
Abstract
After spinal cord injury (SCI), precise assessment of motor recovery is essential to evaluate the outcome of new therapeutic approaches. Very little is known on the recovery of kinematic parameters after clinically-relevant severe compressive/contusive incomplete spinal cord lesions in experimental animal models. In the present study we evaluated the time-course of kinematic parameters during a 6-week period in rats walking on a treadmill after a severe thoracic clip compression SCI. The effect of daily treadmill training was also assessed. During the recovery period, a significant amount of spontaneous locomotor recovery occurred in 80% of the rats with a return of well-defined locomotor hindlimb pattern, regular plantar stepping, toe clearance and homologous hindlimb coupling. However, substantial residual abnormalities persisted up to 6 weeks after SCI including postural deficits, a bias of the hindlimb locomotor cycle toward the back of the animals with overextension at the swing/stance transition, loss of lateral balance and impairment of weight bearing. Although rats never recovered the antero-posterior (i.e. homolateral) coupling, different levels of decoupling between the fore and hindlimbs were measured. We also showed that treadmill training increased the swing duration variability during locomotion suggesting an activity-dependent compensatory mechanism of the motor control system. However, no effect of training was observed on the main locomotor parameters probably due to a ceiling effect of self-training in the cage. These findings constitute a kinematic baseline of locomotor recovery after clinically relevant SCI in rats and should be taken into account when evaluating various therapeutic strategies aimed at improving locomotor function.
Introduction
So far, little is known about the recovery of kinematic parameters following partial SCI in rat, especially after contusive or compressive lesions. Primary observations showed that after a moderate spinal contusion at C7, T9 or L2 level, rats walking freely on a transparent flat surface adapt their locomotion by increasing the stance period probably to improve weight support, lateral balance and propulsion which compensate for the postural impairment induced by SCI (Collazos-Castro et al., 2005; Collazos-Castro et al., 2006). Indeed, when these deficits are compensated by a bipedal robotic device, contused rats decreased the stance length and the paw elevation during swing (Nessler et al., 2006). However, the number of steps is not correlated with functional scores after spinal contusion in rats and this parameter does not seem to be relevant in evaluating the locomotor recovery in contrast to weight bearing and interlimb coordination which are highly related to locomotor performance (Nessler et al., 2006). This last aspect is critical because the coupling between fore- and hindlimbs is a key parameter to reach higher BBB scores. Determining such interlimb coordination based on visual inspection can be misleading since there can be progressive shifts between the fore and hindlimbs which are difficult to detect by eye. In addition, partial spinal lesion models, especially contusion or compression, may entail a wide range of locomotor impairments even in animals submitted to the same injury protocol. Some of the changes might be diluted in a multivariable scale and therefore it is important to evaluate quantitatively some discrete parameters (e.g. phase coupling, foot placement, joint excursion,…) and establish group averages so that these can be compared with other assessment methods such as footprint analysis, grid test and narrow beam test or even BBB scores (Chang et al., 2010; Fouad et al., 2000; Nessler et al., 2006; Pereira et al., 2009).
To our knowledge, no study has addressed the recovery of locomotion following spinal cord compression using detailed hindlimb kinematics. In the present study, we used a modified aneurysm clip (Rivlin and Tator, 1978) to produce a clinically relevant moderately severe thoracic spinal cord compression injury in rats (Poon et al., 2007). In this context our main objective was to precisely document the time course of locomotor recovery during a 6 week period after the spinal clip compression using high-resolution kinematic analysis on treadmill. Secondly, we assessed the effect of daily treadmill training on the recovery of these locomotor parameters. Precise evaluation of the kinematic features after partial SCI could provide a much needed baseline against which to compare the beneficial or deleterious effects of various therapies on the evolution of hindlimb locomotion in a context of clinically relevant spinal lesion model. These data are also necessary to assess the relevance of short-term treadmill training and other approaches to influence activity-dependent plasticity in rodent models with SCI and other therapeutic interventions.
Methods
Animal care
Twenty-two adult female Wistar rats (250–275 g) from Charles River Laboratory (Quebec, Canada) were involved in this study and altogether, 19 rats survived until the end of experiment. Animals were housed in standard plastic cages at 22°C before spinal cord injury (SCI) and 25°C after SCI in a 12:12h light/dark photoperiod. Food (Agribrands Purina®, Ontario, Canada) and drinking water were available ad libitum. Hardwood sawdust bedding (PWI brand, Quebec, Canada) was used before SCI and then was replaced by soft paper bedding (Diamond Soft Bedding #7089, Harlan™ Teklad) after SCI to prevent skin lesions. Animals were examined daily and evaluated by a veterinarian when necessary. After SCI, the bladder was expressed three times daily until the recovery of spontaneous bladder function, which generally occurred between 7 and 14 days post-injury. All the animal procedures included in the present study were approved by the University of Montreal Research Ethics Board and were conducted according to the Canadian Guide to the Care and Use of Experimental Animals (Canadian Council on Animal Care).
Experimental design and groups
After a one-week period of quarantine in the animal facility, rats were habituated to walk consistently on the treadmill at different speeds. Three weeks later, kinematics baselines were recorded for all rats during a second three week period. Then, animals were randomized into two experimental groups: 1) Trained group (n=10), in which a specific treadmill training program was provided after SCI and 2) Untrained group (n=10), which was not trained on the treadmill after SCI (one rat died in this group 1 week before the end of experiment). A spinal cord clip compression injury was then performed on each rat and, afterwards, kinematics was recorded weekly during six weeks. Finally, spinal cords were harvested for histology. The chronological sequence of experimental procedures is presented in Figure 1B. Except for treadmill training sessions and data recording, all rats could move freely in their cage.
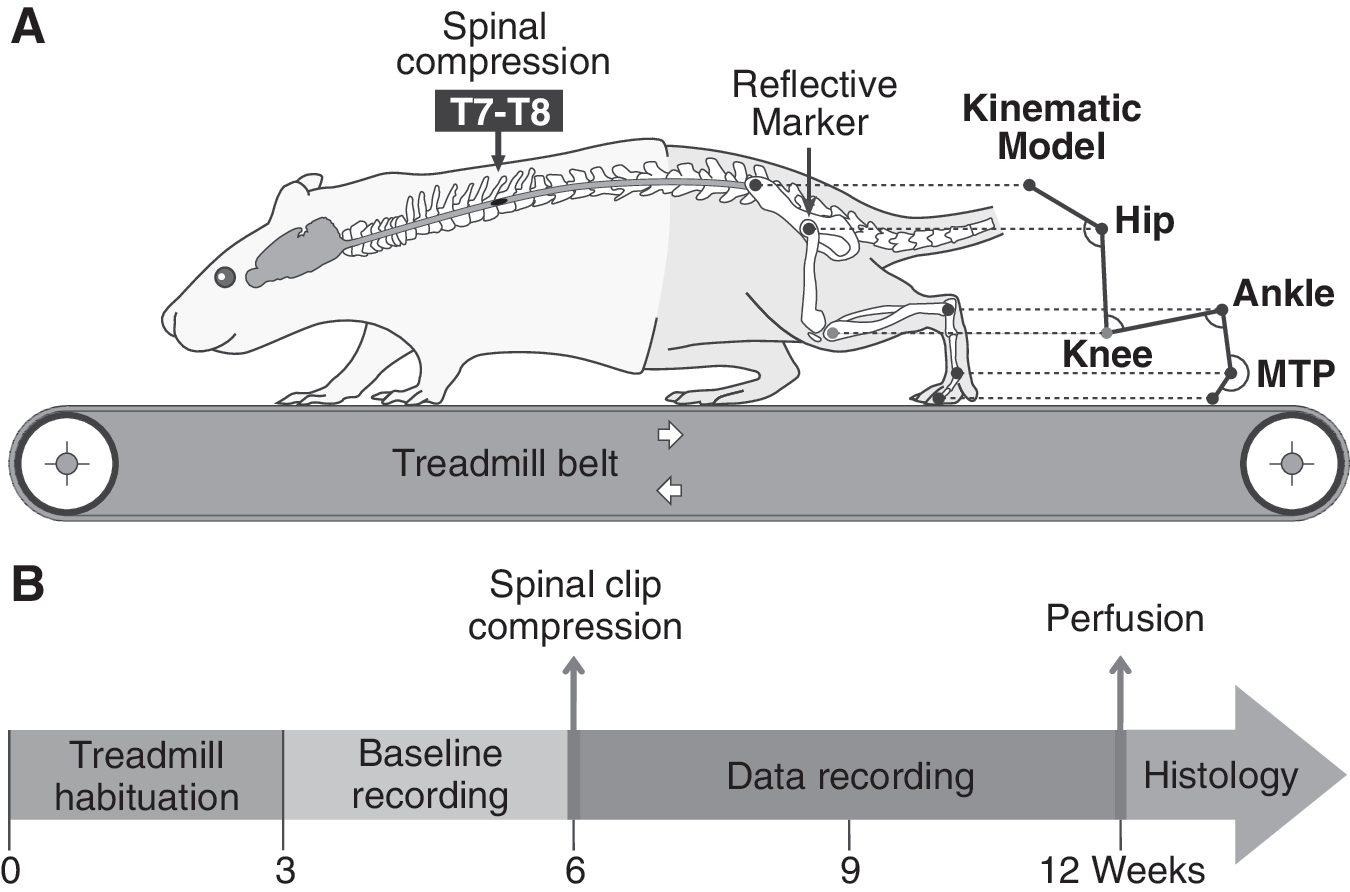
Schematic overview of the experimental protocol.
Treadmill apparatus and habituation
The treadmill belt was laid under a Plexiglas box (Length=41 cm×Depth=9.3 cm×Height=14 cm) with a removable top. All rats were progressively familiarized to walk consistently on the treadmill at speeds ranging from 14 m.min–1 (low) to 30 m.min–1 (high) during the three weeks training program before SCI. Each session lasted 15 minutes and the number of sessions depended on the ability of each rat to perform the task. A soft plastic stick was used when necessary to stimulate the rats and induce appropriate locomotion by touching their hindquarters. Progressively, rats learned to walk freely and regularly in the locomotor device using all 4 limbs at each speed of the range defined above.
Surgical procedures
All surgeries were performed in aseptic conditions and under general gas anesthesia consisting in a mixture of O2/isoflurane (1–4%) given through a mask integrated in a surgical stereotaxic frame. Immediately after surgery, rats were placed under a heating lamp until they fully recovered consciousness. Animals were given systematic postoperative analgesia (40 μg/kg Temgesic®, Schering-Plough, Hertfordshire, UK) and saline (5 ml) subcutaneously to prevent pain and dehydration, then they received antibiotic in drinking water (Clavamox® drops, Pfizer Animal Health) for 5 days. During each session of surgery, animals from both experimental groups were alternately operated.
The aneurysm clip spinal compression model has been extensively described previously (Fehlings and Tator, 1995; Karimi-Abdolrezaee et al., 2004; Karimi-Abdolrezaee et al., 2006; Karimi-Abdolrezaee et al., 2010; Nashmi and Fehlings, 2001; Rivlin and Tator, 1978). Briefly, the hair of the skin over the back of the rat was shaved and disinfected with a 1:1 mixture of 70% alcohol and povidone-iodine (Betadine®, Purdue Pharma L.P., Stamford, USA). A skin incision was made above the spinal processes between T4-T10 (≈3 cm). The superficial back muscles from both sides were cut along the spine and retracted from each side of the surgical area. A laminectomy was then performed between T6-T8 and the spinal cord was compressed by inserting extradurally a calibrated 23.8 g C-shape clip for 1 min at the level of T7. Thus the clip compressed the cord from the ventral to the dorsal side and induced a moderately severe incomplete SCI (see figure 9A and B). Next, a piece of sterile absorbable gelatin sponge (1.5×1×0.7 cm, Gelfoam®, Pfizer Inc.) was placed over the dura between T6-T8 and finally, muscles and skin were sutured.
Treadmill training program
Rats from the T group were all trained to walk on the motorized treadmill belt from one day to six weeks following SCI. The training program was performed 6 days/week and consisted in a walking session of 15 min daily on the treadmill at various speeds (from 14 to 26 m.min–1) depending on the locomotor recovery level of each rat. Early after SCI, rats were incapable of autonomous hindlimb stepping, and were therefore stimulated by pinching the perineum. This evoked, in most cases, some hindlimb locomotion adapted to the belt speed but sometimes only flexion/extension alternation without plantar paw placement. During the training session, the belt speed was set at 14 m.min–1 and the trunk of the animal was manually maintained to limit the lateral imbalance as long as the animal required perineal stimulation to walk. As soon as the rat was capable to walk without perineal stimulation, the belt speed was incremented in steps of 2 m.min–1 every 2 minutes until the maximum speed tolerated by the rat was reached (i.e. in the range defined above). The weight support and lateral balance were visually assessed during the whole experiment. The trunk and/or tail of the animals were manually sustained when necessary.
Kinematic recordings
Kinematic baseline data were obtained at 14, 20 and 26 m.min–1 for each rat to obtain control values. Then, the locomotor performance of each rat was evaluated weekly for 6 weeks following SCI (Fig. 1B). Each week after SCI, the untrained group was assessed during short sessions to avoid the training effect of the evaluation itself. Before each recording session, the hindquarter of the rat was shaved and five markers were set on the skin of the left hindlimb at the bony landmarks levels of the lateral side (3 black dots from permanent marker on ilium, hip and ankle joints, and 2 reflective markers from 3M™ Scotchlite™ Reflective Tape Silver over the metatarsophalangeal joint (MTP) and the tip of the third toe, see Fig. 1A). Two additional markers separated by 10 cm were stuck on the left side of the locomotor setup (i.e. in the field of the video camera) and used to calibrate the spatial scale of the kinematics model throughout the video processing. During the recording session, a left side view of the rat walking on the treadmill was captured using a high frequency numerical video camera (120 Hz, shutter speed=2 ms, model DS-41-300K0262, Dalsa corporation, Waterloo, Canada). All the kinematic parameters described in this study were generated from the coordinates (x, y) of each marker and from the paw contact/lift events using specific custom-made software. Because, in the rat, the knee joint area is located under a voluminous muscle mass which slips under the skin, triangulation was necessary to extract the knee position from hip and ankle joint markers. The trunk and/or tail of the rats were manually supported during data recording for the whole recovery period.
Criteria to define locomotion on treadmill
Since rats in each group exhibited different levels of locomotor recovery at the end of the experiment, we first defined a simple treadmill locomotor scale consisting of 4 different scores: 0=no locomotion, 1=deficient, 2=good and 3=best (table 1). These scores were based on three criteria: 1) the number of consecutive step cycles without downtime, 2) the paw placement and 3) the bilateral alternation. Each criterion cited above includes 4 levels of performance associated with the 4 locomotor scores (see table 1 for details). When locomotor sequences on the treadmill met each of the criteria for a given locomotor score, they were associated, but as locomotor recovery is nonlinear, sometimes one sequence satisfied criteria associated with different locomotor scores. In such case, the criterion reaching the lowest performance was retained and the sequence was associated with the corresponding locomotor score. Every video of rats walking on the treadmill in each condition was evaluated according to the table 1 and given a corresponding locomotor score. Only rats that reached the score 2 or 3 at 14 m.min−1 6 week after SCI were kept for the kinematic analysis and those that reached the score 0 or 1 were considered not to have recovered locomotion.
Ground Projection of the Hip Joint.
Intra-individual variability calculation
In most studies comparing animal groups, the standard deviation (SD) or the standard error of the mean (SEM) of each group are commonly displayed on charts to represent the variability of individual data around the group mean, also named inter-individual variability. However, the intrinsic variability of each animal for the studied parameters (i.e. intra-individual variability) is rarely taken into account in the locomotion studies while this information reflects the motor control capability of animals and seems crucial to evaluate the efficiency and stability of the motor system responses after SCI. In the locomotion studies using kinematics, the group mean is usually based on the individual mean of each animal in the group and the individual mean is based on a defined number of consecutive step cycles from walking animal. In the present study, in order to represent the intra-individual variability, the coefficient of variation (CV) based on the individual SD of the phase duration parameters was calculated as following:
The CV of each animal was then averaged by group and expressed as mean±SD in charts.
Inter-limb coupling evaluation
In order to evaluate the coordination between fore- and hindlimbs (i.e. homolateral) and between the both hindlimbs (i.e. homologous) during locomotion, the inter-limb coupling was evaluated before and during 6 weeks after SCI. The coupling value (i.e. phase expressed from 0 to 1) between two limbs represents the temporal position of the ground contact from one foot relatively to the period of the simultaneous cycle from the other one. For example, a coupling value of 0.5 means that the paw contact of a limb occurs at 50 % of the step cycle of the other limb and value can be represented as 180° on a polar plot. The averaged homologous hindlimbs coupling for trained and untrained animals was represented on a polar chart in which the size of each dot is proportional to the polar dispersion equivalent to the standard deviation in the Cartesian model (Fig. 8A). After spinal cord injury the inter-limb coupling can change and the new temporal relationship between limbs is reflected in the phase measurement. However, most of the time antero-posterior decoupling occurs and the stepping frequencies of the fore- and hindlimbs become different entailing a progressive drift of the coupling values and the presence of recurring 2:1 stepping ratio (i.e. double-contact of forelimbs during one step of the hindlimbs). In this case, it is possible to discriminate different drift (i.e. decoupling) levels calculated on sufficient number of consecutive steps. In the present study, when 2:1 (forelimb:hindlimb) stepping ratio occurred for the first time in the stepping sequence, we subtracted 1 to the phase value of the second forelimb step and the following ones until the next occurrence of the 2:1 stepping ratio in which 2 was subtracted from the phase of the second step, 3 for the third 2:1 ratio and so on until the end of the sequence. The converted polar values were then plotted against the number of forelimb step cycle in scatter chart as represented in the figure 8C and the linear regression was calculated for each animal. The slope value of the regression line representing the drift velocity (i.e. decoupling level) was then averaged by group before and 6 weeks after SCI.
Tissue processing and analysis
At the end of experiments, rats were deeply anesthetized with sodium pentobarbital (80 mg/kg, intraperitoneally) and then were perfused transcardially with 60 ml of cold phosphate buffered saline (0.1
Statistical analysis
Individual kinematic data from each animal at every time point and speed was averaged from a minimum of 10 consecutive locomotor cycles. Analysis of linear data and regressions were performed using PASW18 software (SPSS Inc., Chicago, USA). Chi-Square test (χ 2) was used to test the difference in the frequency of observations from groups at each time point. A paired t-test was used to compare data from different time point (i.e. * symbol in graphics when H0 is rejected) or velocity (i.e. + symbol in graphics when H0 is rejected) in each group and unpaired t-test was used to compare data from each group at different time point (i.e. # symbol in graphics when H0 is rejected). Unpaired t-tests were also used to compare morphometric data of the spinal cord lesion (i.e. lesion cavity and residual tissue). Paired comparison of polar data was analyzed using Watson-William F-test (Oriana 3.13, Kovach Computing Services, Pentraeth, UK). Paired comparison of regression slope and intercept from cycle duration plotted against swing or stance duration was done using Centurion XVI software (Statgraphics, Warrenton, USA). Grouped data are reported as means±SDs, SEMs or polar dispersion in polar plots and the significance threshold was set to p≤0.05 (represented by *, # or + in charts), p≤0.01 (represented by **, ## or ++ in charts) and p≤0.001 (represented by ***, ### or +++ in charts).
Results
Overview of locomotor recovery
At the end of the experiment, only one rat in each group recovered partial weight bearing during locomotion and none of the rats recovered lateral balance. External stabilization of the trunk and/or manual support of the tail were thus used to assist the rats during training (for trained animals) and the kinematic recording sessions (for all animals) throughout the recovery period. We used the simple locomotor scale defined in the materials and methods part of the present article to globally assess the hindlimb movements of the rats during the locomotor sequences and discriminate objectively those exhibiting a walking pattern from others.
Figure 2 shows the averaged locomotor levels of trained and untrained rats reaching levels 0 or 1 and those reaching levels 2 or 3 before and 6 weeks after SCI at 14 m.min–1. The first week after SCI none of the rats were able to walk on treadmill corresponding to the zero level of the scale. At this time, the rats moved using only the forelimbs and exhibited a total flaccidity of the hindlimbs. From two to six weeks following SCI, the locomotor level of most animals increased progressively. At the end of study, around 75% of the rats in both groups exhibited a stepping pattern of the hindlimbs with plantar foot placement when manual support and/or stabilization was provided (solid lines in Fig. 2) while around 25% producing only flexion-extension alternation towards the back with no paw placement (dash lines in Fig.2). For these few rats the locomotor recovery was very poor and their locomotor level remained close to 0 until the end of the study. Only rats reaching the levels 2 or 3 at 14 m.min–1 6 weeks following SCI were kept for the kinematics analysis. This corresponded to 75% of the rats i.e. 7 for trained group and 8 for untrained group represented by the solid lines and full circle symbols in figure 2. This allowed us to focus on rats which had the potential to recover their locomotor capability after spinal cord compression. No significant difference was found between trained and untrained rats for the locomotor level during the whole recovery period. Moreover, in agreement with the study of Fouad et al. (2000), no relevant difference was found between trained and untrained animals for all the kinematics parameters measured throughout this study.
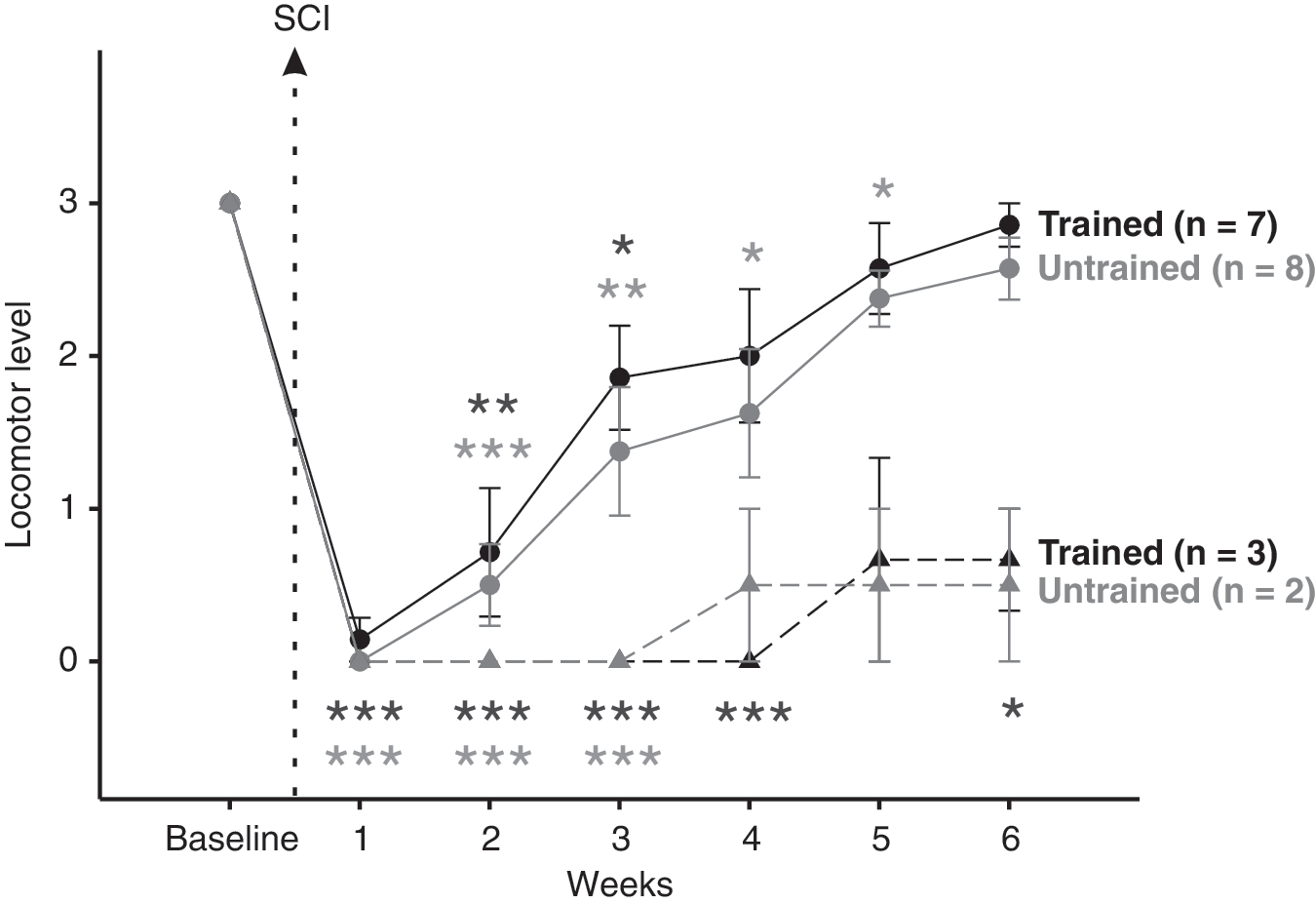
Averaged treadmill locomotor level at 14m.min−1 and at different time point following SCI. The time course of averaged locomotor level representing an overview of the treadmill locomotor recovery was recorded for trained and untrained groups using scale defined in table 1 (see details in the part entitled: Criteria to define locomotion on treadmill). Two subgroups were identified and correspond to animals associated with failing or no locomotor capability (dash line) and those associated with good or best locomotor pattern (solid line) at 14 m.min–1 6 weeks post-SCI. Only rats reaching the level 2–3 were kept for kinematic analysis. Evaluation of the averaged locomotor level in both trained and untrained groups showed no significant difference between two groups all along the post-SCI period. The absence of statistical difference with baseline at the later data points for trained 0–1 and untrained 0–1 subgroups are explained by the low number of animals (i.e. n=3 and n=2 respectively). Data are expressed as mean±SEM, * symbol represents data point compared to baseline.
Change in the angular excursions of the hindlimb joints
The minimum and maximum peaks of angular excursion and total amplitude angular excursion were measured for the hip, knee, ankle and MTP joints before and during 6 weeks after SCI. Angular excursion of the hindlimb joints and corresponding stick figures from a representative rat at each time point are given as an overview in Figure 3 while the detailed data of both groups during the post-injury period are reported in Figure 4.
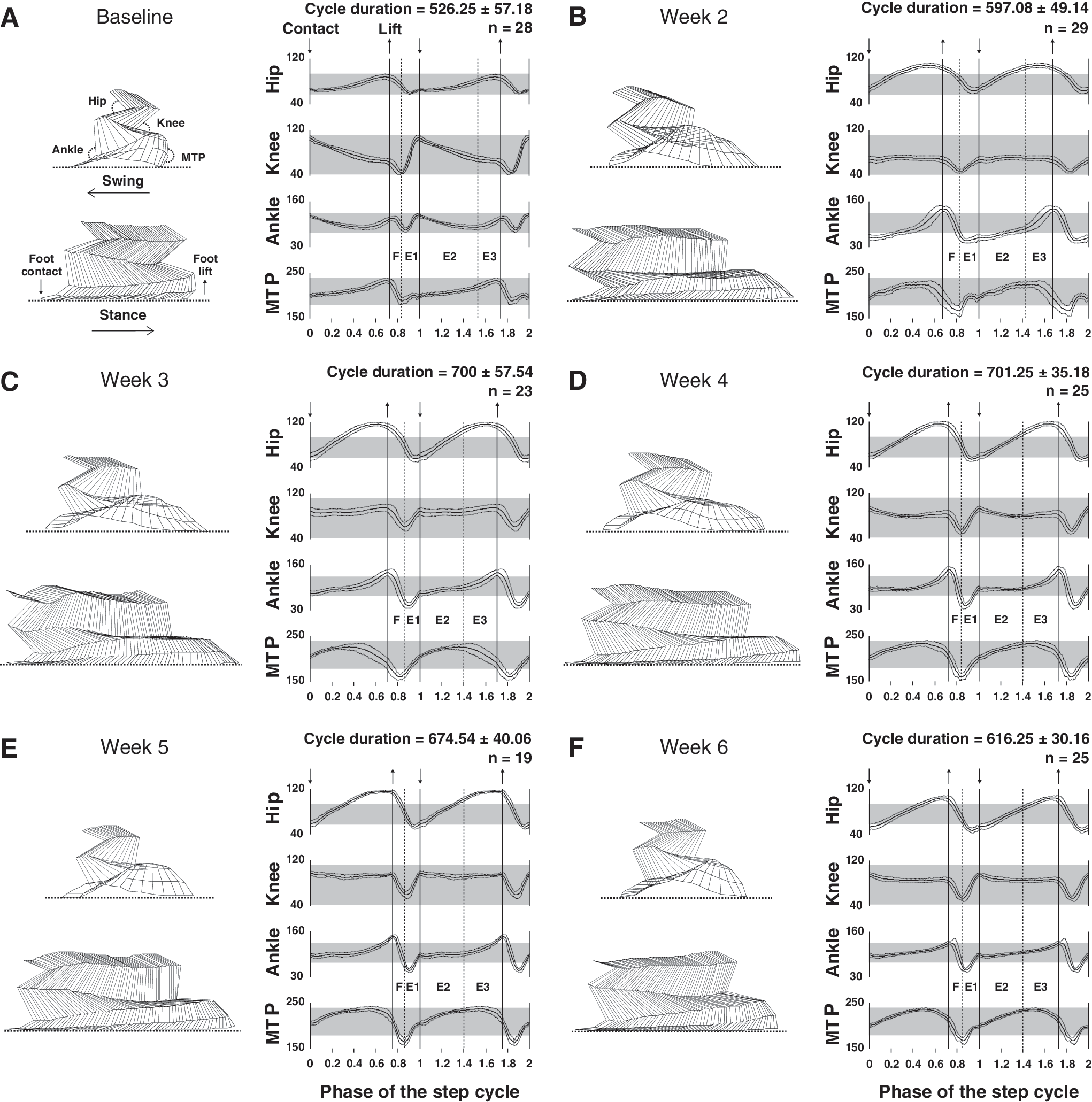
Locomotor pattern of a representative rat before and at different delays after SCI.
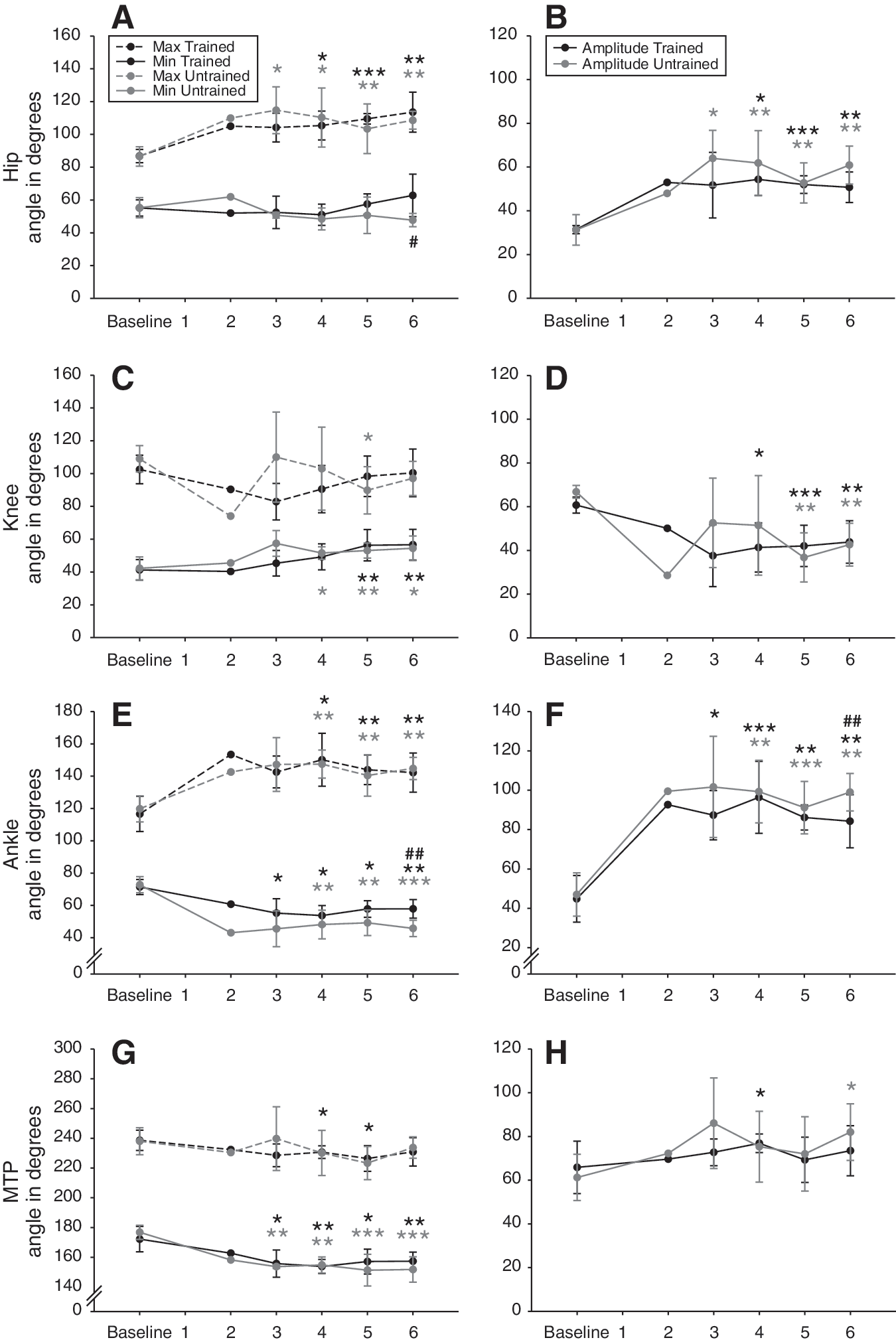
Angular excursion of the four hindlimbs joints.
As soon as rats were capable to walk on the treadmill after SCI (i.e. 2 weeks), persistent abdominal hypotonicity led to anteversion of the pelvis (i.e. increase of angle between hip and spinal column in the sagital plane) during locomotion. In addition, a deficit of the hindlimbs extension at the end of swing (i.e. E1 period of the Philippson locomotor cycle, Philippson, 1905) occurred (Fig. 3B). Consequently, the paw made contact with the ground more caudally than it normally does relative to the hip joint so that the step cycle was actually shifted backwards relative to the hip (Fig. 3B left panel).
Our results also show a general overextension of the hindlimbs at the end of stance which persisted until the end of the study as previously described by Fouad et al. (2000). Moreover, a short inactive period (≈9 % of the total stance duration), in which the back of the foot dragged passively on the treadmill belt, occurred at the stance to swing phase transition (represented by the last sticks at the end of the stance in Fig. 3B and C left panel) and disappeared at week 4. These combined factors tended to increase the stance phase duration during the whole recovery period (left panels in Fig. 3B, C, D, E and F). The swing was also affected by an important paw drag 2 weeks after SCI (Fig. 3B left panel) which decreased progressively to the point of disappearance by week 6.
During the recovery period, the locomotor pattern improved gradually. As early as 3 weeks after SCI, the general extension of the hindlimbs increased at the foot contact especially hip, knee and ankle joints (Fig. 3C right panel). In addition, the trajectories of each joint were more homogenous at around 4 weeks and close to the baseline at the end of study (Fig. 3F left panel).
Hip joint
At 14 m.min–1 the hip joint amplitude increased immediately after SCI to reach a plateau maintained until the end of experiment and significantly higher to baseline 6 weeks post-SCI (p≤0.01; Fig. 4B). This variation of angular amplitude is explained by a greater extension of the hip joint angle during locomotion which also plateaued until 6 weeks post-SCI (p≤0.01; Fig. 4A). These data are consistent with the overextension of the hindlimbs during the transition from stance to swing phase describe above. The flexion value remained significantly constant from baseline to the end of experiment. At 20 and 26 m.min–1 the angle excursion recovery of the hip joint was equivalent to 14 m.min–1.
Knee joint
Contrary to the hip, the knee joint amplitude decreased instantly after SCI, then it fluctuated until 4 weeks to eventually stabilize and remained lower than the baseline 6 weeks post-SCI (p≤0.01; Fig. 4D). In parallel, the flexion of the knee angle progressively decreased until 5 weeks and remained lower than the baseline 6 weeks post-SCI (p≤0.05; Fig. 4C). Although the maximum knee angle value showed a high variability between animals during 4 weeks after SCI, no difference with baseline was found at 6 weeks (Fig. 4C). The decrease of knee amplitude at 14 m.min–1 (around – 35 % 6 weeks post-SCI) is due to the decrease of flexion capability during the F period (Philippson, 1905) while the extension is not significantly affected. At the highest speeds the amplitude of the knee was also decreased in the same proportion but the extension was decreased during E1 at the end of the experiment (p≤0.05 at 20 and 26 m.min–1) and became thus primarily responsible for the decrease of amplitude contrary to the observations at the lowest speed.
Ankle joint
The ankle angle was the most affected during treadmill locomotion after SCI. Amplitude increased immediately and drastically to reach values much greater than baseline at 4 weeks and maintained until 6 weeks (p≤0.01; Fig. 4F). The maximum flexion and extension of the ankle are both increased after SCI and they jointly participated to the increase of ankle amplitude until 6 weeks post-SCI (p≤0.01 for minimum and maximum ankle values; Fig. 4E). At 20 and 26 m.min–1 the evolution of the minimum, maximum and amplitude data of the ankle angle following SCI were similar.
MTP joint
MTP amplitude was less affected by SCI compared to the other hindlimb joints and its value tended to slightly increase all along the recovery period (Fig. 4H). The flexion of MTP joint during locomotion increased after SCI and remained larger than the baseline from 3 weeks to the end of study (p≤0.01 at 6 weeks; Fig. 4G). The value of the maximum angle tended also to decrease during the experiment but this change remained not significant at the end of the recovery period (Fig. 4G). At 20 and 26 m.min–1 the evolution of the MTP angle was similar to 14m.min–1.
Change in the length and duration of the step cycle
Figure 5 shows the timing and length characteristics of the locomotor cycle from both groups at 14m.min−1.
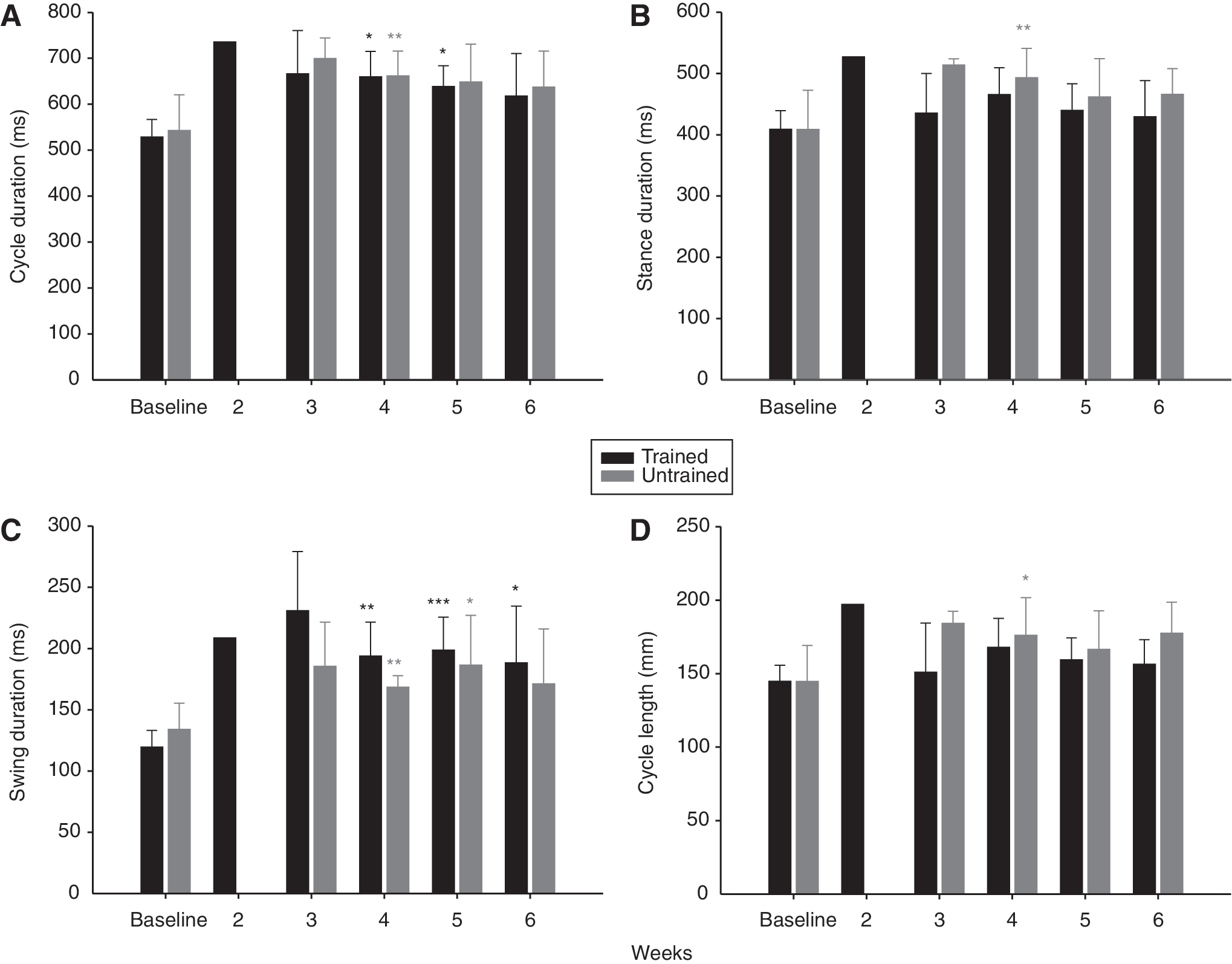
Evolution of the duration and length features of the step cycle after SCI at 14m.min−1.
Step cycle, swing and stance phases duration
Early after SCI, the duration of step cycle, stance and swing tended to increase at 14m.min−1 (Fig. 5A, B and C) as well as the highest treadmill velocities studied. From 4 weeks following SCI, the step cycle, swing and stance durations were significantly greater than baseline at 14m.min−1 (Fig. 5A, B and C). Step cycle, swing and stance duration then returned progressively toward the baseline values during the recovery period (Fig. 5A, B and C) although the duration of all components of the cycle tended to remain higher at the end of study probably due to the residual overextension of the limb at the phase transition. At 20 and 26m.min−1 the global profile of step cycle, swing and stance duration recovery was similar to 14m.min−1 with an increase of values as soon as rats were capable to walk on treadmill after SCI and a progressive return towards the baseline values until the end of study.
Step cycle length
The increase of step duration early after SCI is associated with a tendency to increase step cycle length (Fig. 5D). As for step cycle duration, the step length returned toward the baseline value but a non-significant trend to increased step cycle length present in both groups 6 weeks after SCI remained (Fig. 5D). At the speeds of 20 and 26m.min−1 the general shape of step cycle length recovery was analogous to 14m.min−1. An increase of step length occurred early after SCI. After the peak value was reached, a return toward the intact step length occurred from 4 weeks after SCI and no significant difference with baseline was found at the end of the recovery period.
Relationship between step cycle duration and stance/swing duration
The relationship model (i.e. regression line) between swing/stance duration plotted against step cycle duration for baseline, week 3 and week 6 following SCI at 14m.min−1 is given in Figure 6. Before injury, the intrinsic variation of the step cycle duration was mainly regulated by the adjustments of the stance phase (full circle in Fig. 6A) while the duration of the swing phase shown only little change relative to the variation of the step cycle duration (trained slope=0.1 and untrained slope=0.19; Fig. 6A). These data confirm the stability of the swing duration (≈130 ms) whatever the change of step cycle duration in a normal state. After compressive SCI, the relationship of the swing/stance versus step cycle duration changed so that there was a greater influence of swing in cycle time variation at the expense of stance. Three weeks post-SCI the slope of the swing/cycle duration relationship was increased compared to the baseline (p≤0.001; Fig. 6B) while the slope of stance/cycle duration was decrease (p≤0.001; Fig. 6B). At the end of the recovery period these modifications of the relationship between swing/stance and cycle duration persisted toward a significant higher influence of swing in the regulation of the step cycle duration (p≤0.001; Fig. 6C) and no significant difference with data from 3 weeks following SCI was found. Furthermore, no significant difference was found between groups for the phases/cycle duration relationship during all the post-injury period.
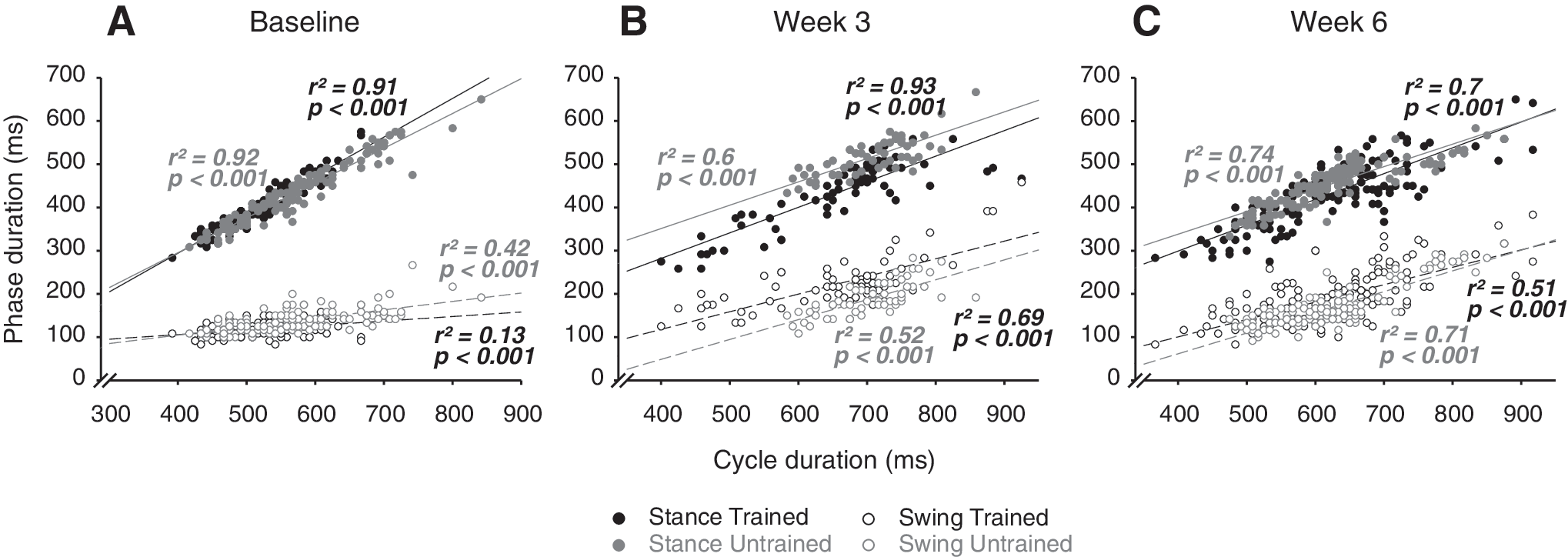
Temporal relationship between swing and stance phases versus step cycle. The raw data from the stance (full circles) and swing (empty circles) phases duration of each animal are plotted relative to the raw data from step cycle duration for trained (black circles) and untrained (grey circles) groups, before
Change in the intra-individual variability of the swing and stance phases duration
The averaged intra-individual variability of the stance and swing duration before and 6 weeks following SCI is given for each group in Figure 7.
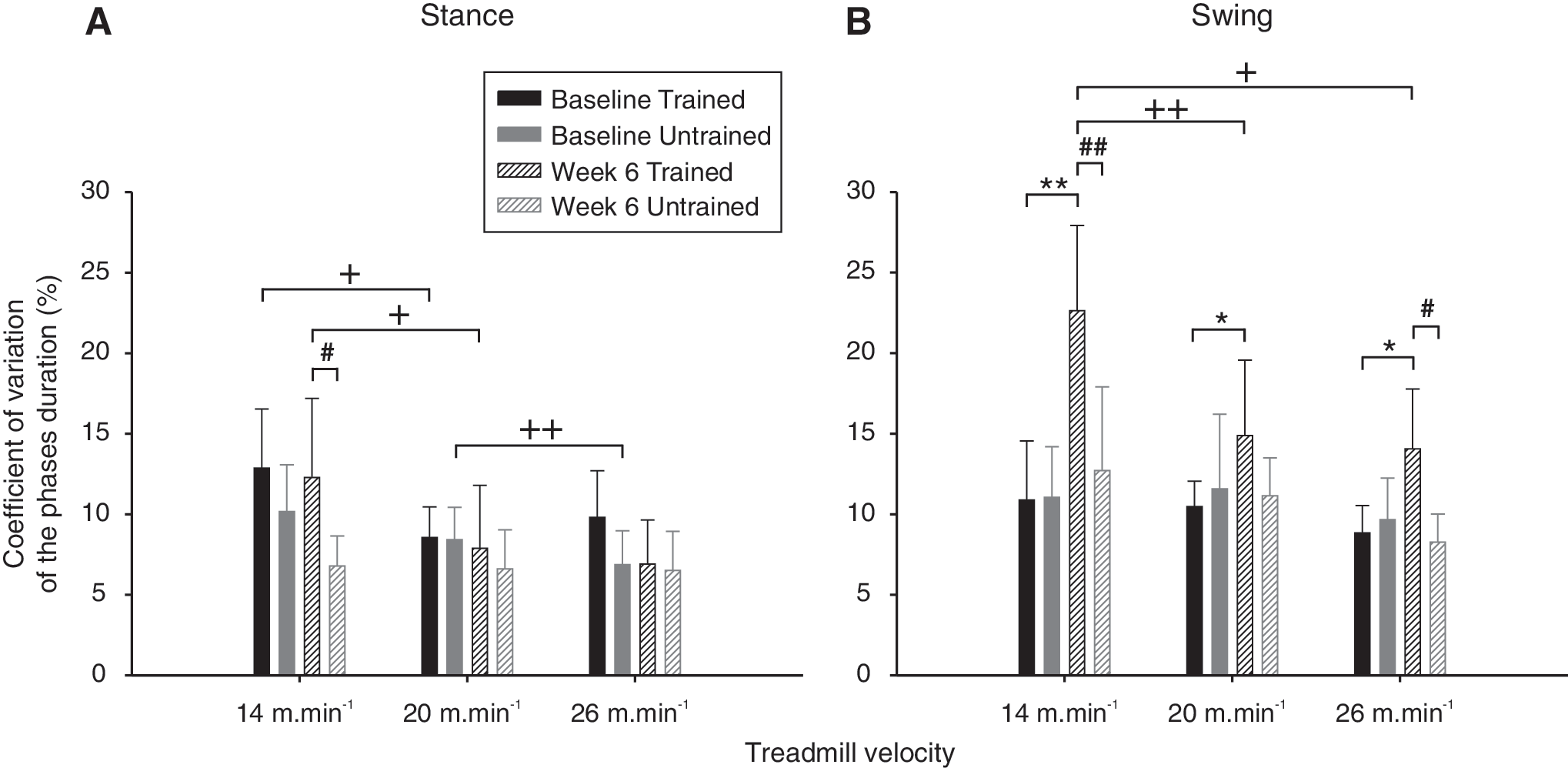
Duration variability of the swing and stance phases.
In intact state, the variability of the stance and swing duration was homogenous for each of the treadmill velocity studied (Fig. 7A and B). After SCI the trend has changed and the stance variability became significantly lower than the swing in both groups (p≤0.05; Fig.7A compared to B) and was significantly higher in trained group than in untrained group at the lower speed (p≤0.05; Fig. 7A). In addition, no difference was present for the stance variability at all velocity in untrained group while this parameter tended to decrease when the speed increased in trained group, especially from 14 to 20m.min−1 (p≤0.05; Fig. 7A).
After SCI the swing variability became significantly higher in trained compared to untrained group at 14 m.min−1 (p≤0.01), 26 m.min−1 (p≤0.05) and remains higher but non-significant at 20 m.min−1 (fig. 7B). As for the stance, the swing variability of the injured trained group decreased significantly when the velocity increased especially from 14 to 20 m.min−1 (p≤0.01) and from 14 to 26 m.min−1 (p≤0.05) while no difference was present in untrained group (Fig. 7B). In addition, conversely to the stance, treadmill training increased the intrinsic swing variability at all speed after SCI as shown in Figure 7B. Taken together, these findings suggest that swing variability become velocity- and activity-dependent after spinal compression.
Change in coordination during locomotion
The averaged homologous coupling of the hindlimbs is reported in Figure 8A while the left homolateral coupling is illustrated in figure 8B, C and D for the velocity of 14 m.min−1.
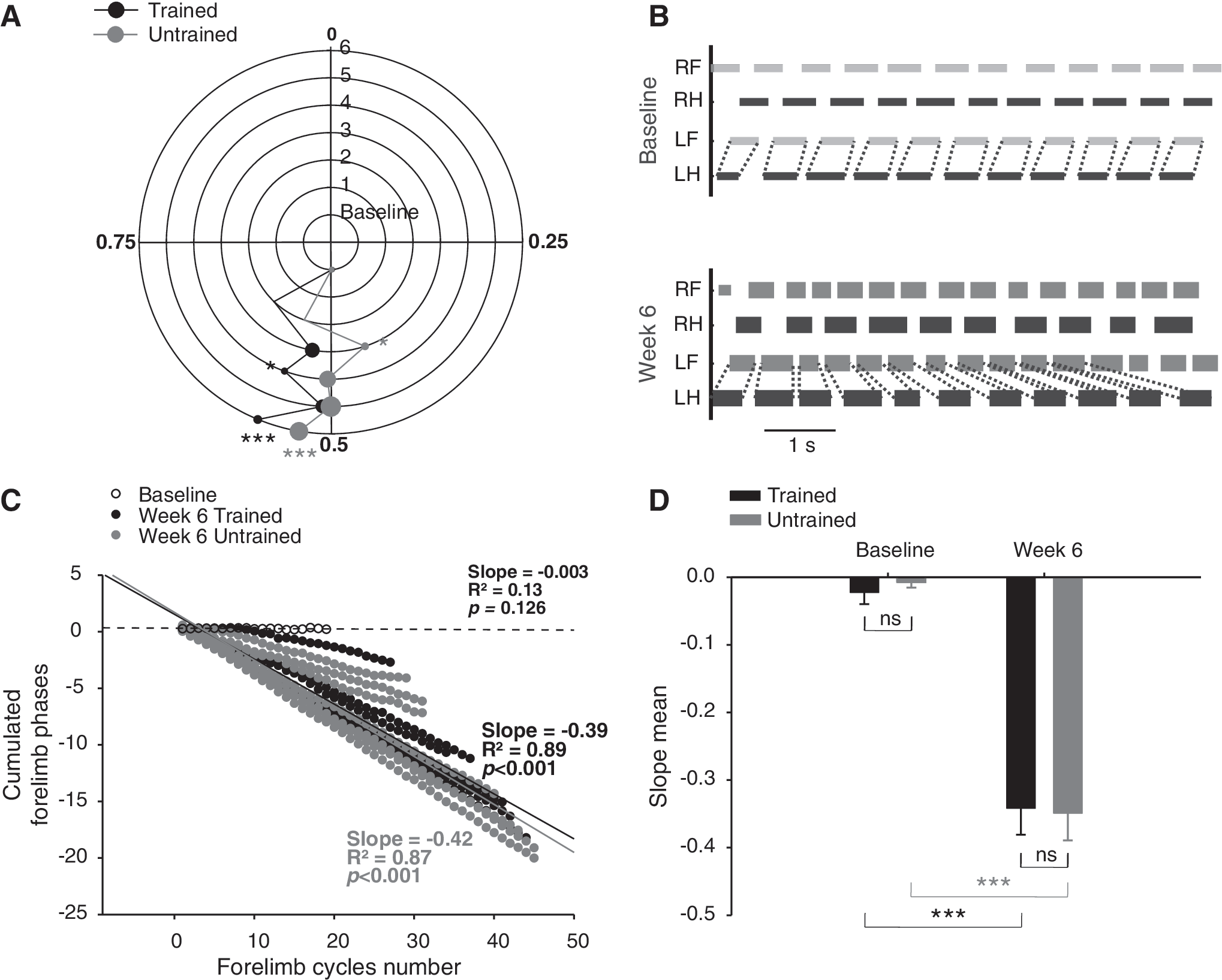
Homologous and homolateral coupling during locomotion. Polar plots in
Before SCI, the paw contact of one hindlimb occurred at the middle of the contralateral hindlimb step cycle as reported in the figure 8A. One week after SCI, only sporadic and isolated hindlimb flexions and extensions movements occurred in some rats but neither coordination nor locomotor pattern were present in either group. As soon as rats were able to exhibit a walking pattern on the treadmill, a return of the hindlimb alternation occurred immediately but a shift of the homologous coupling was present in both groups until week 5 (Fig. 8A). After hovering around the normal hindlimb coupling value of 0.5 during 5 weeks, both groups presented a significant shift of the homologous coupling towards a higher value at the end of study (p≤0.001; Fig. 8A).These inconsistent coupling values all along the post-injury period were probably due to the postural deficits increasing the coupling variability rather than specific adaptive mechanisms.
The homolateral coupling was around 0.3 in normal rats (baseline values in Fig. 8C) and the stepping ratio was 1:1 all along the locomotor sequences (Fig. 8B top panel). Although the SCI model in the present study was a partial lesion, the rats never recovered a constant 1:1 homolateral stepping ratio after the thoracic spinal compression. This deficit is illustrated by the drift of the duty cycles relationship between left fore- and hindlimb from a representative rat in the bottom panel of the figure 8B and by the linear regularity of the phase dispersions from each animal at week 6 in Figure 8C. However, different levels of decoupling were measured into groups. For instance, the progressive coupling drift of the less decoupled animal of the study was slow and the proportion of cycles with 1:1 ratio could be as great as 89% in a sequence of 27 forelimb steps (represented by the black full circles line closest to the baseline in the figure 8C) wrongly suggesting that locomotion was coupled when it was visually assessed during short and non-consecutive sequences of overground locomotion. Animals from both groups were significantly decoupled at the end of the recovery period compared to the baseline and no significant difference was present between groups at the different delays (Fig. 8D).
Morphometric analysis of the spinal cord lesion
We performed morphometric assessments to quantify the degree of cavitation and tissue preservation seven weeks after compressive SCI in both trained and untrained groups (Fig. 9). Table 2 gives values for the mean±SEM percentage of cavity size and residual tissue for each examined distance in trained and untrained groups. Our results revealed no significant difference in the size of lesion cavity or the area of residual tissue at any distance between trained and untrained groups (Fig. 9E). In addition, no major difference was noted in the virtual representation of transversal lesion extent between rats involved in the kinematic analysis and those not retained, in both cases the lesions could be considered as severe (Fig. 9F). Damaged tissue, composed of cystic cavity and astrocytic scar (Fig. 9D) were localized in the central, ventral, ventrolateral and to a lesser extent in the dorsal part of the spinal cord (Fig. 9F) affecting principally the vestibulo- and reticulospinal descending fibers. The rubro- and dorsal corticospinal tracts were also affected. Quantification of regional spared white matter (SWM) at epicenter of the lesion revealed no significant difference between trained and untrained groups in the dorsal and ventral areas (Fig. 10A). However, analysis confirmed that the ventral part of the spinal cord was significantly more affected by the clip compression than the dorsal part (p≤0.01; Fig. 10A).
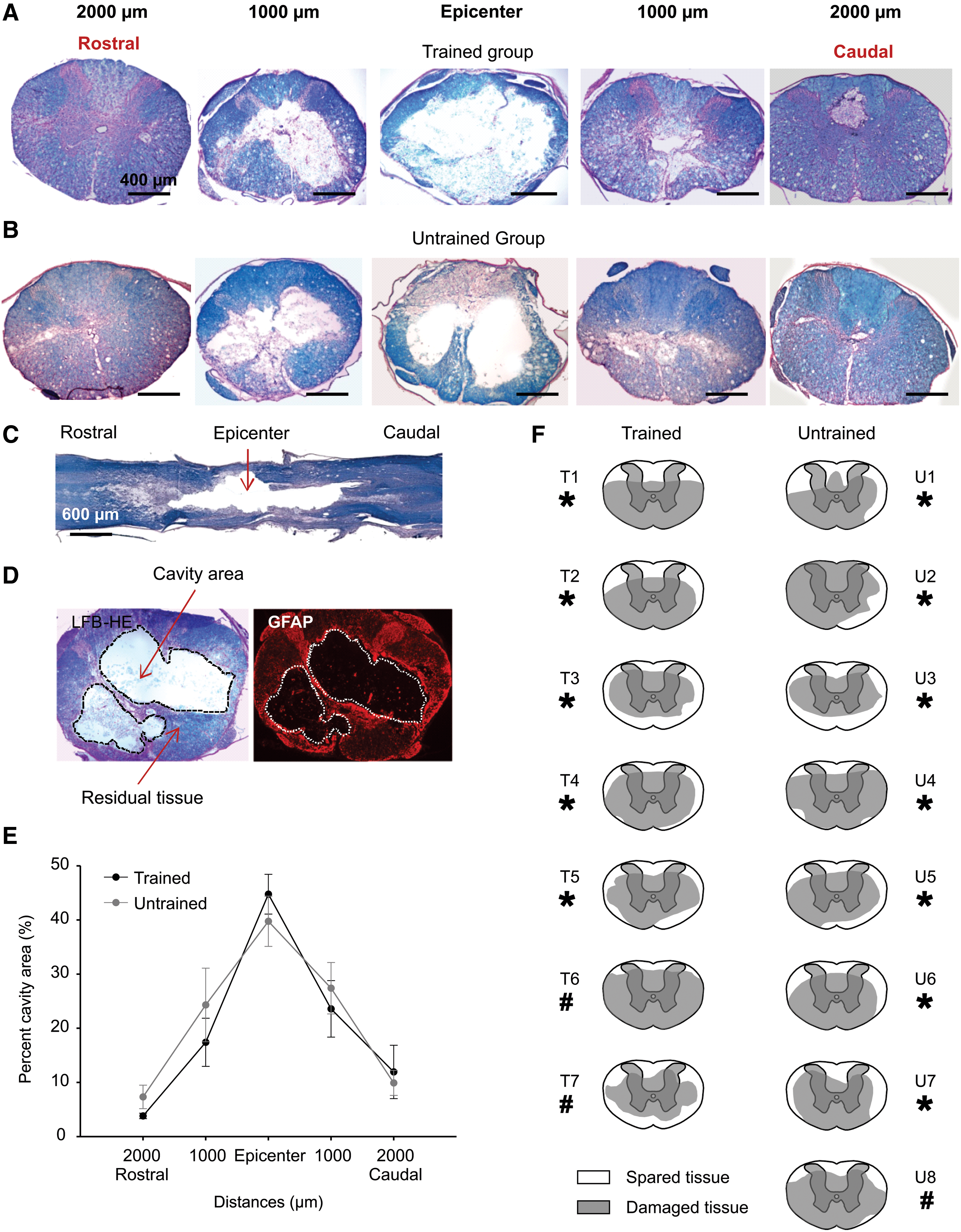
Histological assessment of the SCI lesion.

Spared white matter and relationship with kinematic variability. Bar chart in
Data are expressed as Mean±SEM.
Relationship between lesion extent and behavioural outcomes
Given that there was no difference between groups in dorso-ventral spared tissue and the full rostro-caudal extent of the lesion, linear and polynomial (order 2) regressions were performed on data, regardless of the treatment groups, to explore the relationship between regional SWM (dorsal, ventral, left and right) and behavioural outcomes. No significant relationship was found between the value of kinematic parameters at the end of the recovery period and the local or full SWM at the epicenter of the lesion. However, the variability level of homologous coordination between hindlimbs and hindpaw left contact position was significantly related to the total amount of SWM (p≤0.01; Fig. 10B and C). The variability of the hindlimb contact position is best related with the SWM by a curvilinear regression (Fig. 10C; R2=0.77, p≤0.01) while the best estimation of the relationship between coordination variability and SWM is linear (Fig. 10B; R2=0.54, p≤0.01). These relationships suggest that greater individual variability occurred when the area of total SWM decreased.
Discussion
Evaluating precisely the evolution of locomotor characteristics after SCI is essential to define a baseline for evaluating the benefit of the future studies focused on therapeutic strategies. Obviously, rats after spinal cord clip compression injury have some clear recovery of locomotor capabilities and any therapeutic interventions should demonstrate a clear improvement above that seen in the lesioned untreated group. We show here that after a moderately severe thoracic spinal clip compression SCI in rats, some consistent and robust spontaneous hindlimb locomotor recovery occurs even with defective lateral balance and weight bearing assuming that some external compensatory stabilization is provided during walking. Moreover, rats never recovered the antero-posterior coupling after SCI but we found that different levels of decoupling were present and they were indistinguishable using only a visual evaluation. Finally, although treadmill training had little effect on the recovery of locomotor parameters, the increased variability of swing in the trained group suggest that compensatory neural mechanisms may be activity-dependent and could perhaps lead to an even better recovery if applied for longer time periods.
Postural alterations after spinal clip compression
Contusive devices as the OSU, NYU or Horizons impactors strike the dorsal part of the spinal cord and damage the dorsolateral region and the grey matter to various extents depending on the impact severity. Due to the dorsal approach of such devices, the ventral and ventrolateral parts of the spinal cord are may be less affected. However, in our model, the spinal clip compresses the entire spinal cord between two curved blades and the whole cord is compressed transversally. Our histological results show that the ventral, ventrolateral and mediolateral parts of the spinal cord were severely damaged by the clip compression while the dorsal and dorsolateral parts were injured to a lesser extent (Fig.10A). Consequently, the characteristics of the moderately severe spinal clip compression model could be related to the impairment of the balance control, usually attributed to the vestibulospinal pathways and the deficit of the weight bearing partially supported by the reticulospinal pathways, both located in the ventral region of the spinal cord in rat (Fehlings and Tator, 1995; Fouad and Pearson, 2004; Rossignol et al., 2009; Schwartz et al., 2005).
Recovery of weight bearing and lateral balance is a prerequisite to the re-expression of efficient autonomous locomotion. Rats with partial dorsal section of the spinal cord at mid-thoracic level recover locomotion and weight bearing quite well even if a postural deficit remains present during stepping (Ballermann et al., 2006; Fouad et al., 2000). When the lesions are more extensive, exogenous body weight support is needed to allow locomotion and it can be provided by manual assistance as in the present study or by mechanical devices with added pharmacological or electrical stimulation (Boulenguez and Vinay, 2009; Courtine et al., 2009). However available bearing devices allow only bipedal hindlimb stepping (Courtine et al., 2009; de Leon et al., 2002; Ichiyama et al., 2009; Timoszyk et al., 2005). Such devices may change the biomechanical configuration of the body and probably introduce modifications of the spinal network excitability increasing afferent inputs due to the overextension of the hip. In addition, it has been reported that manually supported locomotor training provide more robust recovery than automated stepping devices (Fouad and Pearson, 2004; Harkema, 2001; Hillyer and Joynes, 2009).
Change in locomotor features and compensatory strategies
Accordingly to previous work on rats with partial spinal cord injury (Basso et al., 1995; Fouad et al., 2000), one of the most evident kinematics deficits after spinal cord clip compression is the ensuing overextension of the limbs at the swing/stance transition. This could be a consequence of the weight bearing deficiency associated with a lesion of the corticospinal tract as suggested in previous study in rats with a bilateral dorsal transection (Ballermann et al., 2006). In fact, the deficit in the range of motion of hindlimb joints angle not only led to an increase of the extension at the end of the stance but also entailed a decrease of hindlimb extension at contact resulting in earlier stance onset in the back of normal contact position (Collazos-Castro et al., 2006). Taken together, these kinematic adaptations resulted in a shift of the whole locomotor cycle toward the back of the hip and a maintained capability to match the treadmill belt at 14 m.min−1.
In contrast to cats with partial lesions (Barrière et al., 2008; Barriere et al., 2010; Brustein and Rossignol, 1998), rats, early after partial SCI, tend to increase the cycle and phases duration concomitantly with a tendency to increase the step cycle length (Heng and de Leon, 2009). Although this increase was non-significant at the end of the recovery period in the present study, a trend towards higher values persisted. The prolongation of the weight support period and consequently, the overextension of the hindlimbs at the end of stance are probably primarily responsible for the increase of the step cycle duration in rats with deficient weight bearing. In addition, some observations suggest that all rats in the present study were self-trained and it has been shown that training increase the stance phase in fully spinalized rats (Cha et al., 2007) suggesting similar compensatory mechanisms.
In the intact rats the swing duration is fixed during locomotion while the variation of the stance length (and consequently stance duration) allows the step-to-step adjustments to meet the belt velocity (Fig. 6A). In the present study, the structure of the step cycle was modified after spinal compression and particularly the relationship between phases and step cycle durations switching toward a greater involvement of swing in the regulation of the step cycle duration at the expense of stance. This finding could be a consequence of decrease in reflex responses of flexor muscle afferents at the swing to stance transition as previously demonstrated in fully spinalized cat (Frigon and Rossignol, 2008). Indeed, the stretch of specific flexor muscles during locomotion (increasing group I afferents activity) promote the onset of swing (McCrea, 2001). Moreover, in normal cats, activation of group I extensor afferents during stance inhibit the initiation of swing to maintained the stance phase until the onset of loading on the contralateral limb (McCrea, 2001). This precise balance between sensory adjustments could be impaired after severe spinal clip compression and this would explain the delay of the swing onset entailing a prolongation of ground contact period. Consequently, the duration of stance would become related to the swing duration as reflected by their almost parallel regression lines in figure 6B and C.
After partial SCI, the interlimb coupling could be differently affected depending on the localization and the extent of the lesion. Although the corticospinal and propriospinal tracts are implicated in the synchronization between fore- and hindlimbs, the coordination between limbs from the same girdles (i.e. homologous coupling) is supported by the Central Pattern Generator (CPG) itself. Consistently, in cat with very large lesion of the dorsal and dorsolateral part of the spinal cord, affecting essentially the cortico- and rubrospinal pathways, the antero-posterior coupling was abolished and the step frequency in both girdles was independent while the coupling between both forelimbs and both hindlimbs was preserved (Barrière et al., 2008; Barriere et al., 2010). Similarly, the antero-posterior coupling was lost while homologous coupling was maintained in all animals of the present study for which at least a part of the corticospinal tracts was severed. These data are consistent with results from studies in rat involving different severity of spinal contusion which inevitably destroy the corticospinal tracts located in the dorsal columns (Basso et al., 1995; Basso et al., 1996). Different levels of continuous antero-posterior decoupling were measured in the present work (Fig. 8C) and some of which were low with only 10 % of 2:1 stepping ratio in a sequence of ≈30 steps indicating a slight and slow coupling drift. The stepping ratio is the only available coupling information during visual observation of locomotion and the low levels of decoupling measured here were quasi-undetectable without quantitative assessment of phase relationship between limbs especially during the short continuous sequences of ground locomotion. The objective measurement of decoupling level is fundamental to avoid serious misinterpretation of general locomotor capability when using BBB scale alone.
The coupling between both hindlimbs represent the temporal position of the paw contact of one limb relative to the step cycle duration of the contralateral limb and is expressed in phase values between 0 and 1 in a polar plot. In the present study, although the averaged homologous hindlimbs coupling was not significantly changed 6 weeks after SCI, we show that the intra-individual variability of hindlimbs coupling was negatively correlated with the area of the spared white matter at the epicenter of the spinal lesion (Fig. 10B). In addition, the same trend for the intra-individual variability of the hindpaw contact position was observed (Fig. 10C), attributing the variability of coupling in the present study to the control of the paw contact. Previous studies showed that supraspinal pathways and especially dorsal tracts are essential to the control of the paw in challenging tasks (Kanagal and Muir, 2007; Kanagal and Muir, 2009; Schucht et al., 2002; Whishaw et al., 1993). Our findings suggest that supraspinal pathways could be also necessary to finely tune the control of the limb upon contact with the ground even in standardized and repetitive treadmill locomotion. However, given the robotic gait associated with low variability in spinal transected animals for which the supraspinal intputs are totally removed, our results probably show only a part of the relationship between spatiotemporal variability of the gait and the lesion extent, excluding the extreme portions of the curves. Moreover, according to our results on swing duration variability (Fig. 7B), treatments could change the relationship between gait variability and lesion extent. However, the quantification of spared white matter does not reflect the contribution of individual tracts and does not necessarily reflect the total amount of spared axons by excluding demyelinated tissue. Axonal tracing studies (Fehlings and Tator, 1995) could be done in the future to address this point.
Absence of treadmill training effect on locomotor performance
Currently, the locomotor training on treadmill remains the gold standard method to improve the intrinsic locomotor capacities after SCI in human and animal because this rehabilitation strategy is safe, non-invasive and devoid of side effect. Even if the beneficial effects of treadmill training after SCI are suggested in human, this efficiency is non-consensual in animal after partial spinal lesion and few previous studies on rats showed unclear or contradictory results (Fouad et al., 2000; Goldshmit et al., 2008; Heng and de Leon, 2009; Multon et al., 2003; Stevens et al., 2006; Thota et al., 2001).
In the present work, the daily treadmill training did not improve the recovery of the kinematic parameters after SCI, at least over the time period chosen to analyze locomotor recovery. Given that training provides good outcomes on human with contusive spinal lesions and on fully transected cats, our results question the causes of such discrepancies with spinal clip compressed rats. Rather than a possible disparity in the basic mechanisms of spinal cord plasticity between humans or cats and rats, the absence of locomotor improvement shown here is probably due to a ceiling effect of the self-training in the cage as previously discussed (Fouad et al., 2000; Heng and de Leon, 2009; Kuerzi et al., 2010). Indeed, unavoidable use of wheelchair in everyday life by paraplegic humans entails a concomitant decrease of legs activity, afferent feedbacks and muscle mass which contribute together to amplify the locomotor impairment of the lower limbs. Similar outcomes were demonstrated recently in adult rat by the team of Magnuson (Caudle et al., 2011). These interrelated consequences potentiate the effects of treadmill training rehabilitation in human by pulling down the baseline performances of the legs. However, as soon as rats are capable to produce hindlimbs locomotor movements after partial SCI, they permanently move their limbs in the cage to satisfy their natural behaviors as drinking, eating or exploring the environment. Consequently, the CPG of these animals is continuously stimulated by proprioceptive and remnant supraspinal afferent feedbacks which might be responsible of locomotor improvement until a ceiling threshold beyond which the locomotor performance could not be increased significantly by treadmill training. In addition, in contused rats trained to swim, major swimming features as posture and hindlimbs kinematics returned closed to normal and hindlimb activity was increased (Magnuson et al., 2009; Smith et al., 2006). These results not only indicate that the locomotor behavior in the cage does not interfere with the swimming performance but also that this particular task can be optimized by specific training (Magnuson et al., 2009). Taken together, these data suggest that, as in cat, spinal network plasticity in rat is also activity-sensitive after partial SCI and reinforce the ceiling effect hypothesis.
Consequences of training on the gait variability
In healthy adults, a small step-to-step variation of locomotor parameters is present even in fixed conditions (see baseline data in Fig. 7) and reflects the intrinsic variability of the locomotor system (Hausdorff, 2005). A variety of pathology or traumatism can influence the level of the intrinsic locomotor circuitry fluctuations (for review Hausdorff, 2007). For example, deficit of the anterior cruciate ligament of the knee decrease significantly the variability of the walking pattern on treadmill (Moraiti et al., 2007). It was suggested that this decrease of intrinsic variations was related to the decrease of the system complexity which affects the ability to adjust the gait to environmental demands. In fact, this hypothesis could also be valid in fully spinal transected cats trained to walk on treadmill for which the variability of their robot-like gait is very low (Barbeau and Rossignol, 1987; Barriere et al., 2010; Rossignol, 2000). Despite the differences that exist between ligament lesion and SCI, the underlying basic neural mechanism may be similar and the decrease of sensorimotor complexity due to the spinal clip lesion could explain that untrained rats with a severe SCI showed a low variability six weeks after SCI.
A recent study in spinalized rats under quipazine showed that allowing variance in the limb trajectory during robot-assisted bipedal training resulted in the recovery of a well-coordinated pattern compared to fixed trajectory (Ziegler et al., 2010). In addition, several evidences indicate that after partial SCI the repetitive quadrupedal training on treadmill could optimize the remnant supraspinal connections as well as their cortical projections increasing their influence on the CPG (Girgis et al., 2007; Hoffman and Field-Fote, 2007; Knikou, 2010; Krajacic et al., 2010; Mano et al., 2003; Muir and Steeves, 1997; Nishimura and Isa, 2009; Raineteau and Schwab, 2001; Rossignol et al., 2004). During locomotion, the swing duration is dependent of the limb trajectory which is finely controlled by the motor system. In the present study, the increase of swing variability in trained animals might thus be explained by the strengthening of supraspinal information that would result in more fluctuations of the basic walking pattern.
Conclusions
This work provides four significant contributions to the field of spinal cord injury in rats.
Footnotes
Acknowledgments
We thank Maryse Menard for her excellent assistance with data recording and data processing; Philippe Drapeau and Claude Gagner for their technical assistance in software design and electronics as well as Desiree Schut and Jian Wang for their assistance with histological staining. This work was supported by research grants from the Craig H. Neilsen Foundation and the Christopher and Dana Reeve Foundation to Soheila Karimi-Abdolrezaee and Michael Fehlings, the Krembil Chair in Neural Repair and Regeneration (Michael Fehlings) the Canada Research Chair on the Spinal Cord and grants from the Canadian Institutes of Health Research (CIHR) to Serge Rossignol. Olivier Alluin was funded by a postdoctoral fellowship from the Multidisciplinary Team on Locomotor Rehabilitation of the CIHR (Regenerative Medicine and Nanomedicine, CIHR Strategic Initiative).
Author Disclosure Statement
No competing financial interests exist.