
Abstract
Traumatic brain injury (TBI) induces transient or persistent dysfunction of gait and balance. Enhancement of cholinergic transmission has been reported to accelerate recovery of cognitive function after TBI, but the effects of this intervention on locomotor activity remain largely unexplored. The hypothesis that enhancement of cholinergic function by inhibition of acetylcholinesterase (AChE) improves locomotion following TBI was tested in Sprague-Dawley male rats after a unilateral controlled cortical impact (CCI) injury of the motor-sensory cortex. Locomotion was tested by time to fall on the constant speed and accelerating Rotarod, placement errors and time to cross while walking through a horizontal ladder, activity monitoring in the home cages, and rearing behavior. Assessments were performed the 1st and 2nd day and the 1st, 2nd, and 3rd week after TBI. The AChE inhibitor physostigmine hemisulfate (PHY) was administered continuously via osmotic minipumps implanted subcutaneously at the rates of 1.6–12.8 μmol/kg/day. All measures of locomotion were impaired by TBI and recovered to initial levels between 1 and 3 weeks post-TBI, with the exception of the maximum speed achievable on the accelerating Rotarod, as well as rearing in the open field. PHY improved performance in the accelerating Rotarod at 1.6 and 3.2 μmol/kg/day (AChE activity 95 and 78% of control, respectively), however, higher doses induced progressive deterioration. No effect or worsening of outcomes was observed at all PHY doses for home cage activity, rearing, and horizontal ladder walking. Potential benefits of cholinesterase inhibition on locomotor function have to be weighed against the evidence of the narrow range of useful doses.
Introduction
Experimental animal models of TBI have explored pharmacological effects on learning and memory deficits, but little is known about pharmacological modulation of locomotor function (Hoane and Barth, 2002; Hoane et al., 2008; Sharma et al., 2006). The prominent alterations in cognitive function observed after severe brain trauma (Arciniegas, 2003; Geffen et al., 1991; Gualtieri and Cox, 1991), taken together with the established role of cerebral cholinergic mechanisms in learning (Chrobak et al., 1988; Dawson et al., 1991; Luine and Hearns, 1990) have aroused interest in the exploration of the role of acetylcholine (ACh) in the pathophysiology of TBI.
A systematic study of the consequences of cerebral trauma on the cerebral cholinergic system is still lacking. The effects of modulation of cholinergic transmission on behavioral outcomes following brain trauma are still controversial, with depression or enhancement of cholinergic transmission having conflicting effects depending upon the agent or time after injury.
The recent reports on accelerated cognitive recovery from brain trauma after treatment with acetylcholinesterase (AChE) inhibitors (Cardenas et al., 1994, Chen et al., 1998; Khateb et al., 2005; Sasaki et al., 1987; Silver et al., 2006, 2009; Taverni et al., 1998; Tenovuo, 2005; Zhang et al., 2004), as well as the dose-related effects of cholinesterase inhibition on electrophysiological abnormalities among persons with TBI (Arciniegas and Topkoff, 2004), support the role of cholinergic factors in the functional impairments following TBI and offer promise as a therapeutic strategy. However, the possible modulation of motor function by cholinergic interventions has remained largely unexplored.
This work was undertaken with two objectives: 1) to determine which motor functions were most severely and persistently altered after a cortical impact injury and 2) to explore the ability of enhanced cholinergic transmission by inhibition of AChE to ameliorate those deficits. In addition, the influence of AChE inhibition on motor function of animals with no TBI was undertaken to determine the possible contribution of confounding variables, specifically the possible negative effects of this intervention on the same motor tests used in animals with TBI.
Methods
Animals
Sixty-eight Sprague-Dawley male rats (250–300 g body mass, Harlan Sprague Dawley, Placentia, CA) were used. Thirty-five of them received TBI and 33 received a sham intervention as described subsequently. TBI animals were divided into five groups receiving saline (n=9) or physostigmine (PHY) continuous subcutaneous infusion at the rates of 1.6 (n=6), 3.2 (n=6), 6.4 (n=7), or 12.8 (n=7) μmol/kg/day. Matching groups were set up for sham intervention rats that received saline (n=9) or PHY at 1.6 (n=6), 3.2 (n=6), 6.4 (n=6), or 12.8 (n=6) μmol/kg/day. The research environment and protocol for animal experimentation were approved by the Institutional Animal Care and Use Committee (IACUC) of the West Los Angeles Veterans Affairs Healthcare System and the USC School of Medicine. The animal facilities at these Institutions are accredited by the Association for Assessment and Accreditation of Laboratory Animal Care (AAALAC), International. Animals were group housed during a 1-week period of acclimatization to the vivarium prior to experimentation. Rats were individually housed following surgery, with free access to water and standard rodent chow for the duration of the study.
Controlled cortical impact (CCI)
Rats were anesthetized with isoflurane (induction 3.5%, maintenance 2.0%). The scalp was shaved and cleansed with Ioprep® and the animal mounted in a stereotaxic apparatus in the skull-flat position. Following sterile technique, a right parasagittal skin incision was performed, the galea incised and reflected, and a craniotomy fashioned, with a 6.2-mm trephine, centered at 1.2 mm anterior and 3.5 mm lateral (right side) to bregma. Severe trauma of the left cerebral cortex was produced as previously reported (Scremin et al., 2006), with a modification of the method of Allen (Allen, 1914) and Feeney and associates (1981). We have previously shown that in a 2-week follow-up of this CCI injury, there remain profound alterations in Ach and choline dynamics in the region of impact, with lesser, although significant, changes in Ach turnover and choline uptake in peripheral regions (Scremin et al., 2006). The apparatus consisted of a vented stainless steel guide tube mounted on a micromanipulator through which a 20 g weight was dropped from a height of 35 cm to strike a stainless steel circular footplate (4.5 mm diameter) resting on the exposed dura mater of the right motor-sensory cortex, with its center 1.2 mm anterior and 3.5 mm lateral to bregma. This site was selected to coincide with the maximum activation of the primary motor cortex observed in rats during treadmill walking (Holschneider et al., 2003). The indentation produced in the cortex by the footplate was limited to 3.0 mm. After trauma, the galea was sutured to cover the craniotomy and the skin closed with 2-0 prolene stitches. With this technique, the cortex subjacent to the striking plate undergoes infarction with later development of a cyst (Scremin et al., 1997). Functional imaging indicates widespread depression of function in the cerebral cortex on the traumatized side (Scremin et al., 2007). Sham intervention animals were anesthetized as described previously, and skin and galea incisions were made and then sutured, but no craniotomy and cortical impact were produced. Animals were provided a source of heat to prevent postoperative hypothermia. Antibiotic (cefazolin Na, 100 mg/kg i.m.) and analgesic therapy (flunixin meglumine, 1.1 mg/kg, s.c.), twice daily, was maintained during 3 days after TBI or sham interventions.
Behavioral testing
Animals were familiarized with the behavioral tasks 3 days prior to surgery by daily exposure, with the final day serving as the pre-surgery baseline. Overnight testing of home cage locomotor activity preceded the daytime testing in the open field, followed by the horizontal ladder, and the Rotarod. Testing occurred on postsurgical days 1, 2, 7, 14, and 21.
Rotarod test
Coordination, balance, and strength were evaluated on the Rotarod treadmill (Columbus Instruments, Columbus, OH). Animals were familiarized with the task 3 days prior to surgery by daily exposure, with the final day serving as the pre-surgery baseline. Constant speed and acceleration paradigms were used, which evaluated the maximum time and speed, respectively, that animals can walk on the rotating cylinder prior to falling. Constant speed was 20 rpm (7.6 cm/sec) with a cutoff for the trial duration of 300 sec. In the acceleration paradigm, the initial rate was 5 rpm (1.9 cm/sec.) and accelerated at a rate of 5 rpm/min with a cutoff for the trial duration of 360 sec. Two trials were undertaken for each animal, with a 20-min rest period in between trials, with the constant speed testing performed in the morning and the acceleration paradigm performed in the afternoon.
Horizontal ladder
Animals were placed on an apparatus (Foot Misplacement Apparatus, Columbus Instruments, Columbus, OH) that consisted of a set of stainless steel horizontal bars that configure a horizontal ladder (length 1.25 m, width 20.3 cm, 3.8 cm inter-rung distance, 0.8 cm rung diameter, 48.9 cm height above floor, lighting 750 lux). At one end of the ladder, a dark compartment was placed to entice the animal to walk toward the perceived “safety” of the dark cover. The animal was placed on one end of the ladder, and the number of missteps as it moved toward the dark compartment at the other end of the ladder was counted automatically by detecting the change of resistance between the ladder and a metal plate below it. Each time the animal missed one of the rungs of the ladder and touched the metal plate below, a very small electric current was detected, amplified, and counted by a separate computerized counter. The animal was not aware of this current, as it was far below the threshold of the animal's sensitivity. Three trials were performed per day with an inter-trial duration of 10 min. Missteps, as well as the time to cross, were assessed.
Home cage activity
Ambulatory activity and total activity (ambulatory+stereotypic movement) of individually housed rats was recorded in the animal's vivarium home cage for 12 h (dark phase, 7 p.m.–7 a.m.) using triple axis infrared activity monitors placed around the animal's home cages (Opto-M3, Columbus instruments, Columbus, OH). Horizontal (XY) and vertical (Z) activity was recorded.
Open field rearing
Animals were placed in a clear, cylindrical, Plexiglas cage (28.5 cm diameter, 30.5 cm height, lighting 750 lux). Rearing activity was recorded by video camera over 5 min with the observer out of the room. Scoring of rearing, and left/right paw use during rearing was scored manually from the video.
Administration of PHY
Physostigmine hemisulfate (Sigma Aldrich, St. Louis, MO) and ethylenediaminetetraacetic acid disodium salt dehydrate (EDTA, Sigma Aldrich, St. Louis, MO) were dissolved at equimolar concentrations in double distilled boiled water, and 2 mL of the solution were used to fill each osmotic pump (Alzet Model 2ML4, Durect Corp. Cupertino, CA). The pumps were cylindrical in shape, with length of 3 cm, diameter of 0.7 cm, weight of 1.1 g empty and delivered 2.5 μL/h continuously during 28 days. They were inserted into a subcutaneous space created by blunt dissection of the interscapular region of animals under isoflurane anesthesia. The skin was closed with 2-0 prolene sutures. The molar infusion rates of PHY were 1.6, 3.2, 6.4, and 12.8 μmol/kg/day. Drug delivery was confirmed on explantation of the pump at the end of the experiment by aspiration of the remaining drug, as well as weighing of the pumps.
Regional brain activity of AChE
Animals were euthanized by decapitation while under deep isoflurane anesthesia (3.5% in 30% O2 balanced with N2O). The brain was rapidly removed and flash frozen in methylbutane cooled to –55°C. One coronal brain slice was obtained between coordinates 1 mm rostral and 4 mm caudal to bregma. Two cortical blocks were dissected on each hemisphere, including the primary motor (M1) and primary somatosensory (S1) areas and portions of secondary motor (M2) and secondary somatosensory (S2) areas. These tissue samples were homogenized, and aliquots of these homogenates were used to determine tissue AChE activity with the kinetic method of Ellman (Ellman et al., 1961).
Data analysis
Behavioral data for each animal were averaged at each postsurgical follow-up evaluation when duplicate or triplicate trials were performed on the same day. Two analysis strategies were used. A repeated measures analysis of variance (ANOVA) was performed for each group, test, and dose tested. If the F ratio for the within factor “time after intervention” was statistically significant (p<0.05), a Dunnett multiple comparisons test was performed to determine significance of differences between the pre-TBI and sham means and means at days 1, 2, 7, 14, and 21 days after those interventions. In another analysis, ANOVA was performed for every test with factor “dose” with four levels (1.6, 3.2, 6.4, and 12.8 μmol/kg/day) at each time after TBI or sham intervention (1, 2, 7, 14, and 21 days). If the F ratio was significant, a Dunnett multiple comparisons test was performed to determine significance of differences of the corresponding outcomes between each dose and the “no-drug” condition.
Results
Effects of PHY on cerebral AChE
AChE activity in the cerebral cortex decreased as a function of PHY dose (Fig. 1). At the PHY infusion rates of 1.6 and 3.2 μmol/kg/day, no signs of cholinergic toxicity were observed at any time. At 6.4 μmol/kg/day, tremor and muscle fasciculations were observed in the head and body during the first hours after surgery for osmotic pump implantation. No such phenomena were observed at these doses after the first day post-implantation. At 12.8 μmol/kg/day, in addition to tremor and muscle fasciculations, an increment in salivary secretion was observed during the few hours that followed surgery for osmotic pump implantation. After the first day post-implantation, these effects disappeared. The toxic manifestations were observed in animals with TBI, as well as in the sham injury controls.
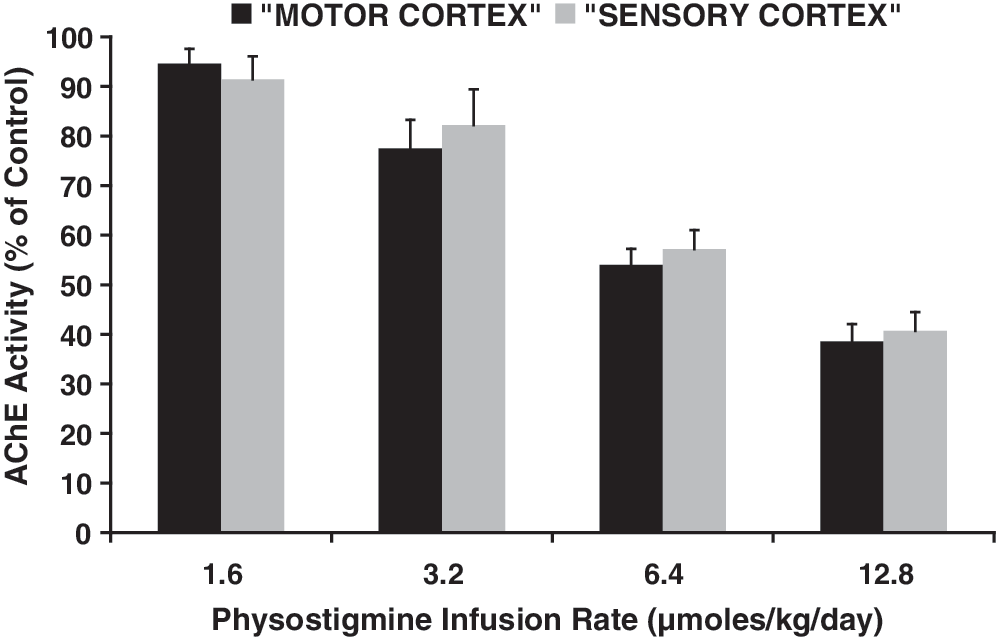
Mean and S.E. of acetylcholinesterase (AChE) activity in motor and sensory cortex of four groups of rats that received 1.6, 3.2, 6.4, or 12.8 μmol/kg/day of physostigmine by continuous subcutaneous infusion with an implanted osmotic pump during 3 weeks. Data are expressed as percent of values in a group of normal controls. Regression of these values on log dose gave Y intercept=107.9±2.6, slope=−63.65±3.5, R2 =0.99, p=0.003 for motor cortex; and Y intercept=106.5±5.0, slope=−58.88±6.8, R2 =0.97, p=0.013 for sensory cortex.
Effects of TBI or sham interventions on locomotion
All measures of motor function were affected after TBI in the animals that received saline treatment (Figs. 2 –5, Tables 1 and 2). The maximum speed achieved on the accelerating Rotarod treadmill was significantly less than the value pre-TBI from day 1 of follow-up and until day 21 (Fig. 2A). No significant differences from pre-intervention variable means were found during follow-up in sham animals (Fig. 2B).
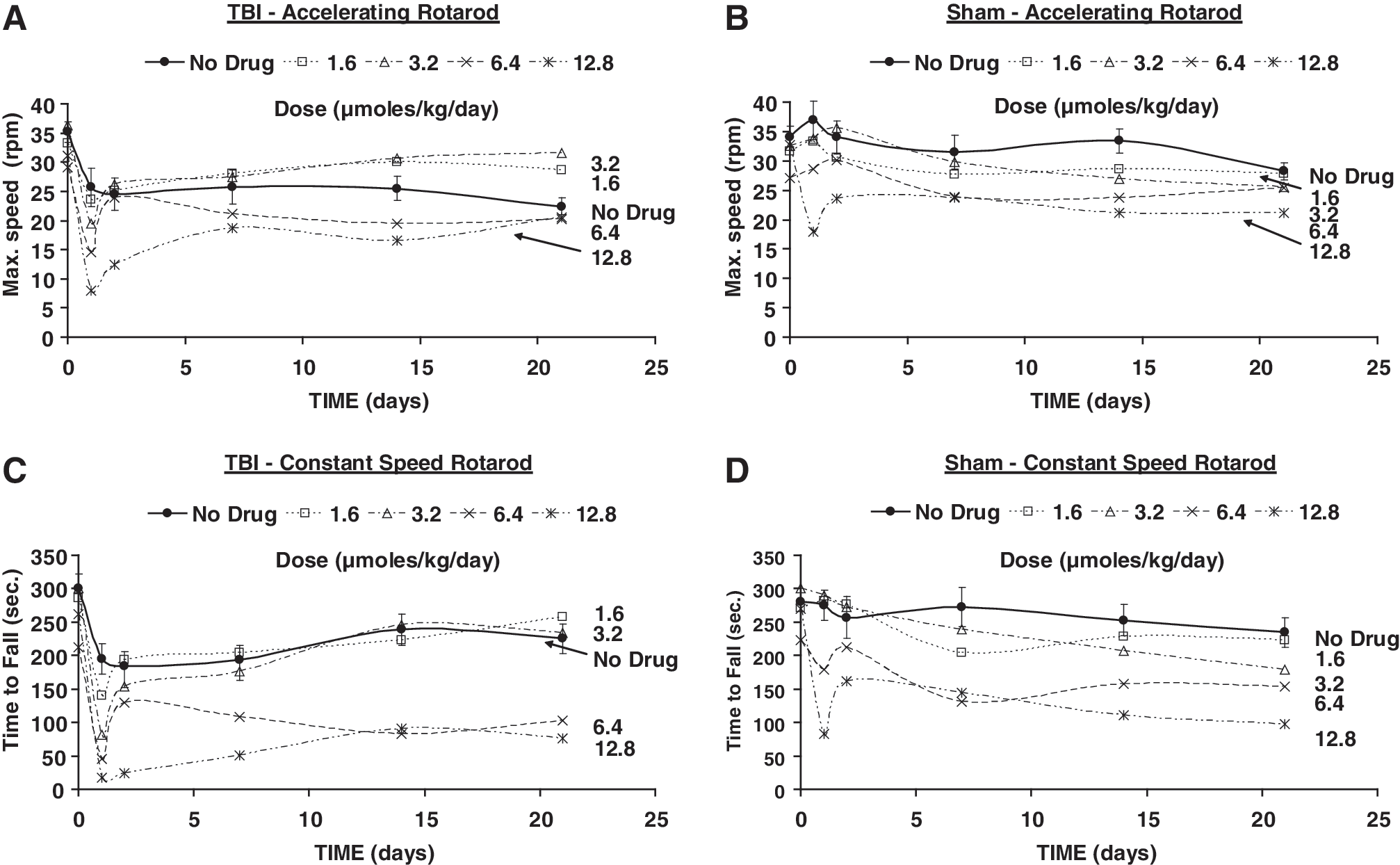
Means of the maximum speed (rpm) achieved on the accelerating Rotarod paradigm (
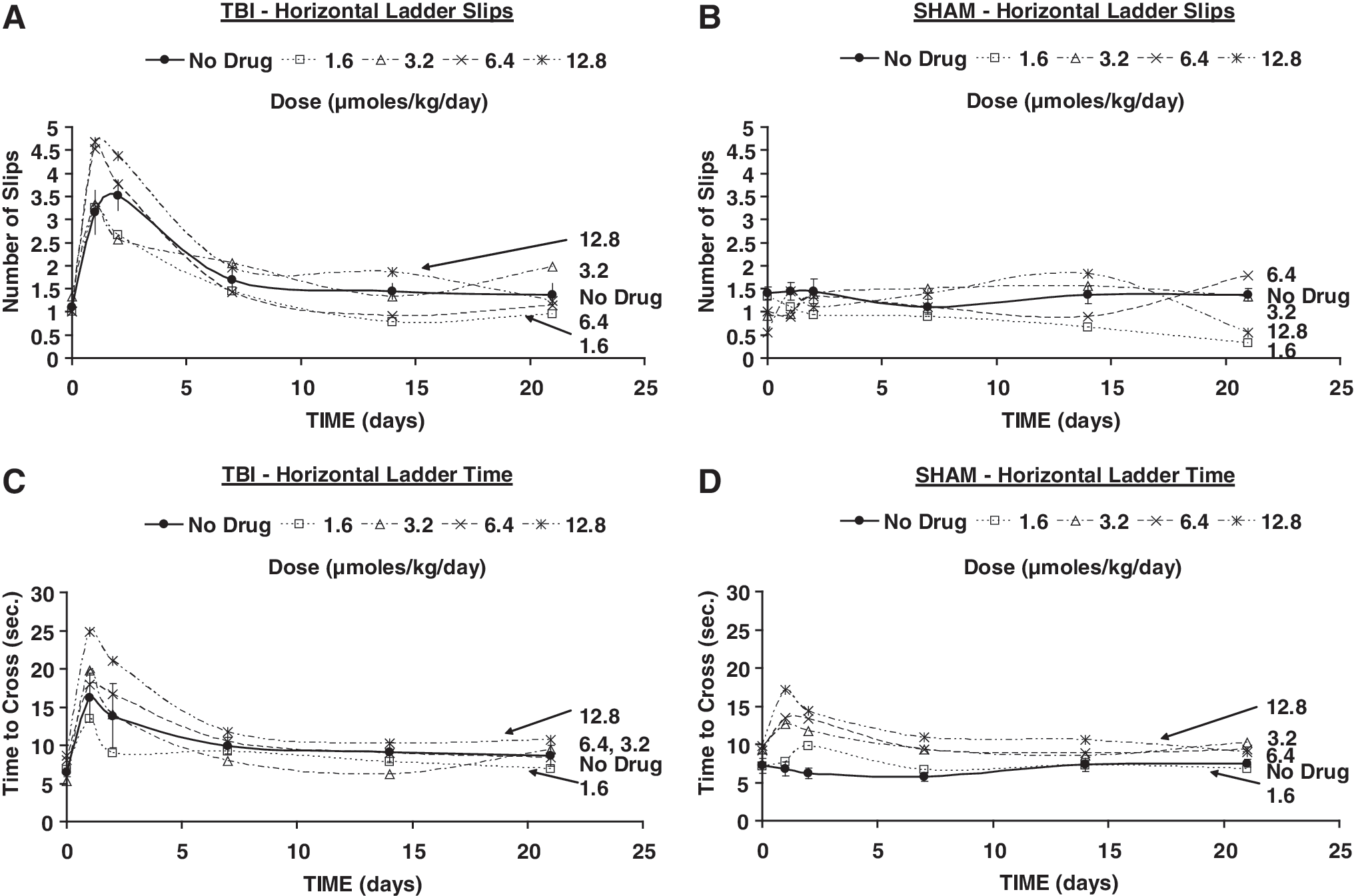
Means of errors (slips) in the horizontal ladder (
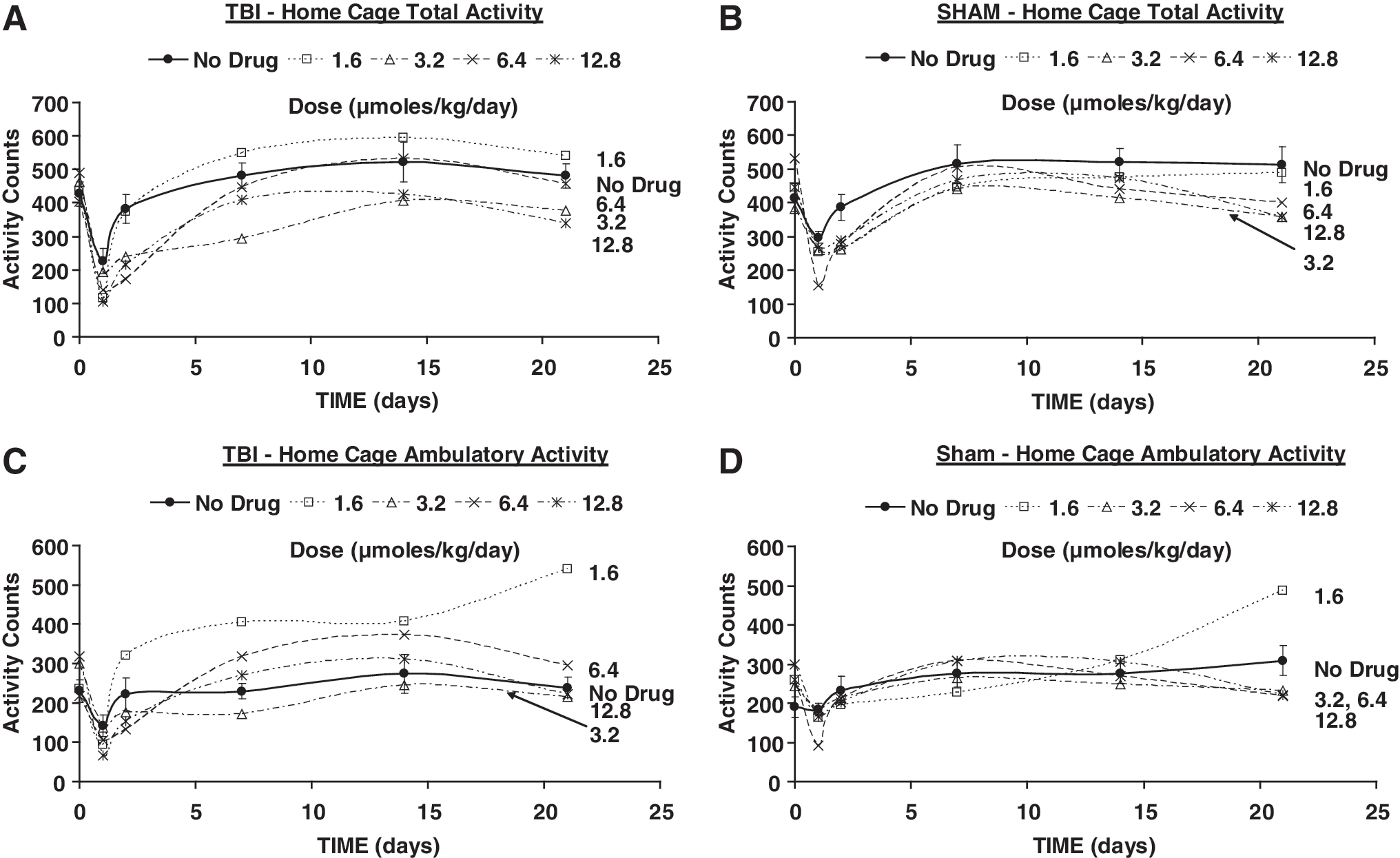
Means of home cage total activity (
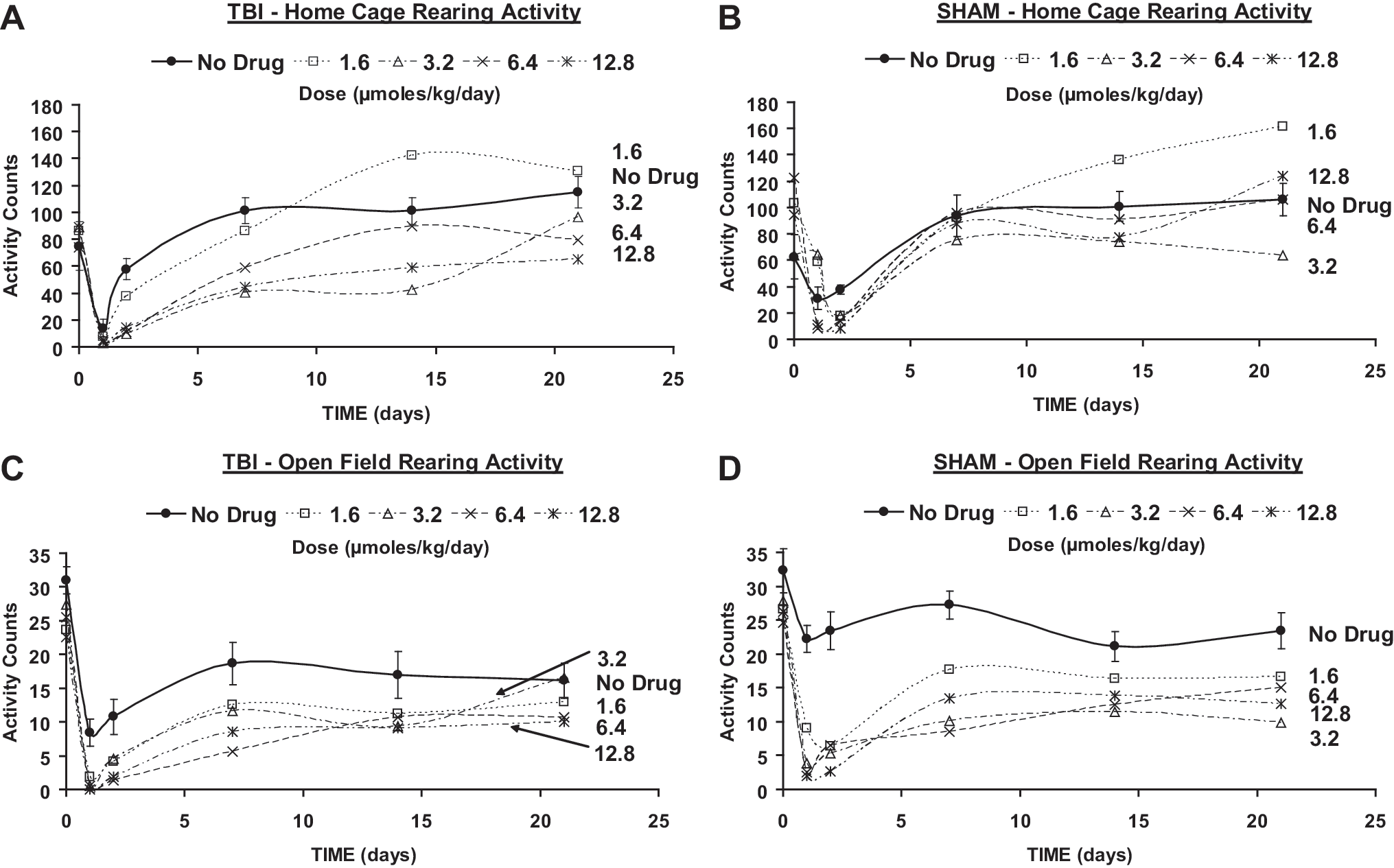
Means of home cage rearing activity (

Repeated measures analysis of vairance was performed for each group and variable tested. If the F ratio for the within factor “time after intervention” was statistically significant (p<0.05), a Dunnett multiple comparisons test was performed to determine significance of differences between the pre-TBI or pre-sham means and means at 1, 2, 7, 14, and 21 days after those interventions. Black background denotes significant deterioration (p<0.05), gray background denotes significant improvement (p<0.05), and white background denotes no significant change.

ANOVA was performed for every test with factor “dose” with 4 levels (1.6, 3.2, 6.4, and 12.8 μmol/kg/day) at each time after TBI or sham intervention (1, 2, 7, 14, and 21 days). If the F ratio was significant, a Dunnett multiple comparisons test was performed to determine significance of differences of the corresponding outcomes between each dose and the “no-drug” condition. Black background denotes significant deterioration (p<0.05), gray background denotes significant improvement (p<0.05), and white background denotes no significant change.
Time to fall from the constant speed Rotarod treadmill was significantly shorter than prior to TBI only during the first week of follow-up, with recovery thereafter (Fig. 2C), whereas no significant changes were found in sham animals (Fig. 2D).
The number of errors (slips) on the horizontal ladder (Fig. 3A) and the time to cross it were significantly greater than in the pre-TBI condition during the first 2 days of follow-up only (Fig. 3C). No significant changes were found in sham animals throughout follow-up on these variables (Figs. 3B and D).
Total activity in the home cage was significantly lower than in the pre-TBI condition only on the first day of follow-up (Fig. 4A). No significant changes were found in home cage ambulation after TBI (Fig. 4C). Sham animals showed no significant differences from pre-sham at any time during follow-up in home cage total activity. No significant change in home cage ambulation of sham animals was noted, except during week 3, when a small increase was seen (Figs. 4B and D).
Rearing behavior in the home cage in TBI animals was significantly lower than pre-TBI the first day of follow-up, with a rebound to a significantly higher value at day 7 and thereafter. Sham animals showed a decrease in home cage rearing only on the first day of follow-up, with a significant rebound thereafter above pre-sham level at day 21.
Rearing during the open field test was significantly decreased in TBI animals throughout follow-up (Fig. 5C), and the same was found in sham animals, with the exception of day 7 (Fig. 5D).
Effects of PHY on locomotion
The maximum speed achieved on the accelerating Rotarod in TBI animals that received PHY treatment recovered to values not significantly different from those pre-TBI, starting at day 2 with 1.6 μmol/kg/day, and after the first week with 3.2 μmol/kg/day. This result contrasted with that of TBI animals that received no drug, as described previously, that dropped from the Rotarod at significantly earlier times than pre-TBI throughout follow-up. At all higher PHY infusion rates the performance worsened, with significantly lower speeds achievable than pre-TBI, except for day 2 with 6.4 μmol/kg/day (Fig. 2 A). In sham animals, no significant differences from pre-sham were found at any follow-up time with 1.6 μmol/kg/day, but at 3.2 μmol/kg/day, fall speeds were significantly lower than pre-sham at days 14 and 21, whereas at 12.8 μmol/kg/day performance significantly deteriorated throughout follow-up (Fig. 2B). Performance of TBI animals on the constant speed Rotarod was diminished relative to the pre-TBI baseline at postoperative days 1 and 2 with PHY 1.6 μmol/kg/day. At higher infusion rates, a progressively persistent postsurgical impairment was noted, such that impairment was noted throughout the follow-up period at a dose of 12.8 μmol/kg/day, and at all follow-up points except on day 2 for the 6.4 μmol/kg/day dose (Fig. 2C). There was no significant improvement elicited by PHY at any dose relative to vehicle in TBI animals. The performance of sham animals was worsened at 14 and 21 days of follow-up by 3.2 μmol/kg/day of PHY, whereas with 12.8 μmol/kg/day they fell earlier than pre-sham at all times (Fig. 2D).
Errors (slips) on the horizontal ladder showed no difference between TBI animals that received saline or PHY at any dose (Fig. 3A). Time to cross the ladder improved at an earlier time (day 2) in TBI animals with 1.6 μmol/kg/min than in those that received saline, but no difference was observed at higher doses (Fig. 3C). In sham animals, significantly more errors were found with 12.8 μmol/kg/day at 1, 7, and 14 days post- sham (Fig. 3B). In sham animals, time to cross the ladder was longer than pre-sham at days 1 (12.8 μmol/kg/day) and days 1 and 2 (6.4 μmol/kg/day) (Fig. 3D).
The decreases in total activity and ambulatory activity in the home cage were extended to day 7 in TBI animals that received PHY at 3.2 μmol/kg/day (Fig. 4A and C). Decreases in home cage total activity and ambulation were also present in sham animals that received 6.4 μmol/kg/day PHY up to day 2 and day 1, respectively (Figs. 4B and D).
Home cage rearing in TBI rats was significantly decreased during the first 2 weeks at 3.2 μmol/kg/day of PHY, with the effect decreasing at higher infusion rates (Fig. 5A). Sham animals (Fig. 5B) showed diminished home cage rearing relative to the pre-sham baseline at all doses, although deficits persisted only through postoperative day 2.
In TBI rats, significantly fewer episodes of open field rearing were observed throughout the follow-up period at all infusion rates (Fig. 5C), with greatest differences noted relative to vehicle during the first postoperative week. Similar findings were observed in sham animals. In the open field, sham animals likewise showed a significant decrease in rearing episodes in response to PHY during post-sham days 1 and 2, with a significant increase noted only at 3 weeks in the saline controls (Fig. 5D).
Discussion
Cortical impact injury induced a deficit of motor function of variable intensity and duration depending upon the modality tested. The accelerating Rotarod test is perhaps the most challenging of those used in the present work. This test involves a combination of strength, endurance, balance, and adaptation to a continuously varying progression speed. This is the motor test that showed the most persistent deficit after injury and also the one in which AChE inhibition was most effective in accelerating recovery.
The measurements of AChE activity included the M1 and S1 areas and adjacent motor and sensory secondary association areas, which are preferentially activated during the Rotarod treadmill task. Our prior work using PHY has shown that the distribution of this AChE inhibitor reaches the entire brain and, therefore, the changes in AChE activity reported for the sampled areas should be representative of other cortical and subcortical areas engaged in the control of locomotion. Moreover, we have found that the cerebral concentration of PHY is correlated with the regional cerebral blood flow (rCBF) during the uptake phase of intravenous pulse administration (Scremin et al., 1990). This flow-dependent characteristic ensures that the areas that are activated during a task (i.e., with increased rCBF) will receive the greatest relative amount of PHY when it is administered continuously, as in the present experiments.
Administration of PHY with osmotic pumps induced a decrease of AChE activity that was linearly related to the log of the dose (Fig. 1). However, the effects of such changes in AChE activity on functional outcomes were far from linear. The beneficial effect of AChE inhibition was only manifest at the lower infusion rates of PHY that reduced cerebral cortical AChE activity to between 95 and 78% of control, with higher infusion rates being associated with worsening of performance in this as well as in other tests. AChE inhibitors in general and PHY in particular are known to show inverted u-shaped dose effects for cognitive impairment in Alzheimer's disease, that is, they are effective at an optimal dose with lack of effect at lower and higher doses (Stern et al., 1987). These agents are also known to have a low margin between therapeutically useful doses and those that induce undesirable effects. Both characteristics are evident in the present data. Although clinical manifestations of cholinergic toxicity (tremor, skeletal muscle fasciculations, and salivation) were only present in the immediate postoperative period at doses of ≥6.4 μmol/kg/day, and never during follow-up beyond day 1, lack of improvement or worsening of outcomes were found at these high doses in Rotarod performance, ambulatory activity, and rearing in the home cage. Deterioration of performance at these high doses was also found, although to a lesser extent, in sham animals.
Rearing in the open field was depressed at all follow-up times in the absence of PHY administration, and this phenomenon was enhanced at all doses of PHY in TBI and sham animals. AChE inhibitors are known to induce decrease in rearing frequency (Wright et al., 2010), and the presence of the phenomenon indicates the effectiveness of PHY in inducing central nervous system effects even at the lowest doses used. The phenomenon of depressed rearing, a natural occurrence in exploratory activity of a novel environment, has been interpreted in the context of the behavioral depression associated with the action of AChE inhibitors (Janowsky et al., 1972). It is interesting that the phenomenon also occurred in the familiar home cage environment, although to a lesser extent than in the novel open field environment.
A rebound (greater activity than before TBI or sham intervention) of ambulatory activity in the home cage was observed at 21 days of follow-up with the lowest dose of PHY in TBI and sham animals. This biphasic effect is in line with previous observations of rebound in the depressant effect of cholinomimetics on behavioral arousal, and it has been proposed to be caused by an interaction between cholinergic and adrenergic systems (Fibiger et al., 1971), a hypothesis that has to be reconciled with recent developments on the complex mechanisms of the effect of cholinesterase inhibitors on arousal and cognition (Hasselmo and Sarter, 2011).
Some limitations of this study and its interpretation should be addressed. It is possible that beneficial effects on motor function of higher doses of PHY may have been masked by deleterious effects on arousal. A gradual dose escalation of PHY in animals over a longer period of time might have minimized such impairments in arousal, and hence motor performance. This hypothesis remains to be tested. Likewise, it remains to be proven if higher doses of PHY may be beneficial after more severe TBI, that is, if injury directly affects subcortical structures such as the cholinergic nucleus basalis of Meynert, which plays a role in arousal. Indeed, following direct neurotoxic injury of this nucleus, PHY at higher doses has been shown to reverse attentional deficits (Muir et al., 1995). If true, the translational implications of this might be that PHY may show enhanced efficacy in TBI with broad cortical and subcortical cholinergic deficits, especially if it minimizes side effects through a gradual dose titration. Future work would have to explore such possibilities, either using the current CCI model or fluid percussion and blast injury models, which may show different dose responses.
Conclusion
In conclusion, impact injury of the motor-sensory cortex induced transient deterioration of some locomotor functions (number of missteps and time to cross a horizontal ladder, time to fall from a Rotarod treadmill at a constant speed, and home cage total and ambulatory activity) and long-term impairment of others (time to fall from the accelerating Rotarod treadmill and rearing). Continuous administration of the AChE inhibitor PHY induced improvement of performance in time to fall from the accelerating Rotarod treadmill at low doses, with lack of effect or worsening as dose was increased. Drug administration enhanced the deficit at all doses in the case of rearing. The benefit of AChE inhibition on the most complex locomotor function tested in this study suggests a potential for this treatment strategy in the management of motor deficits in TBI, but it has to be weighed against the evidence of the narrow range of useful doses.
Footnotes
Acknowledgments
This work was supported by a Department of Veterans Affairs Rehabilitation Research & Development Merit Review (VA RR&D) #B5089R and a VA RR&D Research Career Scientist award (O.U.S).
Author Disclosure Statement
No competing financial interests exist.