
Abstract
The overlapping pathologies and functional outcomes of blast-induced TBI (bTBI) and stress-related neurobehavioral disorders like post-traumatic stress disorder (PTSD) are significant military health issues. Soldiers are exposed to multiple stressors with or without suffering bTBI, making diagnosis and treatment as well as experimental modeling of bTBI a challenge. In this study we compared anxiety levels of Naïve rats to ones that were exposed to each of the following conditions daily for 4 consecutive days: C I: transportation alone; C II: transportation and anesthesia; C III: transportation, anesthesia, and blast sounds; Injured: all three variables plus mild blast overpressure. Following behavioral testing we analyzed sera and select brain regions for protein markers and cellular changes. C I, C II, and C III animals exhibited increased anxiety, but serum corticosterone levels were only significantly elevated in C III and Injured rats. C III and Injured animals also had elevated interferon-γ (IFN-γ) and interleukin-6 (IL-6) levels in the amygdala (AD) and ventral hippocampus (VHC). Glial fibrillary acidic protein (GFAP) levels were only significantly elevated in the VHC, prefrontal cortex (PFC), and AD of Injured animals; they showed an apparent increase in ionized calcium-binding adapter molecule (Iba1) and GFAP immunoreactivity, as well as increased numbers of TUNEL-positive cells in the VHC. Our findings demonstrate that experimental conditions, particularly the exposure to blast acoustics, can increase anxiety and trigger specific behavioral and molecular changes without injury. These findings should be taken into consideration when designing bTBI studies, to better understand the role of stressors in the development of post-traumatic symptoms, and to establish a differential diagnosis for PTSD and bTBI.
Introduction
P
Earlier works have shown that bTBI is a specific form of neurotrauma (Cernak et al., 2001a,2001b, 2011; Chavko et al., 2011; Pun et al., 2011; Risling et al., 2010). Neurobehavioral changes often result from bTBI and share many PTSD symptoms like altered anxiety levels and memory impairment (Okie, 2005; Ryan and Warden, 2003). Such changes have been frequently observed after survivable bTBI, both moderate and severe. Accumulating evidence shows that even exposure to mild blast overpressure can result in lasting neurobehavioral changes (Elder and Cristian, 2009; Elder et al., 2010; Rosenfeld and Ford, 2010).
Exposure to repeated mild blast overpressure poses special diagnostic challenges since there are typically no visible injuries (Belanger et al., 2007; Brenner et al., 2009), and acute neurobehavioral assessments indicate no significant impairment. Soldiers return to their duties and many are deployed numerous times. The consequences of repeated exposure to mild explosive blasts are an especially pressing issue for the military healthcare system for several reasons. These include the sheer number of soldiers exposed to repeated blasts, the apparent cumulative effect of the exposures, and the delayed onset of neurobehavioral symptoms (Rosenfeld and Ford, 2010).
Repeated exposure to low levels of blast overpressure can lead to repeated mild bTBI (mbTBI; Elder and Cristian, 2009; Elder et al., 2010; Rosenfeld and Ford, 2010). Repeated mbTBI is currently not very well defined because the observed neurobehavioral abnormalities overlap with those of PTSD (Belanger et al., 2007; Brenner et al., 2009; Vasterling et al., 2009). Importantly, it appears that there is a relationship between the number of exposures to mild explosive blast and the manifestation of neurobehavioral symptoms like anxiety (Rosenfeld and Ford, 2010). Increased anxiety suggests damage to specific neuroanatomical structures, including the ventral hippocampus (VHC), the amygdala (AD), and the prefrontal cortex (PFC), brain structures that are also indicated to be affected in PTSD (Jaffee and Meyer, 2009).
The complexity of battlefield environments combined with potential individual variability makes modeling and identifying the physical and psychological impacts of repeated mbTBI challenging. Earlier works have demonstrated that exposure to high-energy impulse noise alone can trigger distinct cellular and molecular changes (Säljö et al., 2001,2002, 2008). Moreover, blast sounds (without direct exposure to high-energy impulse noise) may act as a powerful stressor capable of inducing adverse physiological changes that increase anxiety and alter behavior. This is consistent with soldiers that develop post-traumatic symptoms without sustaining any prior physical injury on the battlefield.
Attempting to mimic field conditions can help us analyze the individual contribution of each environmental factor in the development of neurobehavioral symptoms of repeated mbTBI. In this study, we aimed to control for several experimental variables (i.e., conditions) repeated for 4 consecutive days: Naïve: no experimental manipulation; C I: transportation alone; C II: transportation and anesthesia; C III: transportation, anesthesia, and blast sounds; Injured: all three variables plus exposure to mild levels of blast overpressure (Table 1).
The tabulated values for duration, temperature, SPL, blast pressure, and apnea are 4-day averages. Anesthesia duration and concentration, distance travelled, and frequency are exact. Each group was exposed to a different combination of the described experimental variables. All groups (except Naïve animals) were exposed to their designated conditions daily for 4 consecutive days.
psi, pounds per square inch; TBI, traumatic brain injury; dB, decibel.
Each of these experimental variables can potentially play a role in the observed behavioral, cellular, and molecular changes. Soldiers are exposed to various stressors, that include transportation (with the potential danger of roadside bomb detonation), heat, and the acoustic cues of blast with or without injury (Jaffee and Meyer, 2009). On the other hand, anesthesia (e.g., by isoflurane) has been shown to be a neuroprotective (Kurz et al., 1997; Xiong et al., 2003), and in turn a confounding factor in studies that include ischemia.
We found that repeated exposure to standard experimental conditions, particularly the acoustic cues of blast, can significantly increase anxiety and trigger cellular and molecular changes, even without injury. These findings suggest that the consequences of multiple blast exposure are influenced by numerous factors; direct exposure to blast overpressure is only one of them. These findings need to be taken into consideration when designing, analyzing, and interpreting bTBI studies for their implications in the overlapping pathologies of psychological stress and blast-induced physical injury.
Methods
Animals and housing conditions
A total of 34 male Sprague-Dawley rats weighing 245–265 g (Charles River Laboratories, Wilmington, MA) were used. The rats were housed 2 per standard rat cage with free access to food and water in a reverse 12-h/12-h light-dark cycle. The animals were handled according to a protocol approved by the Institutional Animal Care and Use Committee (IACUC) at the Uniformed Services University (USU). Behavioral testing was conducted during animals' dark cycle.
Injury and experimental conditions
After 5 days of acclimation and handling, all animals except Naïve rats (n=6) were transported daily for 4 consecutive days from USU (Bethesda, MD), to Walter Reed Army Institute of Research (WRAIR, Silver Spring, MD), and exposed to the various experimental conditions (Table 1). For the duration of the study Naïve animals were kept in the animal facility at USU with a constant room temperature of 69°F and 68% humidity; the average sound pressure level (SPL) was measured at 50±10% dB. Normal and peak SPL were measured using a Precision Impulse Integrating Sound Level Meter (Model 1800; Quest Electronics, Oconomowoc, WI). Condition I animals (C I, n=6) were transported to WRAIR 2 per cage, but were neither anesthetized nor exposed to the acoustic cues of blast. The rats remained in the vehicle (with food and water ad libitum) and were not brought into the facility; the temperature inside the vehicle ranged from 85–92°F, humidity was 54–88%, and SPL was 47–75 dB. Condition II animals (C II, n=6) were transported and anesthetized, but not exposed to the acoustics of blast. The rats were placed in an induction chamber and anesthetized for 6 min with 4% isoflurane (Baxter Healthcare Corporation, Deerfield, IL). Following anesthesia, C II rats were removed from the facility prior to any blast exposures and returned to the vehicle; temperature in the holding room was 73°F, humidity was 62%, and SPL was 55±10% dB. Condition III animals (C III, n=8) were transported, anesthetized, and exposed to blast sounds. Blast sounds were generated by the blast tube during Injured animals' exposure to mild blast. C III rats were kept in the adjacent holding room for the length of the Injured animals' exposures. Peak SPL during blast overpressure was measured at 95±10% dB. Injured animals (n=8; weight ∼300 g) were transported, anesthetized, and exposed to blast sounds, as well as blast overpressure using a compression-driven shock tube (Long et al., 2009). Following anesthesia the rats were placed in the shock tube holder in a transverse prone position and exposed to whole-body blast overpressure while wearing chest protection. Blast procedures were conducted as previously described (Kovesdi et al., 2011). The duration of apnea was measured for Injured animals, after which they were returned to their cages. After completion of the blast exposures, all animals were transported back to the USU animal facility.
Behavioral test: Elevated plus maze (EPM)
On the last day of the exposures, basic motor activity and anxiety were measured with an EPM (Kovesdi et al., 2011; Kwon et al., 2011). Briefly, the rats were placed one by one in the center of the maze facing one of the open arms; each animal was allowed to explore freely for 5 min while its movement was video tracked. Total distance travelled (meters) and time spent (seconds) in the center and in each arm was recorded using ANY-maze 4.2 Software (Stoelting Company, Wood Dale, IL).
Tissue collection and processing
One day after behavioral testing, the animals were placed inside an induction chamber saturated with isoflurane and deeply anesthetized. For the reverse phase protein microarray (RPPM) and enzyme-linked immunosorbent assays (ELISA; Naïve=4, C I=4, C II=4, C III=5, and Injured=5), the rats were decapitated and the brains were immediately removed and placed in ice. The AD, PFC, and VHC were then dissected, flash frozen in dry ice, and stored at −80°C.
For histology (Naïve=2, C I=2, C II=2, C III=3, and Injured=3), the rats were placed under deep isoflurane anesthesia and transcardially perfused with cold phosphate-buffered saline (PBS) followed by 4% paraformaldehyde solution. The fixed brains were then immersed in cold 15% and 30% sucrose in 1×PBS (consecutively) for cryoprotection. The frozen brains were sectioned coronally at 20 μm thickness using a cryostat (Cryocut 1800; Leica Microsystems, Bannockburn, IL), and sections containing the VHC were kept at −80°C until use.
Protein measures
Preparation of samples
Sample preparation, printing, scanning, and data analysis were performed using RPPM technology as previously described (Gyorgy et al., 2010). Frozen brain sections were immersed in 500 mL T-PER solution with protease and phosphatase inhibitors and sonicated on ice. Serum samples were centrifuged for 15 min at 4°C, and the supernatants were aliquotted and stored at −80°C. Tissue samples were diluted in print buffer to 1 mg/mL, and then diluted in an 11-step dilution in 96-well plates. The samples and dilutions were added to Genetix 384-well plates (X7022; Fisher Scientific, Pittsburgh, PA) using a JANUS Varispan Integrator and Expanded Platform Workstation (PerkinElmer, Waltham, MA), which was programmed to add the samples in a specific pattern for the Aushon Arrayer (Aushon Biosystems, Billerica, MA). The plates were added to the Aushon Arrayer and printed on single nitrocellulose pad slides.
Printing parameters
The Aushon Arrayer was programmed to use 16 pins (4×4 pattern). The 384-plate sample configuration and Aushon program were set to print the 12-point dilution in a triplicate 3×12 block. Tissue samples were printed two touches per spot, while serum samples were printed with one touch.
Immunochemical detection
Primary antibodies were diluted to 10×the optimal Western blot analysis concentration in antibody incubation buffer. Primary antibodies were diluted to the recommended concentrations: vascular endothelial growth factor (VEGF) 1:100 and glial fibrillary acidic protein (GFAP) 1:5000, and then incubated overnight at 4°C with a cover-slip. On the following day the slides were washed and then incubated with Alexa Fluor® 635 goat anti-mouse (cat. no. A-31574), goat anti-rabbit (cat. no. A-31576), or rabbit anti-goat IgG (cat. no. A-21086) secondary antibodies from Invitrogen (Carlsbad, CA) at 1:6000 dilution in antibody incubation buffer for 1 h at room temperature. After drying, the slides were scanned and data were imported into a bioinformatics program. More information on the primary and secondary antibodies can be found in detail elsewhere (Gyorgy et al., 2010).
Data analysis
Data from the scanned images were imported into a Microsoft Excel-based bioinformatics program developed in house for analysis (Gyorgy et al., 2010). The tool calculates the net intensity of the primary antibody compared to the secondary antibody, and graphs it in a linear logarithmic regression format as previously described (Gyorgy et al., 2010).
Corticosterone (CORT) assay
CORT levels were measured using Cayman's Corticosterone Enzyme Immunoassay (EIA) Kit according to the manufacturer's instructions (Cayman Chemical, Ann Arbor, MI). Each sample was diluted 1:500 and measured in triplicate.
Interleukin-6 (IL-6) and interferon-γ (IFN-γ) assays
IFN-γ and IL-6 levels were measured from brain tissues using the rat interferon-γ ELISA and the rat IL-6 ELISA kits (both from Thermo Fisher, Waltham, MA). The assays were performed according to the manufacturer's instructions.
Histology
Immunohistochemistry
Immunohistochemical stainings were performed as previously described (Kwon et al., 2011). Frozen sections containing the VHC were selected and incubated in mouse anti-GFAP (1:400; Millipore, Temecula, CA) and goat anti-Iba1 (1:1000, ionized calcium-binding adapter molecule; Abcam, Cambridge, MA) overnight at 4°C. After washing with 1×PBS, the sections were incubated with corresponding secondary antibodies (1:100 Alexa Fluor 488 goat anti-mouse IgG and 1:100 Alexa Fluor 488 donkey anti-goat IgG; Invitrogen) for 1 h at room temperature. The sections were washed with 1×PBS and cover-slipped using anti-fading media (Vectashield; Vector Laboratories, Burlingame, CA).
TUNEL assay
The extent of DNA fragmentation was determined using a terminal-deoxy-transferase-mediated dUTP nick-end labeling (TUNEL) in situ cell death detection kit, POD (Roche, Indianapolis, IN), according to the manufacturer's instructions and as previously described (Kwon et al., 2011). The TUNEL reaction was performed for 1 h at 37°C; TUNEL-positive cells were visualized by 3,3′-diaminobenzidine (DAB) substrate, and were counted from four sections per animal.
Histological data acquisition
The sections were viewed in an Olympus IX-71 microscope, and images were collected using a SPOT digital camera (Diagnostic Instruments Inc., Sterling Heights, MI), and processed as previously described (Kwon et al., 2011).
Statistical analysis and data comparison
Behavioral and proteomics results were analyzed with analysis of variance (ANOVA) and Tukey post-hoc tests. TUNEL histology results were analyzed with a two-tailed Student's t-test. Statistical analyses were performed using Graph Pad Instat software (GraphPad Software, San Diego, CA). Data are reported as the average±standard error of the mean. For each of our numerical measurements, we determined whether the differences among experimental groups were statistically significant (p<0.05 is depicted by one asterisk, p<0.01 by two asterisks, and p<0.001 by three asterisks).
Results
Basic motor function and anxiety
We found that animals in experimental groups C I, C II, and C III travelled somewhat shorter distances than Naïve rats as measured in the EPM. The differences among the three conditions were not statistically significant when compared to Naïve rats or to one another. On the other hand, Injured animals travelled significantly shorter distances than Naïve rats (Fig. 1A).

Anxiety and basic motor function of the animals in the various experimental groups. An elevated plus maze (EPM) was used to measure changes in basic motor function and anxiety levels. (
C I, C II, and C III animals spent significantly less time in the open arms of the maze compared to Naïve rats. The differences between the three experimental conditions were not significant. Injured animals also spent less time in the open arms of the maze compared to Naïve rats, but the difference was not statistically significant (Fig. 1B).
All experimental groups spent more time in the closed arms of the maze than Naïve rats. Injured animals spent the longest time in the closed arms. Due to substantial variations among animals there were no statistically significant differences among any of the experimental groups (Fig. 1C).
We also measured the time the animals spent in the central area of the maze. We saw a trend that somewhat mirrored what we observed in the closed arms; Injured animals spent the least amount of time in the center compared to Naïve or C I rats (Fig. 1D).
Serum corticosterone levels
Serum CORT levels of C I and C II rats were not significantly different from those measured in Naïve rats (Fig. 2). However, C III and Injured animals had significantly increased CORT levels, more than 2× and ∼ 2.5× higher than Naïve rats, respectively.
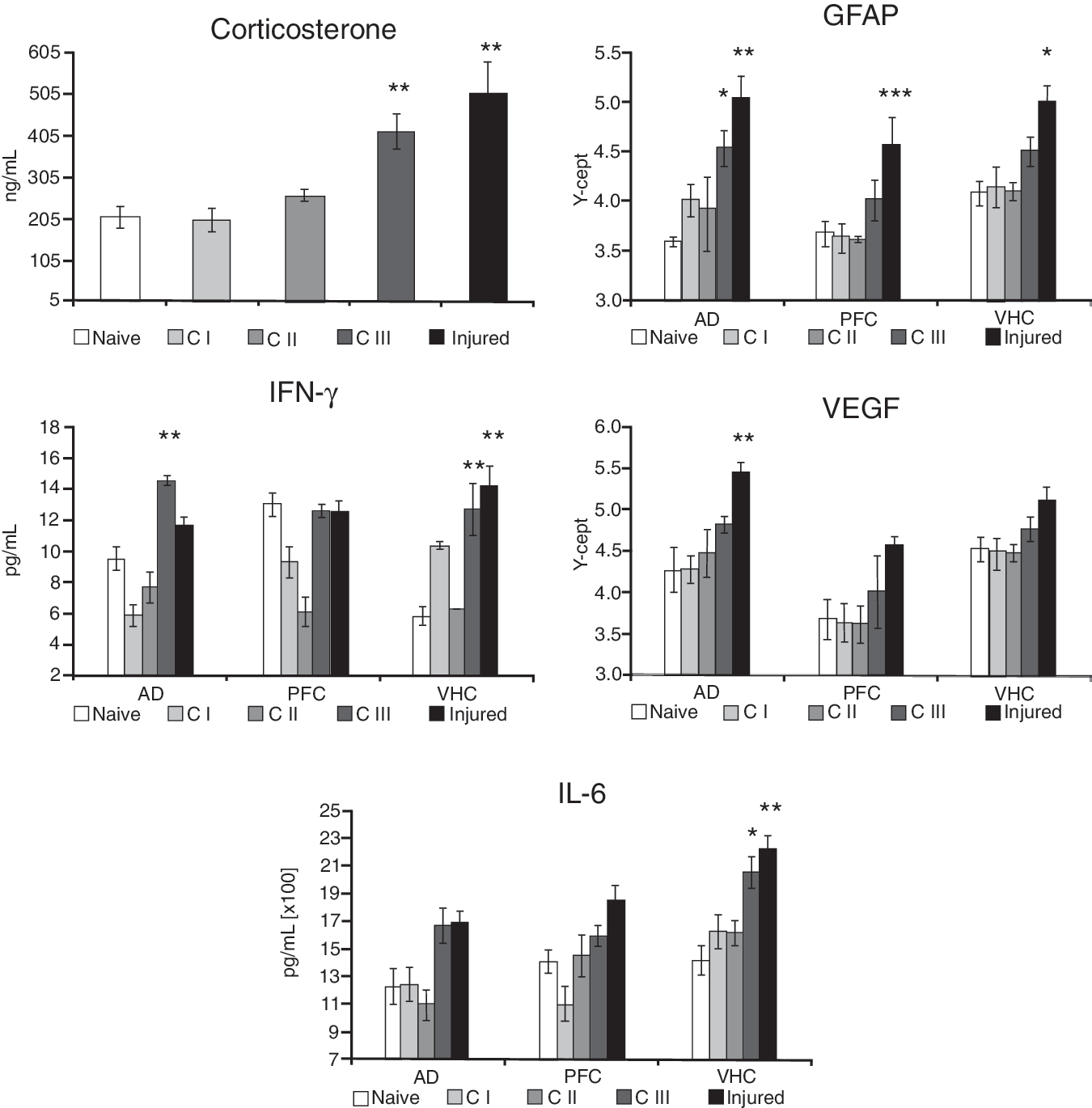
Protein markers in the AD, the PFC, and the VHC of animals in the various experimental groups. Tissue extracts were prepared from dissected brain regions of Naïve, C I, C II, C III, and Injured rats. Tissue levels of GFAP and VEGF were assayed using RPPM; IL-6, IFN-γ, and corticosterone using ELISA. The Y-axis intercept (Y-cept) and pg or ng/mL indicate measured protein levels (*p<0.05, **p<0.01, and ***p<0.001; data are presented as mean±standard error of the mean; IFN-γ, interferon-γ; IL-6, interleukin-6; GFAP, glial fibrillary acidic protein; VEGF, vascular endothelial growth factor; AD, amygdala; VHC, ventral hippocampus; PFC, prefrontal cortex; RPPM, reverse phase protein microarray; ELISA, enzyme-linked immunosorbent assay).
Protein markers in select brain regions
GFAP levels were significantly higher in all measured brain regions of Injured animals only, except for C III animals, which also had significantly elevated GFAP tissue levels in the AD compared to Naïve animals (Fig. 2).
We found the highest levels of IFN-γ in the AD and the VHC of Injured and C III animals (Fig. 2); IFN-γ levels in the same brain regions of C I and C II animals were not significantly different from those of Naïve animals. Interestingly, IFN-γ levels in the PFC of C I and C II rats were much lower than the values in the Injured and C III animals.
VEGF showed a similar trend to GFAP levels across all experimental groups, with VEGF values being highest in the brains of Injured animals and lowest in Naïve animals (Fig. 2). VEGF levels were only significantly elevated in the AD of Injured animals.
IL-6 values were significantly higher in the VHC of the Injured and C III groups compared to Naïve animals (Fig. 2). We found that IL-6 levels in the AD and the PFC showed a somewhat similar trend, but the differences in IL-6 tissue levels of C III and Injured rats were not statistically significant.
Cellular changes
We saw elevated Iba1 immunoreactivity in the VHC of Injured rats (Fig. 3A). Of the other experimental groups, only C II animals displayed a slight increase in Iba1 immunoreactivity, but to a lesser degree than in the Injured animals.

(
The changes we observed using GFAP immunohistochemistry were similar to those measured by proteomics in the VHC. We found an apparent increase in GFAP immunoreactivity in the VHC of Injured animals (Fig. 3B). Interestingly, GFAP expression also appeared to be slightly elevated in the VHC of C III animals.
We assessed changes in the number of TUNEL-positive cells in the hilus and the granular cell layer (GCL) of the VHC. We found that the number of TUNEL-positive cells was increased significantly in the GCL of the VHC of Injured animals only (Fig. 4A). Interestingly, the animals in groups C I and C III had lower numbers of TUNEL-positive cells in the hilus of the VHC.
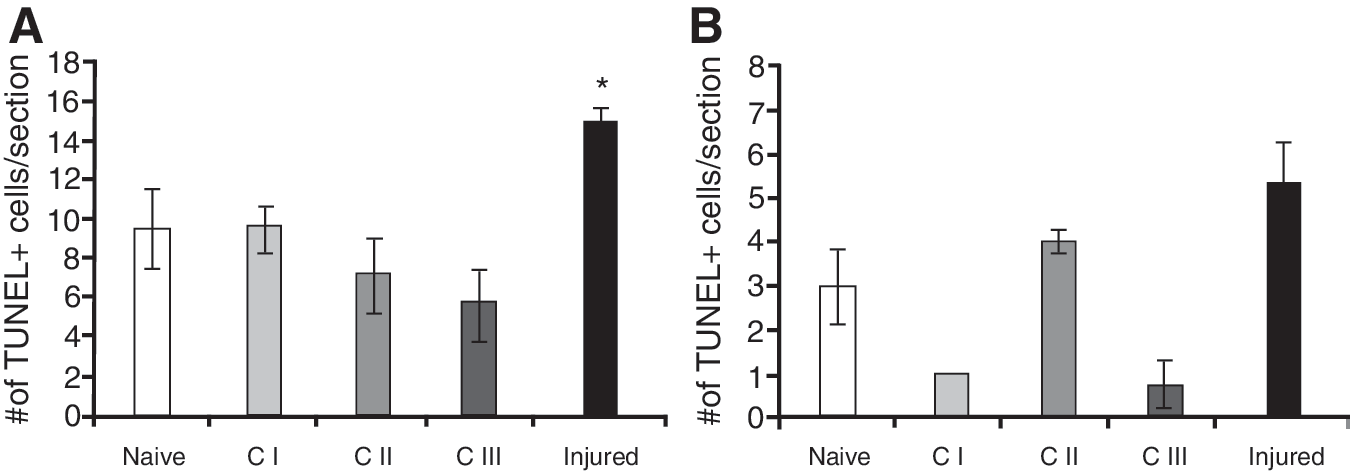
TUNEL apoptotic cells in the GCL (
Discussion
Standard experimental conditions that model select battlefield environments, particularly repeated exposure to the acoustic cues of blast, can significantly increase anxiety and induce cellular and molecular changes, even without injury. However, the observed gliotic response and significant cell death is caused by repeated exposure to mild blast overpressure. These findings support and extend our previous observations about the complexity of factors affecting and/or contributing to the outcome of bTBI (Kwon et al., 2011). At the experimental level, our findings highlight the importance of using appropriate sham groups when designing bTBI studies. We believe that our findings will contribute to a better understanding of the similarities and differences between the pathobiologies associated with environmental stressors and blast-induced physical injury.
Stressors like daily travel, handling, and repeated exposure to the acoustic cues of blast (without injury) significantly increase anxiety. Elevated anxiety may in turn increase vulnerability to and/or worsen the outcome of physical injury (e.g., explosive blast). We have shown earlier that exposing chronically stressed animals to mild blast overpressure prolongs the period of increased anxiety (Kwon et al., 2011). Although the contribution of repeated stress and mild blast exposure to the development of PTSD-like symptoms is currently unknown, repeatedly exposing animals to the acoustic cues of blast can increase serum CORT levels almost as significantly as when combined with blast injury. Abnormal serum CORT levels can alter the inflammatory response (Garcia-Bueno et al., 2008; Miller et al., 2009; Sorrells et al., 2009).
IL-6 and IFN-γ were both significantly elevated in the VHC and the AD of animals repeatedly exposed to the acoustic cues of blast without injury (C III). Repeated exposure to mild blast further increased serum CORT as well as brain IL-6 and IFN-γ levels; these changes corresponded very well with the measured functional outcome (i.e., increased anxiety). In addition to the VHC, the AD is a major neuroanatomical substrate of anxiety (Bannerman et al., 2004; Holscher, 2003). However, the relationship between IL-6 and IFN-γ levels in these brain regions and neurotransmitter systems is not clear at this point (Blalock, 1989; Haddad et al., 2002). Elevated IL-6 levels may contribute to increased cell death due to its neurotoxic effect (Lenzlinger et al., 2001; Stoll et al., 2000).
We found significantly increased numbers of TUNEL-positive cells in the VHC of Injured animals, but not in the VHC of C III animals. This is consistent with our previous finding that blast injury triggers a significant increase in the number of TUNEL-positive cells, while chronic stress has no significant effect (Kwon et al., 2011). In addition to an increase in DNA fragmentation, our histology indicated that the increase in GFAP immunoreactivity is another pathological change specific to injury. Increased GFAP immunoreactivity, combined with a stellar morphology, is a hallmark of practically all TBIs (Eng and Ghirnikar, 1994; Fitch and Silver, 2008).
We previously hypothesized that elevated tissue levels of VEGF are another injury-specific marker (Agoston et al., 2009; Kwon et al., 2011). In this experiment, we only found elevated tissue levels of VEGF in the VHC, PFC, and AD of Injured animals; the rats in all other experimental conditions had tissue levels similar to the VEGF levels measured in Naïve rats. A key factor in increasing VEGF levels is hypoxia; Injured animals sustained a moderate period of apnea, lasting for 15 sec on average. The resulting hypoxia may be responsible for the increase in VEGF levels and the downstream pathological cascade that potentially distinguishes stress from injury associated with apnea. VEGF is a multifunctional molecule (Rosenstein and Krum, 2004) involved in mediating various pathological processes, including increased vascular permeability following TBI (Agoston et al., 2009; Skold et al., 2005).
Increased membrane permeability, early gene induction in certain brain regions, and possible microglia activation have been shown to result from a single exposure to impulse noise at around 200 dB (Säljö et al., 2001,2002,2003). It is important to note that in our experimental set-up, C III and Injured animals were not exposed to such high intensity noises (Table 1); the acoustic cues C III and Injured animals were exposed to likely act as additional stressors rather than direct pressure wave energy experienced as impulse noise (Dancer et al., 1998). As discussed above, we found significantly elevated levels of IL-6 and IFN-γ in the VHC of C III animals. Similarly, serum CORT levels were significantly elevated in the same group of animals. Our findings suggest that repeated exposure to the acoustic cues of blast may serve as an effective stressor that triggers distinct neurobehavioral changes implicated in the development of post-traumatic symptoms, and possibly shell-shock or PTSD in soldiers (Guy, 2004; Rosenfeld and Ford, 2010).
Although the detected functional change was not matched by changes in tested cellular and molecular markers, transportation alone had a significant effect on anxiety in C I animals. In a military blast environment, travel in potentially dangerous areas adds a considerable amount of stress on soldiers. Experimentally, confounding factors (e.g., anesthesia) associated with animal studies make it more challenging to ascertain the exact pathological and symptomatic contribution of stress and injury.
Anesthetics like isoflurane can exert complex neuroprotective effects on the injured brain (Xiong et al., 2003). Moreover, repeated exposure to isoflurane can provide significant dose-dependent neuroprotection against the effects of ischemia, including reduced neuronal death (Zhang et al., 2010). However, in our current study, all animals except Naïve animals were exposed to the same duration, concentration, and frequency of anesthesia, thus ruling out anesthesia as a major confounding factor. Similarly, the relative contribution of the transportation stress is equal across all experimental groups.
In summary, we found that repeated exposure to standard experimental conditions, particularly the acoustics of blast, increases anxiety and triggers specific cellular and molecular changes. Importantly, it is the direct exposure to blast overpressure that triggers the gliotic response and cellular death. These findings need to be taken into account when designing, analyzing, and interpreting bTBI studies. When independently verified, our findings will help in the development of tools to distinguish between psychological and physical traumas and their overlapping pathologies as evidenced in bTBI and PTSD.
Footnotes
Acknowledgments
We thank the Neurotrauma Team (WRAIR) for their technical help during the exposures. This work was supported by Center for Neuroscience and Regenerative Medicine (CNRM) grant no. G1703F.
Author Disclosure Statement
The views, opinions, and/or findings contained herein are those of the authors and should not be construed as an official position, policy, or decision of the Department of the Army or the Department of Defense. The authors have no financial disclosures.
Animal handling and treatments were conducted in compliance with the Animal welfare Act and other Federal statutes and regulations related to animals and experiments involving animals, and adhered to principles stated in the Guide to the Care and Use of Laboratory Animals, National Research Council. The facilities are fully accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International.