
Abstract
The goal of the current investigation was to evaluate the mechanisms through which administration of a selective cannabinoid-2 (CB2) agonist (O-1966) modifies inflammatory responses and helps to improve function following spinal cord injury. A comparison of motor function, autonomic function, and inflammatory responses was made between animals treated with O-1966 (5 mg/kg IP) and animals treated with vehicle 1 h and 24 h following contusion injury to the spinal cord. Motor function was significantly improved in the treated animals at each time point during the 14 days of evaluation. The percentage of animals able to spontaneously void their bladder was also greater over the entire study period in the group treated with the selective CB2 agonist. Seven days following injury there was a significant reduction in both hematopoietic and myeloid cell invasion of the spinal cord, and a reduction in the number of immunoreactive microglia. The results of the evaluation of chemokine/cytokine expression and inflammatory cell invasion also demonstrated a significant effect of treatment on inflammatory reactions following injury. Two days after injury, animals treated with O-1966 had significant reductions in CXCL-9 and CXCL-11, and dramatic reductions in IL-23p19 expression and its receptor IL-23r. Treatment with O-1966 also caused inhibition of toll-like receptor expression (TLR1, TLR4, TLR6 and TLR7) following injury. These results demonstrate that the improvement in motor and autonomic function resulting from treatment with a selective CB2 agonist is associated with a significant effect on inflammatory responses in the spinal cord following injury.
Introduction
D
Currently therapeutic options available for treatment of spinal cord injury are extremely limited. The current mainstay of medical therapy for acute injury is high-dose methylprednisolone. However, there is considerable debate as to whether the adverse effects of high-dose steroids outweigh the potential benefits of their use (Coleman et al., 2000; Hurlbert, 2000; Qian et al., 2000). Modulation of the endocannabinoid system by the administration of exogenous agonists and selective antagonists has been shown to have potential to attenuate the contribution of inflammation to secondary injury in the CNS (Zhang et al., 2009b). The endocannabinoid system consists of endogenously-produced cannabinoids, their receptors, and the enzymes responsible for their synthesis and degradation. The two most well-defined receptors are the CB1 and CB2 receptors. Cannabinoid-2 (CB2) receptors are primarily located on cells of the immune system, and can have significant influence on inflammatory responses. While CB1 receptors are primarily located at synapses within the CNS, there is emerging evidence that they also influence inflammatory responses. Previous studies in our laboratory have demonstrated the therapeutic potential for modulation of the activity of these receptors in improving outcome following spinal cord injury in the mouse (Heller et al., 2009). It should be recognized that there are differences between rats and mice in pathologic responses to spinal cord injury. The major difference is the formation of cystic cavities in the rat, which does not occur in the mouse. Rather than developing cystic cavities, mice instead develop a dense connective tissue matrix. The contribution of inflammatory cells in these two species is relatively similar. The reactions of microglia and macrophages are comparable, with a peak of invasion and activation occurring at approximately 7 days. There is a difference in the timing of T-cell invasion between these two species. In rats the peak of T-cell invasion occurs between 3 and 7 days, whereas in mice T-cell infiltration starts much later (14 days), but continues over the next several weeks (Sroga et al., 2003).
The goal of the current investigation was to determine the mechanisms through which administration of a selective CB2 agonist (O-1966) modifies inflammatory responses and helps to improve function. O-1966 is a highly selective synthetic CB2-receptor agonist. The affinity of O-1966 for CB1 and CB2 receptors was reported to be 5055±984 and 23±2.1 nM, respectively. O-1966 stimulated 35S-GTPγS binding, with an EC50 of 70±14 nM, and an Emax of 74±5% (percent of maximal stimulation produced by the full agonist CP 55,940). IV administration of O-1966 to mice failed to produce effects in the tetrad test in doses up to 30 mg/kg, consistent with its very low CB1-receptor affinity. The ED50's of the CB1/CB2 cannabinoid receptor agonists used as controls are approximately 1–2 mg/kg. Based on these characteristics, O-1966 was defined as a full CB2 agonist (Wiley et al., 2002).
Methods
Animals
A murine spinal cord injury (SCI) contusion model was utilized with 7- to 9-week-old female C57BL/6 mice weighing approximately 16–21 g (Taconic, Hudson, NY). All procedures, interventions, and animal care were done in accordance with the protocol approved by the Temple University Institutional Animal Care and Use Committee, following the National Institutes of Health Guide for the Care and Use of Laboratory Animals. The animals were housed for 1 week prior to surgical intervention for acclimation and observation. A light/dark cycle of 12 h was maintained, and the mice were allowed free access to food and water including hydrogel at all times.
Surgical procedures
The mice were weighed and then anesthetized using an IP injection of a 1:1 combination of ketamine (100 mg/mL) and xylazine (20 mg/mL) at a dose of 1 mL/kg. Once under anesthesia, the back hair was shaved, the ears and tails were definitively marked, and protective eye gel was applied. Body temperature was maintained at 37±0.5°C during the procedure and recovery period with a heating pad and lamp. The surgical site was prepped with povidone-iodine solution. The ribs were used to localize the T8–T10 laminae. Using a combination of sharp and blunt dissection, the paraspinal musculature was dissected free from the laminae between T8 and T10. The mouse was then carefully held by the lateral aspects of the T7 vertebra using Adson forceps. Using an operative microscope for better visualization, laminectomies were performed at the T8 and T9 levels using fine microscissors and laminectomy forceps. The ligamentum flavum was gently dissected free using a cotton swab. Care was taken not to injure the spinal cord and to ensure adequate width of laminectomy.
The mice were then transferred to the Infinite Horizons (IH) impactor device (PSI Inc., Lexington, KY), where they were suspended via modified Adson forceps clamped to the lateral aspect of the vertebra above and below the level of the laminectomy. The impactor tip was positioned directly above the exposed dura, and then raised to a height of 3 mm. The device was set to deliver a 60-kdyn force to the spinal cord. The actual force, displacement, velocity, and injury time was recorded, as well as injury characteristics such as the presence or absence of tail flip at the time of injury and severity of the bruise. The spinal musculature was reapproximated with 4-0 silk suture, and the dorsal fat pad placed back in its normal position. The skin was closed with clips.
A selective CB2 agonist (O-1966) was dissolved with a 1:1:18 ratio of anhydrous ethanol (Sigma-Aldrich, St. Louis, MO), emulphor, and 0.9% saline. The mice were randomly divided into drug-treated, vehicle-treated, and sham groups.
Treated animals received 0.2 mL vehicle or O-1966 (5 mg/kg) by IP injection. The choice of this dose was based upon the results obtained in previous studies of ischemia/reperfusion injury in the brain (Zhang et al., 2009a). Mice euthanized at 24 h received one injection of either drug or vehicle 1 h post-contusion. Mice which were kept over 24 h received two injections, at 1 h and 24 h post-contusion. Sham mice had lamina removed without contusion.
Postoperative care
At the conclusion of surgery the mice were given SC injections of fluid (0.9% NSS; 0.5 mL). The mice were placed in a recovery cage under a heating lamp until they were well recovered from anesthesia. All cages were kept on a heating pad on the first postoperative night. The mice were also given SC injections of fluid (0.9% NSS; 0.5 mL), and buprenorphine (0.03 mg/kg) twice daily, and antibiotics (enrofloxacin, 2.5 mg/kg) once daily for the first 3 postoperative days. The mice had their bladders emptied twice daily via the Credé maneuver until recovery of autonomic function.
Motor function evaluation
The mice were evaluated for motor function recovery using two scales for open-field assessment of locomotion: the 9-point Basso Mouse Scale (BMS), and the 17-point Basso-Beattie-Bresnahan (BBB) locomotor scale modified for mice by Dergham (Basso et al., 1996a,1996b,2006; Dergham et al., 2002). There were 14 animals included in the control group and 10 in the O-1966-treated group for evaluation of motor and autonomic function. Scoring on each of these scales relies on lower limb movement, the ability to plantar-place the hindpaw, stepping with weight support, coordination in ambulation, and trunk stability. Each mouse was evaluated on postoperative days 1, 3, 7, and 14. The investigators were blinded to treatment during the scoring.
Autonomic function evaluation
All mice had autonomic impairment with urine retention following SCI. To relieve their bladders and to assess autonomic function recovery, urine was expressed twice daily via suprapubic pressure (Credé maneuver), and urine mass was determined. The mice were considered to have recovered autonomic function once the total urine mass expressed was less than 500 mg/d for 3 consecutive days.
Evaluation of inflammatory cell invasion
The mice were anesthetized with 40 μL of a 1:1 solution of xylazine and ketamine IP, and perfused through the left cardiac ventricle with 30 mL HBSS without Ca2+ or Mg2+ (Invitrogen, Carlsbad, CA) plus 1 mM EDTA (Promega, Fitchburg, WI). The control of perfusion was assessed by the paleness level of the liver. The organs were dissected and the area of contusion on the spinal cord (1 cm around the impact site) was separated. Spinal cord tissue and the spleen were passed 5×through an 18-gauge needle and digested in HBSS with a Ca2+- or Mg2+-based enzyme solution of DNAseI (0.1 mg/mL; Roche, Indianapolis, IN) and Liberase TL (0.03 mg/mL; Roche) for 45 min at 37°C with shaking. Ten milliliters and 5 mL of enzyme solution were used for spleen and cord, respectively. Enzyme digestion was followed by adding ice-cold blocking solution (10% FCS and 10 mM EDTA in HBSS) for 5 min with 40 and 20 mL per spleen and cord, respectively.
Spinal cord samples were subjected to centrifugation at 4°C at 1100 rpm (200g) for 10 min (low brake). The tissue was pelleted and resuspended in 10 mL of 30% isotonic Percoll (GE Healthcare, Little Chalfont, U.K.) diluted with 10×HBSS without Ca2+ and Mg2+ and distilled water. The tissue was underlaid with 5 mL of 70% isotonic Percoll. Mononuclear cells were isolated from the 30/70 interphase after gradient centrifugation (2000 rpm [1000g] for 20 min, slow brake at room temperature). The collected cells were washed three times in 7×volume of FACS buffer (1 mM EDTA and 2% FCS in PBS), pelleted, and counted. For the 7-day time point 3–4 animals were pooled. Two pooled groups (n=2) were included in the drug-treated and vehicle-treated groups.
To ensure that the changes in cellular invasion were not simply the result of a systemic decrease in peripheral immune cells, the white blood cell content of the spleen in CB2-treated and vehicle-treated animals was also compared. Spleen samples were subjected to centrifugation at 4°C at 1100 rpm (200g) for 10 min (low brake). Red blood cell lysis was done with RBC lysis buffer (eBioscience, San Diego, CA) according to the manufacturer's instructions. The collected cells were pelleted and counted.
All cells were resuspended in FACS buffer and stained with anti-mouse CD16/CD32 (BD Biosciences, East Rutherford, NJ) for 5 min, and afterward with a combination of anti-mouse monoclonal conjugated antibodies (Abs): CD45-APC, CD11b-PE, Ly6G, Ly6C (Gr-1)-FITC, and CD3-PeCy7 for 30 min at 4°C. For staining control purposes the samples were also stained with the appropriate isotypes. All antibodies were purchased from BD Biosciences.
All samples were then immediately analyzed using a BD FACSCalibur and/or BD FACSCanto. Data analysis was done with FlowJo 7.2.5 software (Tree Star, Ashland, OR). For the 7-day time point, spleens from 7 animals were included in the drug-treated and vehicle-treated groups.
Evaluation of proinflammatory chemokine and chemokine receptors and toll-like receptor
RNA extraction
Anesthetized mice were perfused with 30 mL of PBS administered intracardially. The injured group consisted of 3 mice for each time point and treatment, while the naïve and sham groups consisted of 2 mice per time point. The spinal cords were removed by dissection, and 1 cm of cord around the site of injury was homogenized in Trizol, then RNA was extracted according to the manufacturer's protocol. The RNA pellet obtained was resuspended in DEPC water. RNA concentration was determined by measuring optical density at 260 nm using a nanodrop spectrophotometer.
Reverse transcription
For reverse transcription 1 μg equivalent of RNA was diluted in 16 μL of DEPC water and heated at 65°C for 5 min to denature the secondary structures. The tube was cooled immediately on ice to prevent the formation of secondary structures. Then 14 μL of master mix consisting of 300 U of Moloney murine leukemia virus reverse transcriptase, 40 U of RNasin, Moloney murine leukemia virus buffer, 500 ng of random primers, and 30 μg of BSA. Then 15 mM of dNTP mix was added to make up a total volume of 30 μL. Reverse transcription was carried out at 42°C for 90 min to obtain cDNA, which was then used for real-time PCR analysis.
Real-time PCR
The cDNA was diluted 10-fold in DEPC water. Then 4 μL of the diluted cDNA was added to 16 μL of master mix consisting of SYBR green-containing PCR master mixture (2×), and 250 nM of forward and reverse primers. Real-time RT-PCR was performed using the Stratagene Mx3005P (Agilent Technologies, Santa Clara, CA). Cycling conditions were 1 cycle at 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec, 75°C for 30 sec, 57°C for 30 sec, and a melting point determination or dissociation curve. Each sample was amplified with primers for the target and the housekeeping gene β-actin. The amount of cDNA for each target was calculated by the number of cycles required for the fluorescence emission to reach threshold level (cT value), and was normalized to the housekeeping gene. Data are expressed as fold difference to naïve cord. The primers used: CXCL-1: forward 5′-TCGCGAGGCTTGCCTTGACC-3′ and reverse 5′-GCGTTCACCAGACGGTGCCA-3′; CCL-2: forward 5′-CACAGTTGCCGGCTGGAGCA-3′ and reverse 5′-CAGCAGGTGAGTGGGGCGTT-3′; IL-10: forward 5′-CCTGGTAGAAGTGATGCCCC-3′ and reverse 5′-TCCTTGATTTCTGGGCCATG-3′; CXCL-9: forward 5′-CAAAATTTCATCACGCCCTT and reverse 5′-CCAGACAGCTGTTGTGCATT-3′; CXCL-10: forward 5′-ATTCTTTAAGGGCTGGTCTGA-3′ and reverse 5′-CACCTCCACATAGCTTACAGT-3′; CXCL-11: forward 5′-GGGCGCTGTCTTTGCATC and reverse 5′-AAGCTTTCTCGATCTCTGCCAT-3′; p19: forward 5′-TGCTGGATTGCAGAGCAGTAA-3′ and reverse 5′-ATGCAGAGATTCCGAGAGA-3′; IL-23: forward 5′-ACATTGGACTTTTGTCGGGAA-3′ and reverse 5′-AAAATCGGCAACATG-3′; and β-actin: forward 5′-TCCACCACCACAGCTGAGAGG-3′ and reverse 5′-CAGCTTCTC TTTGATGTCACG-3′.
RT2 Profiler PCR array
The expression of proinflammatory chemokines and chemokine receptors and mediators of the TLR-signaling pathway was evaluated using the RT2 Profiler PCR array based on the SYBR green-containing PCR technique. First, 20 μL of cDNA was diluted with double-distilled H2O to a total of 100 μL. The experimental mixture was prepared by mixing the following components: 1225 μL of 2×SYBR green-containing PCR master mix, 98 μL of diluted cDNA, and 1127 μL of double-distilled H2O. A total of 25 μL of the mixture was loaded in each well of a 96-well plate pre-coated with primers for the different genes. The PCR array was performed using the Stratagene Mx3005P detector. PCR cycling conditions were 15 sec at 95°C, and 1 min at 60°C for 40 cycles, followed by a melting point determination of dissociation curves. Cycle threshold values were determined by automated threshold analysis, and the results were normalized to 5 housekeeping genes, which included glyceraldehyde-3 phosphate dehydrogenase (GAPDH), hypoxanthine guanine phosphoribosyl transferase (HPRT1), glucuronidase beta (Gus-β), heat-shock protein 90 alpha (hsp90sb1), and β-actin.
Histology immunohistochemistry and cell counting
A comparison was made between O-1966- and vehicle-treated mice (5 animals in each group). The mice were euthanatized 7 days post-contusion, perfused with PBS to rinse blood from the vascular system, and fixed with a solution of 4% paraformaldehyde in PBS. A 5-mm segment of the spinal cord surrounding the epicenter of injury was extracted and stored in 4% paraformaldehyde in PBS solution for less than 24 h, then transferred to a solution containing 30% sucrose in PBS to dehydrate. After dehydration the tissue was embedded in embedding medium and frozen at −80°C until sectioning. The tissue was cut into 16-μm serial sections. Sections 1.5 mm rostral and caudal from the epicenter of the contused spinal cord were stained with primary antibody (Iba-1, rabbit anti-mouse, 1:500; Wako, Richmond, VA) overnight, and secondary antibody (FITC-goat anti-rabbit IgG, 1:500; Invitrogen) for 1 h, and then washed with PBS, followed by 4′,6-diamidino-2-phenylindole (10 ng/mL DAPI; Sigma-Aldrich) staining for 10 min. The stained tissue sections were observed with a fluorescence microscope and photographed using a digital camera. The images of the Iba-1 and DAPI staining were digitally merged. The numbers of Iba-1 and DAPI double-stained cells observed in a cross-section of spinal cord 1.5 mm rostral and 1.5 mm caudal from the epicenter of injury were counted. The surface area of the cross-section was measured using Image J software, and the number of cells/mm2 was calculated.
Results
Motor function: Open-field assessment
Results for open-field assessment of motor function on the BMS and BBB modified for mice are depicted in Figure 1. The mice in the O-1966-treated group demonstrated statistically significant improvement over the control group in both BMS and modified BBB scores for open-field assessment of locomotion at 3, 7, and 14 days post-injury.
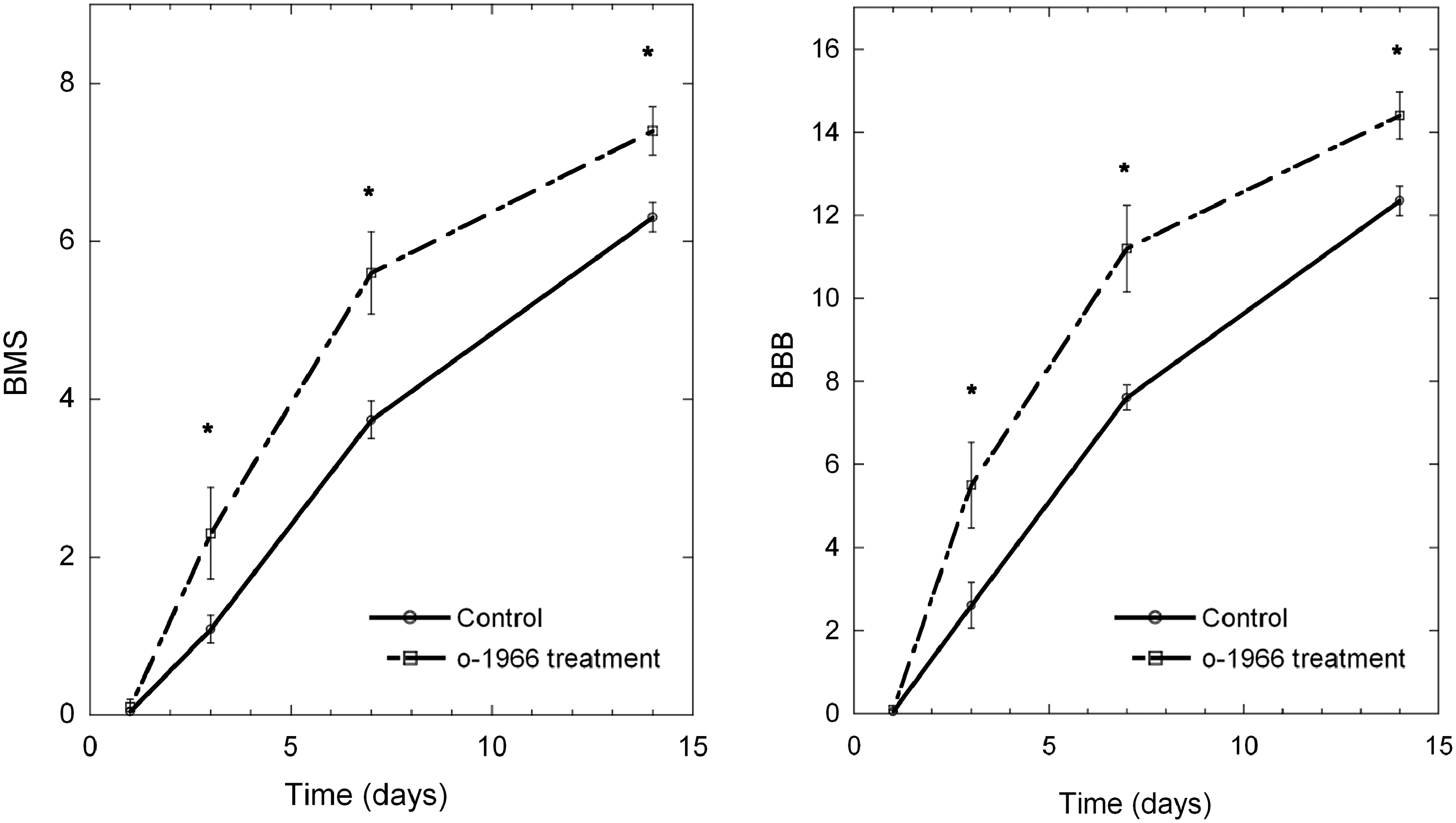
Evaluation of locomotor recovery by the Basso Mouse Scale (BMS) and the Basso-Beattie-Bresnahan (BBB) scale post-contusion, comparing vehicle-treated to CB2-agonist-treated (O-1966, 5 mg/kg) animals. Treatment significantly improved locomotor function at 3, 7, and 14 days post-contusion (*p<0.05, n=14 for the control group; n=10 for the O-1966-treatment group; CB2, cannabinoid-2).
Recovery of bladder function
Animals treated with the selective CB2 agonist had significantly improved ability to spontaneously void (Fig. 2). On the day 3 post-injury none of the vehicle-treated animals were able to spontaneously empty their bladder, while 10% of the CB2-agonist-treated animals were able to spontaneously void. By the day 7 post-injury approximately 50% of the O-1966-treated animals had spontaneous voiding, while less than 5% of the vehicle-treated animals were able to empty their bladder. By day 14 post-injury approximately 60% of the CB2-agonist-treated animals could spontaneously void, while only 25% of the vehicle-treated animals had this ability.
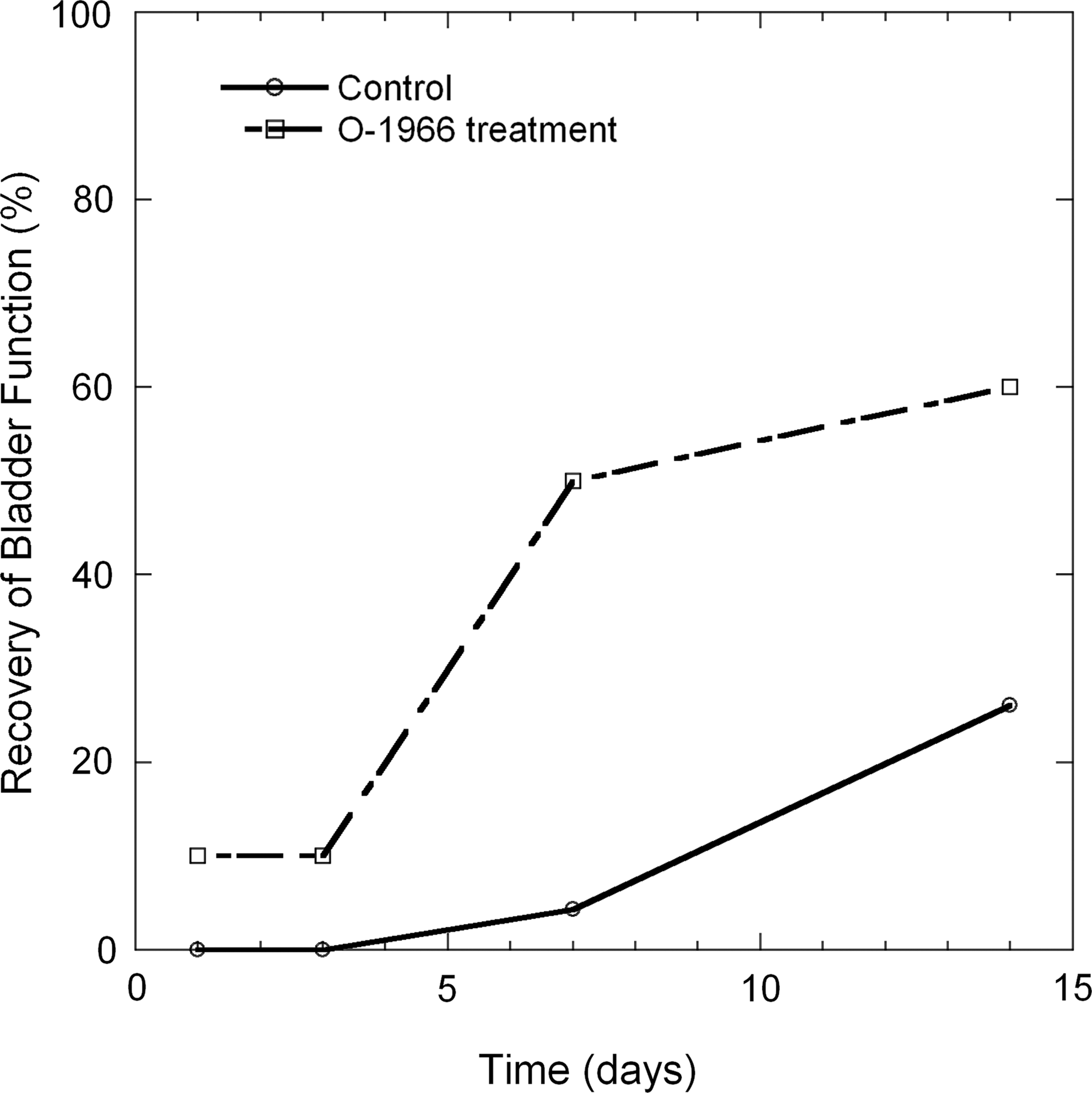
The percentage of animals in the vehicle-treated and O-1966-treated groups that were able to spontaneously empty their bladder following injury as a function of time (n=14 in the vehicle-treated and n=10 in the O-1966-treated groups).
Inflammatory cell invasion
To assess the invasion of the spinal cord by blood-derived leukocytes we used the levels of CD45 to discriminate between blood-derived infiltrates (CD45high) and resident microglial cells (CD45low), as previously described (Sedgwick et al., 1991). To further characterize the details of the observed populations we used several surface markers: hematopoietic cells (CD45+), myeloid cells (CD45hiCD11bhi), monocytes (CD45hiCD11bhiGR-1lo), neutrophils (CD45hiCD11bhiGR-1hi), microglia (CD45loCD11bhi), and T cells (CD45hiCD11b-CD3+). The usefulness of Gr-1 for distinguishing between neutrophils and monocytes has previously been shown by Stirling and Yong (2008).
Changes in white blood cell invasion resulting from drug treatment did not reach statistical significance at the 12-h and 48-h sampling time points. However, at 7 days post-injury in the spinal cord contusion site, the dominant treatment-related changes were decreases in hematopoietic (p<0.05; Fig. 3) and myeloid cell invasion (p<0.01; Fig. 3). A trend toward a decrease was also observed for monocytes, neutrophils, and microglia (Fig. 3). T cells were also present in the spinal cord, but their overall numbers were too small for an accurate assessment. To determine the potential effects of O-1966 on immune cells, in addition to the injury site in the cord, we analyzed the spleen for total cells, monocytes (CD45hiCD11bhiGR-1lo), neutrophils (CD45hiCD11bhiGR-1hi), and T cells (CD45hiCD11b-CD3+). There was no significant difference in terms of numbers of monocytes, neutrophils, and T cells between vehicle-treated and O-1966-treated animals.
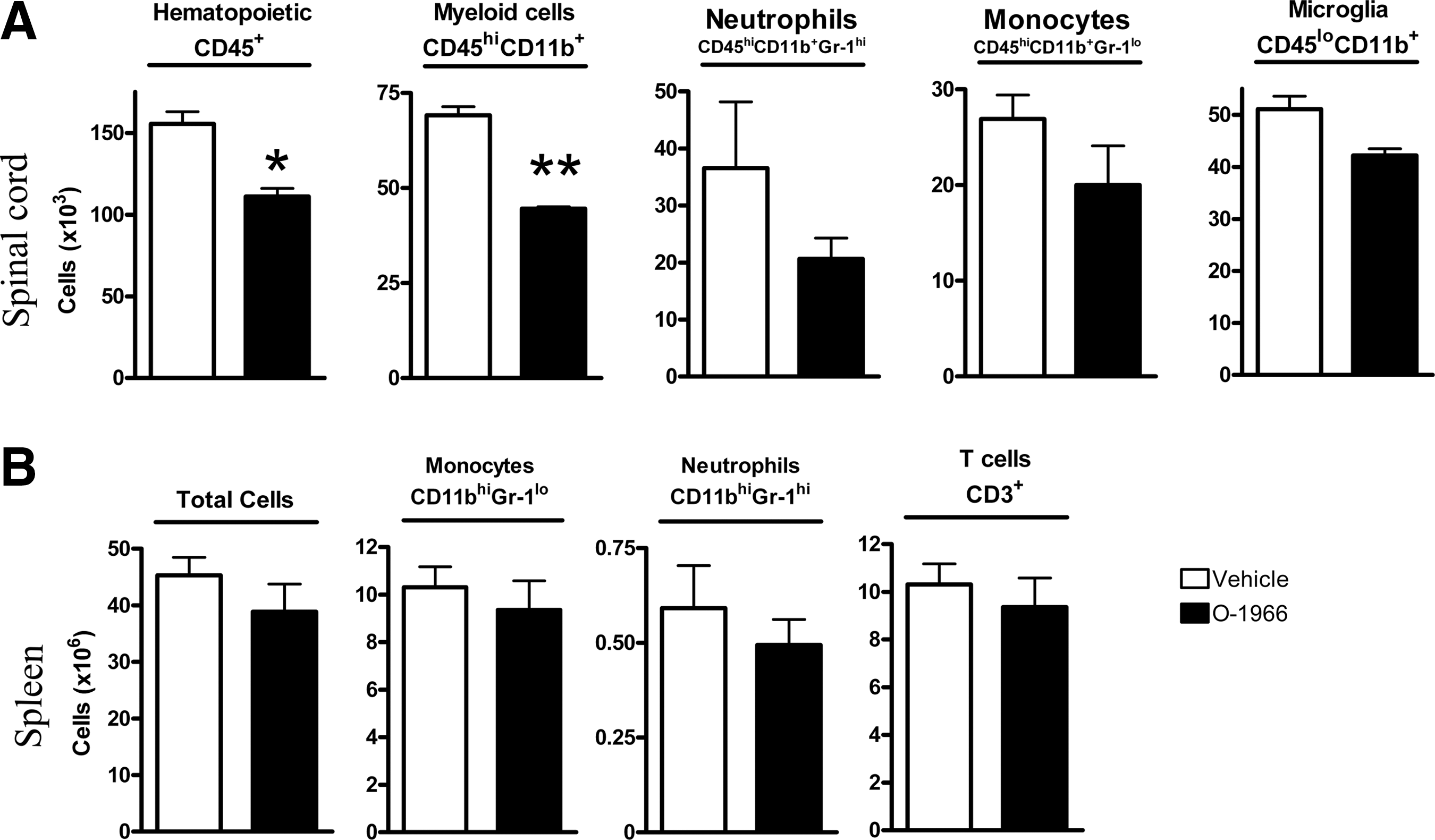
Cellular composition and cell numbers of the injured spinal cord area post-injury (7 days) and spleen (7 days). Open bars indicate vehicle-treated) animals, and solid bars indicate O-1966-treated animals. (
Chemokine/cytokine expression
At 6 h post-spinal cord injury various chemokines and cytokines were upregulated in injured cords compared to cords of sham-operated animals. PCR array analysis showed a significant increase in the expression of CCL-2, CCL-3, CCL-4, CXCL-10, CXCL-1, IL-6, IL-1α, and IL-10 in injured cords, as shown in Figure 4A. Animals treated with O-1966 1 h post-injury did not show any attenuation in the expression of these molecules. Instead, there was an increase in the expression of CCL-2, CXCL-1, and IL-10, which was confirmed by real-time PCR (Fig. 4B).
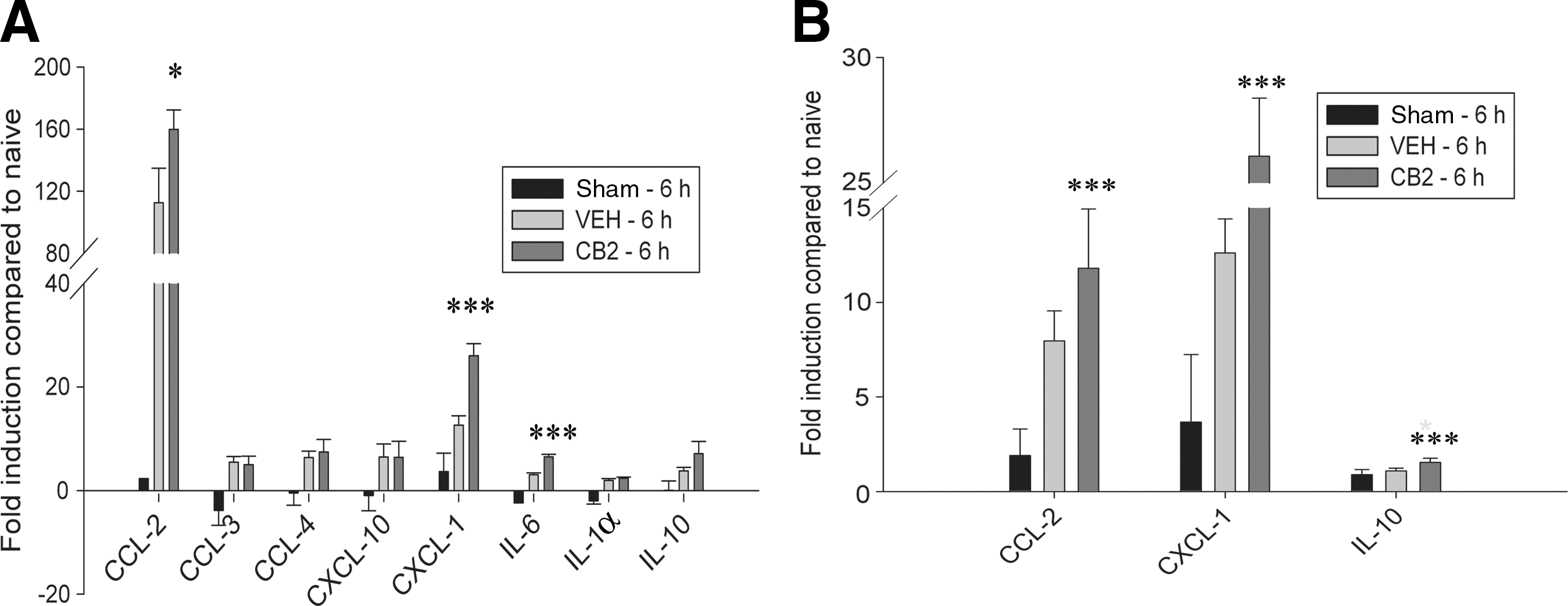
Analysis of mRNA expression of cytokines and chemokines via PCR-array (
At 48 h post-spinal cord injury, expression of many of the chemokines and cytokines seen at 6 h had returned to baseline levels, while there was an upregulation in the expression of a different set of molecules. PCR array analysis showed a significant increase in the expression of CXCL-9, CXCL-11, IL-23p19 subunit, IL-23 receptor, CCR-1, CCR-2, CCR-3, and CXCR-2 in the injured animals (Fig. 5A). The early chemokines CCL-2, CCL-3, and CCL-4 remained elevated in comparison to cords of sham-operated animals. Animals treated with O-1966 1 h and 24 h post-injury showed significant decreases in the expression of CXCL-9, CXCL-11, IL-23p19, and IL-23 receptor compared to animals treated with vehicle. These findings were confirmed by real-time PCR, as shown in Figure 5B. There was also a trend toward a decrease in the expression of CCR-1, CCR-2, CCR-3, and CXCR-2 in the treated cords, though the data did not reach statistical significance. However, there was no difference in the expression of CCL-2, CCL-3, or CCL-4.
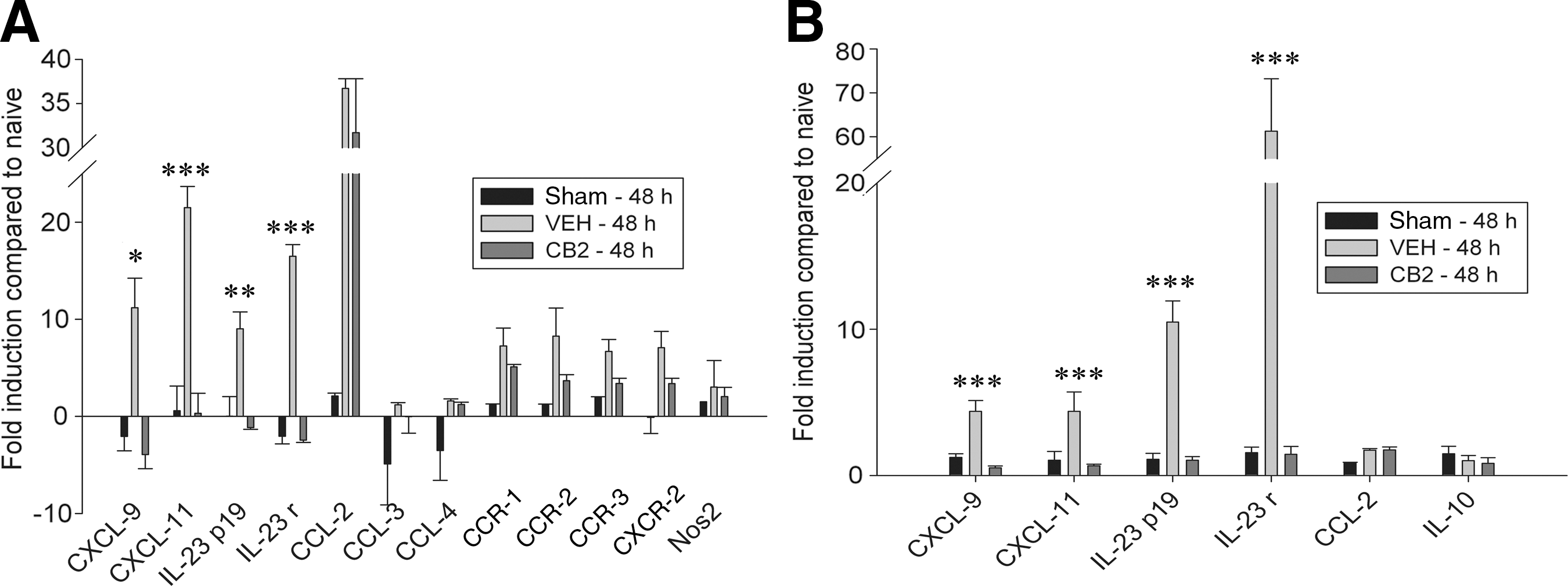
Analysis of mRNA expression of cytokines and chemokines via PCR-array (
Expression of two of the most common inflammatory cytokines, tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-1β), was also analyzed via real-time PCR at both 6 and 48 h post-injury (data not shown). At 6 h there was a small increase in expression of these cytokines in injured animals compared to sham animals. However, there was no difference between treated and untreated animals. These cytokines were not expressed at 48 h post-injury.
Spinal cord injury also led to upregulation of various toll-like receptors. At 6 h post-injury TLR1 and TLR2 were upregulated (Fig. 6A), while at 48 h post-injury TLR1, TLR2, TLR4, TLR6, and TLR7 were upregulated compared to sham-operated animals (Fig. 6B). Treatment with O-1966 had no effect on the expression of TLR2 at either time point. However, it significantly attenuated the expression of TLR1, TLR4, TLR6, and TLR7 at 48 h post-injury.

Analysis of mRNA expression of toll-like receptors (TLRs) via polymerase chain reaction (PCR) array (
Immunoreactive microglia
When evaluated 7 days post-injury, evaluation by the merging of the DAPI and Iba-1 images and counting Iba-1-positive cells showed that CB2-treated animals had significantly fewer immunostained microglia at a distance of 1.5 mm rostral and 1.5 mm distal from the epicenter of injury (Figs. 7 and 8), confirming the trend toward fewer reactive microglia measured in the flow cytometry studies.
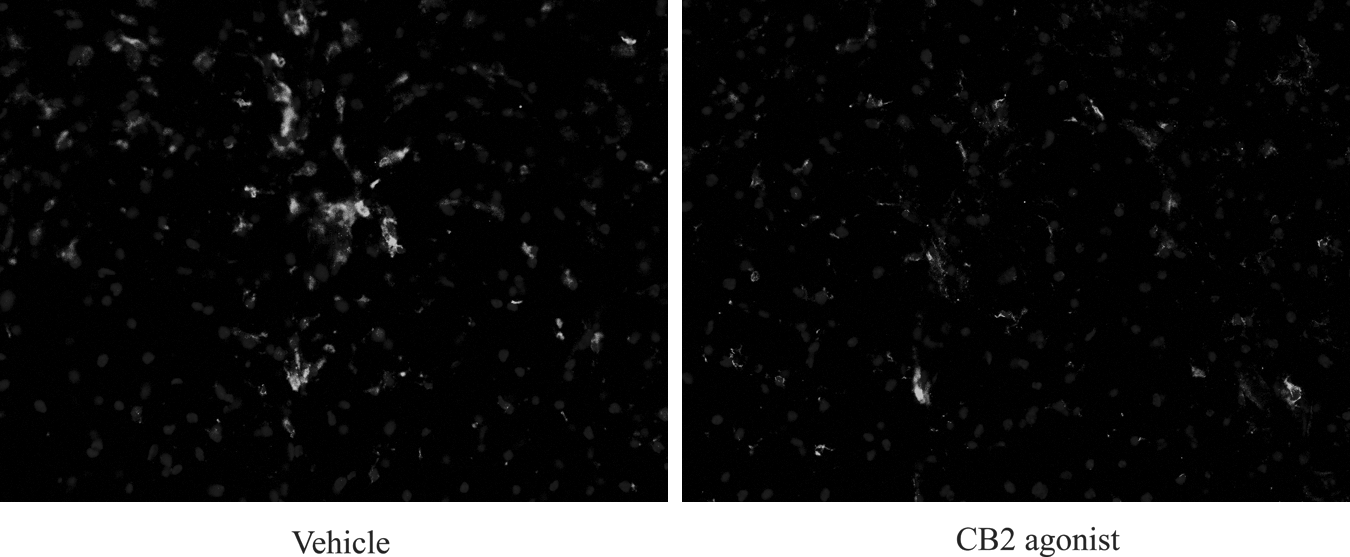
Immunostained microglia in a vehicle-treated and O-1966-treated (CB2) animal 1.5 mm from the epicenter of injury shown at 10×40 magnification (CB2, cannabinoid-2).

Number of Iba-1-positive cells in vehicle-treated and O-1966-treated (CB2) animals 1.5 mm rostral and 1.5 mm caudal from the epicenter of injury.(***p<0.005; CB2, cannabinoid-2).
Discussion
Physical damage to the spinal cord is accompanied by delayed secondary pathology, commonly termed “secondary injury.” Included among the components of secondary injury are neuronal and glial apoptosis, increased blood–spinal cord barrier (BSCB) permeability, and prolonged neuroinflammatory responses (Donnelly and Popovich, 2008; Gal et al., 2009; Gordon, 2003). Post-traumatic inflammatory reactions in the spinal cord are even more pronounced than that occurring following traumatic brain injury (Schnell et al., 1999). Since secondary injury is a major contributor to long-term damage and spinal cord dysfunction, modulation of secondary injury has been viewed as an important therapeutic target. Considerable efforts have been directed toward modulation of the inflammatory component of secondary injury in both experimental animals and humans. Although there is significant evidence that the inflammatory component of secondary injury enhances the magnitude of damage, and that attenuation of inflammatory responses can preserve neuronal function and enhance recovery, there is also evidence that components of the inflammatory response also play an important role in enhancing recovery (Alexander and Popovich, 2009; Elkabes et al., 1996; Perry et al., 1987; Yong and Marks, 2010). Inflammation therefore functions as a two-edged sword. As a result of its dual role in both exacerbating damage and promoting recovery, generalized broad-based suppression of inflammation is not an appropriate therapeutic strategy following spinal cord injury. Selective inhibition of the damaging components of inflammation, along with sparing or enhancing the components that promote recovery, represents the ideal strategy for treatment. Unfortunately, our understanding of the role played by the many components of the inflammatory response after injury is still poorly understood, and therapeutic agents that have this dual role have yet to be discovered.
One therapeutic strategy for better selective manipulation of the inflammatory response following spinal cord injury is modulation of the endocannabinoid system. We were the first to demonstrate in previous studies that acute activation of the CB2 receptor and/or inhibition of the CB1 receptor is associated with attenuation of damage and improved recovery following CNS injury (Baty et al., 2008; Zhang et al., 2007,2009b). In the current investigation we were able to show that administration of a selective CB2 agonist 1 h and 24 h following contusion of the spinal cord resulted in improved motor and bladder function over the 14-day period of observation. Most impressive was the return of spontaneous bladder emptying in the animals treated with the CB2 agonist. These results are very similar to those previously reported by our laboratory, when a lower dose of the CB2 agonist was administered prior to injury (Heller et al., 2009). Autonomic dysfunction causing a loss of bladder control following spinal cord injury is a major source of morbidity, and frequently leads to mortality, in spinal cord-injured patients. Whether the improvement in bladder function in the treated animals is the result of better preservation of the spinal cord, or the result of a direct effect on the bladder remains to be determined.
Although there are numerous reports of anti-inflammatory effects of CB2 agonists in a variety of experimental models of CNS injury, the precise mechanisms via which activation of the CB2 receptor enhances recovery following injury to the spinal cord remain to be determined (Alsasua del Valle, 2006; Berdyshev, 2000; Cabral et al., 2008; Dittel, 2008; Guindon and Hohmann, 2008, Heller et al., 2009; Pacher et al., 2006).
One of the mechanisms via which treatment with a selective CB2 agonist may contribute to improvement in spinal cord function is preservation of the BSCB. Increased permeability of the BSCB is thought to initiate a number of changes leading to detrimental inflammatory reactions in the spinal cord. Results from previous evaluations of the effects of a CB2 agonist on the blood–brain barrier following ischemia/reperfusion injury indicate that treatment contributes to preservation of the barrier, and protection of endothelial cells (Zhang et al., 2009b). The preservation of endothelium not only contributes to reductions in edema, but also induces a reduction in invasion of the spinal cord by circulating inflammatory cells. As it has previously been shown that hematopoietic cell invasion is an orchestrated process, we wanted to determine the therapeutic effect of O-1966 on the dynamics of hematopoietic cell invasion into the injured site of the spinal cord, and on local microglial activation (Stirling and Yong, 2008). Inhibition of CNS invasion by macrophages has been shown to reduce damage following spinal cord injury (Bao et al., 2004; Blight, 1994; Giulian and Robertson, 1990; Mabon et al., 2000; Popovich et al., 1999; Satake et al., 2000; Taoka et al.,1997). We have previously demonstrated, in investigations of cerebral ischemia/reperfusion, that administration of a selective CB2 agonist reduces adhesion molecule expression, and white cell adhesion and rolling on venules (Zhang et al., 2007,2009a). We therefore hypothesized that administration of a CB2 agonist following spinal cord injury might reduce white blood cell invasion of the spinal cord, and also influence microglial activation. Interestingly, there was not a significant effect on white cell invasion at 12 h after injury. There was also a trend toward an increase in the number of resident inflammatory cells (microglia) in the CB2-treated animals as determined by flow cytometry. However, at 48 h after injury there was a trend toward reduced white cell invasion and a reduction in the number of microglia. Seven days following injury there was a significant reduction in both hematopoietic and myeloid cells, and a trend toward a reduction in the number of microglia. The lack of a significant change in white cell numbers in the spleen indicates that this was not the result of a systemic decrease in the number of white blood cells. Treatment ending 24 h after injury had a prolonged effect on immune cell invasion.
Immunohistological staining was performed to further evaluate the effect of treatment with a CB2 agonist on microglial activation. The results of these studies demonstrated that at a distance of 1.5 mm from the epicenter of injury, the number of immunoreactive microglia was significantly reduced 7 days following injury. The results of this investigation demonstrate a prolonged targeted effect of a CB2 agonist on the inflammatory response that persists for at least a week following treatment.
Since increased production of proinflammatory cytokines and chemokines have been reported to be associated with enhanced injury, the effect of treatment with the CB2 agonist on cytokine/chemokine upregulation was also investigated through real-time PCR. At 6 h post-spinal cord injury a number of chemokines and cytokines were upregulated in the injured cords compared to cords from sham-operated animals. As was the case with white blood cell invasion, the effects of treatment were more striking 2 days after treatment than at the earlier time point (6 h). When evaluated 6 h after injury, the animals treated with the CB2 agonist had a significant increase in CXCL-1, CCL-2, and IL-10 mRNA expression compared to untreated animals. CXCL-1 and CCL-2 are both proinflammatory and are thought to be chemoattractants for neutrophils and monocytes, respectively, while IL-10 is thought to have anti-inflammatory properties. The expression pattern was significantly altered by 48 h post-injury. In this case CCL-2 was no longer elevated in the treated compared to the untreated animals, and there was a significantly lower expression of CXCL-9 and CXCL-11, both of which function as T-cell chemoattractants. CB2 treatment also resulted in a dramatic reduction in IL-23p19 expression and its receptor IL-23r. These results indicate a potential for CB2 treatment to influence T-cell differentiation, with a reduction in the number of invading IL-17-producing T cells. IL-17 T cells have recently been recognized as important contributors to the exacerbation of spinal cord injury.
The most commonly expressed cytokines TNF-α and IL-1β were moderately increased at 6 h post-injury in injured animals compared to sham controls. However, there was no difference between treated and untreated animals. In our previous study, in which O-1966 was administered 1 h prior to spinal cord injury, we saw a reduction of TNF-α expression in animals treated with the agonist. In this study we treated animals at 1 h post-injury. Both TNF-α and IL-1β are already expressed as early as 1 h post-injury in murine models of SCI (Pineau and Lecroix, 2007). This may explain why we did not see a reduction of these cytokines in animals treated with O-1966 at 1 h post-SCI.
The most striking and surprising result was the effect of CB2 treatment on the expression of toll-like receptors. Toll-like receptors, originally identified as reacting to pathogen-associated molecular patterns from invading organisms, have recently been shown to also be activated by endogenous molecules called danger-associated molecular patterns (DAMPs; Asea et al., 2002; Li et al., 2001; Ohashi et al., 2000; Okamura et al., 2001; Termeer et al., 2002). A number of DAMPs, including heat-shock proteins, necrotic cells, degeneration products of extracellular matrix molecules, mRNA, and high-mobility group box 1, are formed following injury to the spinal cord, and can activate TLR2 and TLR4 (Kigerl et al., 2007). In the injured CNS, the majority of toll-like receptors are located on monocytes and microglia (Laflamme and Rivest, 2001; Zekki et al., 2002). TLR2 appears to be expressed on both astrocytes and microglia/macrophages, while TLR4 is primarily restricted to microglia and macrophages (Kigerl et al., 2007). Activation of toll-like receptors results in nuclear factor-κB (NF-κB) signaling and the production of proinflammatory cytokines (Medzhitov, 2001; Nguyen et al., 2002; Tanga et al., 2005; Zhang et al., 1999). As reported previously (Kigerl et al., 2007), we found a significant upregulation of toll-like receptor expression following injury to the spinal cord. Although there was no significant effect of treatment on toll-like receptor expression at 6 h after injury, CB2 activation showed a remarkable effect at 48 h. Animals treated with the selective CB2 agonist had significant reductions in TLR1, TLR4, TLR6, and TLR7 compared to vehicle-treated animals. The analysis in these studies was based on mRNA expression, not protein expression. To our knowledge, this is the first report of the effect of a CB2 agonist on toll-like receptor expression. These effects on toll-like receptor expression have the potential to have profound influence on the inflammatory cascade and subsequent recovery following spinal cord injury. The precise mechanisms via which these changes influence outcome following spinal cord injury are not clear. Although TLR2 and TLR4 activation have been shown to enhance the production of proinflammatory cytokines, including IL-1β and TNF-α, animals deficient in TLR2 and TLR4 have enhanced injury and poorer recovery following spinal cord injury (Kigerl et al., 2007). Unlike the TLR4-deficient animals, which had an increase in macrophage invasion, the CB2-treated animals had a significant reduction in macrophage activation. These results may again highlight the principle that selective inhibition of inflammation may be beneficial, while more global inhibition can interfere with healing. While CB2 activation attenuated injury-induced increases in TLR4-receptor expression, it did not completely inhibit expression of this receptor. In short, CB2-receptor activation has the potential to benefit recovery in SCI by modulating the immune response.
Footnotes
Acknowledgments
This work was supported by grant no. 82412 from the Craig H. Neilsen Foundation, training grant T32 DA07237 (to S.A.), grant DA P30 13429 from the National Institute on Drug Abuse, and grant AI084065 from the National Institute of Allergy and Infectious Diseases.
Author Disclosure Statement
No competing financial interests exist.