
Abstract
Environmental enrichment (EE) is a way to induce voluntary locomotor training that positively affects locomotor recovery after acute spinal cord injury (SCI). The beneficial effect on SCI outcome is thought to be based on enhanced plasticity in motor pathways, triggered by locomotor-specific sensory feedback to the spinal cord circuitry for locomotion (central pattern generators [CPGs]). In view of chronic SCI, we tested the hypothesis that EE improves motor outcome after SCI in the rat when started after a clinically relevant delay of 3 weeks. At the CPG level (i.e., the spinal L1–L2 level), where EE-related sensory feedback is processed, two key mechanisms of anatomical plasticity were examined: (1) serotonergic innervation, and (2) survival and differentiation of spinal cord progenitor cells. Delayed EE improved interlimb coordination, which was associated with an increased serotonergic innervation of the ventro-lateral grey matter within the L1–L2 segments. Although spinal cord progenitor cells were found to differentiate into both neurons and glial cells, EE did not affect their survival. These results show that EE induces a substantial improvement of motor outcome after SCI when commenced after a clinically-relevant delay. Increased serotonergic innervation of the lumbar CPG area is therefore suggested to play an important role in the EE-induced recovery of interlimb coordination.
Introduction
O
Rehabilitation strategies have been proposed to affect SCI outcome by augmentation of the remarkable plasticity that is present within adult spinal cord networks (Girgis et al., 2007). Synaptic plasticity in pre-existing pathways and the formation of new circuits through collateral sprouting of lesioned and unlesioned fiber tracts indeed occur spontaneously following incomplete SCI. This spontaneous plasticity is considered imperative to the process of spontaneous locomotor recovery following incomplete SCI, which is observed in both patients and animal models (Bareyre et al., 2004; Fouad et al., 2001; Raineteau and Schwab, 2001; Tillakaratne et al., 2002). In particular, plasticity in descending serotonergic fibers is important because of the role of serotonin in the generation of coordinated locomotion. Indeed, serotonin can elicit locomotion via activation of spinal networks (central pattern generators [CPGs]) that coordinate rhythmic stepping movements (Courtine et al., 2009; Liu and Jordan, 2005; MacLean et al., 1998; Madriaga et al., 2004; Mori et al., 1998).
An as yet unexplored mechanism of EE-induced plasticity involves changes in the proliferation, survival, and differentiation of endogenous progenitor cells in the CPG. Indeed, changes in endogenous progenitor cells can be triggered by both SCI and physical activity (Cizkova et al., 2009). To date, the fate of proliferating and surviving endogenous progenitor cells was only investigated close to a spinal injury site and was found to be glial-restricted. Although induction of neuronal fates has been described to occur in the intact spinal cord during the early postnatal period (Kehl et al., 1997), and in adulthood (Shechter et al., 2007), the injured spinal cord has thus far not been found to support neuronal differentiation.
In the present study, we hypothesize that EE, commenced at 3 weeks after T9 spinal cord contusion injury in the rat, provides locomotor-specific sensory input to the lumbar CPG. This afferent input stimulates adaptive plasticity within this spinal locomotor network, which eventually results in enhanced locomotor abilities. We further aimed at investigating whether: (1) the serotonergic innervation, and (2) the survival and differentiation of endogenous progenitor cells, both within the grey matter of lumbar segments L1–L2, are involved in putative EE-induced functional recovery.
Methods
Animal care
Experiments were performed in accordance with the recommendations of the European Commission (European Communities Council Directive of 24 November 1986; 86/609/EEC); all animal protocols were approved by the Maastricht University Institutional Animal Use Committee. Adult male Wistar rats (U:WU:CPB, 11 weeks of age, 240–275 grams) were used for this study (Charles River, Someren, The Netherlands). The animals were housed under a 12:12-h dark:light cycle in a pathogen-free area with free access to water and food.
Spinal cord contusion injury and bromodeoxyuridine injections
The rats were deeply anesthetized by artificial ventilation using an open mask system to provide halothane in a mixture of O2 and N2O (1:2; 5% and 1.5% halothane for the induction and maintenance of the anesthesia, respectively). Moderate SCI was inflicted (n=22 rats) at spinal segment T10 using the New York University impact injury device (Gruner, 1992). A 10-g, 2.0-mm-diameter rod was released from a 12.5-mm height onto the exposed dorsal surface of the T10 spinal cord. Sham animals (n=3), serving as controls, were subjected to laminectomy only. After SCI or sham surgery, muscles and skin were closed with 4-0 silk sutures, and 5 mL of Ringer's lactate was injected subcutaneously. The animals were returned to their home cages for recovery. Bladders were emptied manually twice a day until reflex bladder voiding returned, typically by the end of the first week after surgery. After surgery, the animals were kept under the same preoperative conditions and fed ad libitum. At 3 weeks after surgery, animals received three subcutaneous injections with bromodeoxyuridine (BrdU; 100 mg/kg in PBS; Sigma-Aldrich, St. Louis, MO), with an inter-injection interval of 6 h. At 11 weeks after surgery, the animals were deeply anesthetized using pentobarbital (180 mg/kg body weight IP), and perfused intracardially with 4% paraformaldehyde in 0.1 M phosphate buffer. The L1–L4 spinal cord was anatomically localized, removed, post-fixed overnight in the same fixative, and then cryoprotected by transfer to 25% sucrose in 0.1 M phosphate buffer.
Housing
The animals were housed in standard type 3 macrolon cages during the pre-operative period (social housing; 2 rats per cage), and for the first 3 weeks after surgery (individual housing). From 3 weeks after surgery onwards, the animals were housed either in an EE (n=11; all animals in the same EE cage; Lankhorst et al., 2001), or in standard type 3 macrolon cages (n=11; individual housing). The EE consisted of a trespa cage with a Perspex front (L×W×H=2.00×1.00×0.80 m) and contained various objects (e.g., running wheels, a shelter house, climbing frames, tubes, and additional nesting material).
Locomotor testing
All behavioral testing was performed by blinded observers. The Basso-Beattie-Bresnahan (BBB) rating scale was used as previously reported (Basso et al., 1995). Moreover, a 7-point BBB subscore was measured (Lankhorst et al., 2001), which has been developed to improve the sensitivity of the BBB score by scoring some of the behavioral attributes (predominant paw position, instability, and tail position) independently (Basso, 2004). For CatWalk gait analysis, the rats were habituated and pre-trained (in the week prior to surgery) as previously described (Deumens et al., 2007). Briefly, the rats were trained to make consecutive uninterrupted runs on a glass runway, during which the contact positions of the paws with the internally-illuminated runway were assessed. Walkway crossing was stimulated by rewarding the animals after each run with small food pellets (Noyes Precision pellets PJPPP-0045; Sandown Chemical Limited, Esher, Surrey, U.K.). The CatWalk system allows for the objective assessment of numerous parameters involved in overground locomotion, including coordination-related parameters and paw-specific parameters (Koopmans et al., 2007). For reliable assessment of locomotor performance, three uninterrupted runs per animal per time point were analyzed. In the present study we used the CatWalk to assess interlimb coordination objectively and thoroughly. This is of the utmost importance since coordination is suggested as a milestone in the recovery process (Basso, 2004), and its assessment using the standard BBB rating scale is limited and has shortcomings, because it focuses on a single aspect of coordination, the one-to-one relationship between forelimbs and hindlimbs, and it is highly subjective (Koopmans et al., 2005). Crossings of the walkway with a regularity index (RI) of 100% were considered to be coordinated. The RI grades the degree of coordination as the total number of normal step sequence patterns in relation to the total number of paw placements (Hamers et al., 2001). CatWalk-based coordination scores were furthermore implemented in so-called CatWalk-based BBB scores, as previously described (Koopmans et al., 2005), and used in several investigations (Koopmans et al., 2009; van Neerven et al., 2010). The CatWalk provides another important coordination parameter: the phase lag (Hamers et al., 2006; Koopmans et al., 2006). Phase lag quantifies interlimb coordination by measuring timing relationships between paw placements; the time of initial contact (IC) of one of the paws (the target) is related (expressed as a percentage) to the stride cycle of another paw (the anchor; for details see Hamers et al., 2006; Koopmans et al., 2006). Phase lag can be calculated (1) between limbs on the same girdle (girdle pair), (2) between limbs on the same side (lateral pair), and (3) between the opposite front- and hindlimb (diagonal pair). In the present study, we focused on the variability of the phase lags in girdle and lateral pairs, as well as on the associated number of mismatches in phase lags (i.e., the number of steps that do not fit the constraints set for computation of the phase lag).
Tissue processing and immunohistochemistry
Spinal cords (n=10 each for the EE and normal housing groups; n=3 for sham control group) were embedded in Tissue-Tek® (Sakura Finetek Europe, Zoeterwoude, The Netherlands) and serially cut in 30-μm-thick transverse sections using a cryostat. For 5-HT staining, every 10th section was rinsed in 0.3% Triton X-100 Tris-buffered saline (TBS-T), TBS, and TBS-T (each step 15 min), followed by incubation with anti-serotonin serum ser 1–16 (1:60,000 in TBS-T; Steinbusch, 1981) overnight. After primary antibody incubation, the sections were rinsed and then incubated in secondary antibody (biotinylated donkey anti-rabbit, 1:800 in TBS-T; Jackson ImmunoResearch Laboratories, West Grove, PA) for 90 min, followed by the Vector ABC procedure (Vector Laboratories, Peterborough, U.K.) for peroxidase staining and visualization with 3,3′-diaminobenzidine (DAB). All steps were performed at room temperature. After rinsing in TBS, the sections were mounted on gelatinized slides, dehydrated through alcohols, cover-slipped, and coded. For BrdU staining, a free-floating staining protocol was used. The sections were rinsed in TBS, incubated in 50% formamide at 65°C for 2 h, rinsed in 2×SSC for 15 min, incubated in 2 N HCl at 37°C for 30 min, and rinsed in 0.1 M boric acid for 10 min (pH 8.5). The sections were then rinsed three times for 15 min each in TBS, and non-specific antigen binding was blocked in a solution containing TBS, 3% normal donkey serum, and 0.25% Triton X-100 (TBS++) for 1 h. Subsequently, the sections were incubated in rat anti-BrdU (1:1000 in TBS++; Accurate Chemicals, Westbury, NY) overnight at 4°C. The next day, the sections were rinsed in TBS and incubated in secondary antibody biotinylated donkey anti-rat (1:400 in TBS++; Jackson ImmunoResearch Laboratories) for 2 h followed by the Vector ABC procedure (Vector Laboratories) for peroxidase staining and visualization with DAB. After rinsing in TBS, the sections were mounted on gelatinized slides, dehydrated through alcohols, cover-slipped, and coded.
To determine the fate of the BrdU-immunoreactive (IR) cells, double-fluorescence immunolabeling was performed, combining BrdU labeling with the cell-specific phenotypic markers glial fibrillary acid protein (GFAP for astrocytes) and neuron-specific nuclear protein (NeuN for neurons). First, the sections were pretreated as described above and BrdU was visualized by incubation in Alexa 594-conjugated streptavidin (1:2000 in TBS; Molecular Probes, Eugene, OR) for 90 min. Subsequently, the sections were rinsed in TBS, blocked by TBS++ for 1 h, and incubated with primary antibody in TBS++ at 4°C overnight: mouse anti-NeuN monoclonal antibody (1:50; Chemicon, Temecula, CA), and mouse anti-GFAP polyclonal antibody (1:1600; Sigma-Aldrich). The cell-specific phenotypic markers were revealed by incubation in Alexa 488-conjugated donkey anti-mouse IgG (1:100 in TBS; Molecular Probes) for 2 h at room temperature.
Quantitative analysis of immunostainings
In order to quantify the 5-HT innervation in particular locations of the L1–L2 ventral grey matter (i.e., medial versus lateral portion), photomicrographs were obtained using an Olympus AX-70 (Olympus, The Netherlands) microscope equipped with a 10× objective and a 10× projection lens. Photomicrographs were taken within the ventro-medial and ventro-lateral grey matter (see Fig. 3) at 10× magnification with a Sony Power HAD 3 CCD color video camera. In total, 24 photomicrographs per animal (EE, n=5; N, n=5; sham, n=3) were analyzed (1 photomicrograph×2 areas/ventral horn×2 ventral horns×6 sections; see Fig. 3B and 3C). 5-HT innervation was detected by image analysis using AnalySIS-pro software (Soft Imaging System, Münster, Germany). First, in every image the region of interest was defined (see Fig. 3C). Second, shading correction was performed before measurement to correct for irregularities in illumination in the microscopic field. Third, a differential contrast enhancement filter was applied to selectively enhance weak differences in contrast; the resulting image is more detailed and appears more focused. Finally, background levels were equalized using a defined threshold setting that represents the grey value range in which the area fraction of 5-HT-IR structures was calculated. All settings were kept identical for all analyses.
Quantitative analysis of BrdU-labeled cells
In 13 rats (n=3 sham, n=5 normal-housed, and n=5 EE-housed animals) estimates of the total numbers of BrdU-IR cells in the grey matter of the L1–L2 segments were carried out by means of stereological methods (Gundersen, 1986), and a stereological workstation consisting of an Olympus BX50 microscope, Olympus UPlan objectives (10× and 40×, oil), and an electronic microcator (Heidenhain, Traunreut, Germany), with StereoInvestigator software (MicroBrightField, Williston, VT). The counting of BrdU-IR cells was performed using five sections spaced equally apart over a 1.6-mm block spanning the L1–L2 segments. Within these sections the dorsal and ventral horn were delineated as shown in Figure 4D, both regions were completely sampled to count at least 200 cells per spinal cord region per animal. Cells were considered BrdU-IR only if the staining was clearly present within the nucleus with either a homogenous or clear punctuate labeling pattern. The actual section thickness after histological preparations was measured with the electronic microcator and total volumes spanning the sampled regions were calculated by means of the Cavalieri principle. Estimates of BrdU-IR cell densities were calculated by dividing the total number of BrdU-IR cells by the total volume of the corresponding region.
Quantification of double labeling for BrdU and phenotypic cell markers
Cells double-labeled for BrdU and the mature astroglial marker GFAP or the mature neuronal marker NeuN were quantified using a confocal laser-scanning microscope (TCS SP2; Leica Microsystems, Wetzlar, Germany). For imaging of Alexa-594 fluorescence a helium/neon laser (excitation wavelength 543 nm, detection range 555–700 nm) was used. For imaging of Alexa-488 fluorescence an argon laser (excitation wavelength 488 nm, detection range 458–514 nm) was used. To determine the proportion of BrdU-labeled cells expressing phenotype-specific markers, BrdU+ cells were counted in each of the quantified regions (ependymal cell layer, ventral horn, and dorsal horn; delineation as shown in Fig. 4D, except the for the ependymal cell layer, which is counted individually) in each subject. The specificity of each label was first verified using single-channel scans at 400× magnification, which were subsequently merged into multiple-channel views. To confirm double-labeling confocal Z-stacks were generated. In addition, we analyzed individual cells using two-photon laser scanning microscopy (TPLSM; Denk et al., 1990), which is a fluorescence microscopy method suitable for imaging structures located deep in scattering tissues (Centonze and White, 1998). TPLSM was performed as described previously (van Zandvoort et al., 2004) using a microscope objective (40× or 60×, oil-immersion or water dipping) connected to an upright Nikon E600FN microscope (Nikon Corporation, Tokyo, Japan). When needed, further magnification was achieved by optical zoom in the scan head, and the Kalman filtering procedure was applied to remove noise. As with confocal scanning, the fluorescent secondary antibody Alexa-488 was mainly visible in the green channel, whereas Alexa-594 was only visible in the red channel. Single-channel images were obtained and subsequently merged into multiple-channel images for the confirmation of double-labeling of Z-stacks. The results for each marker are presented as the proportion of all BrdU+ cells per region that are double-labeled for BrdU and NeuN or BrdU and GFAP.
Imaging
The photomicrographs shown in Figure 7 were generated using the Virtual Slice tool of StereoInvestigator software at a stereological workstation. This tool automatically creates seamless image montages composed of approximately 100 individual fields-of-view at high resolution. The photomicrographs shown in Figure 6 were taken with the Olympus AX-70 microscope equipped with a Sony Power HAD 3 CCD Color video camera. The photomicrographs shown in Figures 5, 6, and 7 were acquired using TPLSM. Image stacks were reconstructed into three-dimensional images followed by colocalization analysis with Imaris 4.0 software (Bitplane, Zurich, Switzerland). No additional image processing was performed.
Statistical analysis
All data were analyzed using SPSS statistical software. Data presented for the BBB scores (i.e., standard BBB score and CatWalk-based BBB score) were analyzed using an analysis of variance (ANOVA) (GLM) with between-subjects factor treatment and within-subjects factor time, with Time as a repeated-measures factor. At individual time points a Student's t-test was used. Student's t-test was also used for the statistical analysis of the RI, phase lag mismatch percent, phase lag variability, and the stride length. BBB subscore was evaluated by non-parametric statistics (Mann-Whitney U test) because the data were ordinal in nature. All immunohistochemical data were also analyzed using the non-parametric tests Kruskall-Wallis and Mann-Whitney U. Group differences were considered statistically significant if p<0.05.
Results
Animal inclusion
In the present study, 3 out of 22 injured animals did not fulfil the inclusion criteria at day post-operative (DPO)-21 (i.e., BBB score of 11 or 12); 1 animal scored 10 points and 2 animals scored 13 points on the BBB scale. In addition, there were two animals that failed to perform the CatWalk procedure. One normal-housed animal died at DPO-49, presumably because of bladder or kidney failure. Thus 16 rats were evaluated behaviorally: 8 EE-housed rats and 8 normal-housed rats.
General locomotor performance
All animals recovered rapidly during the first 3 weeks post-injury and reached BBB scores of 11 or 12 at DPO-21; thereafter the animals were randomly divided into two groups, the EE-housed group and the normal-housed group. During the 2-month period of treatment there was no significant effect of the EE (treatment: F1,14=0.049, n.s.). Thus there was no significant improvement in general locomotion according to the uncorrected BBB score in the EE-housed animals (Fig. 1A).
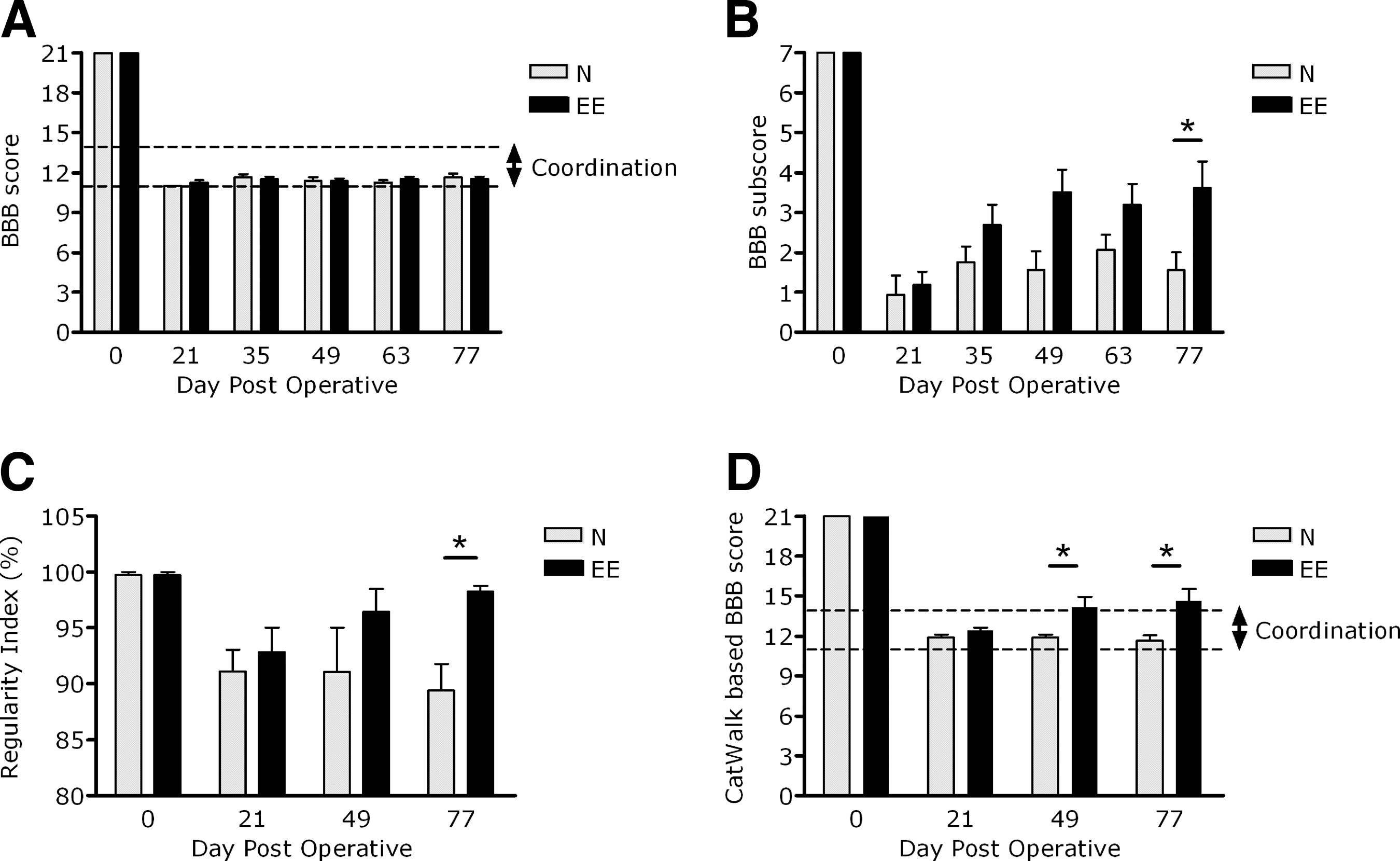
General locomotor abilities as assessed by the Basso-Beattie-Bresnahan (BBB) score and CatWalk score. (
The CatWalk-based BBB score (Koopmans et al., 2005), which integrates the CatWalk parameter RI as an objective measure of coordination into the BBB locomotor scale, changed the general locomotor outcome considerably (Fig. 1D). At the start of treatment (DPO-21) the performance of both groups was comparable (environmental enrichment [EE] 12.4±0.3 and normal [N] 11.9±0.3; mean±standard error of the mean [SEM]; n.s.). From DPO-21 onward the CatWalk-based BBB score showed significant differences between the two housing conditions (treatment: F1,14=8.01, p<0.05; Fig. 1D). After 4 weeks the effect of EE housing became apparent (DPO-49: EE 14.2±0.8 and N 11.9±0.3, mean±SEM; p<0.05). These differences remained until the end of the experiment (DPO-77: EE 14.1±0.7 and N 11.6±0.4, mean±SEM; p<0.05; Fig. 1D). Thus EE housing significantly improves general locomotor performance according to the more sensitive CatWalk-based BBB score.
EE housing ameliorates fine locomotor performance
The BBB subscore (Lankhorst et al., 2001) showed a significant treatment effect at the end of the experiment (DPO-77: p<0.05 by Mann-Whitney U test). Thus EE-housed animals showed significantly better recovery of fine locomotor performance when compared to normal-housed animals as assessed by the BBB subscore (Fig. 1B).
EE housing improves interlimb coordination
Since the recovery of coordination plays a pivotal role in the recovery of locomotor performance after experimental spinal cord contusion, we included the CatWalk system to focus on the recovery of coordination. The following coordination-related parameters revealed interesting results.
Regularity index (RI)
The mean RI at baseline was high (DPO-0: 99.88±0.12, mean±SEM; Fig. 1C), and almost all (47 out of 48) individual CatWalk runs reached an RI of 100%. Three weeks after injury, at the beginning of the treatment period, the mean RI was decreased (DPO-21: EE 93.9±2.2 and N 91.6±2.7; mean±SEM). During the following 2-month treatment period the RI recovered nearly to preoperative levels in the EE animals, whereas the RI did not recover at all in the N animals. At the end of the experiment the animals housed in an EE were shown to have a significantly higher RI than animals under normal-housing conditions (DPO-77: EE 97.9±0.9 and N 89.8±2.5; mean±SEM; p<0.01; Fig. 1C).
Phase lag mismatch percent
At baseline, none of the animals scored a mismatch (Fig. 2A and C). After injury both the girdle mismatch percent (DPO-21: EE 14.1±4.3% and N 12.2±3.2%; mean±SEM), and the ipsilateral mismatch percent dramatically increased (DPO-21: EE 19.8±7.9 and N 26.5±7.3; mean±SEM). Thereafter, both the girdle and the ipsilateral mismatch percentages remained high in animals under normal-housing conditions, but decreased in animals under EE-housing conditions. At DPO-49 the girdle mismatch percent was significantly lower in EE animals compared to N animals (DPO-49: EE 4.3±1.5 and N 18.6±6.5; mean±SEM; p<0.01), whereas at DPO-77 the ipsilateral mismatch percent was significantly lower in EE animals compared to N animals (DPO-77: EE 7.0±2.0 and N 25.9±6.0; mean±SEM; p<0.05; Fig. 2A and C).
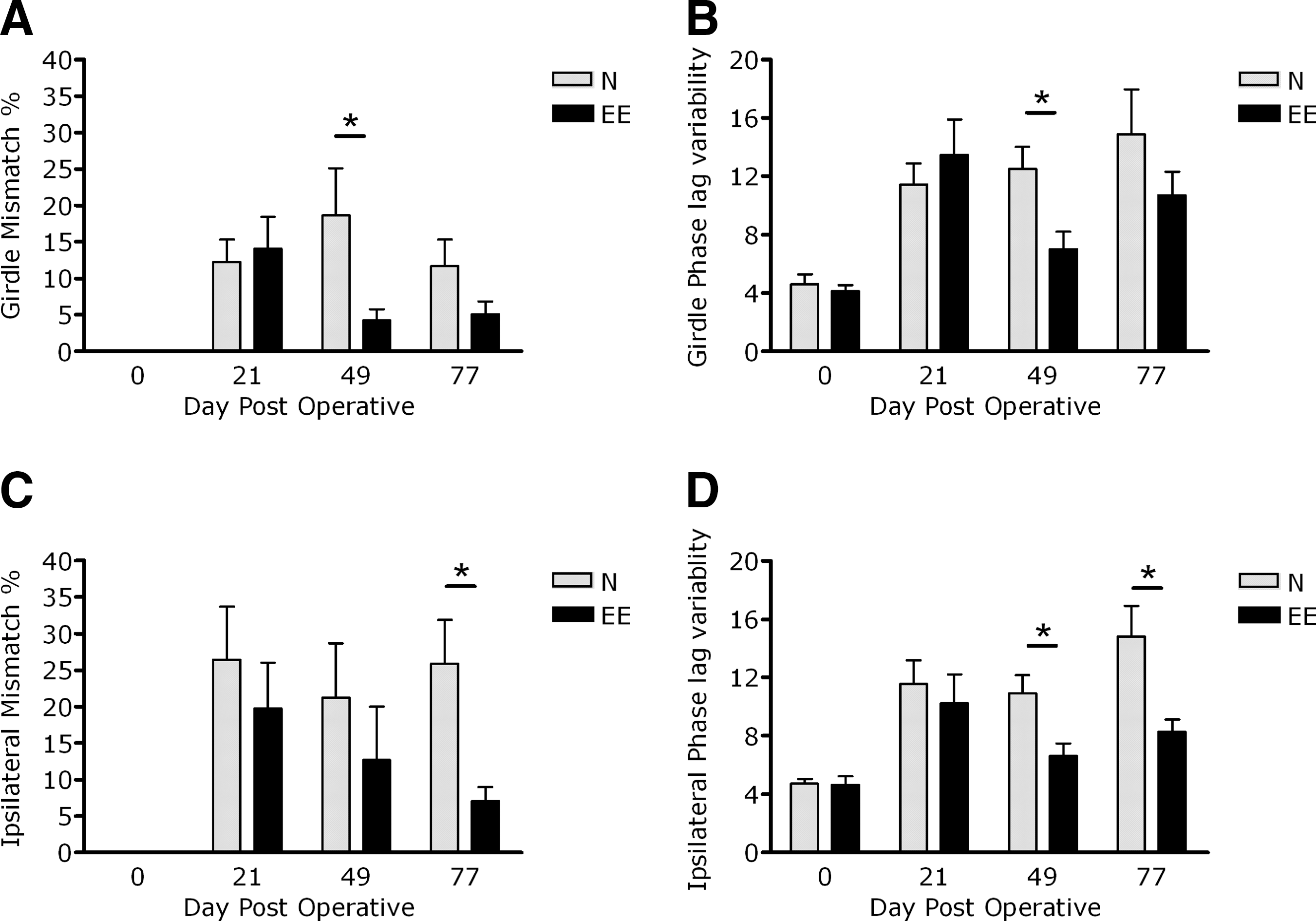
Interlimb coordination as assessed by the CatWalk parameters mismatch percentage and phase lag variability. The girdle (
Phase lag variability
The phase lag variability was about 4.5% at preoperative testing for both the girdle and ipsilateral pairs (Fig. 2B and D). At DPO-21 the girdle variability (DPO-21: EE 13.5±2.4 and N 11.4±1.4; mean±SEM) and the ipsilateral variability (DPO-21: EE 10.2±2.0 and N 11.6±1.6; mean±SEM) were increased. One month later, the girdle variability in animals under normal-housing conditions remained increased, but was decreased significantly in animals under EE-housing conditions (DPO-49: EE 7.0±1.2 and N 12.5±1.5; mean±SEM; p<0.05; Fig. 2B). The ipsilateral variability revealed even more strongly significant differences than the girdle variability, as shown by the complete recovery to preoperative values at DPO-49 (DPO-49: EE 6.6±0.9 and N 10.9±1.2; mean±SEM; p<0.05; Fig. 2B and D). At the end of the experiment, significant differences remained for the ipsilateral variability (DPO-77: EE 8.3±0.9 and N 14.8±2.1; mean±SEM; p<0.05; Fig. 2D), whereas this was not noted for the girdle variability (DPO-77: EE 10.7±1.6 and N 14.9±3.1; mean±SEM; n.s.; Fig. 2B)
Increased serotonergic innervation of the ventro-lateral grey matter after EE
The area fraction of 5-HT-IR structures was shown to be significantly increased exclusively in the ventro-lateral grey matter of lumbar segments L1–L2 after EE housing compared to normal-housed animals and sham-operated animals (p<0.05 by Mann-Whitney U; Fig. 3). No significant differences between the groups were noted in the amount of serotonergic structures in the medial-lateral grey matter of lumbar segments L1–L2.
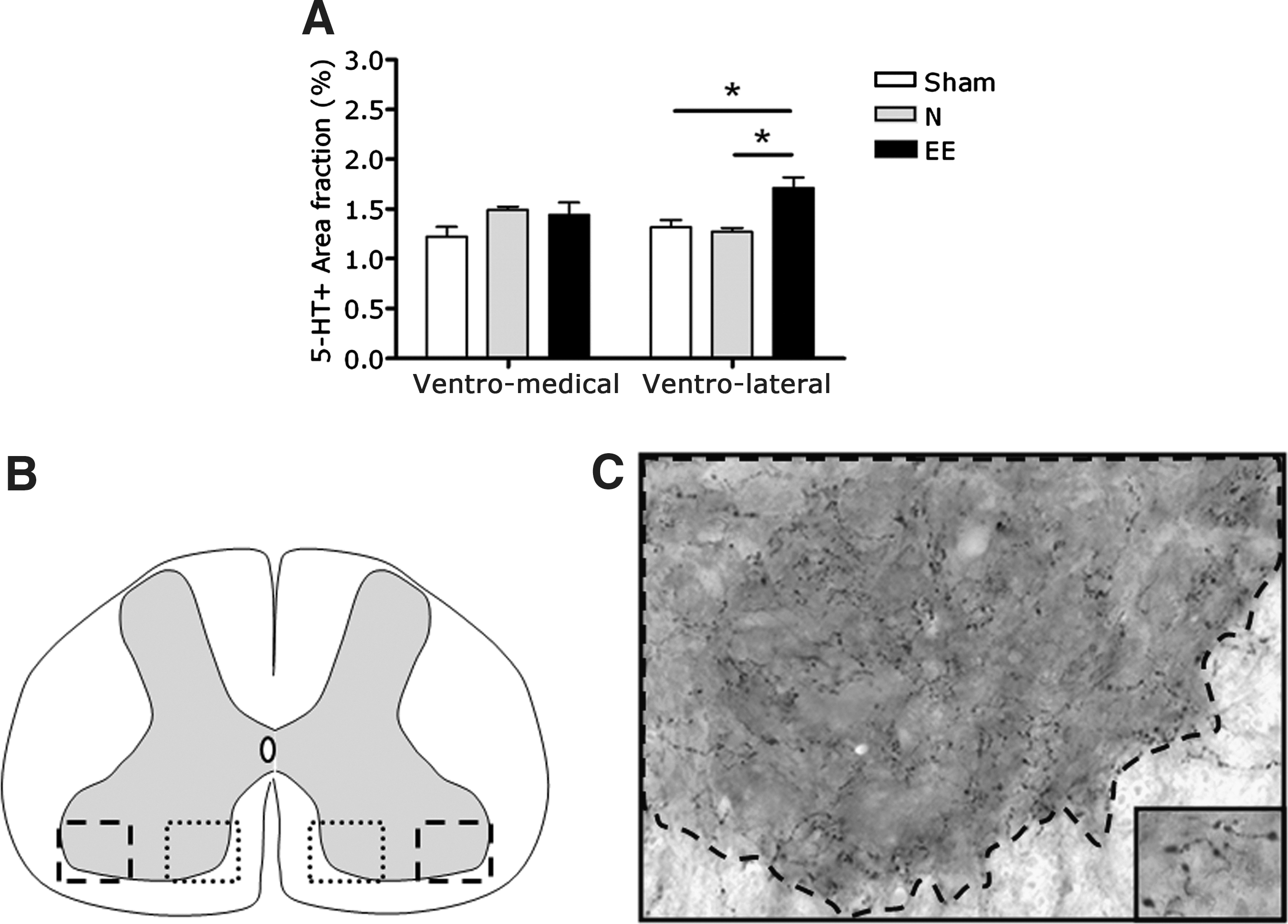
Serotonergic fiber innervation of the ventro-medial and ventro-lateral grey matter of the L1–L2 spinal cord. (
Total number of BrdU-IR cells in the rostral lumbar segments L1–L2 after SCI
At the end of the experiment the total number of BrdU-IR cells within the L1–L2 grey matter, which had been labeled 2 months earlier, were evaluated. Housing conditions had no effect on the number of cells that had proliferated at DPO-21 and survived to DPO-77 (EE 8901±602 and N 11339±1991; mean±SEM; n.s.; Fig. 4A). Neither estimates of the total volume nor the density of BrdU-IR cells within the grey matter revealed significant differences between the two groups (data not shown), implying that the housing condition did not affect the survival of newly divided cells within the rostral lumbar grey matter.

Stereological investigations of the total numbers of bromodeoxyuridine (BrdU)-immunoreactive cells within the grey matter of the L1–L2 spinal cord. Significantly more BrdU-immunoreactive cells were noted in the total grey matter (
Both SCI groups (N and EE animals) were shown to have a higher total number of BrdU-IR cells within the L1–L2 grey matter compared to sham-operated animals (Fig. 4A). This can be explained by significant group differences within the ventral horn (EE 4827±326, N 6800±1449, and sham 1817±97; mean±SEM; p<0.05; Fig. 4B). No significant differences between the groups were noted for the total number of BrdU-IR cells within the dorsal horn (Fig. 4C).
Phenotypic identification of BrdU-IR cells in lumbar segments L1–L2 after SCI
To address the question of whether the differentiation of dividing cells in rostral lumbar segments L1–L2 could underlie the EE-induced improved functional recovery after SCI, we performed cellular double-labeling, which in all cases was verified by confocal microscopy in the xy, xz, and yz planes. In the present study we focused on two specific adult cell markers: GFAP, which identifies mature astrocytes (Table 1 and Fig. 5), and NeuN, which suggests differentiation towards a neuronal fate (Table 2 and Figs. 6 and 7).

Bromodeoxyuridine (BrdU) colocalization with the astroglial marker glial fibrillary acid protein (GFAP) in the L1–L2 spinal cord, with xy, xz, and yz plane images of a typically double-labeled cell.
Bromodeoxyuridine (BrdU) colocalization with the neuronal marker NeuN in the L1–L2 spinal cord ependymal layer. A BrdU-immunoreactive nucleus (

Bromodeoxyuridine (BrdU) colocalization with the neuronal marker NeuN in the L1–L2 spinal cord substantia gelatinosa. Two BrdU-immunoreactive nuclei (
Each value is the raw count of either BrdU+ or BrdU+/GFAP+ cells in 3 sham, 5 normal-housed, and 5 environmental enrichment-housed animals.
CL, cell layer; GM, grey matter; n.o., not observed; BrdU, bromodeoxyuridine; GFAP, glial fibrillary acid protein; IR, immunoreactive.
Each value is the raw count of either BrdU+ or BrdU+/NeuN+ cells in 3 sham, 5 normal-housed, and 5 environmental enrichment-housed animals.
CL, cell layer; GM, grey matter; n.o., not observed; BrdU, bromodeoxyuridine; NeuN, neuron-specific nuclear protein; IR, immunoreactive.
BrdU/GFAP-IR cells were observed throughout the grey matter, with the exception of the ependymal layer. GFAP-IR cells comprised a substantial percentage of the BrdU-labeled cells, with 24%, 35%, and 28% in SCI-EE, SCI-N, and sham-operated animals, respectively. In the ventral horn the proportion of GFAP-labeled cells in both the EE-housed and normal-housed animals was higher compared to the proportion of these cells in the ventral horn of sham-operated animals. No significant differences were noted as a consequence of the housing condition after SCI (i.e., EE or N). In the dorsal horn the proportion of BrdU/GFAP-IR astrocytes was comparable between the groups (Table 1 and Fig. 5).
In sham-operated animals we observed BrdU/NeuN-IR cells in the ependymal layer in 2 out of 3 animals, but no BrdU/NeuN-IR cells were present in the dorsal or ventral horn. However, in the spinal cords of injured animals BrdU/NeuN-IR cells were present in two distinct regions within the lumbar grey matter: (1) in the ependymal layer (Fig. 6), and (2) in the substantia gelatinosa (Fig. 7). In 6 of 11 animals with an SCI, we found BrdU/NeuN-IR cells in the ependymal layer. All spinal cord-injured animals (n=11) contained BrdU/NeuN-IR cells in the dorsal horn. The housing condition after SCI (i.e., EE or N) did not affect the number of BrdU/NeuN-IR cells in either the ependymal layer or the dorsal horn (Table 2). A true neuronal-like morphology of the BrdU/NeuN-IR cells with processes and cell contacts was not observed. However, the BrdU/NeuN-IR cells in the substantia gelatinosa had an identical appearance as the surrounding NeuN-IR neurons. Their morphology was characterized by a more rounded shape and relatively large nuclei, as shown in Figure 6.
Discussion
The present study demonstrates that delayed EE after experimental spinal cord contusion ameliorates the recovery of locomotor performance. The recovery of interlimb coordination, as assessed by sensitive and objective CatWalk parameters, was shown to be significantly improved as a consequence of EE housing conditions. Moreover, a significant increase of 5-HT-IR structures, observed in the ventro-lateral grey matter of lumbar segments L1–L2, might underlie the enhanced locomotor recovery. The survival and differentiation of BrdU-IR cells were shown not to be affected by EE after SCI. Interestingly, we observed BrdU/NeuN double-labeled cells in the ependymal cell layer and in the substantia gelatinosa of lumbar segments L1–L2 in both EE-housed and normal-housed animals after SCI.
Our data show that EE, as a rodent equivalent to human rehabilitation strategies, is effective in improving locomotor outcomes after SCI when started after a clinically-relevant delay of 3 weeks. Further evidence has indicated that delaying the onset of rehabilitation strategies circumvents impairments in untrained functions (Krajacic et al., 2009). Treadmill running is an essential component of EE, and therefore a task-specific training paradigm to enhance locomotor outcome after SCI in the rat (Multon et al., 2003; Hutchinson et al., 2004; Engesser-Cesar et al., 2007; Heng and de Leon, 2009). Moreover, locomotor training in EE is fully voluntary and does not require any specific restrictive measures for the animals. The mechanisms underlying the EE-induced gain of locomotor abilities after SCI have thus far been only poorly investigated.
The present results of the serotonergic innervation of the ventral grey matter suggests that EE housing stimulates adaptive plasticity within the spinal locomotor network, which may eventually result in enhanced locomotor performance. We observed a significant increase in serotonergic innervation exclusively in the ventro-lateral grey matter, and not in the medial-lateral grey matter, of lumbar segments L1–L2 after EE housing. EE particularly induced a recovery of interlimb coordination. Serotonin plays a key role in the generation and organization of CPG-induced rhythmic activity for locomotion, such as interlimb coordination (MacLean et al., 1998; Madriaga et al., 2004; Christie and Whelan 2005; Deumens et al., 2005; Liu and Jordan, 2005; Pearlstein et al., 2005), since locomotor-like activity can be evoked in the isolated rodent spinal cord by a combination of NMDA and serotonin (Cazalets et al., 1995; Kjaerulff and Kiehn, 1996; Branchereau et al., 2000). Recently it has been shown that serotonin is able to generate putative locomotor-like activity by increasing the excitability of ascending commissural interneurons (aCINs), an interneuron population that constitutes the CPG for locomotion (Zhong et al., 2006). These CINs are predominantly located in the ventro-medial (laminae VII, VIII, and X) spinal cord within lumbar segments L1–L3, where the muscle activity between the left and right side of the cord is coordinated (Butt et al., 2002a,2002b; Kiehn, 2006). In the present study we did not observe an effect of EE on the serotonergic innervation of the ventro-medial grey matter within the L1–L2 segments. Neither did we observe an effect of EE on the recovery of coordination between the left and right hindpaws, as assessed by the girdle mismatch percent and the girdle phase lag variability, at the end of the experiment. However, we did observe a significant increase in serotonergic innervation of the ventro-lateral grey matter within the L1–L2 segments of EE-housed animals compared to normal-housed animals. This increase in 5-HT-IR structures provides indirect evidence of serotonergic sprouting (Holmes et al., 2005). Sprouting of 5-HT-IR fibers might originate from spared raphespinal fibers or from the propriospinal pathway, which is known to underlie interlimb coordination in quadrupedal locomotion (Juvin et al., 2005). In particular, plasticity within the latter pathways might explain the observed recovery of bilateral limb coordination, as assessed by the ipsilateral mismatch percent and the ipsilateral phase lag variability (Juvin et al., 2005). Our finding of enhanced serotoninergic innervation of the ventro-lateral grey matter (i.e., the location of motoneuron pools predominantly involved in the innervation of the distal musculature) after EE correlates with the improved coordination-related outcome. As such, these data suggest, but do not indisputably prove, a contribution of the 5-HT system in the observed behavioral benefit.
Locomotor exercise has furthermore been reported to influence cytogenesis in the intact and injured spinal cord (Cizkova et al., 2009; Foret et al., 2010; Krityakiarana et al., 2010). Therefore, the survival and differentiation of spinal cord progenitor cells within the grey matter of the CPG-related L1–L2 segments was investigated as a second putative process influenced by EE. In adult mammals, limited proliferative activity persists in the ependyma, but progenitor cells have also been detected in spinal white matter and in the substantia gelatinosa under normal conditions (Adrian and Walker, 1962; Rakic and Sidman, 1968; Bruni, 1998; Horner et al., 2000; Xu et al., 2008; Hamilton et al., 2009; Barnabe-Heider et al., 2010; Kang et al., 2010). In a single report, neurogenesis was documented within the dorsal part of the intact spinal cord (Shechter et al., 2007). The proliferative response of progenitor cells strongly increases after several types of trauma to the spinal cord (Beattie et al., 1997; Johansson et al., 1999; Namiki and Tator, 1999; Ke et al., 2006; Barnabe-Heider et al., 2010). The approach that allowed characterization of BrdU/NeuN double-labeled cells after SCI in two distinct regions within the spinal cord of both EE-housed and normal-housed animals was methodologically comparable to the approach used to characterize neurogenesis in the adult rodent forebrain (Kempermann et al., 1997; Shors et al., 2001; Gage, 2002). Additionally, this approach was used to examine the fate of proliferating cells after SCI in rats (McTigue et al., 2001; Zai and Wrathall, 2005; Zai et al., 2005), and primates (Yang et al., 2006). These and other studies all revealed that in the spinal injury zone, progenitor cells differentiate into glia (astrocytes and oligodendrocytes), but definitely not into neurons (Takahashi et al., 2003; Mothe and Tator, 2005; Zai and Wrathall, 2005; Barnabe-Heider et al., 2010). Indeed, it is known that environmental cues dictate the fates of endogenous progenitor cells, and that neurogenesis is not favored following SCI. Based on our findings it is likely that a small number of the progeny of neural stem cells or neuronal progenitor cells located within the ependyma of both the injured (EE and N) and uninjured (sham) spinal cords differentiate into a neuronal fate. Whether these cells have the ability to reach a functionally-mature neuronal status remains to be investigated. To the best of our knowledge, we are the first to describe neurogenesis in the injured adult rat spinal cord. Neurogenesis has, however, been reported following experimental autoimmune encephalomyelitis (Danilov et al., 2006), and following a specific combination strategy to induce growth factors and transcription factors in the injured spinal cord (Ohori et al., 2006). Although neurogenesis in the dorsal spinal cord was reported following dorsal rhizotomy in both rats and monkeys (Vessal et al., 2007), evidence for neurogenesis following SCI has thus far been lacking. Importantly, previous investigations have mostly focused on areas close to the spinal lesion, whereas we investigated regions far caudal to the spinal lesion (i.e., the CPG area that is directly associated with hindlimb function). Although the finding of neurogenesis in the lumbar spinal cord following SCI is remarkable in our opinion, it was found not to be affected by EE, and thus does not explain the EE-induced improvements seen in locomotor function after SCI.
The lumbar spinal cord is considered a favorable area for neurogenesis (Weiss et al., 1996). Neural stem cells, characterized by capacities for self-renewal and multipotentiality (Reynolds and Weiss 1992; Morshead et al., 1994; Weiss et al., 1996), have been isolated from the spinal cord ependymal layer (Weiss et al., 1996; Taupin and Gage, 2002). We found that neuronal differentiation of lumbar progenitor cells did not only occur in the ependymal layer, but also in the substantia gelatinosa (lamina II; Rexed, 1952). We also observed that these cells were often clustered, as can be seen in Figure 7. However, BrdU/NeuN-IR cells were only observed in the L1–L2 substantia gelatinosa following SCI. This might be explained by the role that the substantia gelatinosa plays as a major recipient of peripheral inputs from small-diameter primary afferents (Aδ and C), that predominantly convey noxious sensation (Kumazawa and Perl, 1978; Sugiura et al., 1986). SCI-induced changes in sensory inputs to the substantia gelatinosa might be involved in a functional reorganization within this neuronal circuit, possibly even in neuronal differentiation of endogenous progenitor cells.
In conclusion, EE initiated at 3 weeks after SCI improves recovery of interlimb coordination, and it is likely that increased serotonergic innervation of the ventro-lateral grey matter within the L1–L2 segments is behind this observed recovery. Although EE housing affected neither the survival nor the differentiation of BrdU-IR cells after SCI, we remarkably observed BrdU/NeuN-IR cells in the ependymal cell layer and in the substantia gelatinosa of the rostral-lumbar segments L1–L2 in both EE-housed and normal-housed animals after SCI.
Footnotes
Acknowledgments
This work was supported by a grant from the International Spinal Research Trust (ISRT; STR057 to E.A.J.). R.D. was supported by the Dutch Society for Anesthesiology (NVA; Young Investigator Grant to R.D.).
Author Disclosure Statement
No competing financial interests exist.