
Abstract
Infectious complications are responsible for 10–25% of mortality in head-injured patients. In the present work we developed a model of infectious complications in head-injury rats using Escherichia coli (E. coli) with a stable copy of the lux operon, and monitored the infection in vivo by optical imaging. Rats were randomized into three groups: AL (healthy rats), HI (head-injury rats), and HI-EC (HI rats+single enteral bolus of E. coli, 1.3×109/rat given 2 days after HI). Infection was evaluated with a camera at 2 and 6 h after E. coli challenge. Blood and organs were sampled to assess biological parameters. HI was associated with body weight loss, muscle atrophy, and plasma amino acid disturbances, in particular glutamine depletion (AL 919±37 versus HI 647±25 and HI-EC 717±20 μmol/L; p<0.05). In the HI-EC rats, the luminescence signal was observed at T+2 (mean [range]: 34,778 cpm [1617–2,918,810]), and was significantly decreased at T+6 (0 cpm [0–847,922]; p<0.05). Bacterial challenge was associated with a specific body weight loss and a decrease in gastrocnemius protein content, in alanine (AL 512±41 versus HI-EC 395±29 μmol/L; p<0.05), and in sulfur plasma amino acids. In conclusion, we propose a controlled model of HI with infectious complications characterized by specific metabolic alterations. Combined with the in vivo monitoring of the infection by bioluminescence, this model offers a valuable tool to evaluate specific strategies for HI patients.
Introduction
T
The availability of animal models relevant to the clinical situation is essential to develop new strategies in the management of HI patients. To the best of our knowledge, experimental models found in the literature have described either trauma or sepsis, but not both together. Numerous models of head trauma are available in the literature (Morales et al., 2005). Among these, the model of HI induced by fluid percussion (Toulmond et al., 1993) has been well characterized in terms of impairment of cognitive functions (Piot-Grosjean et al., 2001) and metabolic disturbances following trauma (Moinard et al., 2005,2008). For sepsis, models involving the administration of live bacteria have already been developed (Breuillé et al., 1998; Dyson et al., 2009; Fink et al., 1990). However, most of these do not take into account the processes of translocation and dissemination, which are emerging as an important factor of sepsis in stressed patients (MacFie et al., 2006). Finally, a method that allows dynamic in vivo monitoring of the infection as it develops is still lacking in such experimental models.
Against this background, we set out to develop a new rat model combining head injury and infectious complications induced by enteral administration of the pathogenic luminescent strain of Escherichia coli Xen14, which allows in vivo monitoring of infection by optical imaging. This method, which visualizes only live bacteria, offers a good way to follow pathogenic agents in vivo (Jawhara et al., 2004; Kadurugamuwa et al., 2003; Xiong et al., 2005). The advantages of this non-invasive, rapid, real-time monitoring approach over the standard methods were recently reviewed by Contag (Contag, 2008). In our model, the quantization of luminescence images allows the extent and intensity of infection to be determined, and can also provide temporal and spatial information on bacterial dissemination.
Methods
This program was approved by the Comité Régional d'Éthique en Expérimentation Animale Paris Descartes (regional animal ethics committee), and was registered under the number P2.CM.018.06.
Animals and study design
Male Sprague-Dawley rats (300–325 g; Charles River, L'Arbresle, France) were housed individually in a controlled-temperature environment (21±1°C), with a 12-h light/dark cycle (8:00
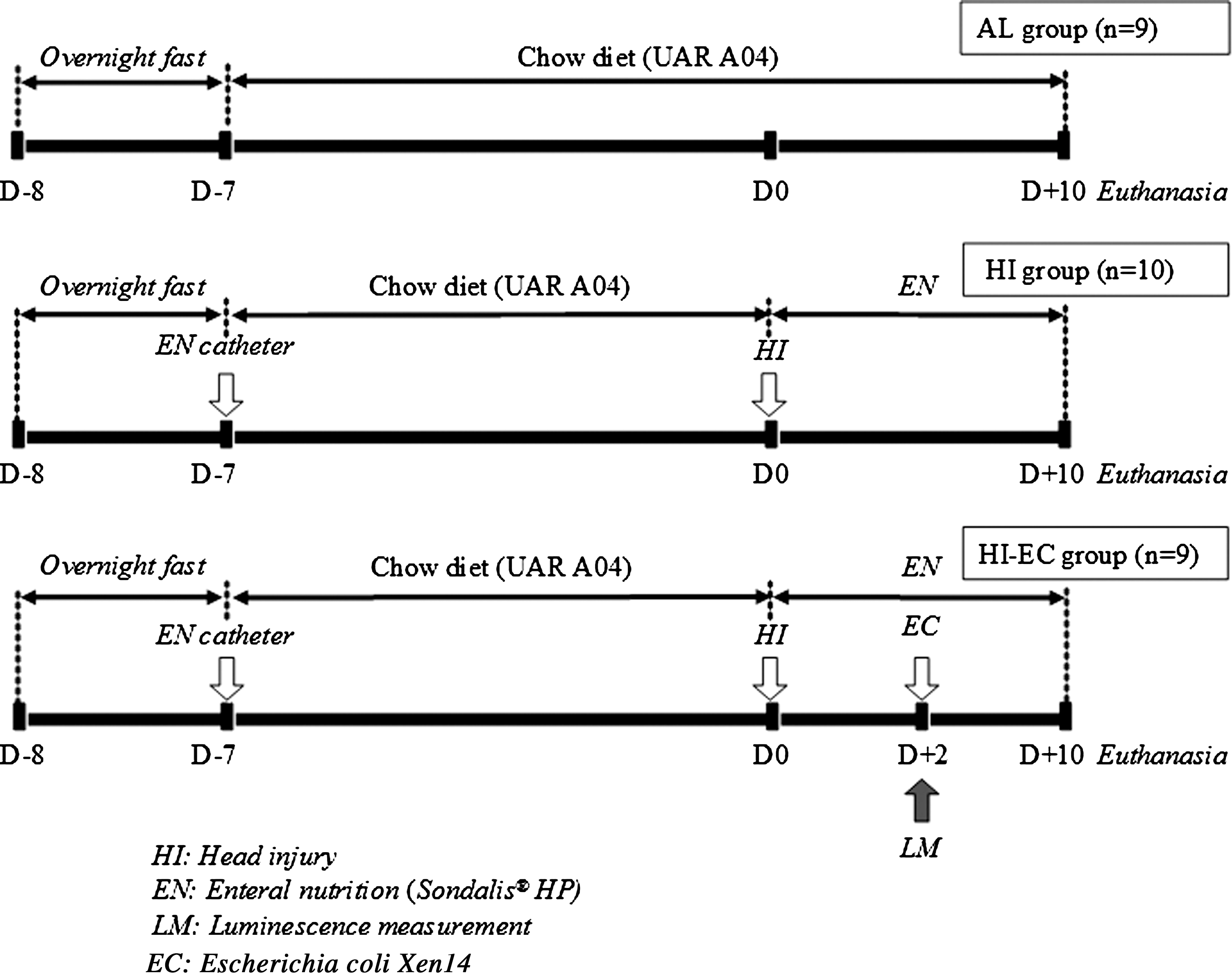
Experimental design.
Since recommendations for enteral nutrition only concern patients unable to maintain adequate oral intake, there is no clinical rationale for using enteral nutrition in healthy subjects. For this and ethical reasons, healthy rats fed ad libitum were chosen as a control group.
Surgical procedure for enteral nutrition
Seven days before HI (D–7), the rats underwent gastrostomy as recently described (Hamani et al., 2007). Briefly, under anesthesia with isoflurane (1.5 L O2/4% isoflurane; Minerve, Esternay, France), the abdominal and neck areas were shaved, and the skin was carefully cleaned with povidone iodine. The animals underwent a left-sided laparotomy, and the greater curvature of the stomach was isolated. A silicon catheter (Fisher Bioblock Scientific, Illkirch, France) was placed inside the stomach and secured by a pursestring suture. The catheter was tunnelled subcutaneously to the neck, and attached to a spring coil-swivel mechanism. A polyethylene tube (Fisher Bioblock Scientific) connected the cannula to a pump (model 1140-101; Harvard Apparatus, Les Ulis, France). Before the surgical procedure, all the rats received a painkiller (Temgesic®; Schering-Plough, Paris, France) subcutaneously at a dose of 50 μg/kg of body weight (BW). The rats were then fed ad libitum and allowed water ad libitum in metabolic cages for a 6-day recovery period.
Head trauma
Seven days after gastrostomy (D0), the rats were anesthetized with chloral hydrate (400 mg/kg IP; Prolabo, Fontenay-sous-Bois, France), and head trauma was induced by fluid percussion as previously described (Hamani et al., 2007).
Nutrition program
HI and HI-EC rats received a standard diet (Sondalis HP®; Nestlé Clinical Nutrition, Noisiel, France) infused continuously by the enteral route at a constant rate of 0.8 mL/100 g BW/h for 10 days, providing 290 kcal/kg/d and 3.29 g of nitrogen/kg/d (Hamani et al., 2007). BW and urine volume were recorded daily.
In vivo evaluation
E. coli challenge and evaluation of bioluminescence
E. coli-Xen14 (Xenogen Corp., Alameda, CA), derived from enteropathogenic E. coli (EPEC) strain WS2572, possesses a stable copy of a luminous bacterium (Photorhabdus luminescens) lux operon on the bacterial chromosome, together with a kanamycin resistance gene. The lux genes code for both the bacterial luciferase and substrate biosynthesis enzymes, which enable the strain to produce luciferase and luciferin simultaneously (Frackman et al., 1990). This results in a photon-emitting state when metabolically active, with no exogenous luciferin substrate requirement.
Challenge was performed with bacteria grown on trypcase soy agar and brain heart infusion broth (Oxoid, Dardilly, France) to their log phase as follows: 2 days after head injury (D+2), the rats in the HI-EC group received a single enteral bolus of E. coli-Xen14 corresponding to 1.3×109 bacteria per rat. The dose was chosen according to preliminary in vivo studies, which evaluated luminescence after enteral administration of increasing doses of E. coli-Xen14 in HI rats (1.3×107 to 1.3×109 bacteria). No luminescence was detectable below 109 bacteria (data not shown). In addition, this dose is in the range of the doses used in classical models of sepsis (Breuillé et al., 1999; Dyson et al., 2009; Heard et al., 1998).
The light emission of the bioluminescent bacteria was detected with a cooled intensified charge-coupled device camera (Biospace; Photon Imager, Paris, France) placed in a black box. This camera allowed real-time measurements (25 acquisitions per second). In vivo bacterial proliferation was evaluated by bioluminescence analysis at 0, 2, 6, and 48 h after bacterial challenge. The operating camera temperature was set at −16°C. Results of luminescence accumulated for 10 min were expressed as the median value [min – max] in cpm in the region of interest, as previously described (Bureau et al., 2010).
Neurological function
A neurological examination was performed in a blinded fashion using a grading scale at D+10. Contralateral sensorimotor functions were examined by assessing placing reactions (leg hanging and visual), grasping reflex, and righting reflex (“head tilted”) in rats placed on a table. The rats were also examined for abnormal postures (forelimb flexion and thorax twisting). The scores for each item were summed and used as a global neurological score (normal score: 1, and deficit: 0), the maximum being 9 for non-operated rats (Besson et al., 2003).
Ten days after HI (D+10) nutrition was stopped and 2 h later all the rats were anesthetized with isoflurane and decapitated.
Biological analysis
Blood collection and tissue removal
Blood was collected in sodium heparinate. Samples of the mesenteric lymph nodes, liver, spleen, lung, kidney, and cecal content were removed for microbiological analysis as previously described (Hamani et al., 2007). Bacterial counts were performed on Drigalski agar for enterobacteria, and on Drigalski supplemented with 30 μg/mL kanamycin for E. coli-Xen14. Several organs (jejunum, ileum, liver, extensor digitorum longus [EDL], tibialis, soleus, gastrocnemius, spleen, and thymus) were removed and weighed as previously described (Charrueau et al., 2009).
Biological parameters
Plasma concentrations of albumin and fibrinogen were determined by immunonephelometry using the Array System 360 analyzer (Beckman-Coulter Instruments, Roissy, France).
For plasma amino acid concentration determination, blood samples were rapidly centrifuged, and plasma was deproteinized with a 30% sulfosalicylic acid solution. The supernatants were stored at −80°C until analysis.
For tissue amino acid concentrations, tissues were ground and deproteinized with 10% trichloroacetic acid containing 0.5 mM ethylenediaminetetraacetic acid (EDTA). The supernatants were stored at −80°C until analysis.
Amino acids were separated and quantified by ion exchange chromatography using the Aminotac autoanalyzer (Jeol, Tokyo, Japan).
For protein determination, the frozen tissue was pulverized and homogenized in ice-cold 10% TCA (1 mL TCA/100 mg tissue) using an Ultra-Turrax T25 tissue disrupter (Ika Labortechnik, Staufen, Germany). After delipidation with ethanol-ether (1:1 vol/vol), the precipitate was dissolved in 1 N NaOH (4 mL/100 mg tissue) for 12 h at 40°C. The total protein content was then determined according to the method described by Fleury and Aberham (1951).
Statistical analysis
Data are expressed as means±SD. Statistical analysis was performed using Statview software (Deltasoft, Grenoble, France). The neurological score was analyzed using a Kruskal-Wallis test and the Mann-Whitney U test. The percentages of enterobacteria that translocated and disseminated, together with the intensity of enterobacterial translocation and dissemination, were analyzed by a Mann-Whitney U test. The luminescence values were analyzed using analysis of variance (ANOVA) and the Kruskal-Wallis test. All the other data were analyzed by ANOVA and the Fisher test. Values of p<0.05 were considered significant.
Results
Behavior of the rats: Mortality and complications
There was no mortality in the HI group. However, mortality following E. coli-Xen14 challenge occurred in 44% of the rats. Thus of a total of 35 rats, 28 were studied: 10 in the HI group, 9 in the HI-EC group, and 9 in the AL group.
Neurological function and biological parameters
Neurological function
Neurological score (over 9) significantly deteriorated following HI (HI: 5.6±0.6 and HI-EC: 5.7±0.8 versus AL: 9±0, p<0.05), with no difference between the HI and HI-EC groups.
Body weight
HI induced a significant decrease in BW (HI versus AL, p<0.05), and bacterial challenge was associated with a further BW loss (HI-EC versus AL and HI, p<0.05) (Fig. 2 and Table 1). After 10 days the injured rats totally recovered.
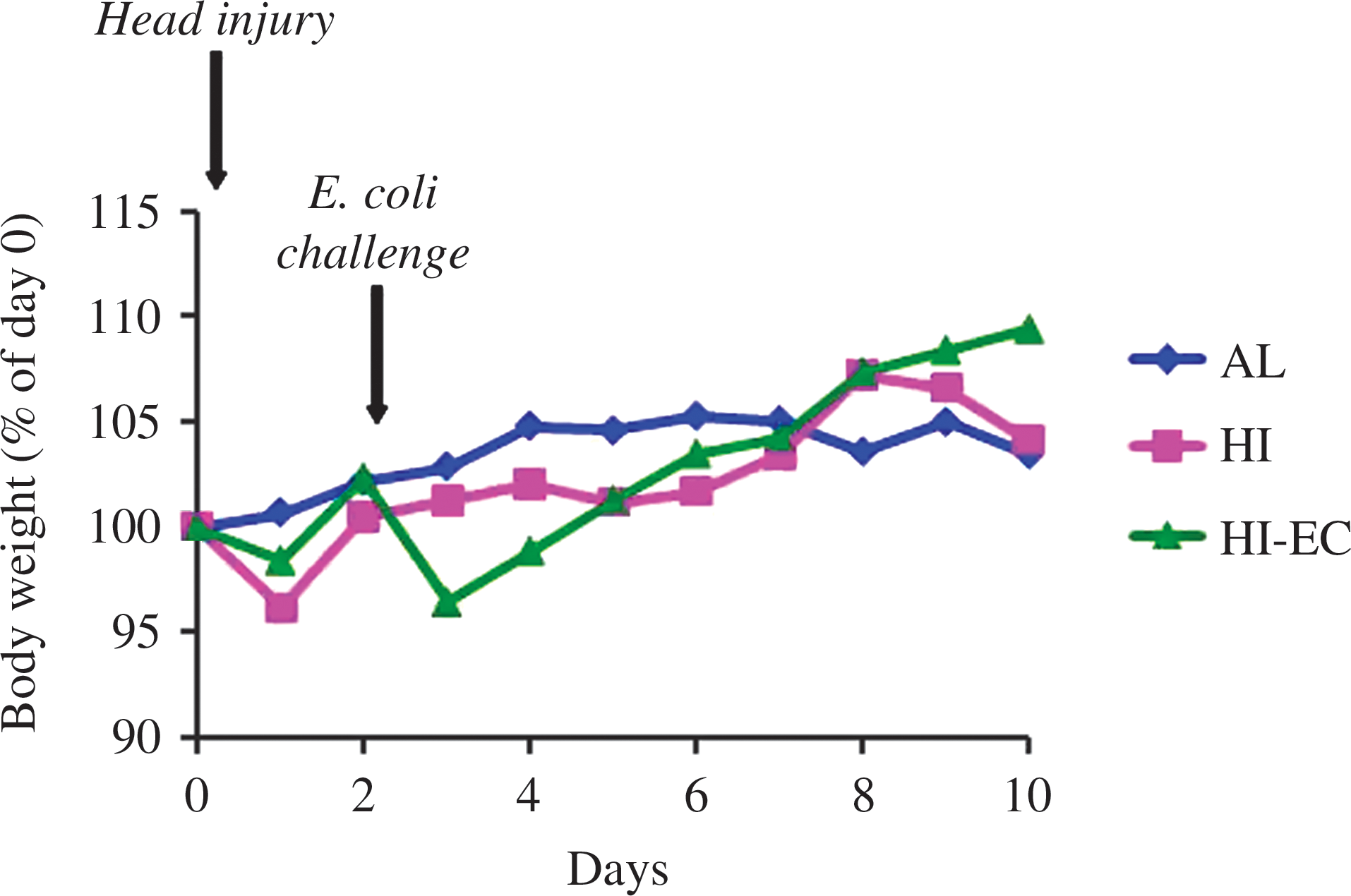
Body weight evolution between day 0 and day 10 in healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats. Results are given as means and are expressed in percent of body weight at day 0.
p<0.05 vs. AL; # p<0.05 versus HI (ANOVA and Fisher test).
Variation of total body weight in healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats.
Results are given as mean±standard deviation, and are expressed in grams.
Organ weight
No modifications in organ weight were observed in any of the groups, except for tibialis and gastrocnemius atrophy in HI rats (HI versus AL, p<0.05), and for spleen hypertrophy in the HI and HI-EC groups (HI and HI-EC versus AL, p<0.05; Table 2).
p<0.05 versus AL (analysis of variance and Fisher test).
Weight of jejunum, ileum, liver, extensor digitorum longus (EDL), tibialis, soleus, gastrocnemius, spleen, and thymus, in healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats.
Results are given as mean±standard deviation and are expressed in grams.
Tissue protein content
HI induced a significant decrease in protein content in the ileum, tibialis, and gastrocnemius (HI and HI-EC versus AL, p<0.05), and bacterial challenge worsened protein wasting in the gastrocnemius (HI-EC versus HI, p<0.05; Table 3).
p<0.05 versus AL; # p<0.05 versus HI (analysis of variance and Fisher test).
Protein content in jejunum, ileum, liver, extensor digitorum longus (EDL), tibialis, soleus, and gastrocnemius, in healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats.
Results are given as mean±standard deviation, and are expressed as mg/cm (jejunum and ileum), or mg/organ (EDL, tibialis, soleus, and gastrocnemius) or g/organ (liver).
Plasma albumin and fibrinogen
HI induced a significant decrease in plasma albumin concentration (HI versus AL, p<0.05), whereas the decrease did not reach significance in the HI-EC group (HI-EC versus AL, p=0.06; Table 4). A significant increase in plasma fibrinogen concentration was observed after HI (HI and HI-EC versus AL, p<0.05). However, the increase was less marked after bacterial challenge (HI-EC versus HI, p<0.05).
p<0.05 versus AL; °p=0.06 versus AL; # p<0.05 versus HI (analysis of variance and Fisher test).
Albumin and fibrinogen concentrations in plasma of healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats. Results are given as mean±standard deviation, and are expressed in g/L.
Amino acid concentration in plasma and tissues
HI induced a significant decrease in plasma glutamine, glycine, taurine, and histidine concentrations, and a significant increase in plasma tyrosine concentration (HI and HI-EC versus AL, p<0.05; Table 5). E. coli challenge was associated with a specific decrease in plasma alanine and sulfur amino acids (HI-EC versus AL, p<0.05).
p<0.05 versus AL; # p<0.05 versus HI (analysis of variance and Fisher test).
Plasma and tissue concentrations of selected amino acids (no significant variations for amino acids are not shown), in healthy (AL), head-injury (HI), and head-injury+infectious complications (HI-EC) rats. Results are given as mean±standard deviation, and expressed in μmol/L (plasma) or μmol/g (tissues).
EDL, extensor digitorum longus.
HI induced a significant increase in tissue concentrations of threonine (jejunum, tibialis, soleus, and gastrocnemius, HI and HI-EC versus AL, p<0.05; EDL, HI versus AL, p<0.05), tyrosine, arginine, and lysine (tibialis and gastrocnemius, HI and HI-EC versus AL, p<0.05; EDL, HI versus AL, p<0.05), and citrulline (jejunum, HI and HI-EC versus AL, p<0.05). HI induced a significant decrease in the gastrocnemius concentrations of branched-chain amino acids (isoleucine and leucine, HI versus AL, p<0.05). E. coli challenge was associated with specific alterations of glycine concentrations, which were increased in the jejunum (HI-EC and HI-EC versus AL, p<0.05), and decreased in tibialis and gastrocnemius (HI-EC versus AL, p<0.05), and glutamine concentrations, which were increased in gastrocnemius (HI-EC versus AL, p<0.05). Branched-chain amino acids significantly increased in gastrocnemius following bacterial challenge (valine, isoleucine, and leucine, HI-EC versus HI, p<0.05).
Microbiological analysis
Bioluminescence after E. coli-Xen14 challenge
In a preliminary experiment, we confirmed the quantitative aspect of the measurement of luminescence by in vitro study. We observed that the luminescence was linearly proportional to the bacterial concentration (r2=0.92; Table 6).
A typical experiment showing the luminescence at the body level, and localized in the gastrointestinal tract, after E. coli administration is depicted in Figure 3. Luminescence was not detectable at T0, and after bacterial proliferation, the luminescence signal peaked at T+2 h, and was significantly decreased at T+6 h, with a median value of zero due to the disappearance of luminescence in 5 out of 9 animals (Fig. 4). No signal was detectable 48 h after bacterial challenge (data not shown).
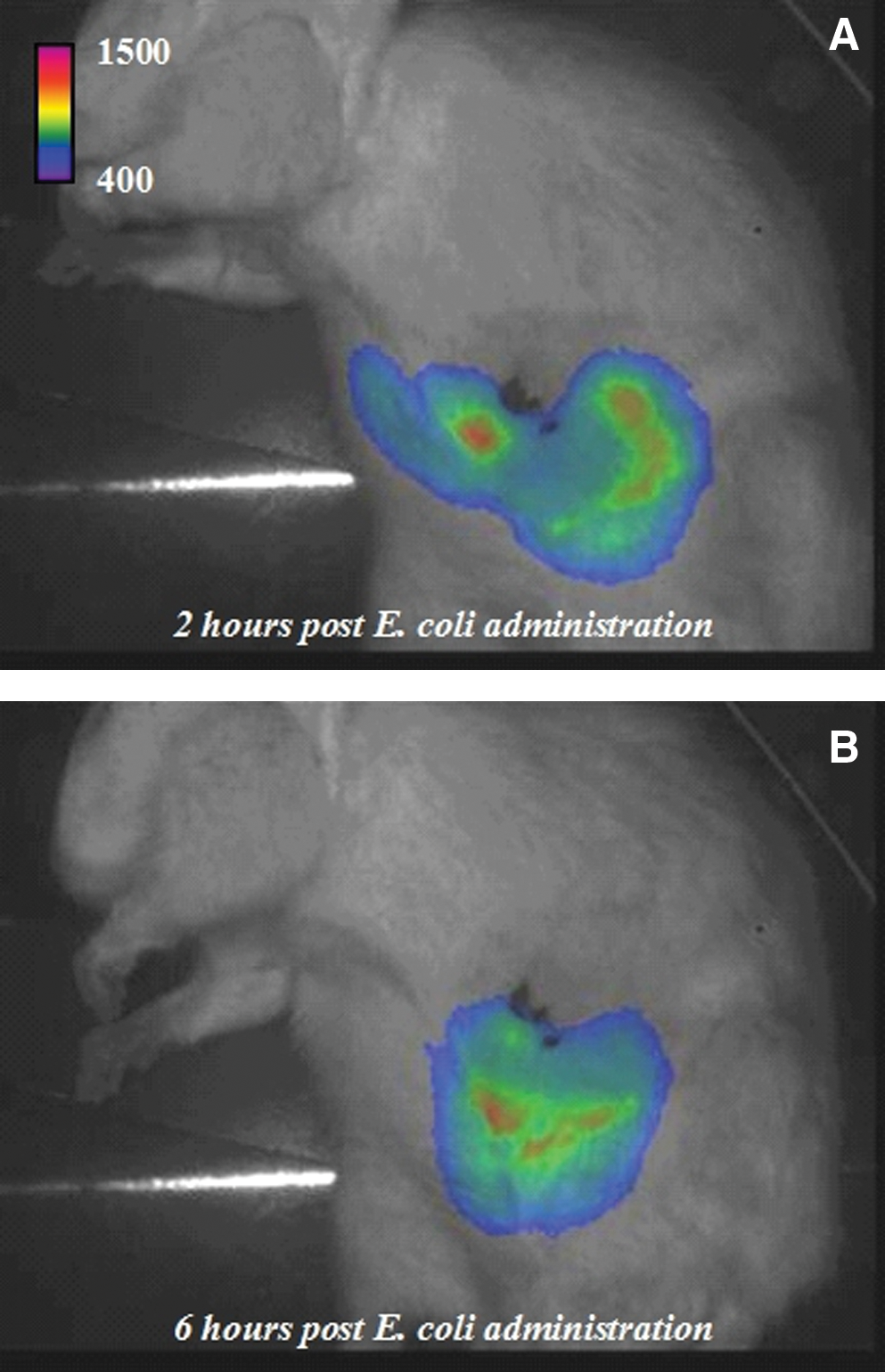
Levels of luminescence 2 h (
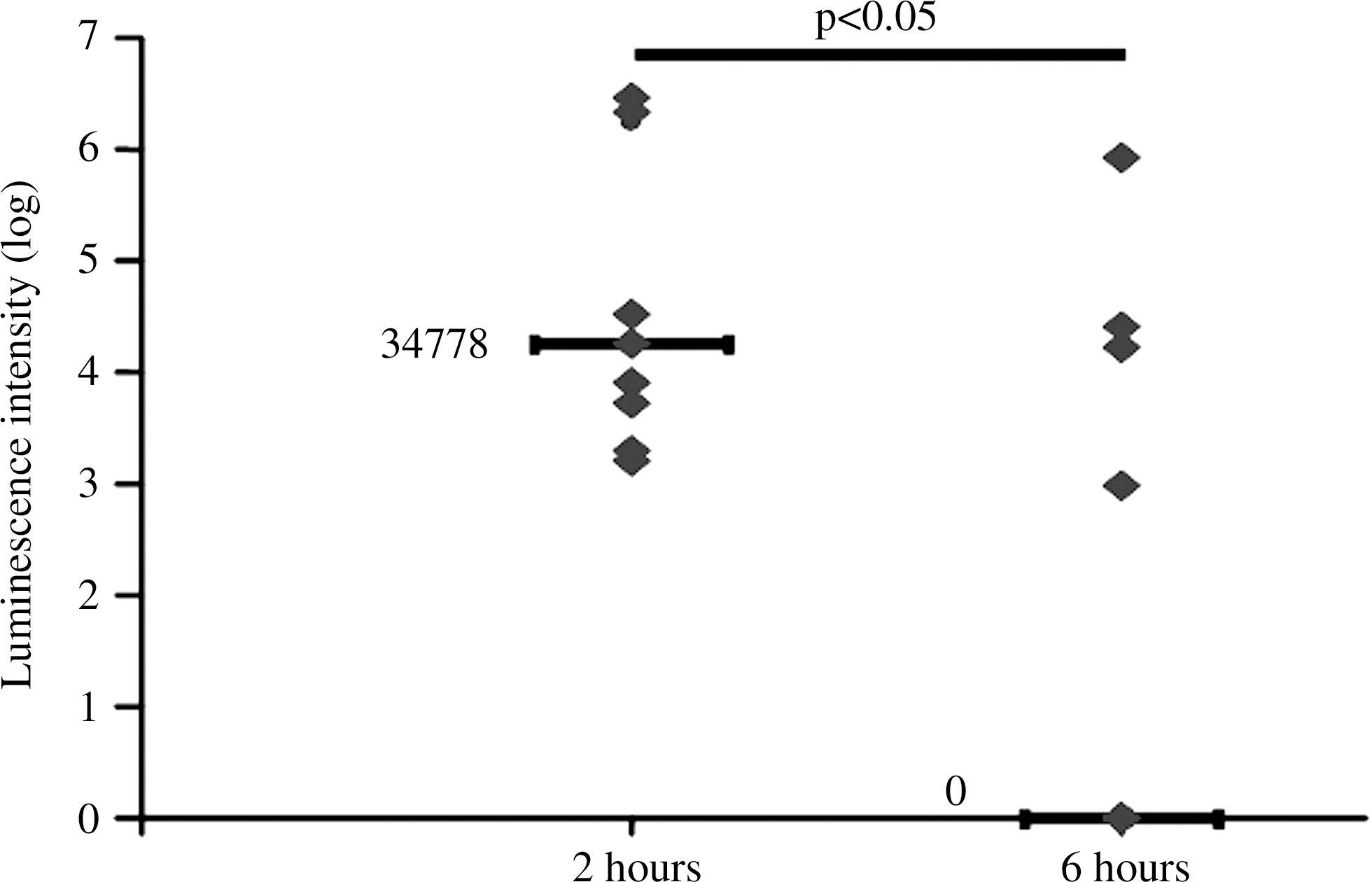
Bioluminescence intensity 2 h and 6 h after E. coli administration in head-injury+infectious complications (HI-EC) rats. Results are given as individual values of luminescence (♦) in cpm in regions of interest, and median value (—, log; p<0.05 by analysis of variance and Kruskal-Wallis test).
Enterobacteria translocation and dissemination
While neither translocation nor dissemination could be observed in the AL group (data not shown), and HI led to similar enterobacterial translocation in the mesenteric lymph nodes, and dissemination in the liver, spleen, and lung in both injured groups (HI versus HI-EC, NS). The bioluminescent E. coli-Xen14 was detected in the spleen, lung, and cecal contents (Table 7).
Level of translocation in mesenteric lymph nodes (MLNs), and level of dissemination in liver, spleen, lung, and kidney, and percentages of animals presenting translocation and dissemination, in head-injury (HI) and head-injury+infectious complications (HI-EC) rats. Level of enterobacteria in cecal content. Levels are expressed as the median [minimum–maximum] in colony-forming units/g.
Enterobacteria, commensal enterobacteria; E. coli-Xen14, bioluminescent E. coli administered at day 2.
Data analyzed by Mann-Whitney U test.
Discussion
Our study validates a new model combining head trauma and infectious complications. In addition, we used an original approach that allows a non-invasive, dynamic evaluation of septic complications.
Classical experimental models of sepsis (e.g., peritonitis models such as cecal ligation and puncture, or administration of endotoxin or live bacteria) reproduce the hormonal and metabolic disturbances observed in septic patients. However, these models, though useful, have several limitations: (1) standardization difficulties, (2) rapid recovery from the catabolic state (usually within 48 h), (3), extremely wide-ranging mortality (from near zero for endotoxemia to almost 100% for peritonitis models), and (4) no clinical relevance in terms of bacterial translocation and dissemination, since these models are applied in otherwise healthy animals.
As classically observed in this head trauma model, HI rats exhibited a BW loss, confirming a catabolic state (Moinard et al., 2005). As expected, E. coli challenge worsened BW loss. Of note, this is not the case when bacteria are administered to healthy rats (C. Moinard et al., unpublished observation). Interestingly, HI induces a high sensitivity to septic shock, with an exacerbation of the metabolic alterations concurrent with head trauma, an aggravation of the wasting syndrome, and an increased mortality (similar to clinical observations in HI patients; Quattrocchi et al., 1990). This result is confirmed by a decrease in muscle protein content in gastrocnemius (a mixed muscle that is very sensitive to muscle wasting; Hasselgren et al., 1989). This shows the relevance of our model to clinical practice, since 50–75% of patients sustaining severe HI develop infectious complications after initial hospitalization.
HI causes malnutrition, and infection promotes wasting. These alterations are thought to be induced by the acute-phase response, with synthesis of proinflammatory cytokines, and then synthesis of inflammatory proteins such as fibrinogen, along with gluconeogenesis and ureagenesis. This leads to muscle protein mobilization and induces a shift of amino acids from the periphery to the central organs, which explains the amino acid disturbances observed (see Charrueau et al., 2010 for a recent review). Interestingly, we observed that infection was associated with specific modifications such as decreased levels of sulfur amino acids (methionine and cysteine). These results agree with those of previous studies performed in septic rats, which clearly demonstrated that cysteine requirement was increased during sepsis (Malmezat et al., 2000). This may be related to an increase in glutathione synthesis (a major factor in oxidative defense during infection; Charrueau et al., 2010). Fibrinogen was lower and albumin was higher in the HI-EC group (vs HI), but we have no ready explanation for this unexpected finding.
In this work, optical imaging was implemented for the monitoring and quantification of bacterial translocation from the gut of live enteropathogenic E. coli-Xen 14, which produces luminescence when metabolically active. In a preliminary study performed in vitro, we observed, as was previously shown (Frackman et al., 1990), a close correlation between the luminescence signal and the number of bacteria, validating the quantitative approach of this method. E. coli-Xen14, which produces luminescence in its growing phase, was visualized 2 h and 6 h after challenge, but was not detectable after 48 h. No signal means that (1) bacteria were no longer in growth phase, or (2) the number of growing bacteria was below the luminescence detection level (Wu et al., 2001). However, they were still detectable by classical culture methods in the spleen, lung, and cecal contents 8 days after bacterial challenge. This bioluminescent imaging enabled us to visualize the spatiotemporal distribution of the bacterial load in this head-injured model. Bacterial translocation is an important early step in the pathogenesis of septic episodes originating from the gastrointestinal tract in head-injured patients. The spatial and temporal differences in the bioluminescence signal can be used to assess the effects of nutrition or other interventions on the biological process, as shown or suggested in other models (Jawhara et al., 2004; Kadurugamuwa et al., 2003; Sharma et al., 2010). Also, since the imaging procedure is non-destructive and non-invasive, experiments can be performed repeatedly in the same animal.
In conclusion, our model successfully reproduced the clinically-relevant infectious complications observed in HI. This model is unique, since septic complications can be evaluated directly in vivo, and are the direct reflection of evolving septic episodes. Also, we clearly observed that the combination of HI and bacterial challenge was characterized by specific metabolic alterations. This model may thus be useful for evaluating the effectiveness of strategies aimed at improving the clinical outcomes of HI patients, who usually develop septic complications.
Footnotes
Acknowledgments
The authors thank Djamel Hamani, who participated in early discussion of the project, and Vincent Pugeault, for participation in animal experiments.
This work was supported in part by an unrestricted grant from Nestlé Clinical Nutrition (Noisiel, France), and by a grant from the Ministry of Research and Technology.
Author Disclosure Statement
No competing financial interests exist.