
Abstract
Manipulation of various components of the endoplasmic reticulum (ER) stress response (ERSR) has led to functional recovery in diabetes, cancer, and several neurodegenerative diseases, indicating its use as a potential therapeutic intervention. One of the downstream pro-apoptotic transcription factors activated by the ERSR is CCAAT enhancer binding protein (C/EBP) homologous protein (CHOP). Recently, we showed significant recovery in hindlimb locomotion function after moderate contusive spinal cord injury (SCI) in mice null for CHOP. However, more than 40% of human SCI are complete. Thus the present study examined the potential therapeutic modulation of CHOP in a more severe SCI injury. Contused wild-type spinal cords showed a rapid activation of PERK, ATF6, and IRE-1, the three arms of the ERSR signaling pathway, specifically at the injury epicenter. Confocal images of phosphorylated EIF2α, GRP78, CHOP, ATF4, and GADD34 localized the activation of the ERSR in neurons and oligodendrocytes at the injury epicenter. To directly determine the role of CHOP, wild-type and CHOP-null mice with severe contusive SCI were analyzed for improvement in hindlimb locomotion. Despite the loss of CHOP, the other effectors in the ERSR pathway were significantly increased beyond that observed previously with moderate injury. Concomitantly, Basso Mouse Scale (BMS) scores and white matter sparing between the wild-type and CHOP-null mice revealed no significant differences. Given the complex pathophysiology of severe SCI, ablation of CHOP alone is not sufficient to rescue functional deficits. These data raise the caution that injury severity may be a key variable in attempting to translate preclinical therapies to clinical practice.
Introduction
T
No single clinical therapy significantly increases functional recovery after SCI in all injury groups. Methylprednisolone and ganglioside GM1 have shown modest efficacy in subsets of human SCI patients, although these data remain controversial (Hawryluk et al., 2008). We recently demonstrated activation of the ERSR in a moderate T9 contusion model of SCI, and significant functional recovery in the hindlimb locomotion of CHOP-null mice. That functional recovery correlated with an increase in white matter sparing and transcript levels of myelin basic protein (MBP) and claudin 11 (Ohri et al., 2011). Similarly, a previous study demonstrated ERSR activation in rats after moderate contusive SCI (Penas et al., 2007). As more than 40% of human SCI are complete and thus defined as severe (
Methods
Animals
All animal procedures were performed in strict accordance with the Public Health Service Policy on Humane Care and Use of Laboratory Animals, Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, National Research Council, 1996), and with the approval of the University of Louisville Institutional Animal Care and Use Committee. CHOP-null mice, completely bred to a 100% C57BL/6 background, were procured from Jackson Laboratories (Bar Harbor, ME). Age- and weight-matched wild-type C57BL/6 female mice (6–8 weeks old) were obtained from Harlan (Indianapolis, IN).
Spinal cord injury
Mice were anesthetized by an IP injection of 0.4 mg/g body weight Avertin (2,2,2-tribromoethanol in 0.02 mL of 1.25% 2-methyl-2-butanol in saline; Sigma-Aldrich, St. Louis, MO). Lacri-Lube ophthalmic ointment (Allergen, Irvine, CA) was used to prevent drying of eyes, and gentamicin (50 mg/kg; Boehringer Ingelheim, St. Joseph, MO) was subcutaneously administered to reduce infection. A laminectomy was done at the T9 vertebra and the mice were placed in a custom stabilizer device, which holds the spinal cord level and steady in the Louisville Injury Systems Apparatus (LISA) as described previously (Beare et al., 2009; Zhang et al., 2008). The LISA uses a laser sensor to measure the velocity and displacement of an injury is obtained by an impactor. Both wild-type and CHOP-null mice received a 0.45-mm-displacement contusion at a velocity of 1.0 m/sec. This injury was predicted to be moderate/severe, but because of new clamps produced a reproducible severe injury as defined by both the Basso Mouse Scale (BMS) and white matter sparing. Injury severity, based on the biophysical parameters of impactor movement, was identical for all animals. Experimental controls included sham animals that remained uncontused and received laminectomy only at the T9 vertebra. Post-surgery, the animals were given 1 cc of sterile saline subcutaneously, 0.1 cc of gentamicin intramuscularly on the day of surgery, and on days 3 and 5 post-surgery, and 0.1 cc buprenorphine subcutaneously on the day of surgery and for the next 2 days. The animals were placed on a heating pad until full recovery from anesthesia. Post-operative care included manual expression of the bladders twice a day for 7–10 days or until spontaneous voiding returned. The animals were sacrificed 6 weeks post-injury.
RNA extraction and reverse transcriptase PCR
Total RNA was extracted from spinal cord tissue of sham animals, injured wild-type (n=4), and CHOP-null mice (n=4) at the injury epicenter (3 mm), and 1 cm away from the injury epicenter for rostral segments (3 mm) using Trizol (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. The RNA was quantified by UV spectroscopy and RNA integrity was confirmed on an ethidium bromide-stained formaldehyde agarose gel. cDNA was synthesized with 1 μg of total RNA using the High Capacity cDNA Synthesis Kit (Applied Biosystems, Foster City, CA) in a 20-μL reaction volume. As controls, mixtures containing all components except the RT were prepared and treated similarly. All cDNAs and control reactions were diluted 10× with water before using as a template for quantitative real time (qRT)-PCR.
Quantitative PCR analysis
qRT-PCR was performed using an ABI 7900HT real-time PCR instrument (Applied Biosystems). Briefly, diluted cDNAs were added to Taqman universal PCR master mix (Applied Biosystems) and run in triplicate. Target and reference gene PCR amplification was performed in separate tubes with Assay on Demand™ primers (Applied Biosystems) as follows: XBP1 (Mm00457359_m1), GRP78 (Mm01333323_g1), CHOP (Mm01135937_g1), ATF4 (Mm00515324_m1), and growth arrest and DNA-damage protein 34 (GADD34; mm00492555_m1). The RNA levels were quantified using the ΔΔCT method. Expression values obtained from triplicate runs of each cDNA sample were normalized to triplicate value for glyceraldehyde-3-phosphate dehydrogenase (GAPDH; reference gene) from the same cDNA preparation. Transcript levels are expressed as fold changes compared with respective levels in sham controls.
XBP1 splicing
XBP1 splicing was performed using HotStarTaq enzyme (Invitrogen) in a final volume of 50 μL containing 2 μL of cDNA and 10 pmol of XBP1 primers (sense: ggccttgtggttga gaaccaggag; antisense, gaatgcccaaaaggatatcagactc) in 1× reaction buffer. Amplification cycles were: 94°C for 2 min followed by 35 cycles at 95°C for 10 sec, 68°C for 30 sec, and 72°C for 30 sec. The final incubation was done at 72°C for 10 min. GAPDH transcript levels were used to normalize against different amounts of input RNA. XBP1 and GAPDH PCR products were resolved on 2.5% and 1% agarose gel electrophoresis, respectively, and detected by ethidium bromide staining.
Immunohistochemical analyses
Mice were anesthetized and transcardially perfused with ice cold PBS. The spinal cords were dissected fresh at different time intervals post-SCI and cryopreserved at −80°C. TBS™ was used as a mounting medium and the cords were sectioned longitudinally at 20 μm and stored at −80°C until further use. For immunostaining, spinal cord sections were post-fixed with ice cold methanol for 10 min and blocked in TBS containing 5% BSA and 0.1% Triton-X-100 for 1 h at room temperature with gentle agitation. Mouse polyclonal anti-NeuN (1:100; Chemicon, Temecula, CA), goat polyclonal anti-Olig2 (1:200; Santa Cruz Biotechnology Inc., Santa Cruz, CA) and mouse monoclonal anti-glial fibrillary acidic protein (GFAP; 1:250; Chemicon) primary antibodies were used for neurons, oligodendrocytes, and astrocytes, respectively. The ER stress primary antibodies used were as follows: Rabbit polyclonal CHOP (1:100; Sigma, Ronkonkoma, NY), rabbit polyclonal ATF4 (1:100; Abcam, Cambridge, MA), rabbit polyclonal peIF2α (1:100; Biosource, Camarillo, CA), rabbit polyclonal GRP78 (1:250; Stressgen, Ann Arbor, MI), and rabbit polyclonal GADD34 (1:100; Santa Cruz Biotechnology). The sections were then incubated with fluorescein- (1:100; FITC) and rhodamine (1:200; TRITC)-conjugated F(ab’)2 secondary antibodies (Jackson ImmunoResearch, West Grove, PA) for 1 h at room temperature. Confocal images were obtained using an Eclipse 90i laser confocal microscope (Nikon, Melville, NY). Negative controls used were species-specific, non-immune IgGs or sera.
Western blot analyses
Protein lysates prepared from 0.4 cm tissue isolated from sham animals and the injury epicenter of contused cords at 6 h and 24 h post-injury in protein lysis solution (20 mm Tris, pH-6.8, 137 mm NaCl, 25 mm B-glycerophosphate, 2 mm NaPPi, 2 mm EDTA, 1 mm Na3VO4, 1% Triton X-100, 10% glycerol, protease inhibitor, 0.5 mm DTT, and 1 mm PMSF) were quantified using a BCA Kit (Pierce Protein Research Products, Rockford, IL). Proteins were separated on SDS-PAGE gels and transferred to nitrocellulose membranes (Whatman, Schleicher & Schuell, London, U.K.). The membranes were processed using standard procedures with the following antibodies: GRP78 (1/1000 dilution; Cell Signaling, Technology, Inc., Beverly, MA), GADD34 (1/500; Santa Cruz Biotechnology), ATF4/CREB-2 (1/500 dilution), phospho-eIf2α (1/1000 dilution; Cell Signaling), eIF2α (1/1000; Biosource International, Wilmington, NC), and GAPDH (1/5000).
Behavioral assessment
Open field BMS locomotor analyses were performed prior to injury for each animal to determine the baseline scores, and weekly following SCI for 6 weeks exactly as defined by Basso and associates (2006). All raters were trained by Dr. Basso and colleagues at the Ohio State University and were blinded to animal groups.
White matter sparing
Six weeks after SCI, wild-type and CHOP-null mice were perfused with 4% paraformaldehyde and spinal cords at the injury epicenter±2 mm were dissected out and stored in 4% paraformaldehyde at 4°C. The next day the cords were cryoprotected in 30% sucrose and kept for at least 3 days at 4°C. TBS™ was used as the mounting medium and the cords were sectioned transversely at 20 μm and stored at −80°C until further use. Eriochrome cyanine (EC) was used to stain myelin to detect the extent of spared white matter at the injury epicenter. The percentage of spared white matter was determined with reference to sham control animals as described previously (Magnuson et al., 2005). Briefly, the slides were allowed to thaw at room temperature for an hour. The sections were then hydrated by submerging in Coppin jars containing xylene and ethanol in the following order: xylene (2×30 min), 100% ethanol (3 min), 95% ethanol (3 min), 70% ethanol (3 min), 50% ethanol (3 min), and distilled water (2 min), followed by staining in EC for 10 min. After dipping the sections in tap water twice to remove excess stain, the sections were differentiated in 0.5% ammonium hydroxide solution for 10 sec, and then underwent another two washes in tap water to stop the differentiation process. The sections were air dried overnight in a biological safely hood at room temperature, mounted the next day using mounting medium, and allowed to dry overnight in the hood. Images were obtained using a Nikon Eclipse Ti inverted microscope, and white matter was traced using Nikon Elements software. The epicenter was defined by the segment with the least amount of spared white matter.
Statistical analyses
For functional assessments after injury, a repeated-measures analysis of variance (ANOVA) with fixed-effects and Bonferroni post-hoc t-test was performed to detect differences in BMS scores and subscores between the sham and injury groups over the 6-week testing period. Statistical analysis of qRT-PCR data was performed using the independent t-test for means with equal or unequal variances or repeated-measures ANOVA (one-way or two-way analysis of variance) followed by post-hoc Tukey HSD test. For all other analyses, the independent t-test for means assuming equal variance was performed.
Results
The ERSR pathway is executed by three ER transmembrane proteins: PERK, ATF6, and IRE-1α. The activation of the PERK arm of ERSR involves phosphorylation of eIF2α at Ser-51, which results in global shutdown of protein translation. Additionally, it also results in the increased expression of ATF4. Analysis of RNA isolated from the injury epicenter and rostral controls of wild-type animals at 1, 6, 12, 24, and 72 h post-severe SCI showed a significant upregulation of the ATF4 transcript levels to 2.5-fold at 6 h, threefold at 12 h, and 2.2-fold at 24 h by qRT-PCR. Upregulation of ATF4 was not observed in the rostral segments of mouse spinal cords, indicating the specificity of the ERSR to the injury epicenter (Fig. 1A). A significant 2.2- to 3.5-fold increase in XBP1 transcript levels at the injury epicenter occurred 6–72 h post-SCI (the latest time point tested), indicating the activation of the ATF6-dependent UPR signaling (Fig. 1B). Interestingly, a significant twofold increase in XBP1 mRNA levels was also observed at 12 h and 72 h in the rostral control segments of the lesioned cord, indicating a spread of the pathological process from the site of primary impact into the adjacent tissue. The presence of both the spliced (226-bp) and unspliced (254-bp) XBP1 transcripts at 6, 12, 24, and 72 h post-SCI compared to shams (laminectomy controls) by semi-quantitative PCR indicated the activation of the IRE-1α-dependent pathway of the ERSR (Yoshida et al., 2001; Fig. 1C). Collectively, these data demonstrate that severe injury to the spinal cord leads to a robust and early activation of all three arms of the ERSR.
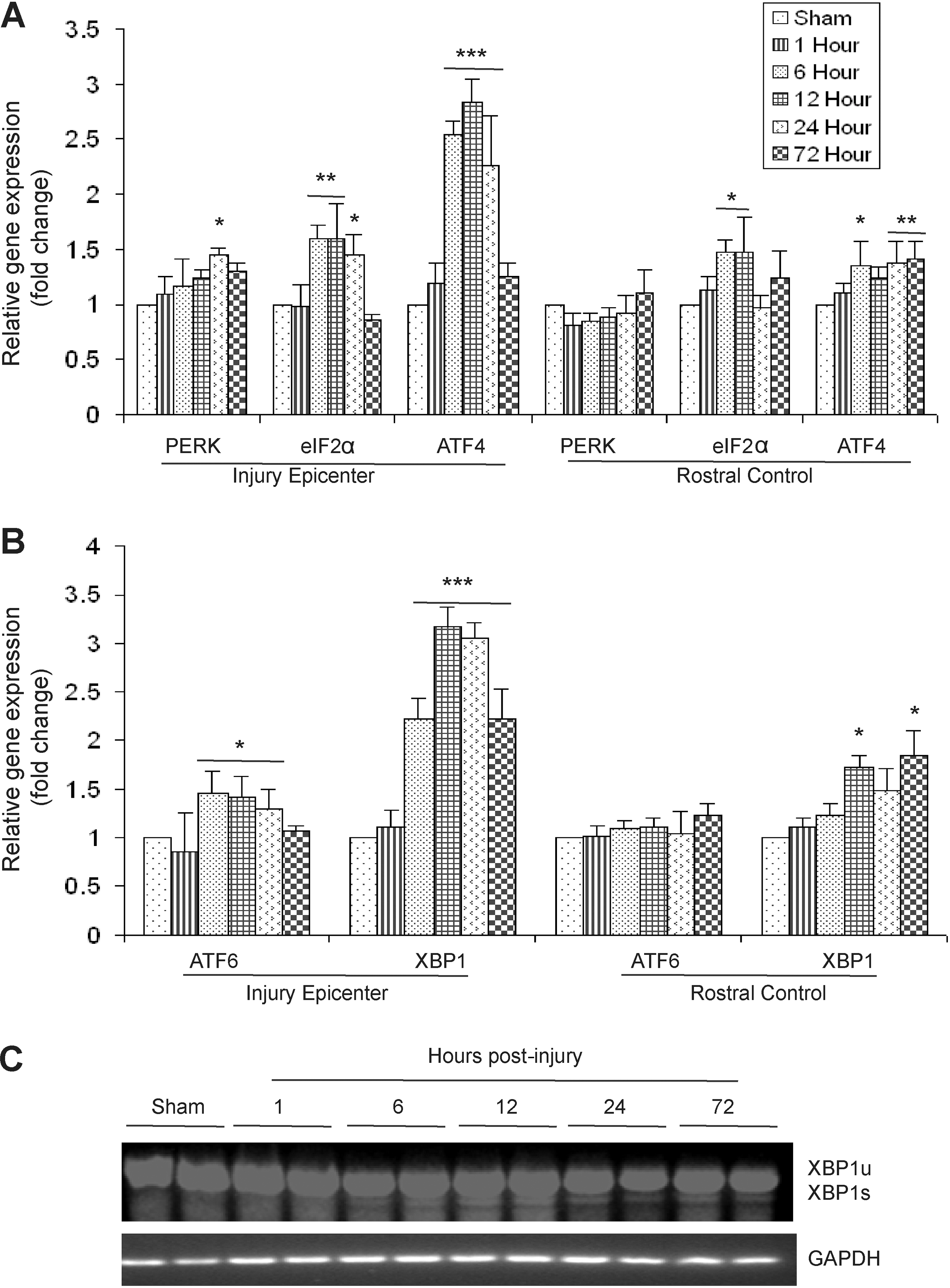
Activation of the unfolded protein response (UPR) in T9 contused spinal cords of wild-type animals after severe spinal cord injury (SCI). (
To gain further insight into the complex transcriptional response of downstream targets of the ERSR/UPR in response to severe SCI, we determined the expression level of CHOP, glucose-regulated protein 78 (GRP78) and GADD34. qRT-PCR data demonstrated a significant 2.5-fold induction of CHOP mRNA as early as 6 h post-injury specifically at the injury epicenter. This induction persisted at least until 72 h (the latest time point examined). In addition, a significant increase in CHOP transcript levels was also apparent in rostral segments at 24 and 72 h post-injury, suggesting the spread of the UPR to adjacent tissue. This increase was not observed after moderate contusive SCI (Ohri et al., 2011). A significant 2.5- to 2.8-fold induction of GRP78 mRNA was observed at 12 h post-severe SCI, which increased to 3.2-fold at 24 h, and then decreased to 2.5- to 2.8-fold at 72 h. In contrast, the rostral segments did not show increased expression of GRP78 mRNA except at 72 h. The expression level of GADD34 mRNA increased to 3.5-fold as early as 1 h post-injury, persisted until 12 h, and then declined to basal levels by 72 h. Upregulation of GADD34 transcript levels was specific to the injury epicenter, as the rostral segments did not show any significant upregulation at any of the time points tested (Fig. 2A). To determine changes in the ERSR protein levels, protein lysates extracted from the epicenter of sham and contused animals after 6 and 24 h post-injury were analyzed on an immunoblot (Fig. 2B). An increase in phosphorylated EIF2α levels was evident at 6 h post-injury, which further increased at 24 h, indicating sustained translation attenuation. Levels of GRP78 were highest at 24 h post-injury, correlating with the qRT-PCR data. Increased levels of ATF4 were observed at 6 and 24 h post-injury. GADD34 protein levels were highest at 6 h post-injury. Collectively, these data indicate that the levels of various ERSR transcripts translate into corresponding alterations in protein levels.
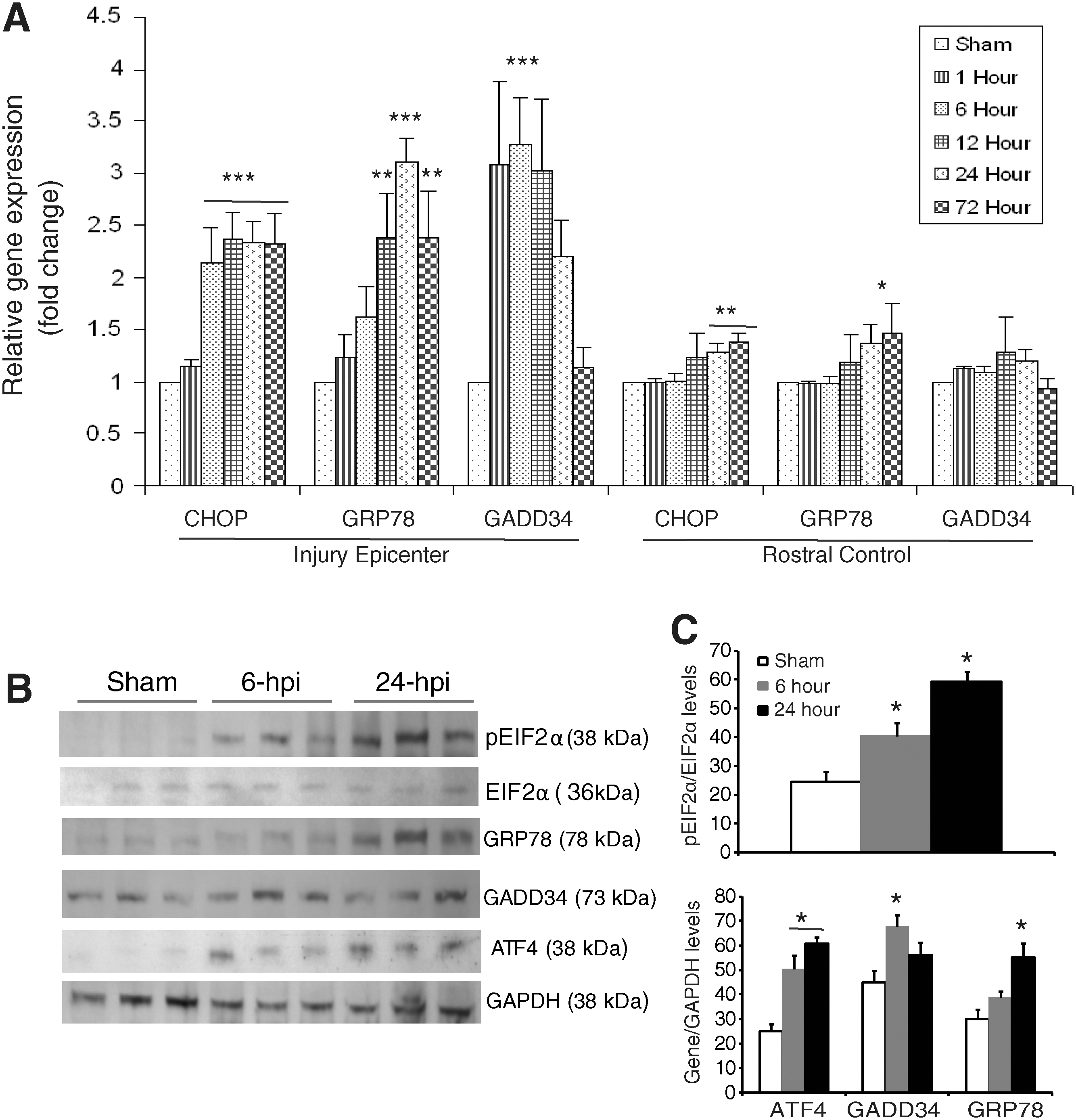
Expression of the unfolded protein response (UPR) response genes in wild-type animals after severe spinal cord injury (SCI). (
Immunohistochemical analyses showed the various key components of the UPR pathway to be co-localized in both neurons and oligodendrocytes. The accumulation of pEIF2α was apparent in both neurons (Fig. 3A) and oligodendrocytes (Fig. 4A) at 6 h post-severe SCI. The expression of GRP78, ATF4, and CHOP was distinctly seen as early as 6 h post-severe SCI in neurons (Fig. 3B–D), and their oligodendrocytic expression was predominantly apparent at 24 h (Fig. 4B–D). In contrast, the non-injury controls did not show any detectable expression of these markers in either oligodendrocytes or neurons (data not shown). Taken together, these results indicate that traumatic severe injury to the spinal cord led to the increased expression of various ER stress response genes temporally in neurons and oligodendrocytes.
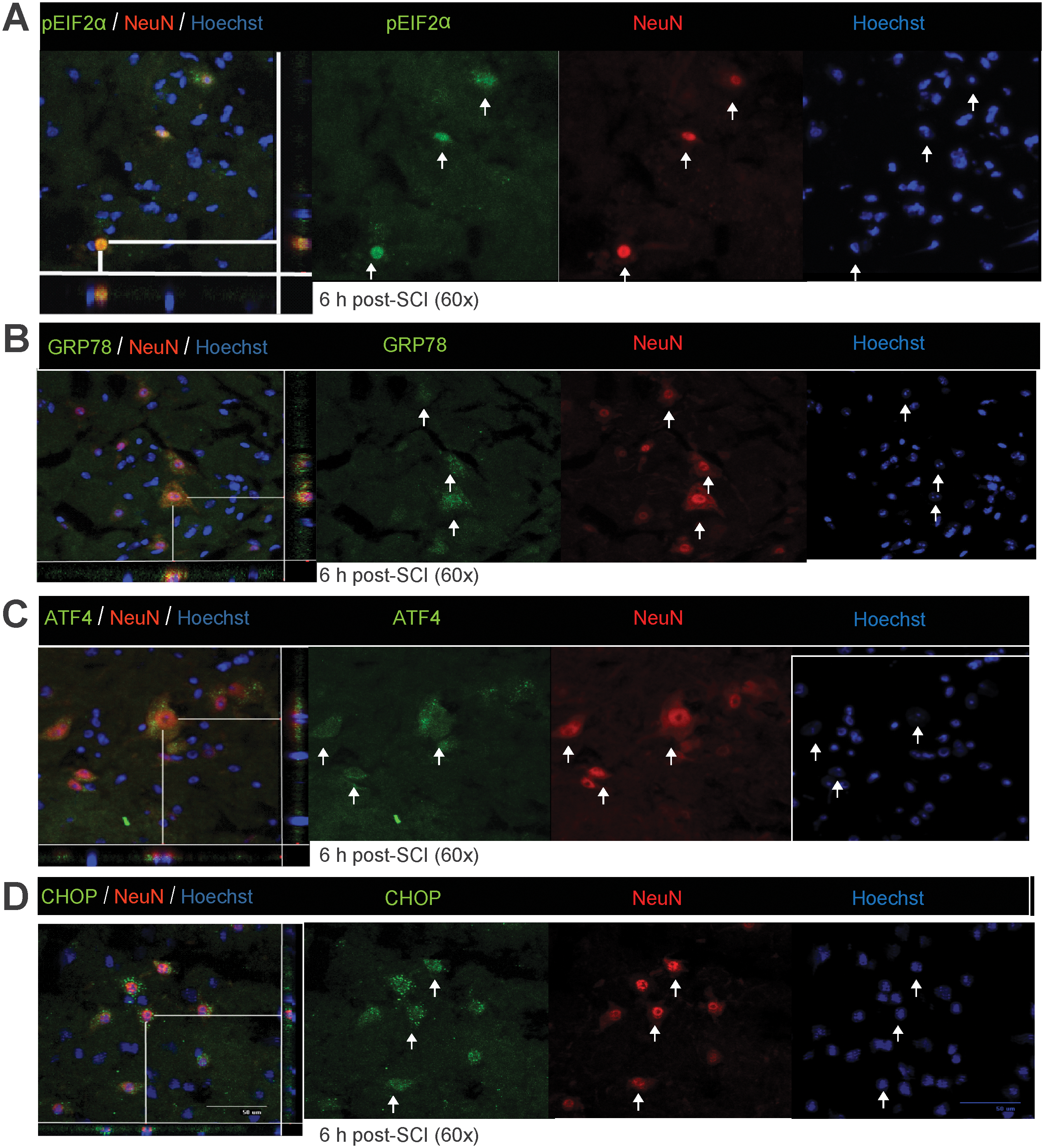
Expression of unfolded protein response (UPR) markers in neurons of wild-type animals after severe spinal cord injury (SCI). Immunohistochemical analyses of longitudinal sections reveal co-localization of pEIF2α (
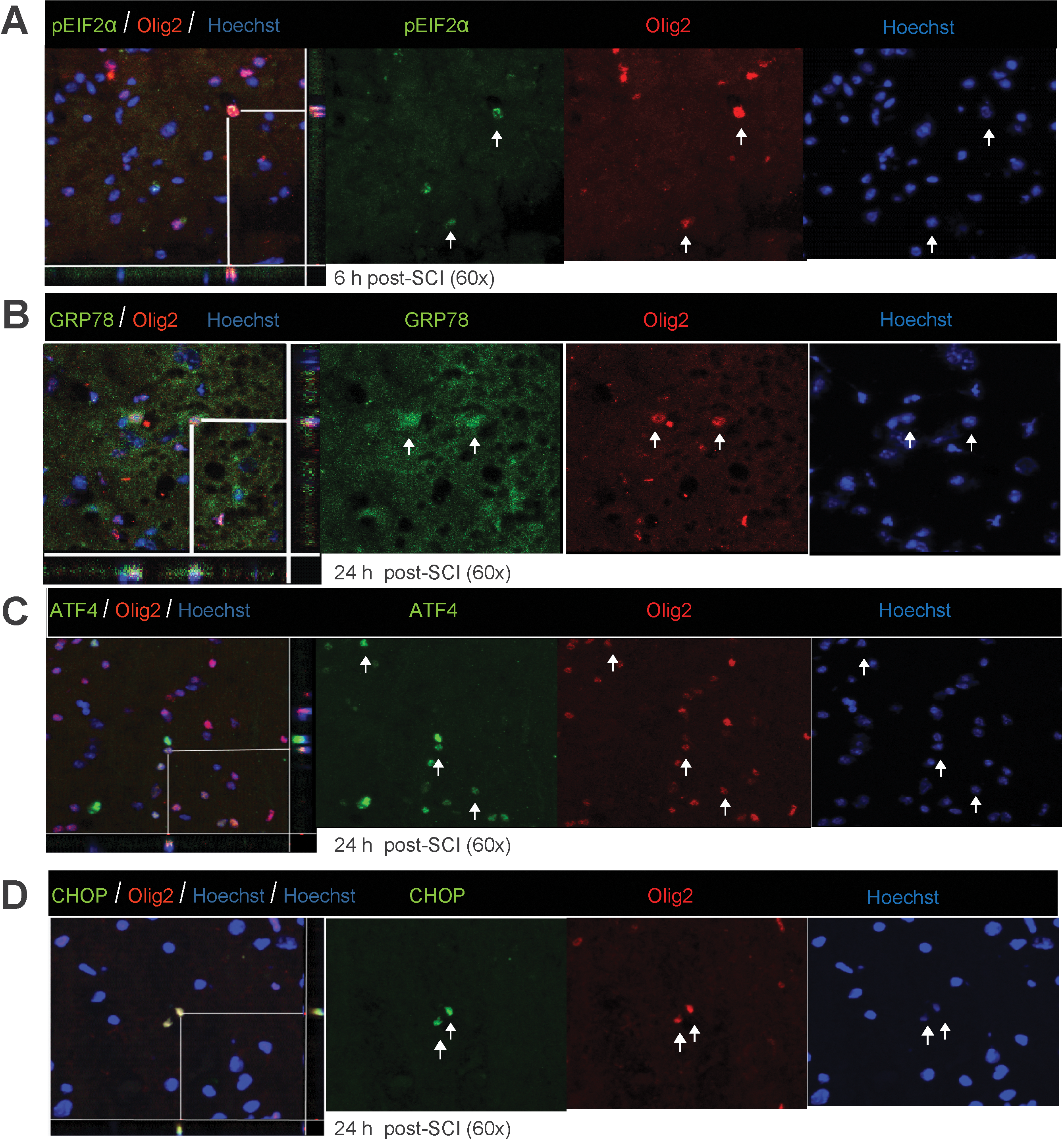
Oligodendrocytic expression of unfolded protein response (UPR) markers in wild-type animals after severe spinal cord injury (SCI). Immunohistochemical analyses of longitudinal sections reveal co-localization of pEIF2α (
Deletion of CHOP does not result in functional recovery in mice with severe SCI
Considering the significant upregulation of CHOP mRNA in contused spinal cords specifically at the injury epicenter, we decided to utilize CHOP-null mice to evaluate its potential role in functional consequences of severe SCI. We first determined the UPR response in the CHOP-null mice 72 h post-severe injury. The relative gene expression values of ATF4, GADD34, GRP78, and XBP1 in wild-type and CHOP-null animals remained similar, suggesting identical UPR responses in both groups of animals to severe injury (Fig. 5A). A lack of CHOP expression in CHOP-null mice after severe injury as assessed by immunohistochemistry and Affymetrix gene expression (data not shown) confirmed that CHOP is truly knocked out. Analysis of BMS scores comparing wild-type and CHOP-null animals displayed scores of 3.17±0.25 and 2.86±0.38, respectively, at week 1, and 3.06±0.16 and 3.14±0.38, respectively, at week 6 (Fig. 5B). A BMS score of 3.0 correlates with a severe injury and is in accordance with an earlier study (Beare et al., 2009). CHOP-null mice did not show any enhanced functional recovery compared to wild-type animals at any of the time points tested. At 6 weeks post-injury, histological analysis showed no significant difference in the percentage of spared white matter in wild-type (27.46±4.6%) and CHOP-null (30±5.2%) mice (Fig. 5C). These data indicate that, unlike moderate contusive SCI, deletion of CHOP is not sufficient to enhance functional recovery after severe injury.

Deletion of CCAAT enhancer binding protein homologous protein (CHOP) does not result in functional recovery after severe spinal cord injury (SCI). (
Discussion
Clinically, more than 40% of human SCI are complete (
CHOP, also known as growth arrest- and DNA damage-inducible gene 153 (GADD153), is a member of the C/EBP family of bZIP transcription factors, and operates as a downstream component of the PERK-ATF4 arm of the ER-stress pathway. Overexpression of CHOP led to cell cycle arrest and apoptosis (Matsumoto, 1996; Oyadomari et al., 2001). In contrast, CHOP-null mice exhibited reduced apoptosis in response to ER stress (Oyadomari, 2002; Zinszner et al., 1998), showed smaller infarcts than wild-type animals subjected to bilateral carotid artery occlusion (Kohno et al., 1997), and displayed less tissue loss after stroke injury (Tajiri et al., 2004). In addition, we recently showed that CHOP-null mice displayed enhanced functional recovery compared to wild-type mice after moderate contusive SCI, an effect suggested to reflect oligodendrocyte protection (Ohri et al., 2011).
The fact that the current study showed no functional differences between the wild-type and CHOP-null animals after severe SCI could be explained by the complex transcriptional and translational interplay of various components of the ERSR pathway and the presence of other tissue-damaging pathways. For example, CHOP-null mice were not resistant to lethal doses of tunicamycin, an ER stress-causing agent (Zinszner et al., 1998), suggesting that other pro-apoptotic pathways are also active. ER stress-mediated cytotoxicity is regulated by at least three independent signaling cascades: transcriptional activation of CHOP, activation of the cJUN NH2-terminal kinase (JNK) pathway through the ASK1 complex (Nishitoh et al., 2002; Urano et al., 2000), and activation of caspase 12 (Nakagawa et al., 2000). In addition, recent studies have implicated Bax and Bak as executioners for ER stress-mediated apoptosis (Wei et al., 2001; Zong et al., 2001). ER stress can also activate the Bcl-2 (Annis et al., 2004; Thomenius and Distelhorst, 2003) and caspase families of proteins (Boyce and Yuan, 2006). ER stress itself can upregulate or activate several BH3-only pro-apoptotic members of the Bcl-2 family, including Bim (Morishima et al., 2004), BIK (Germain et al., 2005; Mathai et al., 2005), and PUMA (Luo et al., 2005). Therefore, efferent signaling from the stressed ER can engage diverse cytotoxic pathways directly, resulting in massive cell death. Compounding this, each of the distinct insults (inflammation, hypoxia, oxidative stress, and trauma) which underlie the complex pathophysiology of SCI (Baptiste et al., 2009; Hall and Springer, 2005) has the potential to induce the ERSR that may possibly result in severe protein misfolding and lead to autophagy. Consistent with this suggestion, severe protein misfolding results in insoluble protein aggregates in the cells that cannot be removed by the ubiquitin/proteaome system and are eliminated by autophagy (Bernales et al., 2006). To identify yet other potential pathways, microarray analyses comparing wild-type and CHOP-null mice demonstrated significant threefold enrichment of the p53 pathway, chemokine signaling pathway, Fc gamma R-mediated phagocytosis, ErbB signaling pathway, Fc epsilon Ri signaling pathway, and B-cell receptor signaling pathway (data not shown), suggesting additional cell-death and inflammatory pathways to be upregulated/enriched in the CHOP-null mice after severe injury.
The clinical significance of the current data is twofold. With increasing injury severity, both the magnitude and duration of the ERSR were increased, and that increase likely activated multiple cytotoxic mechanisms besides CHOP. Identifying additional key effectors downstream of the ER-mediated cytotoxicity pathways and ER-independent pathways may provide a basis for new therapeutic approaches for treating SCI. However, a parallel cautionary interpretation would be that with very severe SCI (e.g., clinically complete), pharmacological interventions to reduce secondary cell death may be ineffective both due to the extent of initial tissue destruction, as well as excessive activation of cell death pathways.
Acknowledgments
The work presented in this manuscript was supported by RR15576, The Kentucky Spinal Cord and Head Injury Research Trust, Norton Healthcare, the Commonwealth of Kentucky Challenge for Excellence, and NS073584 (to M.H. and S.R.W.), NS047341 (to M.H.), and NS054708 (to S.R.W.). We thank Christine Nunn for help with surgical procedures, Kimberly Fentress for assistance with post-operative animal care, and Darlene A. Burke for help with statistical analyses.
Footnotes
Author Disclosure Statement
No competing financial interests exist.