
Abstract
Recombinant adeno-associated viral (rAAV) vectors expressing neurotrophic genes reduce neuronal death and promote axonal regeneration in central nervous system (CNS) injury models. Currently, however, use of rAAV to treat clinical neurotrauma is problematic because there is a delay in the onset of transgene expression. Using the adult rat retina and optic nerve (ON), we have tested whether rAAV gene therapy administered at the time of injury combined with short-term pharmacotherapy has synergistic effects that enhance neuronal survival and regeneration. The ON was transected and a 1.5 cm segment of autologous peripheral nerve (PN) was grafted onto the cut end. At this time, bicistronic rAAV2 encoding ciliary neurotrophic factor (CNTF) and green fluorescent protein (rAAV2-CNTF-GFP) was injected into the injured eye. To provide interim support for axotomized retinal ganglion cells (RGCs) during vector integration and therapeutic transgene expression, rCNTF protein and a cyclic adenosine monophosphate (cAMP) analogue (CPT-cAMP) were injected intravitreally 3 and 10 days postoperatively. For comparison, another rAAV2-CNTF-GFP group received two intravitreal saline injections 3 and 10 days after the PN–ON surgery. A further PN graft group received only postoperative intravitreal injections of rCNTF plus CPT-cAMP. After 4 weeks, regenerating RGCs were retrogradely labelled by applying fluorogold to the distal end of each PN graft. Compared to saline-injected animals, both RGC survival and axonal regrowth were significantly higher in the rCNTF and CPT-cAMP injected rAAV2-CNTF-GFP group; approximately one third of the RGC population survived axotomy, and 27% of these regrew an axon. These values were also higher than those obtained in rats that received only rCNTF plus CPT-cAMP injections. Therefore, we show for the first time that rAAV-mediated gene delivery at the time of, or just after, neurotrauma is most successful when combined with temporary post-injury trophic support, and is potentially a viable treatment strategy for patients after acute CNS injury.
Introduction
R
In animal studies, application of recombinant (r) neurotrophic factors enhances the survival of injured neurons and stimulates axonal regrowth following injury in many parts of the CNS (Alcala-Barraza et al., 2010; Cui, 2006; Lu and Tuszynski, 2008). However, neurotrophic factors often have a short half-life in biological systems (Dittrich et al., 1994) and minipumps or multiple injections are usually required to achieve more long-term protection (Cui and Harvey, 2000; Cui et al., 2003). Sustained delivery using viral vectors may be even more effective. Indeed, in studies in the adult rodent visual system, a system often used to elucidate CNS repair mechanisms (Benowitz and Yin, 2008; Harvey et al., 2006), a single intraocular injection of rAAV2 encoding ciliary neurotrophic factor (CNTF), promotes greater regeneration of injured retinal ganglion cells (RGCs) compared to multiple rCNTF eye injections (Leaver et al., 2006). In the eye, rAAV2 integration and transgene activation has reached high levels of therapeutic protein expression by 2 weeks (Harvey et al., 2002); therefore, in the work of Leaver and associates (2006), the rAAV2-CNTF-GFP vector was injected into the eye 10 days prior to optic nerve (ON) crush or transection. Other visual system studies have injected vectors between 7 and 21 days before ON injury (Cheng et al., 2002; Malik et al., 2005; Planchamp et al., 2008) and in rodent stroke models, rAAV vectors encoding growth factors have almost always been injected prior to acute arterial occlusion (Shen et al., 2008; Zhu et al., 2008; Airavaara et al., 2010). There has been one report describing delivery of rAAV after ischemia (Zhu et al., 2009), and vectors have also been applied at the time of injury in a small number of spinal cord studies (Chen et al., 2007; Ruitenberg et al., 2004).
In experimental animal models, initiation of rAAV-mediated gene expression before an injury provides important information on the therapeutic potential of a particular protein, but the use of gene therapy as a prophylactic for random and unpredictable CNS trauma is clearly not possible in the human context. In the present study, we therefore first compared RGC survival and regeneration using the more clinically relevant approach of injecting rAAV2-CNTF-GFP vectors at the time of ON injury with data obtained from previous pre-injury rAAV2-CNTF-GFP treatments (Hellström and Harvey, 2011; Leaver et al., 2006). As in earlier studies, a peripheral nerve (PN) autograft was sutured onto the cut ON (PN–ON graft) to provide a more favourable environment for axonal regeneration (Aguayo et al., 1991; Harvey al., 2006). We then examined whether additional pharmacotherapeutic treatment applied after the injury and during the time taken for rAAV2 vector integration and transgene activation could elicit even greater RGC protection and regeneration. To provide interim trophic support, rCNTF protein and a cAMP analogue (8-[4-Chlorophenylthio]-adenosine-3′,5′-cyclic monophosphate; CPT-cAMP), a combination known to promote RGC viability and axonal regrowth (Cui et al., 2003; Park et al., 2004), were injected intravitreally at 3 and 10 days post-injury. For comparison, another group of rats injected with rAAV-CNTF-GFP at the time of the PN–ON graft received two intravitreal saline injections 3 and 10 days post-injury; therefore, they did not receive additional temporary trophic support. For further comparisons, an additional PN-grafted group received only postoperative intravitreal injections of rCNTF plus CPT-cAMP. Four weeks after surgery, the number of RGCs regenerating an axon into the grafts was determined using the retrograde tracer fluorogold (FG). Surviving and transduced RGC numbers were estimated using βIII-tubulin+ and GFP+ immunohistochemistry, respectively (Cui et al., 2003).
Methods
Experimental groups, surgical procedures and intravitreal eye injections
Twelve female Wistar rats (8–10 weeks old, Animal Resources Centre, WA) were used for the rAAV-CNTF-GFP experiment. Rats were anesthetized with a 1:1 mixture (1.5 mL/kg) of ketamine (100 mg/mL) and xylazine (20 mg/mL). and intraorbital ON axotomy was followed by attachment of a 1.5 cm length of autologous PN graft, as previously described (Cui et al., 2003; Hellström et al., 2011; Hu et al., 2007). Eye ointment containing atropine sulphate (10 mg/g, Troy Ilium) was applied to protect the cornea during surgery. The grafts consisted of a segment of tibial nerve, a branch of the sciatic nerve, and were sutured onto the cut ON ∼ 1.5 mm behind the optic disk using 10/0 Ethilon suture (Johnson & Johnson, North Ryde, NSW, Australia) without compromising retinal blood supply. Rats also received a subcutaneous injection of buprenorphine (0.02 mg/kg, Temgesic; Reckitt & Colman, Hull, UK) and an intramuscular injection of Benacillin (0.3 mg/kg, Troy Ilium; NSW, Australia). The distal end of the PN graft was placed over the skull and secured to connective tissue.
At the time of the PN–ON graft, 4 μL of rAAV2-CNTF-GFP was intravitreally injected (1 x 1012 GC/mL) into the operated eye. The rAAV2-CNTF-GFP vector was the same vector construct that was used by Leaver and associates (2006), and was precisely the same viral vector batch and titer (1×1012 GC/mL) used for the studies described by Hellström and associates (2011). In brief, the rAAV2-CNTF-GFP was commercially produced by GTC Virus Vector Core (NC, USA) and was generated from the pTRUF12.1 plasmid. The CNTF gene included an NGF signal sequence to enable secretion of the vector-produced CNTF protein (Sendtner et al., 1992). The gene sequence also contained a post IRES site that allowed for a bicistronic GFP expression, hence resulting in two individual proteins (CNTF and GFP). Expression was driven by a cytomegalovirus-chicken β-actin promoter. Previous studies using quantitative polymerase chain reaction (qPCR), Western blot, and immunohistochemistry verified vector-induced mRNA and protein expression of the secretable rAAV2-CNTF construct in the retina (Leaver et al., 2006; Liang et al., 2001).
Under isoflurane anaesthesia, six PN-grafted rats were given a 3 μL intravitreal injection of rCNTF (0.5 μg/μL, Peprotech, Rehovot, Israel) plus CPT-cAMP (0.1 mM, Sigma) at days 3 and 10 post-injury. The other six PN-grafted animals were given saline injections (3 μL) at the same time points. (For detailed explanations regarding practical techniques, see Hellström, et al., 2011; Leaver et al., 2006). One rCNTF plus CPT-cAMP injected rat formed a cataract post-surgery and was excluded from further analysis. Nine additional PN–ON grafted rats (8–10 weeks old) were not injected with rAAV-CNTF-GFP but instead received postoperative intravitreal injections of rCNTF (0.5 μg/μL) plus CPT-cAMP (0.1 mM; total volume 3 μL). This combination was also administered 3 and 10 days after surgery, but these rats also received an additional injection of rCNTF plus CPT-cAMP on day 17, a protocol shown previously to promote extensive RGC survival and axonal regeneration (Cui et al., 2003; Park et al., 2004). All PN–ON grafted rats survived for 4 weeks prior to perfusion and subsequent retinal analysis.
Retrograde labelling, immunohistochemistry, quantification and analysis
Three days before perfusion, rats were anesthetized with a 1:1 mixture (1.5 mL/kg) of ketamine (100 mg/mL) and xylazine (20 mg/mL) and 0.5μL of 4% FG (Fluorochrome LLC, CO, USA) was slowly injected into the distal end of each PN graft to retrogradely label RGCs that had regenerated an axon at least 1 cm into the grafts. After 3 days, rats were deeply anesthetized (Lethabarb, Verbac, NSW; i.p; 2 μL/g) and transcardially perfused with cold phosphate-buffered saline (PBS) (pH 7.4) containing 0.1% heparin, then a phosphate buffer containing 4% paraformaldehyde. Retinas were isolated, postfixed for 1 h in the same fixative and whole mounted onto glass slides and viewed under a fluorescent microscope. In each retinal whole mount, the number of regenerating (FG+) cells was quantified at 250×magnification in 50–80 fields sampled across the entire retina (one field=0.235mm×0.235 mm). The outline of each retina was drawn on a computer screen using a MD2 microscope digitizer (Accustage, MN, USA) and a grid was randomly placed over the drawing. A cursor was placed on each grid intersection and the number of FG-labelled RGCs counted in the field at that point (Cui et al., 2003; Park et al., 2004). The counts per field were then averaged for each animal and extrapolated so that an estimate of the total number of regenerating, surviving, and transduced numbers of RGCs could be estimated from each retina, using 58mm2 as a standard size retina (Danias et al., 2002; Gellrich et al., 2002).
To label and visualize surviving and transduced cells, primary antibodies for βIII-tubulin (mouse α-TUJ1 antibody, Covance, NJ, USA; 1:500; RGCs) and GFP antibody (rabbit α-GFP, Millipore, VIC; 1:300; transduced cells) were used followed by fluorescently conjugated secondary antibodies: α-mouse IgG-Cy3 (Jackson Labs, PA, USA) for TUJ1; 1:400 and α-rabbit IgG-Alexa 488 (Invitrogen, VIC) for GFP; 1:400. Surviving (βIII-tubulin+) cells were quantified the same way as the regenerating (FG+) cells. To minimize bleaching of the whole mounts, the total number of GFP+ cells was estimated from images that were taken of 10 randomly selected areas from across the entire retina of each animal (total photographed area=1.41 mm2). Images were not further post-manipulated and the camera settings were set according to the negative control from each staining. The counts were then averaged and extrapolated using the same method as for the FG+ and βIII-tubulin+ counts.
The rAAV-CNTF-GFP vector was injected into the lower temporal region of the eye and RGC transduction was therefore highest in the corresponding region of the retina. In each rat the transduced retinal area (area where GFP+ RGCs were most frequently encountered) and total retinal area were measured (at 40×using the MD2 microscope digitizer) to estimate the extent of vector diffusion and transduction following each intravitreal rAAV2 injection. Cell counts from transduced areas (counting fields containing GFP-expressing cells) versus non-transduced areas (counting fields with very few if any GFP-expressing cells) were logged separately to allow for later comparison. Representative areas from selected retinas from each experimental group were photographed using confocal microscopy. For comparisons between the two rAAV-CNTF-GFP injected groups, the two-tailed Student's t test (paired, and unpaired) was used (significant levels: *p<0.05, **p<0.01, ***p<0.001). Additional comparison of overall RGC viability and regeneration that also included the non-viral group used Kruskall–Wallis post-hoc comparison following one-way analysis of variance (ANOVA).
Results
When analyzed 4 weeks after PN–ON transplantation, in PN-grafted rats that received three intravitreal rCNTF plus CPT-cAMP injections 3, 10, and 17 days after PN–ON surgery, we obtained a mean of 11842±520 (SEM) βIII-tubulin+ surviving RGCs per retina and on average 5476±654 (SEM) regenerating (FG+) RGCs, values similar to those obtained previously (Cui et al., 2003). In comparison, in rats that received a PN graft and intravitreal rAAV2-CNTF-GFP injection followed by two sham saline injections, there were on average an estimated 15,883±2355 (SEM) βIII-tubulin+ surviving RGCs per retina (n=6; Fig. 1). Of these viable RGCs, 3522±901 (SEM) were FG+ and therefore had regenerated an axon as far as the distal end of the autograft. Therefore, ∼22% of surviving RGCs were induced to regenerate an axon when the AAV vector was delivered at the time of injury without further supplementary trophic support.
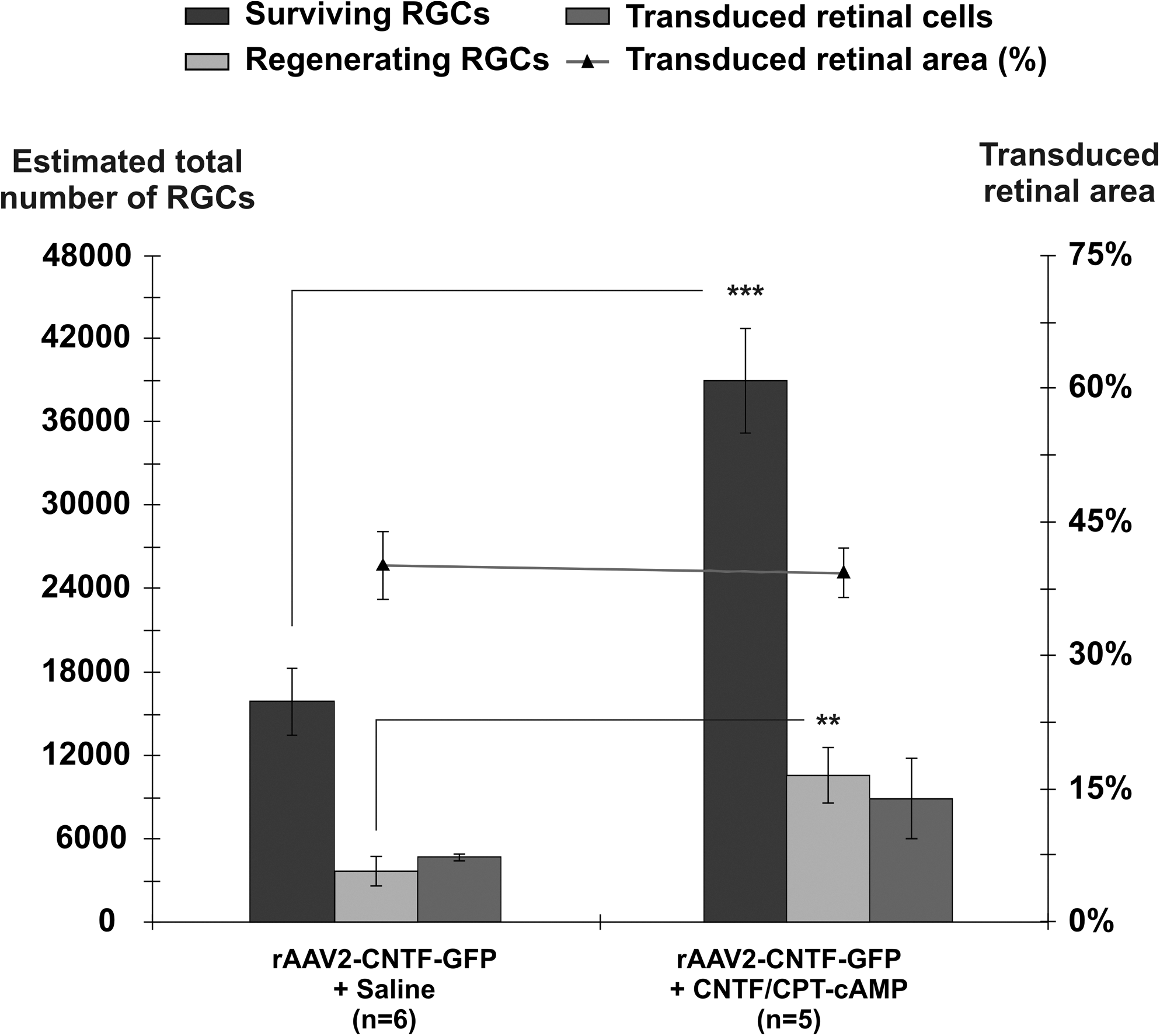
The estimated total number of surviving (βIII-tubulin+), regenerating (fluorogold+) and transduced (GFP+) retinal ganglion cells (RGCs) in retinal whole mounts, 4 weeks after rAAV2-CNTF-GFP vector administration at the time of peripheral nerve–optic nerve (PN–ON) transplantation. Both groups received additional intravitreal injections 3 and 10 days post-injury, of either saline (control) or recombinant CNTF and CPT-cAMP. The numbers of surviving RGCs and RGCs regenerating an axon into the grafts were both significantly increased in the CNTF and CPT-cAMP group. There was no significant difference between estimated transduced retinal areas for the two groups. Two tailed unpaired Student t test; **p<0.01, ***p<0.001. Error bars=+/- SEM.
Overall, a much greater number of RGCs survived the ON injury in the rAAV2-CNTF-GFP injected group that received additional, temporary trophic support (rCNTF and CPT-cAMP) during the period of vector transgene activation (Fig. 1). On average, 38,993 RGCs survived (±3792 SEM; n=5), significantly more than in the group injected post-injury with saline (two-tailed, unpaired Student's t test; ***p<0.001). The estimated total amount of regeneration was also much greater; there were on average 10,567±2035 (SEM) regenerating FG+ RGCs in this group, significantly more than the number seen in the vector plus saline injected group (two-tailed, unpaired Student's t test; **p<0.01) (Fig. 1). These survival and regeneration values were also greater than those seen in PN-grafted rats that received only postoperative rCNTF plus CPT-cAMP injections, despite the fact that the latter group received an additional intravitreal injection of rCNTF plus CPT-cAMP on day 17. Overall, in the combined gene therapy and pharmacotherapy group, ∼27% of the surviving RGC population regenerated an axon.
Because the 4 μL of rAAV vector was injected into the lower temporal region of the eye, as in previous studies, the number of transduced GFP+ RGCs was greatest in this quadrant of the retina, and transduced RGCs were less frequently encountered in retina superior to the optic disk (Hellström and Harvey, 2011; Hellström et al., 2011; Leaver et al., 2006). The areas of retina that were effectively transduced after each rAAV2-CNTF-GFP vector injection were similar between the two gene therapy groups (Fig. 1). Consistent with earlier studies, in both groups the density of surviving and regenerating RGCs was significantly greater in temporal retina where transduction levels were maximal. Representative confocal images of surviving βIII-tubulin+ (red) and regenerate FG+ (blue) RGCs at comparable eccentricities in retinal whole mounts from the two rAAV-CNTF-GFP groups, in both non-transduced and transduced regions of the retina, are shown in Figure 2. rAAV2-CNTF-GFP transduced retinal cells are green (Fig. 2B, D).

Representative fields at comparable eccentricities from retinal whole mounts from PN-grafted eyes after rAAV2-CNTF-GFP injection followed by two saline (
The consistent difference in RGC viability and regenerative capacity in transduced versus non-transduced retinal areas emphasizes the relatively localized nature of rAAV2-related CNTF release on injured and regenerating RGCs (see also Hellström and Harvey, 2011). To quantify this difference, RGC densities in the non-transduced versus transduced areas were calculated for the two vector groups. In the rAAV2-CNTF-GFP plus saline group the mean density was estimated to be 217±27 (SEM) and 472±73 (SEM) RGCs per mm2 in non-transduced versus transduced retinal regions (Fig. 3). By comparison, density in these two regions in the rCNTF plus CPT-cAMP injected group was estimated to be 526±65 (SEM) and 913±105 (SEM) RGCs per mm2, the latter a little less than half the density seen in normal Wistar rat retina. Regenerating RGC densities in the two rAAV-CNTF-GFP groups displayed the same trends (Fig. 3).

Estimated retinal ganglion cell (RGC) densities from rAAV-CNTF-GFP plus saline or rAAV-CNTF-GFP plus recombinant CNTF protein/CPT-cAMP injected groups. Values are shown in RGCs per mm2 retina and were obtained from counts acquired from either the non-transduced (Non-transd.) or transduced (Transd.) retinal areas respectively. Note the high density of surviving RGCs in the estimate derived from counts from the transduced area of the rAAV-CNTF-GFP treated group with additional temporary trophic support. Uninjured rat retina contains a little more than 2000 RGCs per mm2 (Hellström and Harvey, 2011). Two-tailed, paired Student t test; significance levels: *p<0.05, **p<0.01. Error bars=+/- SEM.
Discussion
In research from our laboratory over a number of years, we have typically found that only ∼6000–8000 RGCs survive in adult PN–ON grafted rats in the absence of any other growth-promoting treatments (Cui et al., 2003; Hellström et al., 2011; Hu et al., 2007; Leaver et al., 2006; Park et al., 2004). The new data presented here show that, in PN-grafted animals injected with rAAV2-CNTF-GFP at the time of injury followed by two sham saline injections, on average almost 16,000 RGCs remained viable 4 weeks after the surgery. Therefore, there is a 2.5–3-fold increase in the survival of RGCs following CNTF gene therapy given at the time of or immediately after the injury, even in the absence of interim trophic support. Furthermore, in this group, ∼22% of surviving RGCs regrew an axon into a PN graft.
These new data can be compared with results from adult PN-grafted rats that received intravitreal injections of rCNTF protein in combination with CPT-cAMP (present study, see also Cui et al., 2003; Hu et al., 2007). In our new rCNTF plus CPT-cAMP data, we estimated that ∼12,000 RGCs remained viable 4 weeks after PN–ON surgery, of which ∼46% regenerated an axon. Interestingly, surviving RGC numbers were not significantly different from the rAAV-CNTF-GFP plus saline group; however, in the rCNTF plus CPT-cAMP animals, a significantly greater proportion of RGCs regenerated an axon. These comparisons suggest that early administration of trophic support is especially important in order to maximize the regenerative potential of neurons that survive the initial axotomy. In a study performed in parallel to the work presented here, and using the same parameters, the same surgeon, and an identical viral batch, we obtained higher average values per retina when the AAV2-CNTF-GFP was injected into the eye 2 weeks prior to the PN–ON graft. In that report, we estimated that there were on average 21,457 surviving RGCs and 10,327 regenerating FG+ RGCs (Hellström et al., 2011). Clearly then, application of gene therapy at the time of ON transection does by itself have a measurable therapeutic impact, but rAAV2-CNTF-GFP transduction of RGCs some days prior to the neurotrauma is more effective.
The kinetics of rAAV2 integration and transgene activation after an intravitreal injection is well documented, starting at low levels from ∼ 1 week and reaching high levels of therapeutic protein expression after ∼ 2 weeks (e.g., Auricchio et al., 2001; Harvey et al., 2002; Hellström and Harvey, 2011; Lebherz et al., 2008). In adult rats, RGC death begins 3–4 days after axotomy and is already well advanced 1 week after neurotrauma (Berkelaar et al., 1994; Garcia-Valenzuela et al., 1994; Mansour-Robaey et al., 1994; Nadal-Nicolás et al., 2009), which is during the period of onset of CNTF transgene expression when the rAAV2 vector is injected at the time of injury. Because multiple rCNTF intravitreal injections starting on day 3 post-injury can protect RGCs, an effect further potentiated by co-injection with CPT-cAMP (Cui et al., 2003; Hu et al., 2007; Park et al., 2004), we hypothesized that combining the intravitreal rAAV2-CNTF-GFP injection at the time of injury with short-term pharmacotherapeutic support during the period of vector integration and subsequent therapeutic gene transcription would enhance post-injury RGC survival and axonal regeneration into PN grafts. This was indeed the case; on average almost 39,000 RGCs remained viable at 4 weeks using this paradigm (more than double the sham saline injection value), and ∼27% of these neurons regenerated an axon into a PN graft.
The number of surviving RGCs in the rCNTF plus CPT-cAMP injected group represents approximately one third of the initial RGC population, and is almost double the number of surviving RGCs found in rats injected with rAAV2-CNTF-GFP vector 2 weeks prior to axotomy, using the same PN–ON graft protocol (Hellström et al., 2011). Perhaps surprisingly, however, the number of regenerating axons was similar (means of 10,567 and 10,327, respectively), which we now hypothesize may in fact reflect constraints imposed by the maximal carrying capacity of the tibial nerve branch, a proposal that is currently under investigation.
In all groups, surviving and regenerating RGC densities were significantly greater in temporal retina (Fig. 3), the region where RGC transduction was highest, and corresponding to the vicinity of the initial rAAV2 injection. The average surviving RGC density derived from the transduced area in the two groups receiving additional trophic support was estimated to be 913 RGCs per mm2, which means that almost half of the normal RGC population survived the initial insult in this part of the retina. Localization of transduction following gene therapy with viral vectors is a common limitation after a single intravitreal injection in the adult rat (Hellström and Harvey, 2011). Based on the foregoing theoretical considerations, it is likely that the number of surviving and regenerating RGC numbers would be enhanced still further by giving multiple intravitreal vector injections to achieve effective transduction across the entire retina. This would require sequential injections over several days, because increased intravitreal injection volumes may elevate intraocular pressure which is detrimental to RGCs (Qu et al., 2010; Sampat and Garg 2010). An extended window of therapeutic opportunity may help reduce secondary injury cascades and prove beneficial for clinically relevant ex vivo gene therapy applications such as the use of gene modified cell transplantation protocols after acute CNS trauma (Gögel et al., 2011; Jain, 2009; Kim et al., 2009; Ma et al., 2010).
There is evidence that CNTF can have both direct and indirect effects on injured RGCs, but the relative importance of these effects remains controversial, and perhaps varies depending upon how the cytokine is delivered. Intravitreal rCNTF injections have been reported to increase infiltration of macrophages, which may underlie some of the beneficial effects of this cytokine on RGC regeneration (Cen et al., 2007; Cui et al., 2009; Kurimoto et al. 2010; Yin et al., 2003, 2009). Exogenous CNTF may also act indirectly on RGCs by modulating supportive retinal glia (van Adel et al., 2005) and increasing expression of endogenous cytokines (Müller et al., 2009; Park et al., 2009). Nevertheless, exogenous CNTF does appear to have direct effects on RGC survival and axonal regeneration when applied either as a recombinant protein (Leibinger et al., 2009; Müller et al., 2009) or delivered using rAAV2 (Leaver et al., 2006). Our current and previous observations (Hellström et al., 2011) demonstrate that RGC survival and regeneration is significantly higher in the transduced compared to the non-transduced retinal areas, which emphasizes the localized nature of the rAAV2-related CNTF effect on injured RGCs, and seems to argue against a pan-retinal effect resulting from vector-mediated CNTF release.
After acute neurotrauma or stroke, a gene therapy protocol that allows rapid onset of protection is optimal in order to minimize death of injured neurons, providing greater opportunity for subsequent regenerative repair. Novel, more efficient viral vectors with faster transgene activation are being developed based on different types of viruses (Hermening et al., 2006; Rahim et al., 2009; Thomas et al., 2000; Yanez-Munoz et al., 2006), or on pseudo-serotyped vectors (Hellström et al., 2009; Sweigard et al., 2010; Von Seggern et al., 2003), or vectors that carry the self-complimentary RNA strand (Kong et al., 2010; Natkunarajah et al., 2008; Wang et al., 2003; Yokoi et al., 2007). In the present study, the standard rAAV2 vector was used, first because this serotype is efficient at transducing RGCs (Hellström et al., 2009) and second because of the size of the modified secretable CNTF gene; there are limitations in the packaging capacity of many of the new vector systems that have a faster integration and transgene expression capability (Hellström and Harvey, 2011).
Conclusion
In summary, our new data provide “proof of principle” that rAAV gene therapy can be an effective strategy for the treatment of patients after CNS trauma. rAAV2-CNTF-GFP injection at the time of injury resulted in a reduction in the extent of RGC death following acute ON injury; however, simply by combining the rAAV application with appropriate short-term pharmacotherapeutic support, we obtained a synergistic effect that rescued approximately one third of the original RGC population and induced substantial regeneration of RGC axons. We therefore show for the first time that a major hurdle in the treatment of neurotrauma using current rAAV vector based gene technology, the delay in activation and expression of relevant transgenes, can be overcome using a combined gene therapy and pharmacotherapy approach.
Footnotes
Acknowledgments
This study was funded by the National Health and Medical Research Council (NHMRC) and Western Australia Neurotrauma Research Program. M.H. was supported by an International Postgraduate Research Scholarship and a University of Western Australia Completion Scholarship.
Author Disclosure Statement
No competing financial interests exist.