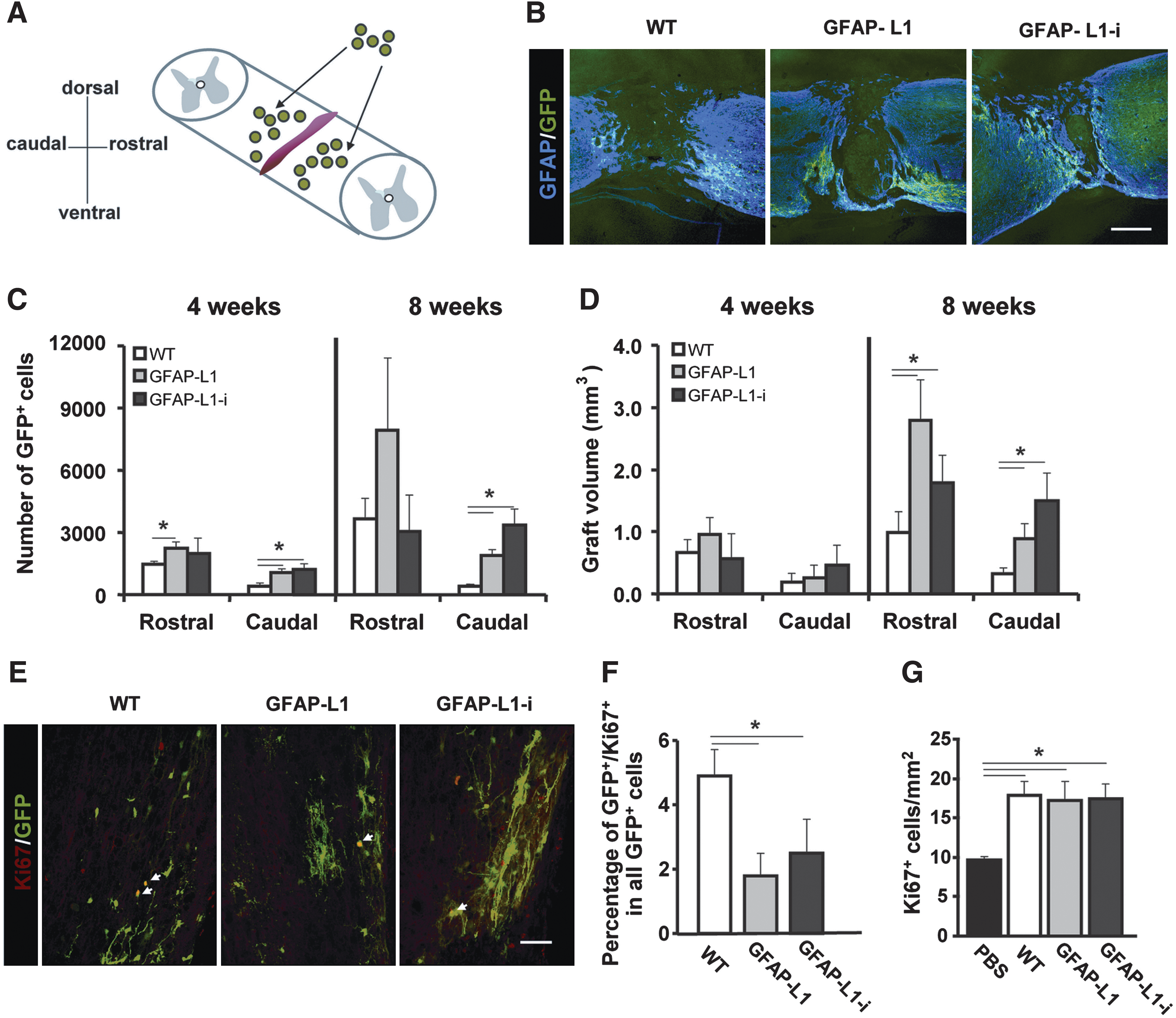
Analysis of number of grafted cells, graft volume, and characterization of proliferation of grafted and host cells after transplantation into the injured mouse spinal cord.
To assess graft survival, the number of grafted cells rostral and caudal to the lesion site was determined 4 and 8 weeks after transplantation. Four weeks after transplantation the GFAP-L1 NSC and GFAP-L1-i group showed enhanced cell numbers versus the WT group both rostral and caudal to the lesion site (Fig. 4C). Eight weeks after transplantation, the numbers of grafted-derived cells caudal to lesion site in the GFAP-L1 and GFAP-L1-i groups remained higher than in the WT group (Fig. 4C), whereas no significant differences were detected rostral to the lesion site.
Estimation of the graft volume showed no significant differences among the groups 4 weeks after injury. Eight weeks after injury, the graft volume was higher in the GFAP-L1 and the GFAP-L1-i group when compared to the WT group both rostral and caudal to the lesion site (Fig. 4D). GFAP-L1 NSCs and GFAP-L1-i cells showed decreased proliferation when compared to WT NSCs 8 weeks after transplantation and, therefore, the enhanced cell numbers in the graft and volume of GFAP-L1 and GFAP-L1-i grafts cannot be attributed to enhanced proliferation of grafted cells (Fig. 4E, F). Interestingly, both, WT and L1-overexpressing (GFAP-L1 and GFAP-L1-i) NSCs led to increased overall cell proliferation in the host tissue of the injured spinal cord when compared to control animals injected with PBS (Fig. 4G). Furthermore, the rostrocaudal and horizontal extent of the grafts defined as the longest distance between grafted cells in opposite directions from the center of the graft 4 and 8 weeks after transplantation were analyzed to estimate migration of the grafted cells. The GFAP-L1 and GFAP-L1-i grafts showed an increased rostrocaudal extent both 4 and 8 weeks after transplantation and an increased horizontal extent 4 weeks after transplantation when compared to the WT group (not shown).
GFAP-L1 NSCs and GFAP-L1-i cells show increased neuronal differentiation and neurite outgrowth after transplantation into the injured spinal cord
To monitor neuronal differentiation of grafted cells' immunohistochemical analysis with antibodies against doublecortin (DCX), a marker for immature neurons, was performed four weeks after transplantation. In mice grafted with GFAP-L1 NSCs or GFAP-L1-i cells, a significantly higher percentage of graft-derived DCX-positive cells was detected than in mice grafted with WT NSCs (Fig. 5A). Eight weeks after transplantation, immunohistochemical analysis with antibodies against the neuronal marker protein β-tubulin III revealed an enhanced percentage of graft-derived β-tubulin III-positive cells in mice transplanted with GFAP-L1 NSCs and GFAP-L1-i cells versus WT NSCs. Neurons from GFAP-L1 NSCs and GFAP-L1-i cells had significantly longer neurites than neurons derived from WT NSCs (Fig. 5B). Furthermore, overexpression of L1 showed a tendency toward decreased astroglial differentiation of grafted cells in the GFAP-L1 and GFAP-L1-i groups versus the WT group (Fig. 5C). Graft-derived CNPase-positive oligodendrocytes were rarely observed in all experimental groups (not shown).
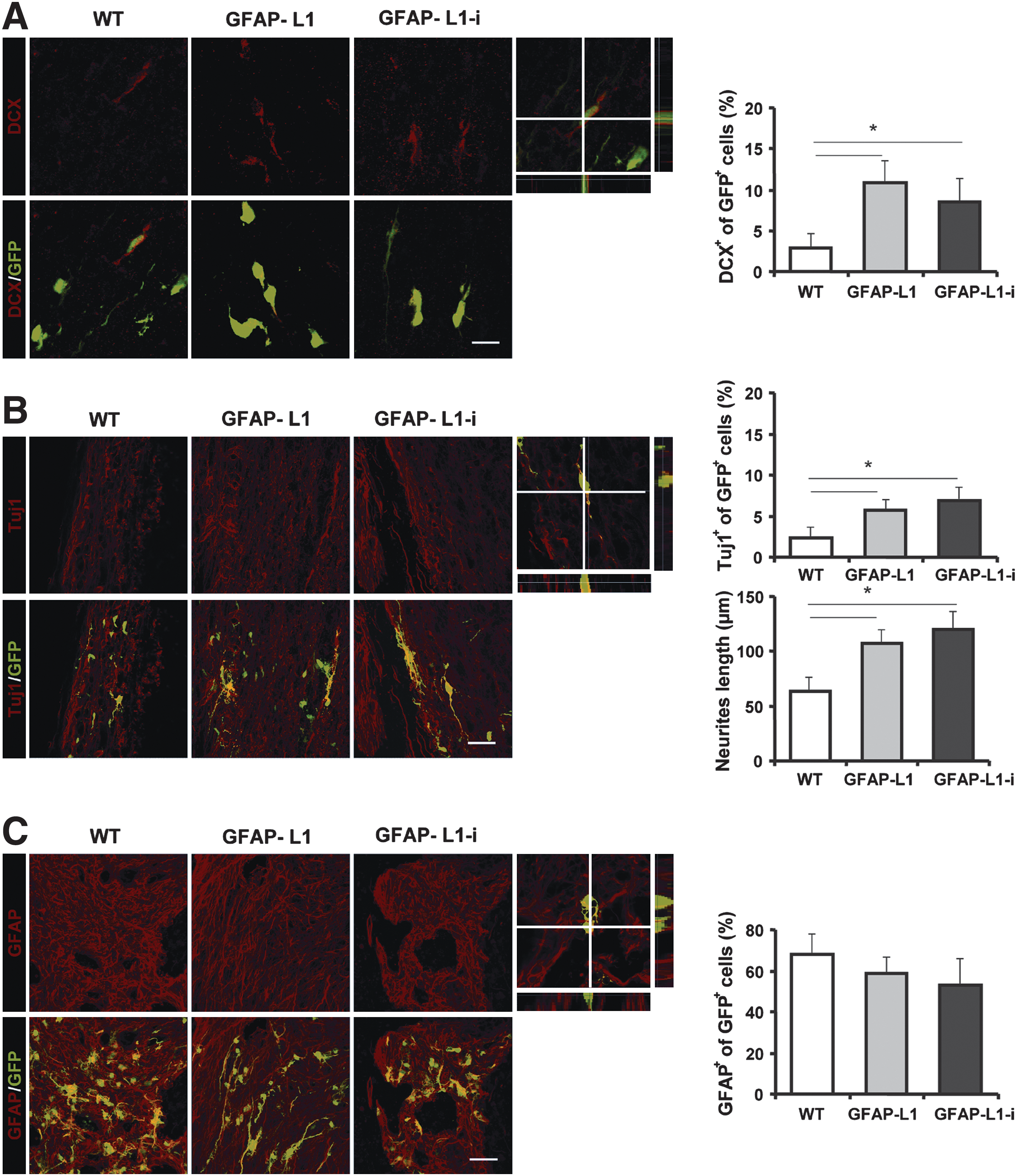
Glial fibrillary acidic protein (GFAP)-L1 neural stem cells (NSCs) and GFAP-L1-i cells show increased neuronal differentiation, decreased astrocytic differentiation, and increased neurite length after transplantation into the injured mouse spinal cord.
Ectopic expression of L1 in NSCs enhances host motoneuron soma size and synaptic input to host motoneurons after transplantation into the injured spinal cord
To analyze the influence of transplanted cells on endogenous motoneurons after spinal cord injury, the size of host motoneurons and the densities of synaptic terminals on the cell bodies of host motoneurons caudal to the lesion site were determined 8 weeks after injury. Immunohistochemical analysis revealed that the mean area of the somata of host motoneurons was significantly enhanced in the GFAP-L1 and the GFAP-L1-i versus the WT and PBS groups (Fig. 6A). Furthermore, the linear density (number per unit length) of large perisomatic ChAT-positive puncta, which have been shown to comprise C-type synapses on motoneurons associated with muscarinic type 2 receptors (Hellstrom et al., 2003), was determined. Coverage of host motoneuron somata by ChAT-positive puncta was significantly enhanced in the GFAP-L1 and GFAP-L1-i groups when compared to the control-injected (PBS) group, whereas the WT group did not significantly differ from the GFAP-L1 and GFAP-L1-i groups (Fig. 6A).
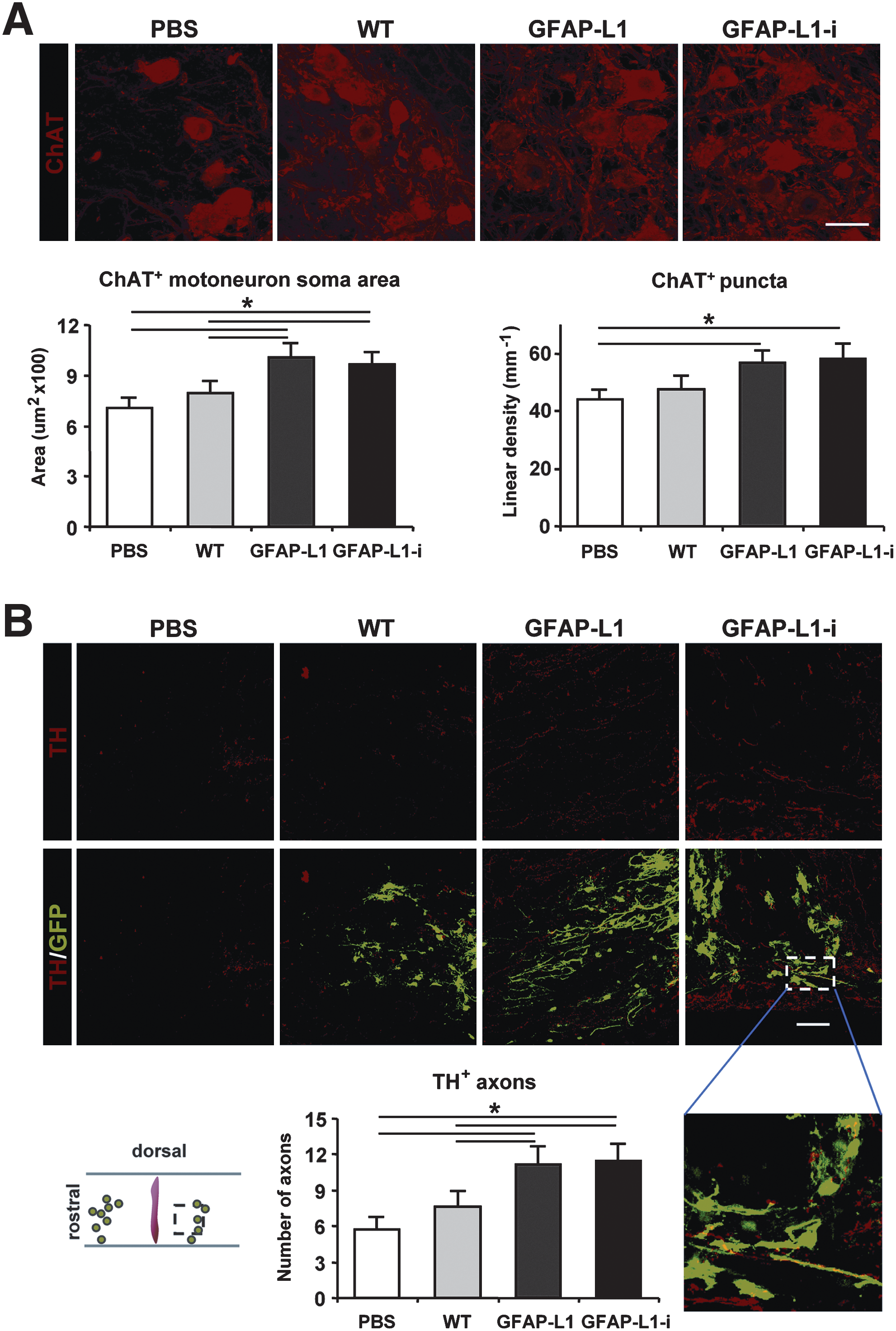
Analysis of soma size and perisomatic terminals of host motoneurons and numbers of tyrosine hydroxylase-positive axons distal to the lesion site in sections from mice transplanted with wild type (WT) neural stem cells (NSCs), glial fibrillary acidic protein (GFAP)-L1 NSCs, or GFAP-L1-i cells, or sham-injected with phosphate-buffered saline (PBS) after spinal cord injury.
Mice transplanted with GFAP-L1 NSCs and GFAP-L1-i cells show increased numbers of tyrosine hydroxylase-positive axons distal to the lesion site
Monoaminergic descending pathways located in the ventral and lateral columns of the spinal cord (Shapiro, 1997) modulate the excitability of spinal cord circuitries capable of initiating and controlling rhythmic coordinated movements (Fouad and Pearson, 2004; Jakovcevski et al., 2007). To assess whether transplantation of NSCs ectopically expressing L1 under control of the human GFAP promoter could enhance monoaminergic reinnervation of the spinal cord caudal to the lesion site, the numbers of catecholaminergic (TH)-positive and serotonergic (5-hydroxytryptamin-positive) axons caudal to the lesion site were determined 8 weeks after transplantation. The number of TH-positive fibers was increased in the GFAP-L1 and GFAP-L1-i groups when compared to the WT and PBS groups (Fig. 6B). Moreover, a tendency toward increased numbers of serotonergic (5-HT-positive) axons in the GFAP-L1 and GFAP-L1-i groups was observed when compared to the WT and PBS groups 8 weeks after transplantation. This difference was, however, not statistically significant (Supplemental Fig. 2A).
After injury, the proximal segments of the axons of the CST, which are required for voluntary movement, retracted from the lesion site (Chen et al., 2007). No significant differences were detected among the four experimental groups regarding regrowth or sparing of CST axons 8 weeks after transplantation (Supplemental Fig. 2B). Since regeneration after spinal cord injury, in particular growth/retraction of the corticospinal axons, strongly depends upon the lesion severity (Apostolova et al., 2006), we also estimated the volume of a fibrous lesion scar in all experimental groups. Lesion scar volumes, measured at 8 weeks after injury were similar among the experimental groups (Supplemental Fig. 2C). Therefore, it is expected that lesion severity was similar in all experimental groups and cell transplants did not influence the scarification process.
Transplantation of NSCs into the lesioned spinal cord does not influence the astroglial reaction
Expression of GFAP in the host tissue was measured 8 weeks after transplantation. No significant differences were detected in the host tissue among the four experimental groups 8 weeks after transplantation (Supplemental Fig. 3).
GFAP-L1 NSCs and GFAP-L1-i cells promote functional recovery after spinal cord injury
Spinal cord compression caused severe disabilities, with the degree of impairment observed 1 week after injury being similar in the four groups (Fig. 7A). Between 1 and 6 weeks after transplantation, the locomotor function was significantly enhanced in mice transplanted with GFAP-L1 NSCs and GFAP-L1-i cells when compared to mice grafted with WT NSCs and PBS-injected mice, as revealed by analysis of the BMS score values (Fig. 7A and E). In addition to the BMS score, the plantar stepping ability of the animals was analyzed by measuring the foot-stepping angle. In agreement with the BMS score, recovery in the GFAP-L1 and GFAP-L1-i groups was enhanced when compared to the WT and PBS groups 3 and 6 weeks after transplantation (Fig. 7B and F). Interestingly, the foot-stepping angle was improved in the GFAP-L1-i versus the GFAP-L1 groups 3 weeks but not 6 weeks after transplantation, indicating accelerated functional recovery in mice transplanted with GFAP-L1-i cells.

Glial fibrillary acidic protein (GFAP)-L1 neural stem cells (NSCs) and GFAP-L1-i cells enhance functional recovery in mice after spinal cord injury. Time course and degree of functional recovery after spinal cord injury in mice transplanted with wild type (WT) NSCs, GFAP-L1 NSCs, or GFAP-L1-i cells, or sham-injected with phosphate-buffered saline (PBS) are shown. Means±S.E.M of Basso Mouse Scale (BMS) scores
Furthermore, the rump-height index was determined as a measure of the ability to support body weight during ground locomotion. No differences in the outcome after transplantation were observed among the four groups (Fig. 7C and G). In addition, the evaluation of the extension–flexion ratio measuring voluntary movements without body weight did not reveal any differences among the four groups (Fig. 7D and H). From the values of the parameters shown in Figure 7E–H, we calculated, on an individual animal basis, overall recovery indices for group mean values (Fig. 7I) and each animal (Fig. 7J). This analysis revealed an overall better functional outcome in the GFAP-L1 and GFAP-L1-i groups compared to the WT and PBS groups at 3 and 6 weeks after transplantation.
Recovery of motor functions in mice is associated with elevation in the H-reflex (Lee et al., 2009). To further confirm the better functional recovery in the GFAP-L1 and GFAP-L1-i groups, electrophysiological H/M reflex recordings were taken. Because the promotion of functional recovery at 6 weeks was similar in the GFAP-L1 and GFAP-L1-i groups, only mice transplanted with GFAP-L1 NSCs, WT NSCs, and control mice injected with PBS were analyzed 6 weeks after transplantation. The H/M ratios in the GFAP-L1 group were significantly higher than in the WT and PBS groups at low frequencies (0.1–0.5 Hz, Fig. 7K) and showed a tendency toward enhanced H/M ratios at higher frequencies ranging from 1 to 20 Hz.
Discussion
Because of the morphological and functional complexity of the human spinal cord, spinal cord injury remains highly difficult to treat even under experimental conditions. As such, no single therapeutic intervention or pharmacological manipulation could be assumed to solely treat spinal cord injury. To explore potential and efficacious therapeutic strategies for spinal cord injury is not only of interest for basic neuroscience, but also for translational strategies in particular.
In the present study, we syngeneically transplanted NSCs derived from transgenic mice that had been genetically manipulated to ectopically express the paradigmatic neural cell adhesion molecule L1 under the control of the human GFAP promoter into the compression-injured murine spinal cord, to monitor the effects on functional recovery. It has been shown that L1 is not expressed in astrocytes and other glial cells in the CNS but by the regeneration-conducive Schwann cells in the peripheral nervous system with their remarkable feature to promote the regrowth of severed axons. Remarkably, the recent observation (Li et al., 2010) that adhesion molecules are important determinants for the mechanically guided lineage selection and differentiation of stem cells, adds to the concept that adhesion molecules are essential for stem cell biology. Therefore, the study of L1 functions in distinct conditions of application is a prerequisite for further steps in therapeutic application.
The human GFAP promoter has been shown to be active not only in astrocytes but also in radial glial cells in the developing mouse brain (Liu et al., 2006; Platel et al., 2009) and has successfully been used to isolate radial glial cells from the developing rodent brain (Malatesta et al., 2000). Therefore, the human GFAP promoter is superior to the widely used nestin promoter, which is expressed during the NSC stage only, but not in astrocytes, the most prevalent progeny of cultured NSCs, because the human GFAP promoter leads to sustained expression of genes of interest in astrocytes and can be used to isolate radial glial cells. As expected, the expression of L1 directed by the human GFAP promoter led to ectopic expression in radial glia and astrocytes in our study, thus demonstrating that this promoter is a valuable tool to generally deliver beneficial genes via NSCs to the lesioned spinal cord. Immunoisolation with antibodies against L1 further promoted the successful application of L1 to increase the fraction of L1-expressing cells for transplantation. In contrast to L1-positive cells isolated from WT NSCs comprising terminally differentiated neurons, L1-positive cells isolated from GFAP-L1 NSCs were able to form neurospheres, and were multipotent, thus exhibiting stem cell properties.
Both GFAP-L1 NSCs and GFAP-L1-i cells showed decreased proliferation, increased migration, and enhanced neuronal and decreased astrocytic differentiation in vitro when compared to WT NSCs. These results are in line with previous studies of NSCs growing on substrate-bound L1 in vitro (Dihne et al., 2003) and of embryonic stem cells overexpressing L1 (Bernreuther et al., 2006). Potential mechanisms may be attributed to L1 ectopically expressed on the surface of radial glial cells and astrocytes that modify the differentiation of non-fate-determined neural precursor cells by heterophilic and homophilic mechanisms as described previously for NSCs in vitro (Dihne et al., 2003). Furthermore, L1-expressing astrocytes derived from GFAP-L1 NSCs increased neuronal differentiation and neurite outgrowth of co-cultured WT NSCs, indicating a heterophilic interaction of L1 expressed by transgenic cells with molecules on the surface of non-transgenic cells in our study.
Considering the acute effect of spinal cord injury leading to spinal shock including hypoxic/ischemic events, and venous stroke lasting for 3 days after spinal cord injury (Belegu et al., 2007), we subsequently transplanted WT NSCs, GFAP-L1 NSCs, and GFAP-L1-i cells 4 days after injury. After transplantation, GFAP-L1 NSCs and GFAP-L1-i cells showed enhanced survival when compared to WT NSCs as shown by the enhanced graft size and cell numbers in GFAP-L1 and GFAP-L1-i versus WT grafts, in spite of a decreased proliferation of grafted cells. This agrees with previous studies showing that L1 enhances the survival of neurons both as soluble and substrate-bound molecule in vitro (Chen et al., 1999) and the survival of transplanted embryonic stem cells in the spinal cord (Chen et al., 2005). The enhanced survival may be mediated via homophilic and heterophilic interactions of L1 with the cell surfaces of neighboring transplanted cells or the extracellular matrix (Brummendorf et al., 1998; Dihne et al., 2003; Rathjen et al., 1984). Furthermore, L1 can be shed from the cell surface by proteolysis (Kalus et al., 2003), thus releasing L1 into the extracellular space to act in an autocrine and/or paracrine fashion on cells that are reached within its diffusion radius. L1 has been shown to interact with ankyrin, members of the ezrin, radixin, and moesin (ERM) family, actin stress fibers, and the AP-2 adaptor complex (for review see Loers et al., 2005). These interactions with the cytoskeleton are also likely to play an important functional role in vivo in the present lesion paradigm mediating not only increased survival and neurite outgrowth but also increased migration of grafted GFAP-L1 NSCs and GFAP-L1-i cells.
Ectopic expression of L1 in NSCs under the influence of the human GFAP promoter increased neuronal differentiation and neurite outgrowth of GFAP-L1 NSCs and GFAP-L1-i cells after transplantation into the injured spinal cord when compared to WT NSCs. Our data show relatively high proportions of doublecortin expressing donor cells at 4 weeks, and β-tubulin III expressing donor cells at 8 weeks after transplantation. We have, however, seen only a few NeuN-expressing donor cells and no ChAT-, TH- or CAMKII-expressing donor cells, whereas some of them acquired a GABAergic phenotype. Although we did not design our study to look primarily for neuronal differentiation of transplanted cells, but rather to deliver L1 into the injured spinal cord using radial glial cells and astrocytes as vehicles, the apparent lack of expression of markers of differentiated neurons in grafted cells was unexpected. A possible explanation could be that in stressed cells, the expression of markers such as NeuN and ChAT is downregulated, and the majority of grafted cells were still located in the vicinity of the lesion site, where the expression of NeuN and ChAT in the host cells was downregulated as well. Previous studies showed that transplanted NSCs differentiate predominantly into a population comprising astrocytes with both beneficial and inhibitory properties for CNS repair (Davies et al., 2006, 2008). Although in our present study there was an increase in neuronal differentiation of grafted GFAP-L1 NSCs and GFAP-L1-i cells in vivo, the overall yield of neurons was low and therefore most probably did not contribute substantially to regeneration. This view is supported by the observation that transplantation of Schwann cells – cells that do not differentiate into neurons – that had been genetically engineered to overexpress L1 led to an accelerated functional recovery after spinal cord injury in mice (Lavdas et al., 2010). However, the predominantly non-neuronal progeny of transplanted GFAP-L1 and GFAP-L1-i NSCs, comprising mainly radial glial cells and astrocytes expressing L1, probably mediated the beneficial effects of L1. Because radial glial cells transplanted early after injury could have significantly contributed to regeneration before they, at later stages, differentiated into astrocytes, we cannot ascribe the beneficial effects of L1 solely to L1-expressing radial glia or to astrocytes, although the accelerated functional recovery mediated by the GFAP-L1-i cells comprising L1 overexpressing radial glial cells alludes to their importance in regeneration.
In our study, ectopic expression of L1 by radial glial cells and astrocytes ameliorated the decrease of cholinergic synaptic input to motoneurons after injury. These cholinergic synapses have been shown to modulate several ionic conductances responsible for repetitive discharges of motoneurons, and significantly contribute to motor behavior (Chevallier et al., 2006), thus correlating with functional recovery after spinal cord injury (Apostolova et al., 2006; Jakovcevski et al., 2007). Moreover, ectopic expression of L1 by radial glial cells and astrocytes rescued endogenous motoneurons indicated by an increased soma size. L1 has previously been shown to be a survival factor for motoneurons in vitro presumably by exerting L1-triggered survival promoting functions in these cells via activating several signaling mechanisms, including the MEK and PI3K pathways and regulation of Ca++ influx, in addition to interacting with cytokine receptors (Schuch et al., 1989). Furthermore, L1-overexpressing NSCs enhanced the regrowth of TH-positive fibers distal to the lesion site. Homophilic or heterophilic mechanisms have been shown to enhance axonal growth of L1-expressing axons that can form L1-mediated fascicles that support each other's growth. L1-expressing dopaminergic descending axons, originating in the substantia nigra, tegmentum, and reticular formation, are located in the ventral and lateral columns of the spinal cord. They mediate the excitability of spinal cord circuitries (Fouad and Pearson, 2004) which correlates with locomotor functional recovery in the injured spinal cord of mice (Jakovcevski et al., 2007). L1 has been shown to promote the survival of dopaminergic neurons (Hulley et al., 1998) and the outgrowth of tyrosine hydroxylase positive neurites (Poltorak et al., 1992) in vitro and to rescue host dopaminergic neurons in the brain after transplantation of L1-overexpressing stem cells in a mouse model of Parkinson's disease (Cui et al., 2010; Ourednik et al., 2009). Furthermore, L1 enhances neurite outgrowth (Brummendorf et al., 1998) as well as axon guidance, fasciculation, and regeneration (Castellani et al., 2000). L1 may thus help the descending dopaminergic neurons to regrow by using this molecule as a conducive homophilic interaction partner between the non-myelinated L1-expressing dopaminergic axons. Therefore, it is not surprising that L1-overexpressing astrocytes and radial glial cells enhanced dopaminergic reinnervation in the spinal cord caudal to the lesion site, thereby contributing to functional recovery. In contrast to TH-positive descending fibers, no significant differences in regrowth or sparing of CST axons that are required for voluntary movement were detected. The use of CAMKII antibodies alone to measure regeneration of the CST has certain limits, as CAMKII labels not only CST axons but also sensory axons in the dorsal spinal cord and, importantly, also the pain fibers that sprout significantly after injury (Fang et al., 2002). However, as we used parasagittal longitudinal sections, there was very little interference in immunostaining from sprouting dorsal root axons in sections where the CST was visualized. It is therefore not likely that the main conclusions of our study would have been different if conventional anterograde tracing had been used to visualize the CST.
In addition, transplantation of GFAP-L1 NSCs and GFAP-L1-i cells did not lead to reduced astrogliosis in the present study. As a previous study showed decreased astrogliosis after virus-mediated overexpression of L1 in host cells in the injured spinal cord (Chen et al., 2007), it is conceivable that ectopic expression of L1 in host cells is necessary to reduce astrogliosis.
The effects of transplantation of GFAP-L1 NSCs and GFAP-L1-i cells on functional recovery after spinal cord injury were assessed by monitoring several behavioral parameters. The current evaluation standard for functional outcome in rodent spinal cord injury after therapeutic interventions is the open-field locomotion rating scale (Basso et al., 1995, 2006; Joshi and Fehlings, 2002) although the necessity of combining the open-field locomotion rating scale score with more specific behavioral and electrophysiological assessments has been postulated (Fouad and Pearson, 2004; Kesslak and Keirstead, 2003). In the present study, to thoroughly explore the recovery of locomotor function, we measured, in addition to the BMS score, three indicative parameters: the foot-stepping angle, the rump-height index, and the extension–flexion ratio (Apostolova et al., 2006). In particular, these methods produce numerical raw data to allow a refined assessment of functional motor recovery. These data, not only presented as raw data, but also as a compiled recovery index, have always led to results in complete agreement with the well- established Basso, Beattie, and Bresnahan (BBB) and BMS scores, as also shown in the present study. The plantar stepping, a spinal function, was monitored by both the BMS score and the foot-stepping angle with similar outcomes (Apostolova et al., 2006; Jakovcevski et al., 2007). Both parameters were enhanced in mice grafted with GFAP-L1 NSCs and GFAP-L1-i cells when compared to WT NSCs and PBS-injected controls. In contrast, the rump-height index, a measure for postural reflexes influenced by supraspinal functions, and the extension–flexion ratio (Apostolova et al., 2006; Jakovcevski et al., 2007) requiring higher levels of supraspinal control, were not improved in the GFAP-L1 and GFAP-L1-i groups. Evaluated by the foot-stepping angle, locomotor activity was significantly enhanced 3 weeks after transplantation of GFAP-L1-i NSCs versus GFAP-L1 NSCs, whereas no differences were detected 6 weeks after transplantation. This indicates that GFAP-L1-i cells exert their beneficial effects at early stages after transplantation. These effects may be ascribed either to the increased amount of L1 delivered to the lesioned spinal cord by the L1-overexpressing radial glial cells in the GFAP-L1-i cell group, which had been immunoisolated from GFAP-L1 NSCs prior to transplantation, or to the described beneficial effects of transplanted radial glial cells themselves (Hasegawa et al., 2005). Because of the predominant astrocytic differentiation that did not differ between GFAP-L1 and GFAP-L1-i cells after transplantation, the difference between GFAP-L1 and GFAP-L1-i groups had most likely disappeared 6 weeks after transplantation. It is reasonable to assume that transplantation of GFAP-L1 and GFAP-L1-i NSCs into the compression-injured mouse spinal cord influences local adaptive responses intraspinally rather than enhancing recovery of voluntary movements during the time span the animals were observed, although the measured parameters cannot clearly separate supraspinal and spinal functions. This view is corroborated by the morphological analyses described previously, revealing enhanced numbers of TH-positive axons, enhanced cholinergic synaptic input (estimated by the linear density of ChAT-positive puncta on host motoneurons), and an enhanced soma size of host motoneurons distal to the lesion site while no improvement in the regrowth of the corticospinal tract was observed.
To confirm the behavioral assessments, we applied electrophysiological H-reflex recordings, a measure of motoneuronal reflex excitability (Lee et al., 2009). Consistent with previous results showing that spontaneous functional recovery after spinal cord injury in C57BL/6J mice was associated with a progressively increasing excitability of the plantar H-reflex (Lee et al., 2009), enhanced H-reflex responses at low frequencies were observed in mice transplanted with GFAP-L1 and GFAP-L1-i NSCs, but not in WT NSCs or sham-injected controls. Consistent with higher monoaminergic and cholinergic synaptic input into the lumbar spinal cord in GFAP-L1 and GFAP-L1-i NSCs injected mice, the finding of enhanced H-reflex responses in these mice suggests that better functional recovery may be mediated via enhanced motoneuronal excitability. This is in line with our previous studies that have demonstrated that augmented functional recovery in tenascin-R- and CHL1-deficient mice was associated with an enhanced H-reflex response, whereas worse functional outcome in tenascin-C-deficient mice was associated with ameliorated H-reflex responses (Chen et al., 2010; Lee et al., 2009).
In summary, we provide not only in vitro beneficial effects of ectopically expressed L1 in glial cells, but, most importantly, also novel in vivo evidence that ectopic expression of L1 in radial glia and astrocytes promotes improved functional recovery of locomotor behavior, when compared to previously studied applications of this beneficial molecule. Our study not only provides evidence that expression of a regeneration-conducive molecule under control of the human GFAP promoter represents a novel strategy for a more general presentation of molecules in NSCs, but underscores the conducive effects of homophilic binding of L1 in the relationship between astrocytes and neurons, which is a prominent driving force in regeneration in the peripheral nervous system of adult mammals.
Footnotes
Acknowledgments
We are grateful to Yulia Tereshchenko for breeding GFP transgenic mice, Eva Kronberg for animal care, and Hyun Joon Lee for help with the H-reflex recordings. This work was supported by the BMBF (01GZ070), the New Jersey Commission for Spinal Cord Research and the Li Kashing Foundation (to M. S.). M.S. is consultant at the Center for Neuroscience at Shantou University Medical College, China.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.