
Abstract
The mammalian target of rapamycin (mTOR) is a serine/threonine kinase that negatively regulates autophagy. Rapamycin, an inhibitor of mTOR signaling, can promote autophagy and exert neuroprotective effects in several diseases of the central nervous system (CNS). In the present study, we examined whether rapamycin treatment promotes autophagy and reduces neural tissue damage and locomotor impairment after spinal cord injury (SCI) in mice. Our results demonstrated that the administration of rapamycin significantly decreased the phosphorylation of the p70S6K protein and led to higher expression levels of LC3 and Beclin 1 in the injured spinal cord. In addition, neuronal loss and cell death in the injured spinal cord were significantly reduced in the rapamycin-treated mice compared to the vehicle-treated mice. Furthermore, the rapamycin-treated mice showed significantly higher locomotor function in Basso Mouse Scale (BMS) scores than did the vehicle-treated mice. These results indicate that rapamycin promoted autophagy by inhibiting the mTOR signaling pathway, and reduced neural tissue damage and locomotor impairment after SCI. The administration of rapamycin produced a neuroprotective function at the lesion site following SCI. Rapamycin treatment may represent a novel therapeutic strategy after SCI.
Introduction
Autophagy is an intracellular catabolic mechanism for the degradation of cytoplasmic constituents in the autophagosomal–lysosomal pathway (Mizushima, 2004). This mechanism plays an important role in homeostasis and it is defective in certain diseases (Hara et al., 2006; Liang et al., 1999; Rubinsztein et al., 2005; Shintani and Klionsky, 2004). Autophagy eliminates and recycles long-lived proteins and unwanted damaged organelles in a cell during development and under stress conditions (Levine and Klionsky, 2004; Shintani and Klionsky, 2004). It has been reported that autophagy is also important for normal cell growth, differentiation, and survival (Reggiori and Klionsky, 2002; Schmelzle and Hall, 2000).
Recent studies demonstrated that rapamycin treatment enhances autophagy and rescues neuronal cells in several neurodegenerative diseases such as Huntington's disease and Parkinson's disease (Malagelada et al., 2010; Ravikumar et al., 2004). In such neurodegenerative diseases, induction of autophagy by rapamycin enhances the clearance of a wide range of aggregation-prone proteins and reduces their toxicity (Berger et al., 2006; Webb et al., 2003). Previous studies demonstrated that the activity of autophagy was increased in damaged neural tissue after traumatic brain injury and cerebral ischemia (Bigford et al., 2009; Diskin et al., 2005; Rami et al., 2008). Administration of rapamycin enhances the autophagic activity and reduces neural tissue damage after traumatic brain injury and neonatal hypoxia–ischemia induced brain injury (Carloni et al., 2008; Erlich et al., 2007a). Several studies also showed that the enhancement of autophagy can reduce apoptosis in damaged neural tissue of the central nervous system (CNS) (Carloni et al., 2008; Pan et al., 2008). Therefore, the promotion of autophagy using rapamycin is considered to provide a neuroprotective function in the CNS.
We previously reported that autophagic activity was significantly increased in damaged neural tissue after spinal cord injury (SCI) (Kanno et al., 2009b, 2011). However, the actual function of autophagy in the damaged neural tissue after SCI is still unclear (Kanno et al., 2009c). Furthermore, whether rapamycin treatment can promote autophagy and induce a neuroprotective function in the damaged neural tissue after SCI remain to be elucidated.
The purpose of the present study was to examine whether rapamycin treatment inhibits the mTOR signaling pathway and promotes autophagy after SCI, using a spinal cord contusion model in mice. We also examined whether rapamycin treatment reduces neural tissue damage such as demyelination, neuronal loss, and cell death, and improves locomotor recovery following SCI.
We show here that rapamycin treatment significantly enhanced autophagic activity by inhibiting mTOR in the injured spinal cord. Furthermore, rapamycin significantly reduced neural tissue damage and improved locomotor function after SCI. Therefore, administration of rapamycin may represent a novel therapeutic strategy after SCI.
Methods
Animals
All experimental procedures were approved by the Institutional Animal Care and Use Committee of Tohoku University. All efforts were made to minimize the number of animals used and to decrease the suffering of the animals used in the study. A total of 58 adult female C57BL/6J mice (10–12 weeks old; Charles River, Japan Inc., Yokohama, Japan) were used in this study. Four or five animals were used for each experimental group at each time point. The mice were housed three or four per cage and kept at a temperature of 24°C with free access to water and food before and after surgery.
Spinal cord contusion injury
The mice were anesthetized with 1.25% halothane in an oxygen/nitrous oxide (30/70%) gas mixture. During surgery, the rectal temperature was monitored and maintained at 37.0±0.5°C by a heating pad. The skin above the vertebral column was shaved and cleaned with antiseptic. A 15-mm mid-line skin incision was made, and the laminae of the T8–T12 vertebrae were exposed. A laminectomy was performed at T10, exposing the dorsal cord surface with the dura intact. The vertebral column was stabilized with angled clamps attached to the T8 and T12 transverse processes. A SCI was induced by a modified New York University Impactor (Basso et al., 1996; Gruner, 1992; Hashimoto et al., 2007; Kanno et al., 2009a; Kim et al., 2001; Okada et al., 2004). A 10 g rod (tip diameter: 1.5 mm) was dropped from 3 mm onto the T10 segment. The impact rod was removed immediately after injury. The muscles and skin were closed in layers. Bladders were expressed twice a day until spontaneous voiding began. The sham-operated animals received the same surgical procedures, but no impact injury was sustained.
Rapamycin preparation and treatment
Rapamycin (Calbiochem) was dissolved in dimethyl sulfoxide (DMSO) (25 mg/mL) and further diluted in 0.5 mL aqueous solution containing 5% polyethylene glycol 400 and 5% Tween 80 immediately before injection. At 4 h after SCI, mice in the rapamycin treatment group were injected intraperitoneally with rapamycin at a dose of 1 mg/kg body weight. Control animals were injected with an equivalent volume of the vehicle.
Behavioral analysis
To evaluate the functional consequences of SCI, a locomotor rating test using the Basso Mouse Scale (BMS) was performed (Basso et al., 2006; Engesser-Cesar et al., 2005). This scale was developed specifically for mice because the characteristics of locomotor recovery are different in mice than in rats. We also analyzed the BMS subscore, because some animals can show improvements in specific aspects of locomotion that do not follow a typical pattern of recovery, and therefore are not reflected in any change in the overall score on the BMS (Basso et al., 2006). Before surgery, the mice were placed individually in a molded plastic open field for 4 min to assure that all subjects consistently obtained the maximum score. The mice were placed in the open field, and well-trained investigators scored them on the BMS in a blinded manner. The BMS scores were measured at 4 and 24 h, and at 3, 7, 14, 21, 28, 35, and 42 days after SCI.
Western blot analysis
The mice were killed at 24 h or 3 days after SCI, and their spinal cords were removed. The spinal cords were homogenized in lysis buffer containing 50 mM Tris HCl (pH 7.6), 20 mM MgCl2, 150 mM NaCl, 0.5% Triton-X, 5 units/mL aprotinin, 5 μg/mL leupeptin, 5 μg/mL pepstatin, 1 mM benzamidine, and 1 mM phenylmethylsulfonyl fluoride. The debris was removed by centrifugation, and the protein levels in the lysates were determined with the aid of the Bio-Rad protein assay (Bio-Rad Laboratories, GmbH, Munich, Germany). The proteins in the lysates were separated by SDS–polyacrylamide gel electrophoresis (PAGE) in 15% gels, and then electrophoretically transferred to polyvinylidene difluoride membranes. The membranes were blocked for 1 h in TBST buffer (0.01 M Tris HCl, pH 7.5, 0.15 M NaCl and 0.05% Tween 20) containing 3% milk, and incubated with rabbit anti-phospho-p70S6K antibody (1:1000; Cell Signaling Technology), rabbit anti-LC3 antibody (1:500; MBL) or rabbit anti-Beclin 1 antibody (1:100; Santa Cruz Biotechnology) diluted in TBST buffer overnight at 4°C. The membranes were washed three times and incubated with secondary antibody linked to horseradish peroxidase (1:1000; Invitrogen) for 1 h at room temperature. The immunoreactive bands were developed using enhanced chemiluminescence reagents (Amersham Corp.) and digitalized by the LAS-1000 Pro (FUJIFILM, Tokyo, Japan). The band densities were quantified using a scanned densitometric analysis and the Image J 1.42q software program (National Institutes of Health). The quantities of the band densities were normalized to β-tubulin, and thereafter, those quantities were compared among the rapamycin-treated mice, the vehicle-treated mice and the sham controls.
Tissue preparation
At 3 days and 42 days after SCI, the mice were overdosed with an intraperitoneal injection of 100 mg/kg sodium pentobarbital. The mice were transcardially perfused with normal saline, followed by 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) at pH 7.4. For immunohistochemical staining, the spinal cord segments containing the injured site were collected, post-fixed in the same fixative overnight at 4°C, and embedded in paraffin. Serial 7-μm transverse sections around the injured site were mounted on slides. A total of 9 sequential sections at 250 μm intervals were collected, spanning a 2000 μm length in the spinal cord centered at the epicenter. The sections were used for immunohistochemical staining as described in the next section.
Immunohistochemistry
Immunohistochemical staining for LC3 and Beclin 1 was performed using the sections obtained 3 days after SCI. The sections were de-paraffinized and rehydrated, and then were washed in PBS for 10 min, followed by washing with PBS containing 0.3% Tween for 10 min, and blocking with 3% milk and 5% fetal bovine serum (FBS) in 0.01 M PBS for 2 h. The sections were incubated with rabbit anti-LC3 antibody (1:100; MBL), rabbit anti-Beclin 1 antibody (1:100; Santa Cruz Biotechnology) or mouse anti-NeuN antibody (1:100; Chemicon) diluted in PBS overnight at 4°C. After rinsing with PBS, the sections were incubated with goat anti-rabbit IgG Alexa Fluor 594 secondary antibody (1:500; Molecular Probes) or goat anti-mouse IgG Alexa Fluor 488 secondary antibody (1:500; Molecular Probes) for 1 h at room temperature. The sections were mounted with Vectashield containing DAPI to label the nuclei (Vector Laboratories). In each experiment, the sections were stained at the same time.
Counting of LC3-positive cells and Beclin 1-positive cells
Each section was scanned using a laser microscope (BX 51; Olympus, Tokyo, Japan) following the immunochemical staining of LC3 and Beclin 1. The scanned image of the transverse section was displayed on a monitor with a grid using the Photoshop Elements Software Program Version 6.0 (Adobe, CA). Then LC3-positive cells and Beclin 1-positive cells in the section were counted using a manual counter. The LC3-positive cells were defined as those displaying punctate LC3 fluorescent dots (Kabeya et al., 2000; Rami et al., 2008). The LC3-positive cells and the Beclin 1-positive cells were counted in the sections at the epicenter, and on the 250 μm and 500 μm rostral and caudal sides. The sum of the numbers in the five sections was compared among the rapamycin-treated mice, the vehicle-treated mice, and the sham controls.
White matter staining
The serial 7-μm transverse sections cut at 250 μm intervals around the epicenter 42 days after SCI were stained with luxol fast blue for myelin. The section with the least amount of luxol fast blue stain was selected as the lesion epicenter. A total of nine sequential sections spanning 2000 μm of spinal cord length, which included four sections both rostral and caudal to the section at the epicenter, were assessed. The images of the stained sections were captured by a digital photographic camera, and the spared white matter area of the spinal cord was analyzed using the Image J 1.42q software program. After performing the luxol fast blue staining, the spared white matter appeared dark blue and isocellular, as seen in healthy neuronal tissue. The damaged or degenerated white matter appeared to be either blanched or replaced by scar tissue having clusters of cells with prominent basophilic nuclei (Bao et al., 2011; Joshi and Fehlings, 2002; Steward et al., 1999). We analyzed the spared white matter area in the rapamycin-treated and the vehicle-treated mice.
Counting of NeuN-positive cells
After the immunohistochemical staining of NeuN as described previously, each section was scanned using a microscope (BX 51; Olympus, Tokyo, Japan). To investigate neuronal loss in the SCI, the number of NeuN-positive cells in each section 42 days after injury was counted. The NeuN-positive cells were defined as cells double labeled with NeuN and DAPI. The section at the epicenter, and 250 μm rostral and caudal sections were chosen for each animal. The cell counting procedure was the same as that described previously. The sum of the numbers in the three sections was compared between the rapamycin-treated and the vehicle-treated mice.
Counting of TUNEL-positive cells
To detect DNA fragmentation caused by cell death in the injured spinal cord, terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) staining was applied using the In Situ Cell Death Detection Kit (Roche, Mannheim, Germany) for the transverse sections obtained at 3 days after injury. The labeled sections at the lesion epicenter and the sites 250 μm caudal and rostral to the lesion were scanned with a BX 51 microscope. The number of TUNEL-positive cells in each section was counted. The TUNEL-positive cells were defined as cells double labeled with TUNEL and DAPI. The cell counting procedure was the same as that described previously. The sum of the numbers in the three sections for each animal was calculated and compared between the rapamycin-treated and the vehicle-treated mice.
Statistical analysis
Significant differences in the number of LC3-positive cells, the number of Beclin 1-positive cells, the number of TUNEL-positive cells, the band densities obtained from the Western blot analyses, and in the spared white matter area, were analyzed using the unpaired t test. The significance of any differences in the BMS scores at each time point was determined by Mann–Whitney's U test. In all analyses, a p value<0.05 was considered to be statistically significant. All statistical analyses were performed using the GraphPad Prism 5.0a software program (GraphPad Software, Inc. La Jolla, CA).
Results
BMS locomotor scores
To evaluate the effect of rapamycin treatment on locomotor recovery after SCI, the BMS total scores and subscores were measured for 6 weeks (Fig. 1A, B). The total BMS scores in the rapamycin-treated mice and in the vehicle-treated mice increased after 4 h and reached a plateau at 28 days after injury. The averages of the total BMS scores were consistently higher in the rapamycin-treated mice than in the vehicle-treated mice from 14 days to 42 days after injury. The rapamycin-treated mice had significantly higher total BMS scores than the vehicle-treated mice from 21 days to 42 days. At 42 days after injury, the total BMS scores of the rapamycin-treated mice were 7–9 (8.4±0.4). Except for one rapamycin-treated mouse with a BMS score of 7, the other four rapamycin-treated mice consistently achieved coordinated plantar stepping with normal trunk stability or with mild trunk instability at 42 days. On the other hand, the BMS scores of the vehicle-treated mice were 5–8 (6.8±0.5). Four out of 5 vehicle-treated mice did not keep their paws parallel when stepping, or they showed severe trunk instability such as leaning, waddling, or near collapse of the hindlimbs.
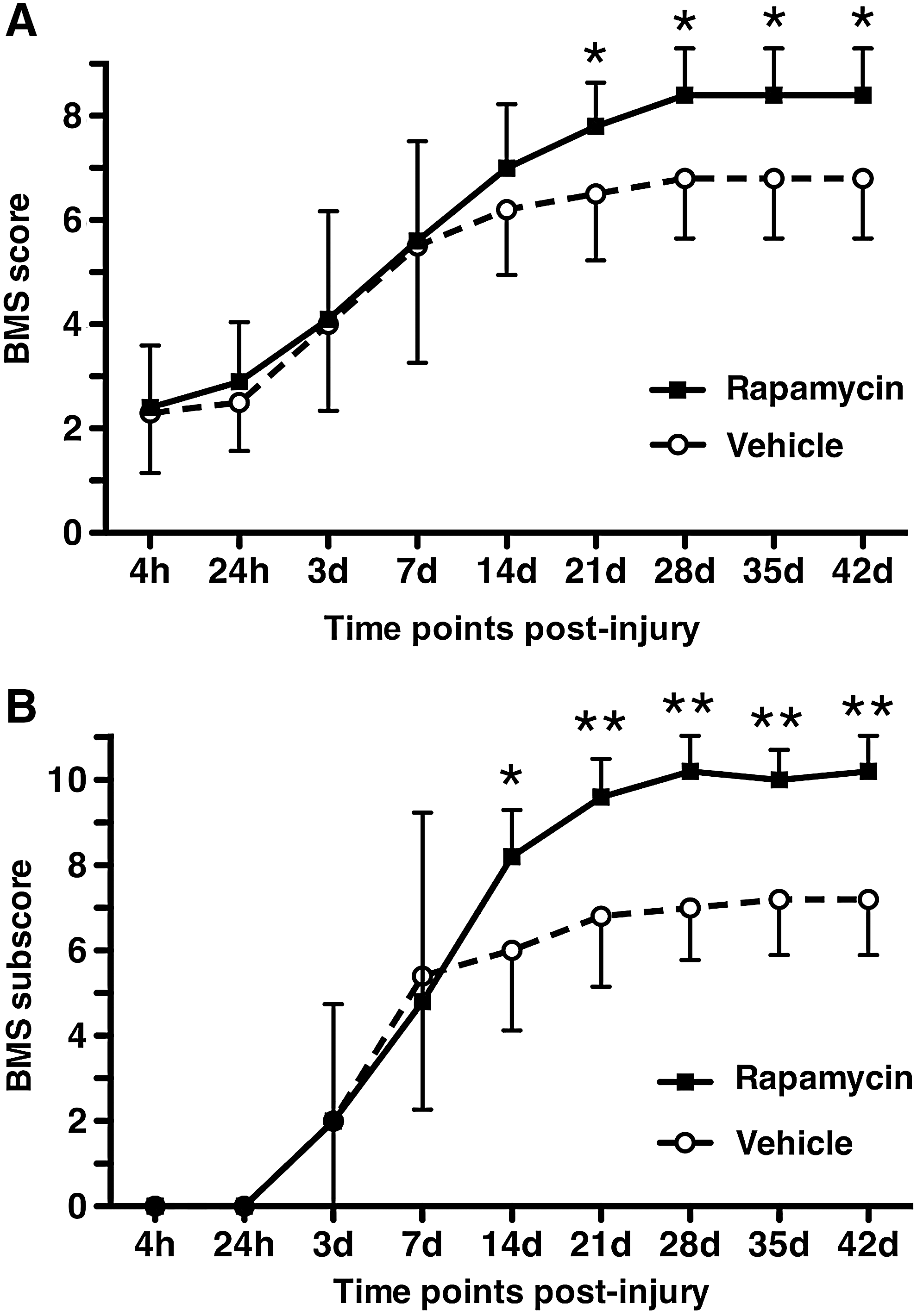
Locomotor recovery after SCI in vehicle-treated mice and rapamycin-treated mice. The BMS scores
The BMS subscores of both the rapamycin-treated mice and the vehicle-treated mice increased after 24 h and reached a plateau at 28 days. The average subscores were consistently higher in the rapamycin-treated mice than in the vehicle-treated mice from 14 days to 42 days. The BMS subscore was significantly higher in the rapamycin-treated mice than in the vehicle-treated mice from 14 days to 42 days after injury.
Phosphorylation of p70S6K
To examine the effectiveness of the rapamycin treatment on the mTOR signaling pathway in the mice, the phosphorylation of p70S6K was evaluated by Western blot analysis. The phosphorylated p70S6K protein was decreased in the rapamycin-treated mice compared with the vehicle-treated mice and the sham controls at 24 h and 3 days after injury (Fig. 2A). Based on the analysis of the band density, the level of phosphorylated p70S6K was significantly lower in the rapamycin-treated mice (2.318±0.866) than in the vehicle-treated mice (8.773±1.083) and in the sham controls (10.000±1.794) (p=0.002 and 0.005, respectively) at 24 h (Fig. 2B). The level of phosphorylated p70S6K at 3 days did not show any statistical differences among the three groups.
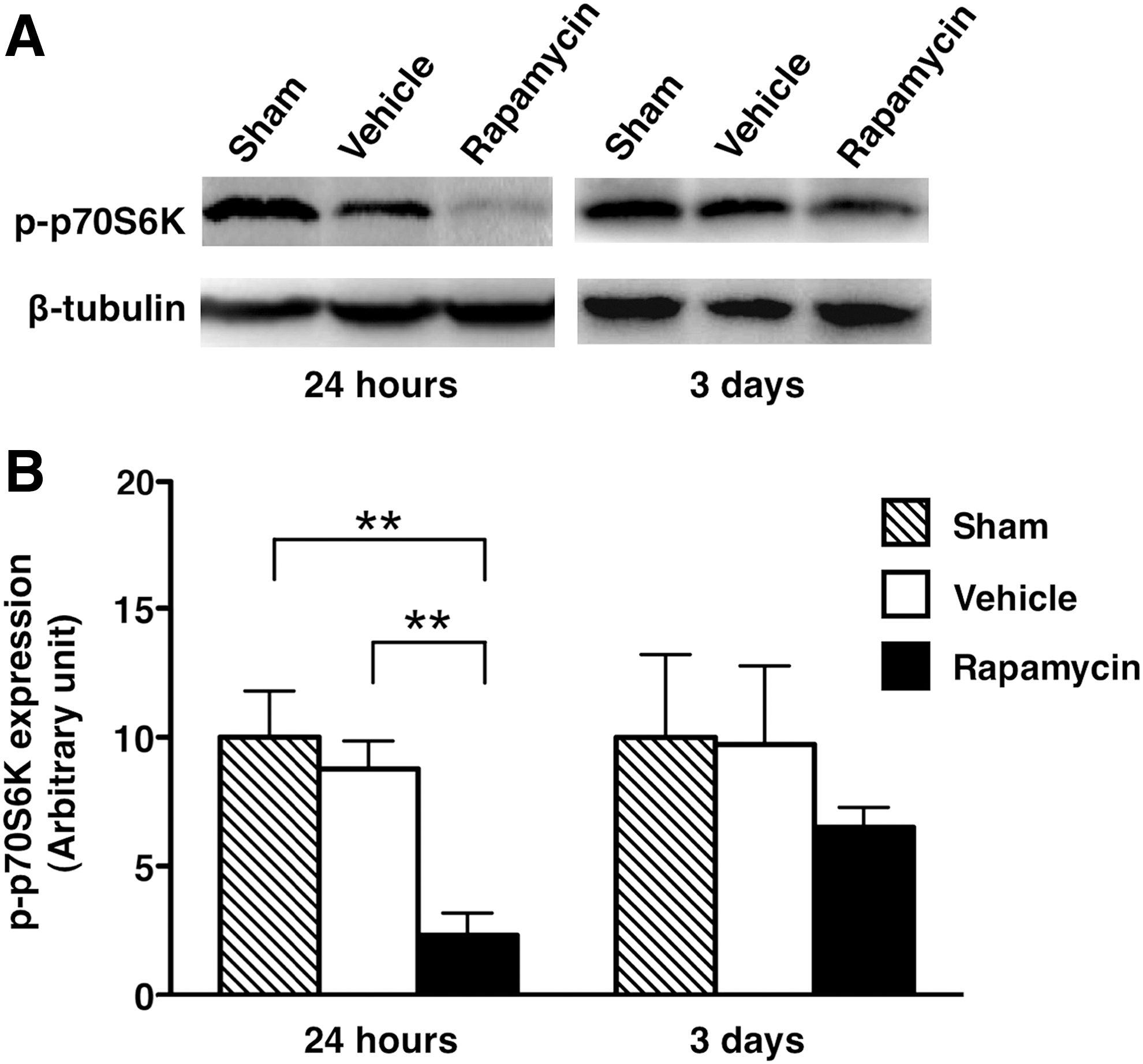
The expression of phosphorylated p70S6K in sham control, vehicle-treated mice and rapamycin-treated mice.
Immunohistochemical and Western blot analyses of LC3 expression
To investigate the activation of autophagy after rapamycin treatment, immunohistochemical staining of LC3 was performed, and the number of LC3-positive cells was counted. The cells expressing LC3 were increased in the rapamycin-treated mice and in the vehicle-treated mice compared with the sham controls (Fig. 3A–I,S). The population of cells expressing LC3 in the rapamycin-treated mice was obviously higher than that in the vehicle-treated mice. Higher magnification revealed that the LC3-expressing cells displayed punctate LC3 dots that were located in the cytoplasm (Fig. 3J–R). The number of LC3-positive cells was significantly increased in the vehicle-treated mice and the rapamycin-treated mice compared with the sham controls (p=0.012 and<0.001, respectively). In addition, the number of LC3-positive cells in the rapamycin-treated mice was significantly higher than in the vehicle-treated mice (p=0.018) (Fig. 3T).
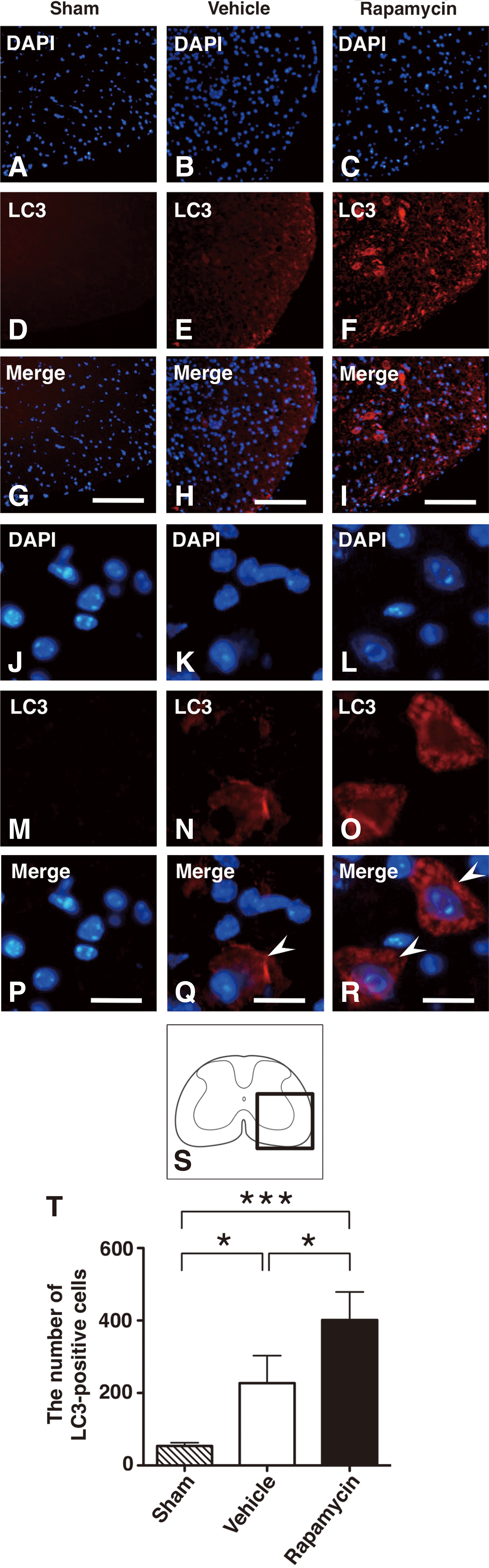
Immunohistochemical staining of LC3 in sham control, vehicle-treated mice and rapamycin-treated mice.
In the Western blot analysis, the expression of LC3-II protein was increased in the rapamycin-treated mice compared with the vehicle-treated mice and the sham controls (Fig. 4A). In the analysis of the band density, the level of LC3-II expression was significantly higher in the rapamycin-treated mice (7.274±0.783) and vehicle-treated mice (4.717±0.532) than in the sham controls (1.000±0.380) (p=0.002 and 0.001, respectively). In addition, the LC3-II expression was significantly higher in the rapamycin-treated mice than in the vehicle-treated mice (p=0.035) (Fig. 4B).
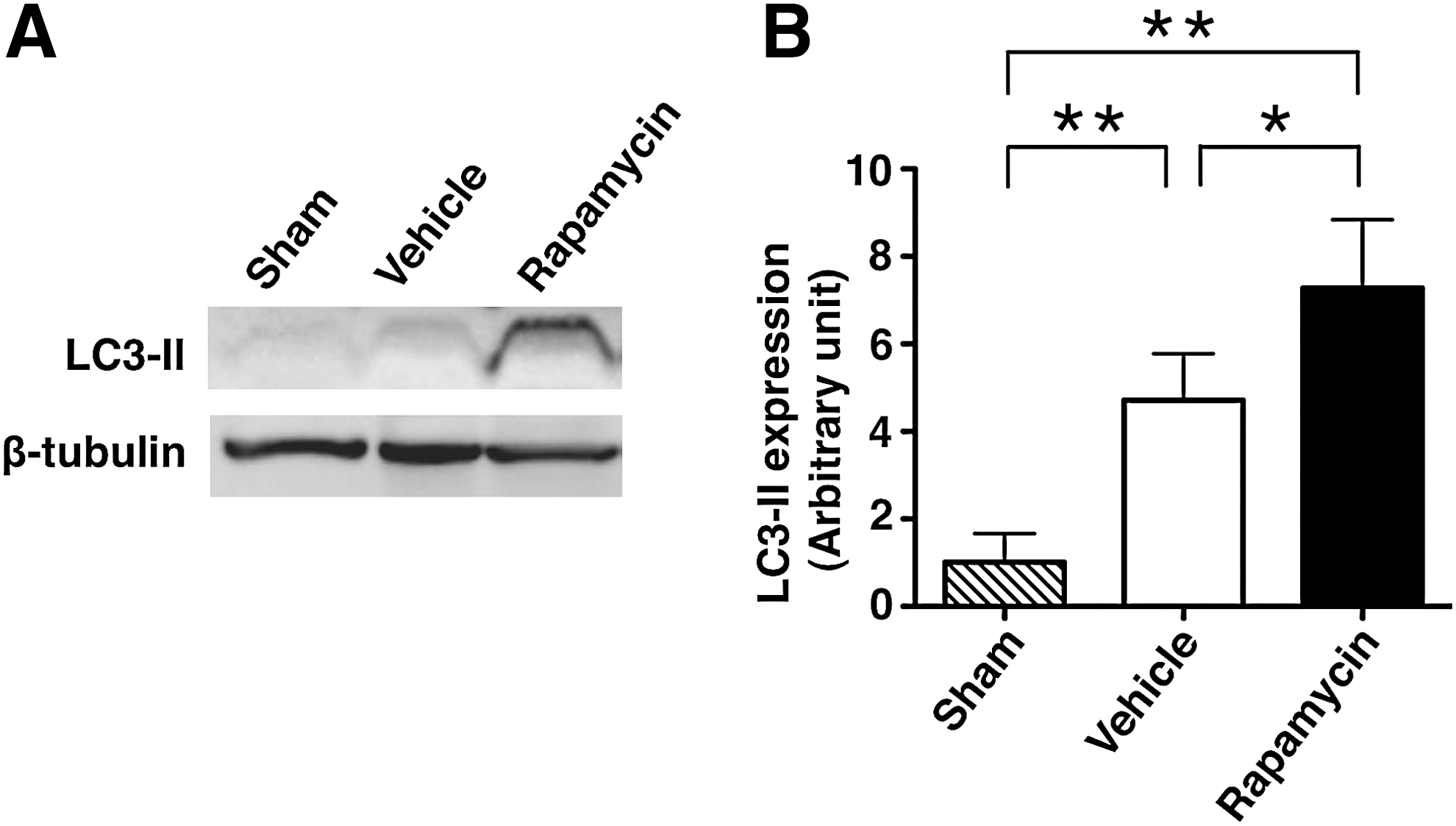
The expression of LC3-II in sham controls, vehicle-treated mice and rapamycin-treated mice.
Immunohistochemical and Western blot analyses of Beclin 1 expression
To analyze the autophagic activity, we also performed immunohistochemistry for Beclin 1, and counted Beclin 1-positive cells. The Beclin 1 expressing cells were clearly increased in the rapamycin-treated mice and in the vehicle-treated mice compared with the sham controls (Fig. 5A–I,S). The number of Beclin 1-positive cells was significantly increased in the rapamycin-treated mice and the vehicle-treated mice compared with the sham controls (p=0.005 and <0.001, respectively) (Fig. 5T).
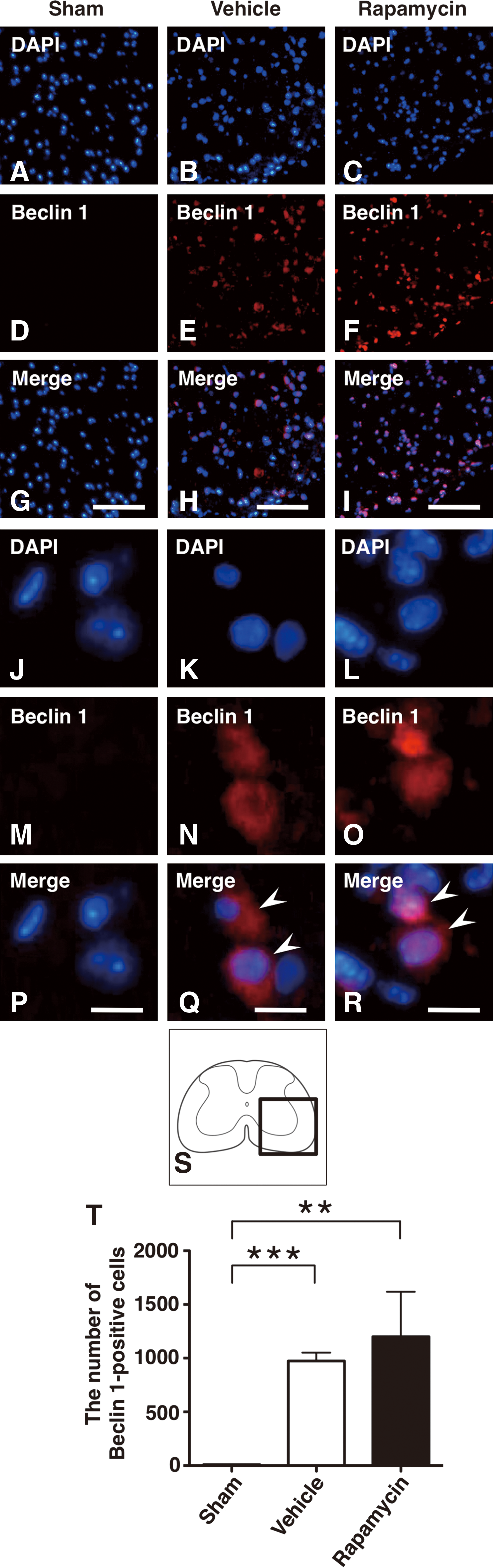
Immunohistochemical staining of Beclin 1 in sham control, vehicle-treated mice and rapamycin-treated mice.
In the Western blot analysis, the expression of Beclin 1 protein was significantly increased in the rapamycin-treated mice and the vehicle-treated mice compared with in the sham controls (Fig. 6A, B). In the analysis of the band density, the level of Beclin 1 expression was significantly higher in the rapamycin-treated mice (7.411±0.882) and vehicle-treated mice (6.506±0.609) than in the sham controls (1.000±0.594) (p<0.001 and 0.001, respectively). In addition, the Beclin 1 expression in the rapamycin-treated mice was relatively higher, but not significantly so, than that in the vehicle-treated mice.
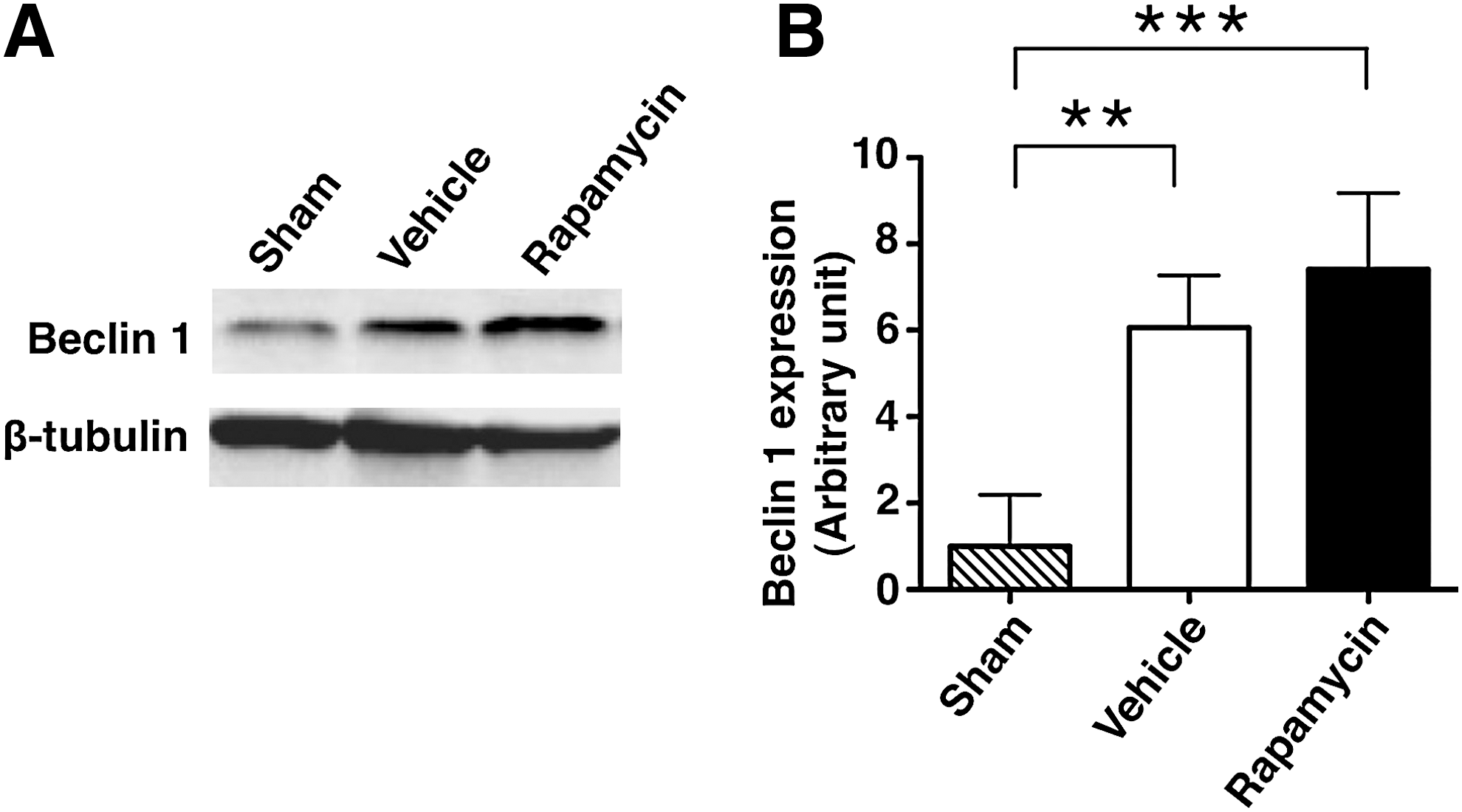
The expression of Beclin 1 in sham control, vehicle-treated mice and rapamycin-treated mice.
Areas of spared white matter
To investigate the differences in the amounts of demyelination after the injury, the spared white matter areas were compared between the rapamycin-treated and the vehicle-treated mice using luxol fast blue staining. The areas of spared white matter in the rapamycin-treated mice were obviously larger than in the vehicle-treated mice, as indicated by the luxol fast blue staining of the sections around the epicenter (Fig. 7A,B). The white matter area was more preserved in the rapamycin-treated mice than in the vehicle-treated mice, especially in the dorsal side of the spinal cord (Fig. 7B). In quantitative analysis of the spared white matter area, the averages of spared white matter area in the rapamycin-treated mice were not significantly but consistently larger than in the vehicle-treated mice at >500 μm rostral and caudal sides from the epicenter (Fig. 7C).
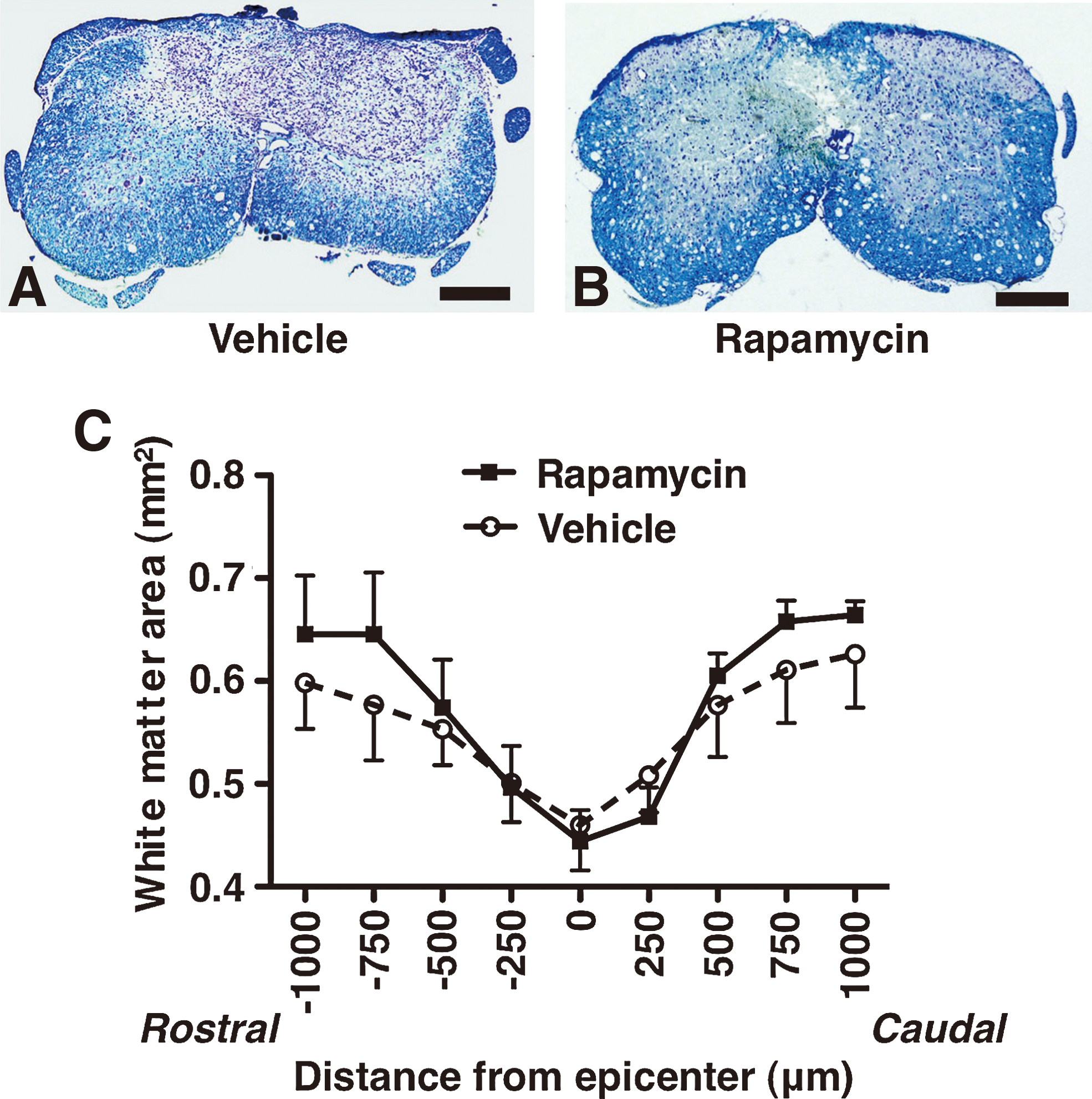
White matter sparing in the vehicle-treated mice and rapamycin-treated mice at 42 days after SCI. Representative spinal cord sections at the 750 μm rostral side from the epicenter showed that the spared white matter area was relatively smaller in the vehicle-treated mice
The number of NeuN-positive cells
To investigate loss of neural cells after injury, the number of NeuN-positive cells was compared between the vehicle and the rapamycin-treated mice by immunohistochemical staining. The NeuN-positive cells were sparsely observed at the epicenter in the vehicle-treated mice. In contrast, the rapamycin-treated mice showed a higher number of NeuN-positive cells compared with the vehicle-treated mice (Fig. 8A–M). The number of NeuN-positive cells in the rapamycin-treated mice was significantly higher than those in the vehicle-treated mice at 42 days (p=0.012) (Fig. 8N).
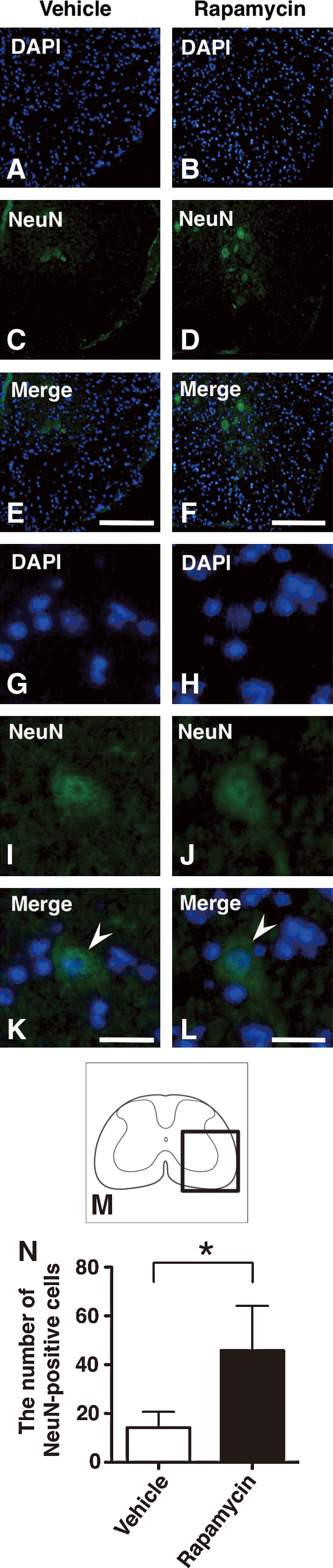
Immunohistochemical staining of NeuN in vehicle-treated mice and rapamycin-treated mice at 42 days after SCI. Representative sections at the epicenter showed that there were fewer NeuN-positive cells in the vehicle-treated mice
The number of TUNEL-positive cells
To investigate the effect of rapamycin on cell death after SCI, we performed TUNEL staining, and the number of TUNEL-positive cells was compared between the vehicle- and the rapamycin-treated mice. In the TUNEL staining of sections obtained at 3 days after injury, the TUNEL-positive cells were obviously decreased in the rapamycin-treated mice compared with the vehicle-treated mice (Fig. 9A–M). When the TUNEL-positive cells were counted, the number of TUNEL-positive cells was significantly lower in the rapamycin-treated mice compared to the vehicle-treated mice (p=0.008) (Fig. 9N).
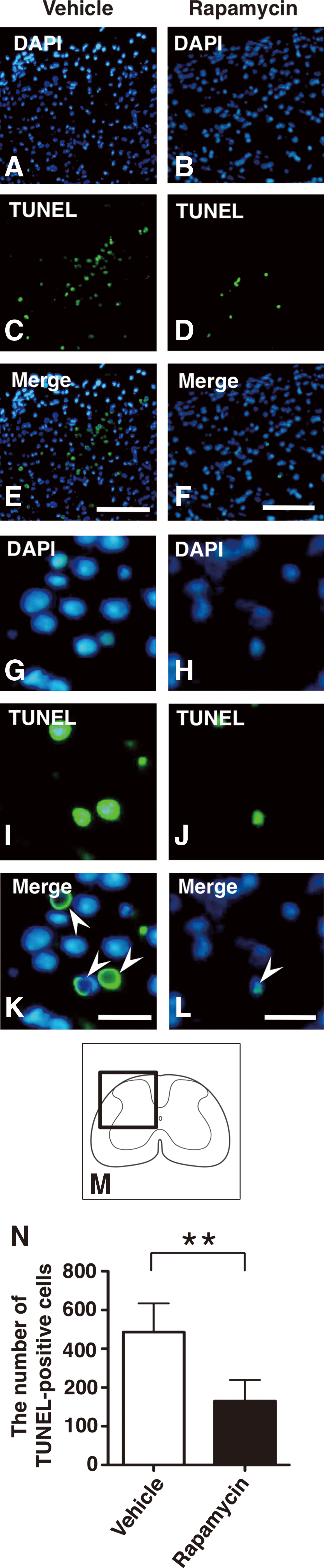
TUNEL staining in vehicle-treated mice and rapamycin-treated mice at 3 days after SCI. Representative sections at the epicenter showed that there were obviously fewer TUNEL-positive cells in the rapamycin-treated mice
Discussion
In the present study, we demonstrated that the administration of rapamycin significantly decreased the phosphorylation of p70S6K protein and led to higher expressions of LC3 and Beclin 1 in the injured spinal cord. In addition, neuronal loss and cell death in the injured spinal cord were significantly more reduced in the rapamycin-treated mice than in the vehicle-treated mice. Furthermore, the rapamycin-treated mice showed significantly higher locomotor function in BMS scores than did the vehicle-treated mice after SCI. These results demonstrated that the administration of rapamycin promoted autophagy by inhibiting mTOR signaling pathway and reduced neural tissue damage and locomotor impairment after SCI. The present study indicated that rapamycin treatment had a neuroprotective effect at the lesion site following SCI.
Previous studies demonstrated that rapamycin treatment inhibits the mTOR signaling pathway and activates autophagy in several neurodegenerative diseases (Pan et al., 2008; Ravikumar et al., 2004). Additionally, rapamycin enhanced autophagic activity after traumatic brain injury and neonatal hypoxia–ischemia induced brain injury (Carloni et al., 2008; Erlich et al., 2007a). These data indicate that rapamycin promotes autophagy by inhibiting mTOR in the CNS. In the present study, rapamycin treatment significantly reduced the phosphorylation of p70S6K protein in the injured spinal cord. The reduction of phosphorylated p70S6K indicated that administration of rapamycin actually inhibited mTOR signaling after SCI. Our results also showed that rapamycin treatment induced upregulation of LC3 and Beclin 1 expression in the damaged neural tissue after SCI. These findings demonstrated that rapamycin promoted autophagy by inhibiting mTOR pathway at the lesion site following SCI.
The autophagic activity is regulated by various molecular factors including mTOR, Beclin 1, Bcl family, DRAM, p53, and NF-κB (Maiuri et al., 2007; Scarlatti et al., 2009). Previous research has suggested that Beclin 1 regulates not only the autophagic activity, but also the cell death mechanism (Maiuri et al., 2007). Beclin 1 interacts with Bcl-2, which is known to be an anti-apoptotic protein (Erlich et al., 2007b). Therefore, the level of Beclin 1 expression may not directly indicate the level of autophagic activity. In this study, the expression of LC3, a marker of autophagy, significantly increased due to the administration of rapamycin after SCI. However, the increase in the Beclin 1 expression after the rapamycin treatment was not statistically significant. These findings suggested that the activation of autophagy after rapamycin treatment therefore did not directly indicate the upregulation of Beclin 1 expression after SCI.
The inhibition of mTOR by rapamycin can cause either beneficial or detrimental effects, depending upon the disease model and timing. Previous studies suggested that rapamycin treatment aggravated renal ischemia/reperfusion injury (Gonçalves et al., 2007; Lui et al., 2006). Rapamycin blocks the cardioprotective effect of ischemic or pharmacological preconditioning for myocardial infarction (Jonassen et al., 2001; Kis et al., 2003). On the other hand, rapamycin treatment had a cytoprotective effect, including reduction of infarct size and decreasing cell death caused by myocardial ischemia/reperfusion injury (Khan et al., 2006). The enhancement of autophagy using rapamycin also produces a neuroprotective function in various diseases of the CNS. Previous studies demonstrated that administration of rapamycin rescued neuronal cells in models of Huntington's disease and Parkinson's disease (Malagelada et al., 2010; Ravikumar et al., 2004). Interestingly, rapamycin treatment reduces neural tissue damage after traumatic brain injury and neonatal hypoxia–ischemia induced brain injury (Carloni et al., 2008; Erlich et al., 2007a). In the present study, rapamycin prevented the neuronal loss after SCI. Moreover, the rapamycin-treated mice showed significantly better locomotor recovery in BMS scores. The present study suggested that administration of rapamycin provided a beneficial effect by reducing the neural tissue damage and locomotor impairment following SCI.
Rapamycin has cytoprotective effects that have been attributed to blocking the apoptotic signaling pathway (Carloni et al., 2008; Khan et al., 2006; Pan et al., 2008). It has been suggested that rapamycin enhances the clearance of mitochondria by autophagy, thereby reducing cytosolic cytochrome c release and downstream caspase activation (Ravikumar et al., 2006). Previous studies demonstrated rapamycin to have a cytoprotective function by reducing apoptosis in various disease models. For example, rapamycin treatment significantly reduced the number of TUNEL-positive cells in myocytes in a myocardial ischemia-reperfusion model (Khan et al., 2006). Rapamycin promoted autophagy and inhibited apoptosis in a model of lactacystin-induced neurodegeneration (Pan et al., 2008). Activation of autophagy by rapamycin also reduced the expression of activated caspase-3 in a model of neonatal hypoxia-ischemia induced brain injury (Carloni et al., 2008). Interestingly, in these disease models of the CNS, the activity of autophagy was upregulated even without the administration of rapamycin and it was further enhanced by rapamycin treatment (Carloni et al., 2008; Pan et al., 2008). These reports suggested that the rapamycin treatment further promoted autophagic activity and reinforced its cytoprotective function to reduce cell death in such diseases of the CNS. In the present study, rapamycin further promoted the activity of autophagy that had been upregulated after SCI. Additionally, the rapamycin treatment significantly reduced the number of TUNEL-positive cells at 3 days after SCI. These findings suggested that rapamycin treatment enhanced the activity of autophagy and thereby prevented cell death in the damaged neural tissue after SCI.
The administration of rapamycin in this study was performed using a single injection at 4 h after SCI. Many previous studies that investigated the pharmacokinetics of rapamycin in vivo showed the half-life of rapamycin to be relatively long (33–63 h) (Chen et al., 2007; Napoli et al., 1997). In fact, several reports demonstrated that a single administration of rapamycin could affect the autophagic activity and cell death for >24 h (Carloni et al., 2008; Sheng et al., 2010). In the present study, the single administration of rapamycin at 4 h actually inhibited the phosphorylation of p70S6K in mTOR signaling pathway at 24 h after SCI, and produced a significant reduction in the degree of neuronal loss and cell death. These results indicated the rapamycin treatment at 4 h could reduce the secondary injury that mainly occurs from 24 h to 3 days after the onset of SCI (Beattie et al., 2000; Yong et al., 1998). The present study suggested that the administration of rapamycin produces a cytoprotective effect at the lesion site. Therefore, the administration of rapamycin may represent a novel therapeutic strategy to reduce secondary injury following SCI.
Recent studies have indicated that the inhibition of mTOR by rapamycin can suppress the regeneration of damaged neural tissue. The inhibition of mTOR by rapamycin suppresses growth cone regeneration after axonotomy (Verma et al., 2005). Axonal regeneration activated by the administration of adenosine-5’-triphosphate (ATP) in the injured spinal cord can decrease because of the administration of rapamycin (Hu et al., 2010). Rapamycin reduces the ability of axonal regeneration that was promoted by the deletion phosphatase and tensin homolog (PTEN) in the CNS (Park et al., 2008). These studies suggested that the inhibition of mTOR by rapamycin suppressed new protein synthesis and cell proliferation to promote axonal regeneration. On the other hand, a few reports have indicated that rapamycin could promote a permissive environment for neuroregeneration. Rapamycin treatment reduced reactive gliosis at the lesion site in a spinal cord ischemia model (Codeluppi et al., 2009). Rapamycin activates autophagy and improves myelination in a demyelinating neuropathy model (Rangaraju et al., 2010). In this study, the inhibition of mTOR by rapamycin during the acute phase of SCI significantly reduced the neural tissue damage and locomotor impairment. Therefore, the administration of rapamycin during the acute phase should induce a cytoprotective effect to reduce the secondary injury of SCI. Further studies may clarify the effect of rapamycin on regulating neuroprotection and neuroregeneration and lead to a novel therapeutic strategy after SCI.
Footnotes
Acknowledgments
This work is supported by Grant-in-Aid for Scientific Research (No. 19591712) from Japan Society for the Promotion of Science, by Japan Orthopaedics and Traumatology Foundation, Inc. (No. 0138), and by Grant in 2007 from The General Insurance Association of Japan. We thank Dr. Koshi N. Kishimoto for useful discussion. We also thank Mr. Katsuyoshi Shoji for technical assistance, and Ms. Teruko Sueta and the animal care team at Institute for Animal Experimentation in Tohoku University for the animal care in this study.
Author Disclosure Statement
No competing financial interests exist.