
Abstract
A mouse model of repeated blast exposure was developed using a compressed air-driven shock tube, to study the increase in severity of traumatic brain injury (bTBI) after multiple blast exposures. Isoflurane anesthetized C57BL/6J mice were exposed to 13.9, 20.6, and 25 psi single blast overpressure (BOP1) and allowed to recover for 5 days. BOP1 at 20.6 psi showed a mortality rate of 2% and this pressure was used for three repeated blast exposures (BOP3) with 1 and 30 min intervals. Overall mortality rate in BOP3 was increased to 20%. After blast exposure, righting reflex time and body-weight loss were significantly higher in BOP3 animals compared to BOP1 animals. At 4 h, brain edema was significantly increased in BOP3 animals compared to sham controls. Reactive oxygen species in the cortex were increased significantly in BOP1 and BOP3 animals. Neuropathological analysis of the cerebellum and cerebral cortex showed dense silver precipitates in BOP3 animals, indicating the presence of diffuse axonal injury. Fluoro-Jade B staining showed increased intensity in the cortex of BOP3 animals indicating neurodegeneration. Rota Rod behavioral test showed a significant decrease in performance at 10 rpm following BOP1 or BOP3 at 2 h post-blast, which gradually recovered during the 5 days. At 20 rpm, the latency to fall was significantly decreased in both BOP1 and BOP3 animals and it did not recover in the majority of the animals through 5 days of testing. These data suggest that repeated blast exposures lead to increased impairment severity in multiple neurological parameters of TBI in mice.
Introduction
R
Explosive blast injury is described by four causative categories: primary, secondary, tertiary, and quaternary, based on physical features of the blast episode (CDC, 2006; Owen-Smith, 1982; Wolf et al., 2009). Primary blast injury is caused by the shockwave; secondary blast injury is caused by high velocity shrapnel and debris; tertiary blast injury is from blast wind and the resulting body acceleration, displacement, and impact; and quaternary blast injury is from heat and toxic smoke exposure associated with explosive detonations. The severities of bTBIs mainly depend upon strength of the explosive, distance from the exposure, reflection of the waves (Mellor, 1988; Wolf et al., 2009), open field, and presence of buildings or vehicles at the explosion sites (Leibovici et al., 1996). Because it is difficult to develop complex animal models involving all four different phases of blast injuries or intensity of bTBIs encountered in the battlefield, more efforts have been primarily focused on developing individual phase blast injury models (Bell, 2008; Chen et al., 2009). Primary blast injury composed of injuries from the shock front is probably the least understood, and is being explored mostly to determine the spectrum of neurobiological effects and biochemical mechanism of injury (Chen et al., 2009; Nakagawa et al., 2011; Scott et al., 2006).
Pre-clinical models using shock tubes driven by compressed air or helium have been increasingly used in the laboratory to understand the pathophysiology of bTBI (Cernak et al., 1996, 2011; Chavko et al., 2007; Gorbunov et al., 1997; Koliatsos et al., 2011; Leonardi et al., 2011b; Long et al., 2009; Saljo et al., 2009, 2010). More compact, convenient, and cost-effective vertical shock tubes are also being used more and more to study the effects of primary blast on the brain (Connell et al., 2010; Courtney and Courtney, 2010; Park et al., 2011; Svetlov et al., 2010). Experiments with single blast exposure revealed tissue and cellular injury including the most common neuronal, axonal, and glial injuries (Bauman et al., 2009; Cernak, 2010; Cernak et al., 2001a,b, 2011; Nakagawa et al., 2008; Readnower et al., 2010; Saljo et al., 2000, 2001, 2002a,b, 2009, 2010; Suneson et al., 1990; Svetlov et al., 2010). Both necrotic and apoptotic cell death also has been implicated in bTBI (Cernak et al., 2001b; Kato et al., 2007; Nakagawa et al., 2008).
There are limited studies on the neurological effects of repeated blast exposures. Most of the initial repeated blast exposure studies were centered on lung and internal injuries and aimed to establish blast safety thresholds (Richmond, 1991). Also, many of the repeated sub-lethal blast exposure studies are focused on the effects on the auditory system (Ylikoski and Ylikoski, 1994). Repetitive blast exposures in Iraq war veterans with post-concussive symptoms were associated with cerebellar hypometabolism (Peskind et al., 2011). In sheep, repeated blast exposure reduced the threshold of injury to lungs and internal organs (Yang et al., 1996). Using pigs, it has been shown that repetitive occupational levels of blast exposure cause brain injury (Saljo et al., 2008). Pre-clinical rat models with single and repetitive blast exposures showed that multiple blast exposure increase the stress levels in the animals (Mazurkiewicz-Kwilecki, 1980). Studies with sheep showed that 60 repeats of 39.5, 130, or 196 kPa blast overpressure (BOP) result in mild, moderate, or severe injury in lungs, respectively indicating that peak overpressure is an important factor in the severity of injury (Yang et al., 1996). Repeated blast exposure can increase the severity or decrease the threshold of bTBI suggesting that there may be a period of vulnerability after initial blast exposure (Saljo et al., 2010, 2011). On the other hand, analyzing cerebrospinal fluid (CSF) and blood biomarkers, a recent study with repeated exposure to gun blasts or detonation showed no indication of blood–brain barrier damage or TBI in military personnel (Blennow et al., 2011). Therefore, the effects of repeated blast exposures in rendering the brain more vulnerable and increasing the severity of TBI are still controversial. In the United States Marine Corps Dynamic entry or breaching training, Warfighters are repeatedly exposed to as many as 12 explosions (0.3 to 10 lb) per day during training exercise or even more numbers per night during military operation (USAARL Report No. 2010-16,
Studies of blast exposure in mice are limited. Mouse models are of choice when investigating biochemical changes to understand the neurobiological mechanisms of injury, because of the availability of a large repertoire of antibodies, well-characterized genes and proteins, and the availability of genetic variants. Recently, a mouse model of single blast exposure involving mild, moderate, and severe brain injury by different peak overpressures has been reported (Cernak et al., 2011; Koliatsos et al., 2011). The majority of studies with repeated blast exposure have involved very low pressures, and there are no studies to date with repeated blast exposures at higher pressures using a shock tube.
We developed a mouse model of repeated bTBI using a compressed air driven shock tube that was used earlier in the development of a rat model of battlefield bTBI (Long et al., 2009). Animals were exposed to various levels of BOP and percent survival, righting reflex time after blast, body-weight loss, brain edema, neuropathology, and neuromotor functions were measured to assess the level of injury caused by multiple blast exposures. The mouse repeated blast injury model will be a valuable tool to study the mechanism of repeated bTBI and brain vulnerability factors, and for evaluation of therapeutics for protection.
Methods
Animals and blast injury model
All animal experiments were conducted in accordance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals, and adhered to principles stated in the Guide for the Care and Use of Laboratory Animals (National Research Council Publication, 1996 edition) with an Institutional Animal Care and Use Committee approved protocol. Male C57BL/6J mice, 8–10 weeks old, that weighed between 22 and 26 g (Jackson Laboratory, Bar Harbor, ME) were used in this study. Mice were housed at 20–22°C (6:00 to 18:00 h light/dark cycle) with free access to food and water ad libitum. A compressed air-driven shock tube available at the Institute was used for BOP exposure of mice (Long et al., 2009) (Fig. 1A). Mice were anesthetized with 4% isoflurane gas (O2 flow rate 1.5 L/min) for 8 min and quickly placed on a holder in prone position. The animals were restrained with a net to minimize potential movements during blast exposure and related injury (Fig. 1B). The holder was placed 2.5 feet inside from the open end of the 15 foot long expansion chamber, and animals were positioned perpendicular to the direction of the shock waves. BOP exposure was generated by rupture of Mylar membranes (DuPont, Wilmington, DE) of various thicknesses (Long et al., 2009). Immediately after the blast exposure, mice were returned to the cage.
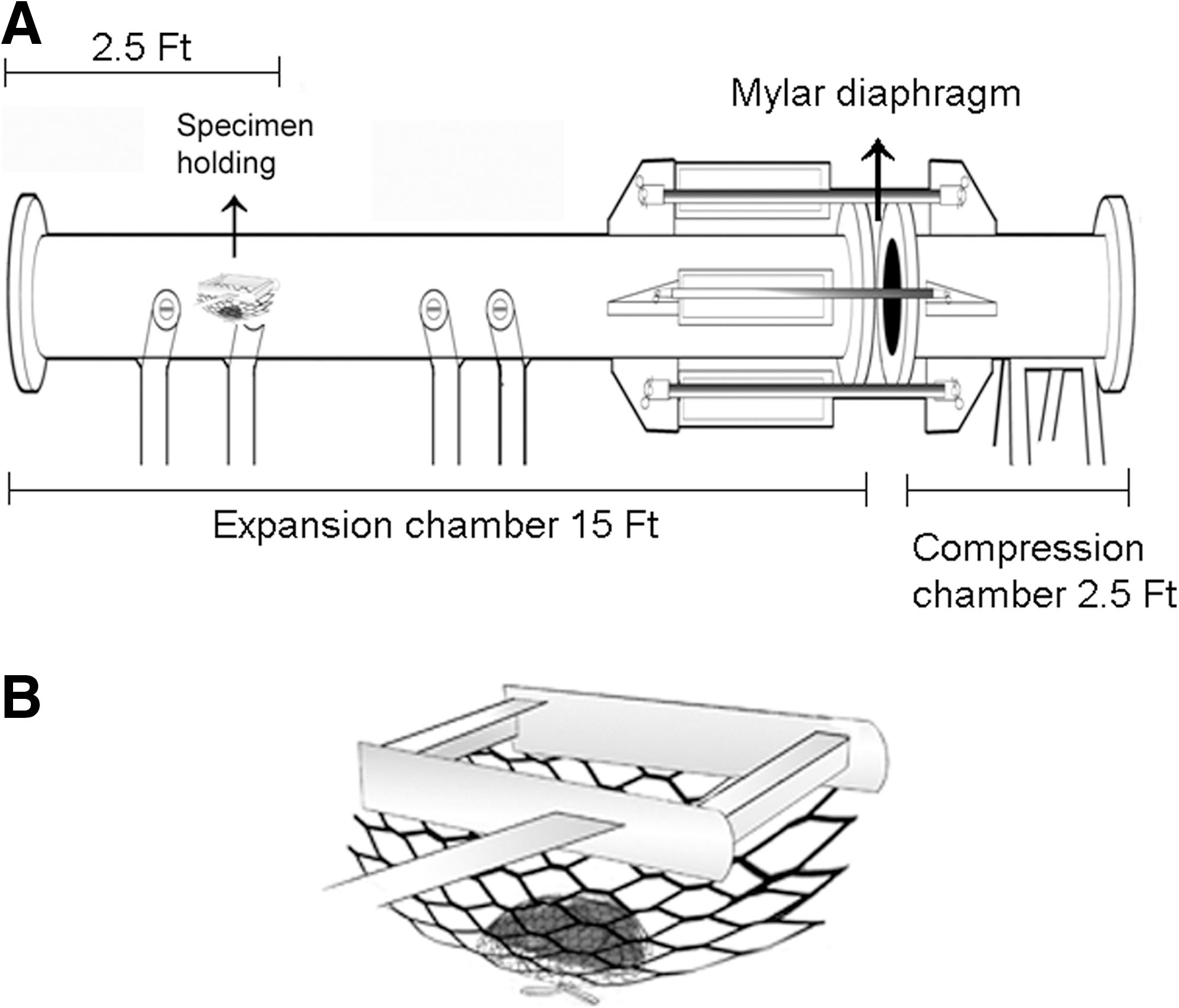
A diagrammatic view of the shock tube and animal restraining for blast overpressure exposure. (
In the pilot study, a total number of 48 mice were used for the single BOP examination. Animals (n=12) were randomly separated into sham control and three different single BOP (13.9, 20.6, and 25 psi) groups. Rupture and static pressure were measured routinely in the laboratory using pressure transducers (Shoge and Long, unpublished data). The static pressure of 20.6 psi was selected for studying the effects of repeated BOP and comparison of the data to single BOP. A total of 210 mice were separated into groups of sham controls (n=60) and BOP groups (n=150). BOP groups involved single BOP (BOP1) (n=43), and repeated BOPs (n=107). For repeated blast exposures, mice were subjected to a second BOP after a 1 min interval, and in some cases to a third BOP (BOP3) 30 min later after additional anesthesia. Sham controls received one or two anesthesia administrations and were handled similarly to blast-exposed mice without the BOP exposure.
Neurological assessment
The righting reflex time was used to measure the length of unconsciousness (Readnower et al., 2010). After anesthesia and blast exposure, the animals were placed on the table in a supine position. The duration of righting reflex was recorded from when the mouse was taken out of the chamber containing 4% isoflurane gas to when the mouse returned to a normal prone position after blast exposure. The animals were also carefully examined for any physical injury caused by blast exposure. Righting reflex time was also measured similarly in sham controls.
Assessment of brain edema
Surviving and paired sham animals were euthanized at 4, 24, or 48 h after the third blast exposure in BOP3 group. The brain was immediately removed and weighed in an analytical balance using pre-weighed aluminum trays to obtain the wet weight. Dry weight of the brain was obtained after drying in an oven at 100°C for 72 h. The percentage of water content of the brain was determined by the formula, (wet weight – dry weight)/wet weight. Brain edema was calculated as the percentage of increase in brain water content (Schoettle et al., 1990).
Measurement of reactive oxygen species (ROS)
Total ROS levels in the cortex of the brains of sham controls and BOP1 and BOP3 animals were measured using 2',7'-dichlorofluorescein (DCF) as described earlier, with minor modifications (Wang and Joseph, 1999a). Briefly, 10 μL of 10% (weight/volume) homogenate of brain cortex prepared in tissue protein extraction reagent (Pierce Chemicals, Rockford, IL) was added to 96-well fluorescence measurement plates containing 200 μL of 5 mM DCF in phosphate buffered saline. The samples were incubated at 37°C for 1 h and the fluorescence was measured at excitation 488 nm and emission 532 nm using a SpectraMax M5 microplate reader (Molecular Devices, Sunnyvale, CA). The fluorescence from each well was captured. The data were expressed as relative fluorescence units/mg tissue.
Neuropathology
Sham control and blast exposed mice were allowed to recover for 6 or 24 h and then deeply anesthetized with pentobarbital (25 mg/kg, i.p.) and transcardially perfused with phosphate buffered saline, followed by 4% paraformaldehyde (PFA), pH 7.4. The brain was removed and immersed in 4% PFA for 6 h, followed by 20% sucrose for 72 h and rapidly frozen in isopentane pre-cooled to −70°C with dry ice. Serial sections were cut coronally from bregma −2.68 mm to −4.84 mm and bregma −5.68 mm to −6.40 mm and were mounted on Superfrost Plus microscope slides. A set of 24 sections (30 μm) was stained with Fluoro-Jade B (FD NeuroTechnologies, Ellicott City, MD). Sections were examined using a fluorescent microscope at excitation 480 nm and emission 525 nm. Fluoro-Jade B labeled degenerating neurons were analyzed by Image-Pro software (MediaCybernetics Inc., Bethesda, MD). Another set of 24 sections (50 μm) was immersed in 0.1 M phosphate buffer containing 4% PFA for 5 days at 4°C and processed with FD Neurosilver™ Kit II (FD NeuroTechnologies, Ellicott City, MD) for the detection of neurodegeneration. Neuronal degeneration was indicated by dense silver precipitates, appearing as black grains when sections were examined under a light microscope with bright field. Silver stained sections were scored based on relative density of silver staining and axonal injury by two experienced pathologists as follows: no damage (-), mild pathology (scattered) (+), moderate pathology (++), and severe pathology (+++).
Assessment of motor function
A microprocessor controlled Rota Rod (Harvard Apparatus, Holliston, MA) was used for motor coordination measurement. Motor function was assessed at 2 h, and 1, 3, and 5 days after single or three repeated blast exposures at 20.6 psi. Before blast exposures, mice were removed from their home cages and placed on the Rota Rod apparatus. With the drums rotating at minimum speed, each animal was placed in its respective position for familiarization with the revolving drum. After three training runs of 3 min at 1 h intervals for 3 days, mice were exposed to BOP3. At indicated time points after blast exposures, mice were tested again in constant speed mode (4 rpm) or in accelerating speed mode (from 1 to 10 and 20 rpm). The sham or BOP- exposed mice were tested three times with 3 min for each trial. The endurance time of each mouse and the latency at which the animal lost its grip and fell were recorded and exported for analysis.
Statistical analysis
Statistical analyses were performed using the GraphPad Prism V4.03 software (GraphPad Software Inc., San Diego, CA). Kaplan–Meier curves and χ2 test were used for the demonstration and analysis of survival rate. One-way ANOVA and Bonferroni's multiple comparison tests were used to calculate the p values. Values were expressed as mean±standard error of the mean. A p-value<0.05 was considered as significant.
Results
Effects of different levels of BOP and repeated blast exposures on mortality
The mortality rates among animals exposed to different levels of BOPs (13.9, 20.6, and 25 psi) are shown in Fig. 2A. Exposures to 13.9 psi BOP (n=12) resulted in no animal death, whereas animals exposed to 20.6 (n=12) and 25 psi (n=12) BOP had 8% and 42% lethality respectively. All the animals that died after blast exposure at 20.6 psi died immediately and there was no delayed death. In contrast, some of the animals exposed to 25 psi BOP died immediately or by 24 and 48 h after blast exposure (Fig. 2A). There was a significant increase in mortality within 48 h after 25 psi BOP compared to sham controls. Animals exposed to different levels of single BOP showed no serious physical signs of injury. A few animals exposed to 20.6 and 25 psi BOP displayed unsteady gait or staggering movements, which lasted for 2 h.
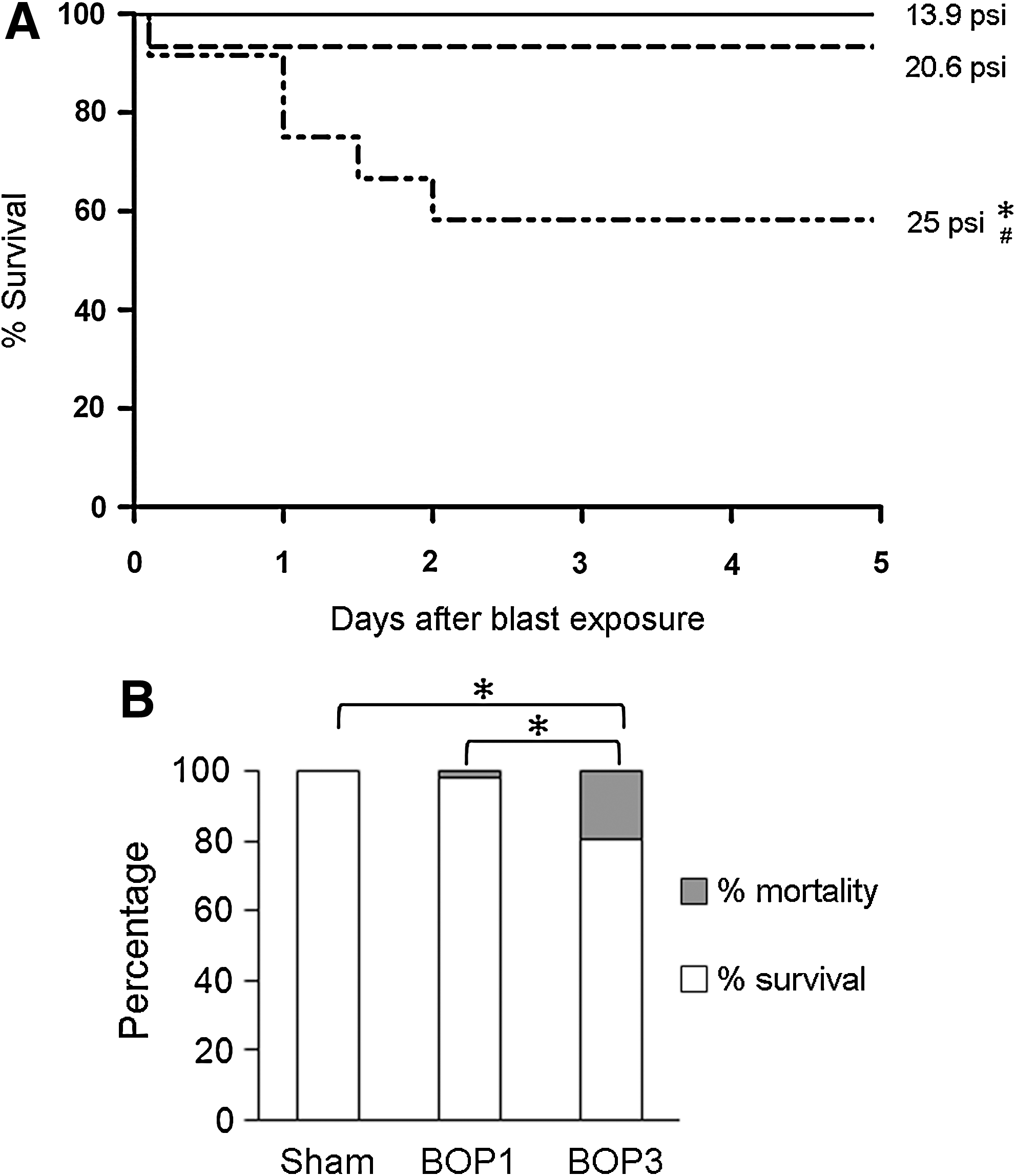
The mortality rate of animals following different levels of single and repeated blast exposures. (
A BOP of 20.6 psi was used for repeated BOP studies and to compare the neurobiological effects with those of a single BOP. A total of 150 mice were subjected to repeated blast exposures. Out of the 150 animals, 3 animals died after BOP1 (2%). A total of 40 surviving animals from BOP1 were used for comparison of the data with three repeated blast exposures. The remaining 107 mice surviving after BOP1 received the second and third blast exposures, which resulted in 21 more deaths (20%). Therefore, the overall mortality was increased from 2% to 20% in BOP3 compared to BOP1 (Fig. 2B). All the animals that died after repeated BOPs at 20.6 psi died immediately after the last blast exposure, and there was no delayed animal death during the 5 days of experiments. Most of the dead animals displayed hemorrhage that included 33% subdural hemorrhage, 21% lung hemorrhage, and 29% abdominal hemorrhage. In surviving animals, 24 h after three repeated blast exposures, 8% of the animals showed subdural hemorrhage and 6% of the animals showed lung injury. Repeated blast exposures did not result in any signs of extremity injuries.
Repeated blast exposures increased the duration of righting reflex time
The righting reflex time of isoflurane-anesthetized animals during recovery after blast exposure is shown in Figure 3. The duration of righting reflex of mice that received isoflurane anesthesia 1 or 2 times during the 30 min interval but no blast exposures (sham 1 and sham 2, respectively) were 103±5, and 102±5 sec, respectively. After a single BOP exposure at 20.6 psi, the righting reflex time was significantly increased to 152±6 sec compared to sham controls. Three repeated blast exposures at 20.6 psi significantly increased the duration of righting reflex to 313±47 sec compared to single BOP and sham controls.
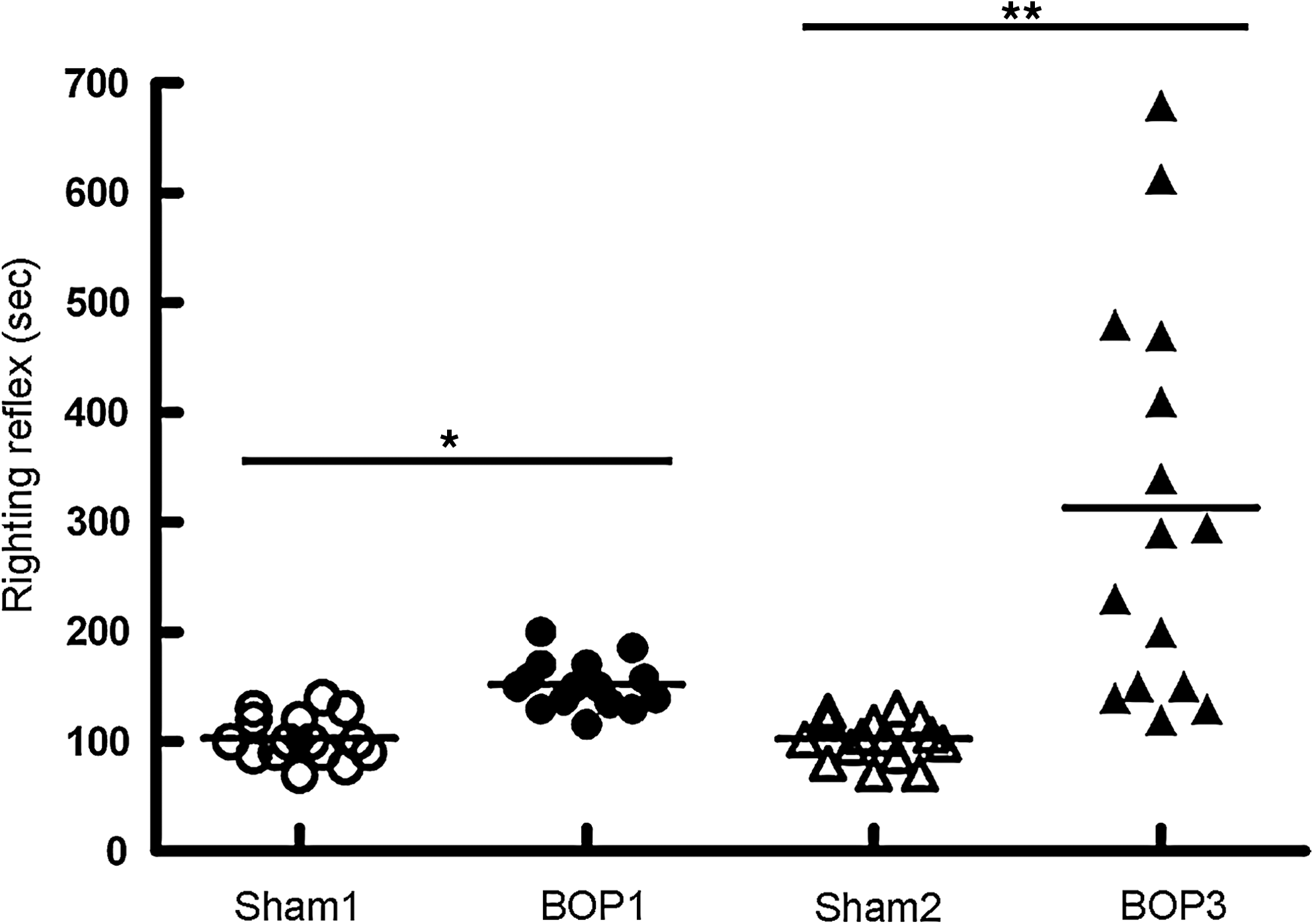
Righting reflex time in repeated blast exposed animals. Mice were exposed to single (BOP1), or three repeated (BOP3) blast exposures at 20.6 psi with an interval of 1 and 30 min. The duration of righting reflex after the last blast was determined and plotted against sham controls that were handled in the same fashion but without blast exposures. *p=<0.01; **p<0.001 (n=20).
Repeated blast exposures increased the body-weight loss in mice
Body-weight loss in sham controls and surviving blast-exposed animals was determined at 24 h intervals after blast exposure up to 5 days. As shown in Figure 4, the body weight was constantly increased over time in sham controls. The sham control animals gained 1.5±0.3% of body weight at 24 h. On the other hand, body weight was decreased in animals exposed to 13.9 psi BOP on days 1 and 2. Animals exposed to 20.6 psi BOP showed 4.1±0.7% body-weight loss on day 1, 1.2±0.7% on day 2, and 0.4±0.3% on day 3. By day 5, the animals exposed to 20.6 psi BOP showed a gain in body weight although it was consistently lower than that seen in sham controls (Fig. 4). Animals exposed to 25 psi BOP showed a loss of 6.74±0.9 % body weight on day 1. The loss of body weight was similar on day 2, but was reduced further on days 3 and 5.
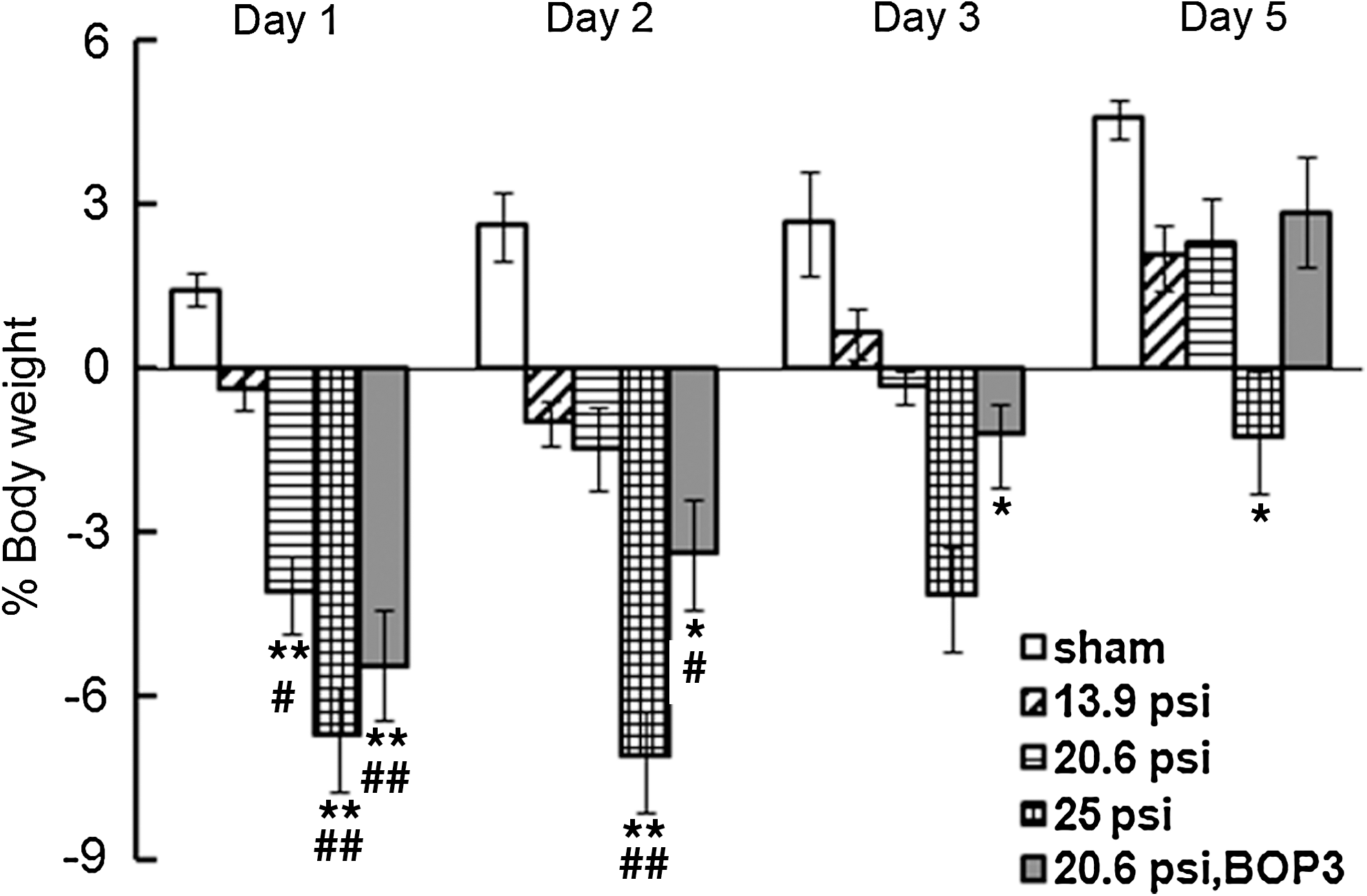
Body weight loss in mice exposed to different levels of single (BOP1) or three repeated (BOP3) blast exposures at 20.6 psi. Mouse body weight was measured before and at days 1, 2, 3, and 5 after 13.9, 20.6, 25 psi BOP1or BOP3 at 20.6 psi. Sham controls showed steady increase in body weight from day 1 to day 5. On the other hand, BOP1 animals at 13.9, 20.6 and 25 psi showed a decrease in body weight on days 1 and 2 that was pressure dependent. On day 5, 13.9 and 20.6 psi blast exposed animals showed a body-weight gain. Animals exposed to 25 psi single blast exposure showed significant loss of body weight from days 1 to 5. Three repeated blast exposed animals at 20.6 psi showed higher body-weight loss than single blast exposed animals at 20.6 psi up to 3 days, but showed body-weight gain on day 5 (n=8-10, *p<0.05 and **p<0.01 compared to sham, # p<0.05 and ## p<0.01 compared to 13.9 psi).
Animals exposed to three repeated blast at 20.6 psi showed higher body-weight loss on days 1, 2, and 3 compared to respective single BOP exposed animals using the same overpressure (Fig. 4). On day 5, repeated blast exposed animals showed a gain in body weight that was similar to single blast exposed animals on day 5 using the same or lower overpressures.
BOP exposure and brain edema
Animals exposed to BOP3 at 20.6 psi showed significant increase in brain water content compared to sham controls at 4 h post-exposure (Fig. 5). However, 24 and 48 h after blast exposures, the brain water content was similar to sham control levels. A pilot study with single blast exposure showed no significant change in brain water content.
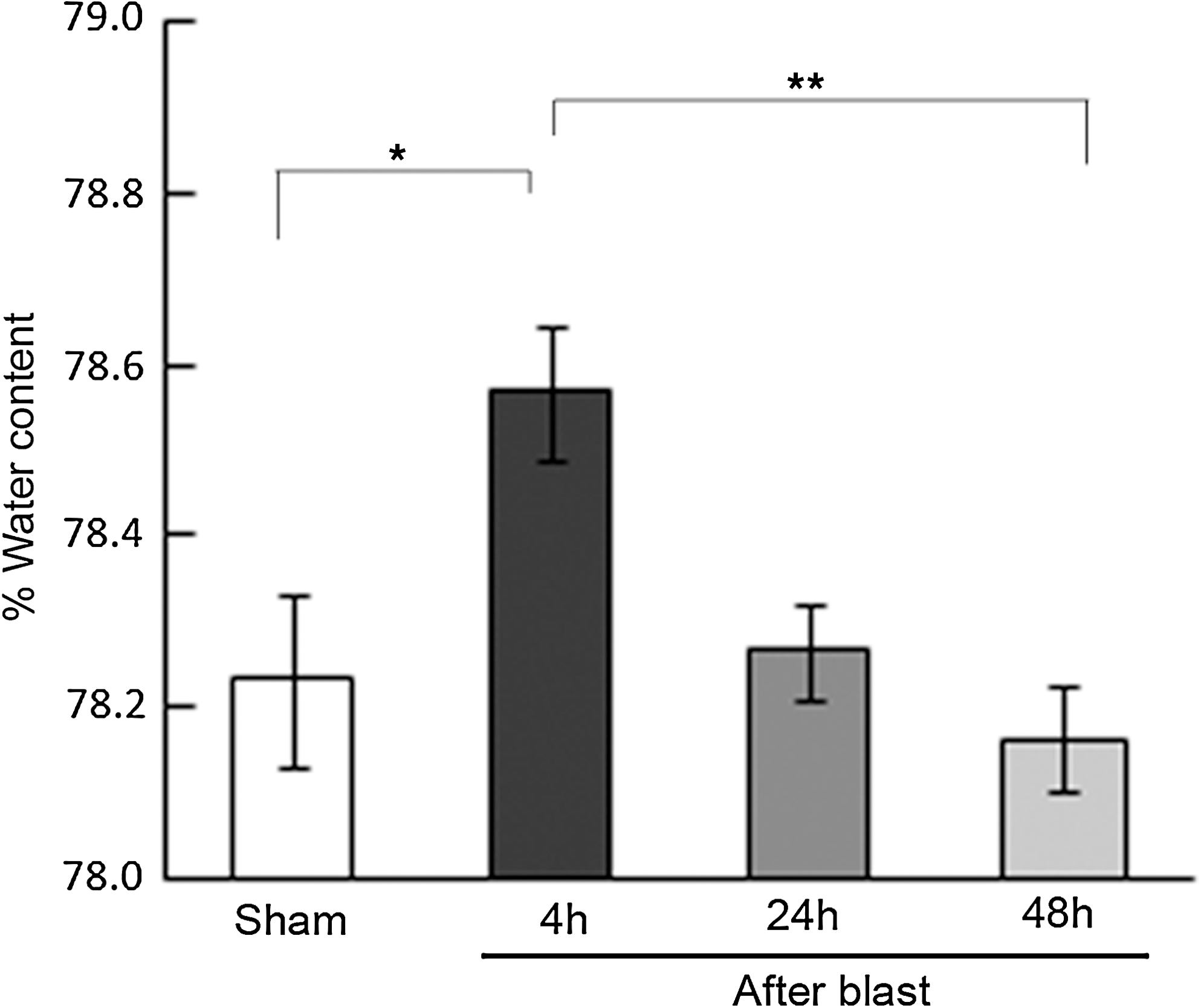
Brain edema in animals exposed to repeated blast overpressures. Histogram showing the percent water content in the brain of sham controls and three repeated 20.6 psi blast exposed animals. The blast exposed animals that were euthanized at 4, 24, and 48 h after three blast exposures are shown. The brain edema at 4 h was significantly increased compared to sham controls and recovered at 48 h (n=8-10, *p<0.05, **p<0.01).
Repeated blast exposure increased ROS
ROS in the cortex of the brain after single or three repeated BOPs at 20.6 psi are shown in Figure 6. ROS was significantly increased at 6 h after single and BOP3 at 20.6 psi compared to sham controls. ROS levels were further increased at 24 h in single and three repeated blast exposed animals. There was no significant difference in the ROS levels between single or three repeated blast exposed animals either at 6 or 24 h.
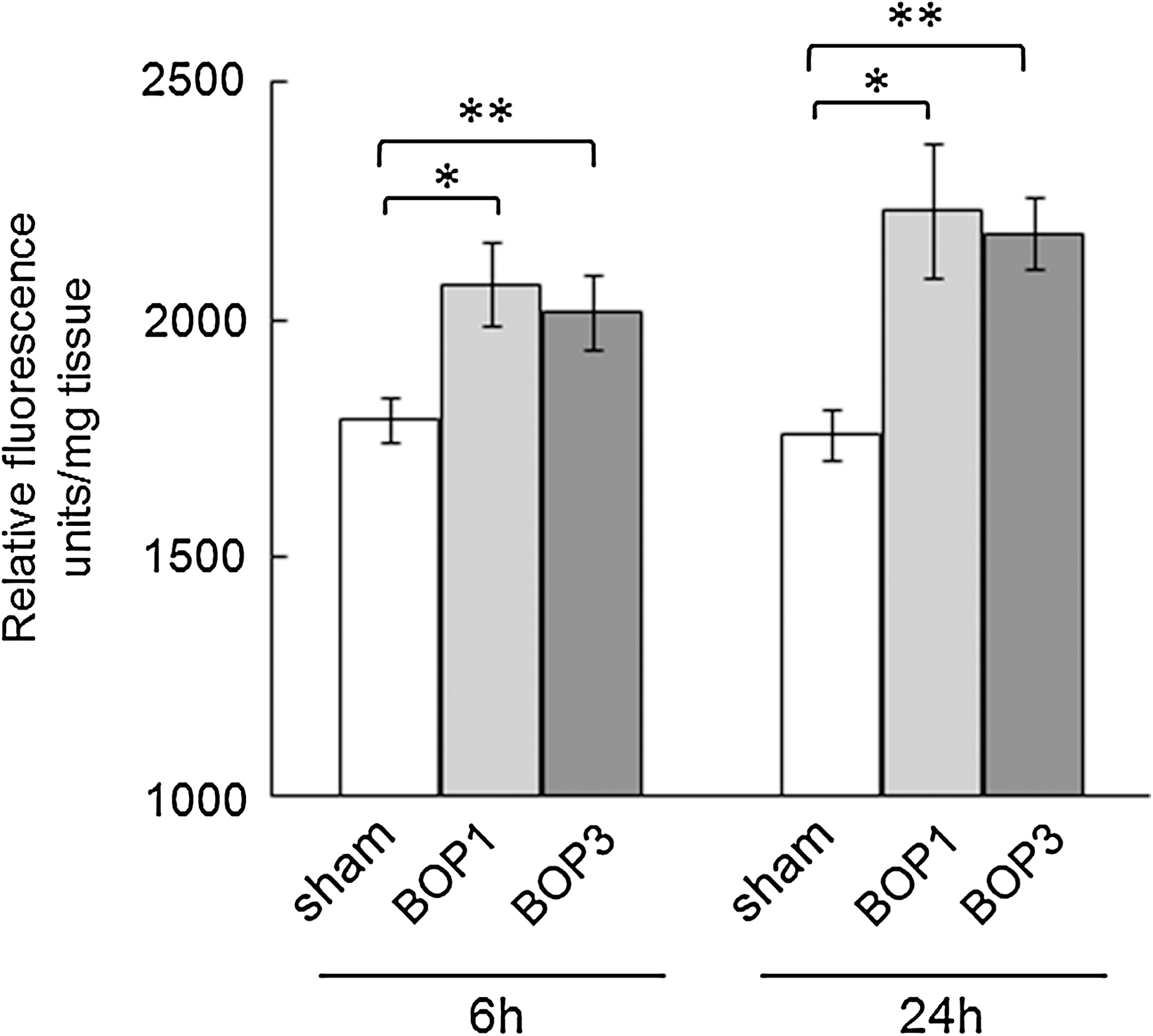
Brain reactive oxygen species (ROS) levels in mice exposed to three repeated blasts. Brain reactive oxygen species were measured as described in the Methods section. Compared to sham controls, brain ROS levels were significantly higher in BOP1 and BOP3 exposed animals at 6 h and 24 h (n=6, *p<0.05, **p<0.01).
Repeated blast exposure increased neuropathology
Silver stained serial brain sections of mice showed severe axonal injury in the BOP3 group compared to the BOP1 and sham groups. At 24 h post-blast, mice that had received BOP1 exposure showed weak stains (scored − to +) in the white matter of the cerebellum (Fig. 7A3) and cortex (Fig. 7B3). Mice that had received BOP3 exposure presented dense silver precipitates (scored ++ to +++) in the cerebellum (Fig. 7A4). Moderate axonal degeneration with scoring of + or ++ was also observed in the basal or piriform cortex (Fig. B4) compared to sham controls (Figs. 7A2, B2).

Histopathology of the brain of blast-exposed mice. Paraformaldehyde fixed brain was sectioned coronally from cerebra through cerebellum and 24 serial sections from each mouse were processed for histopathology.
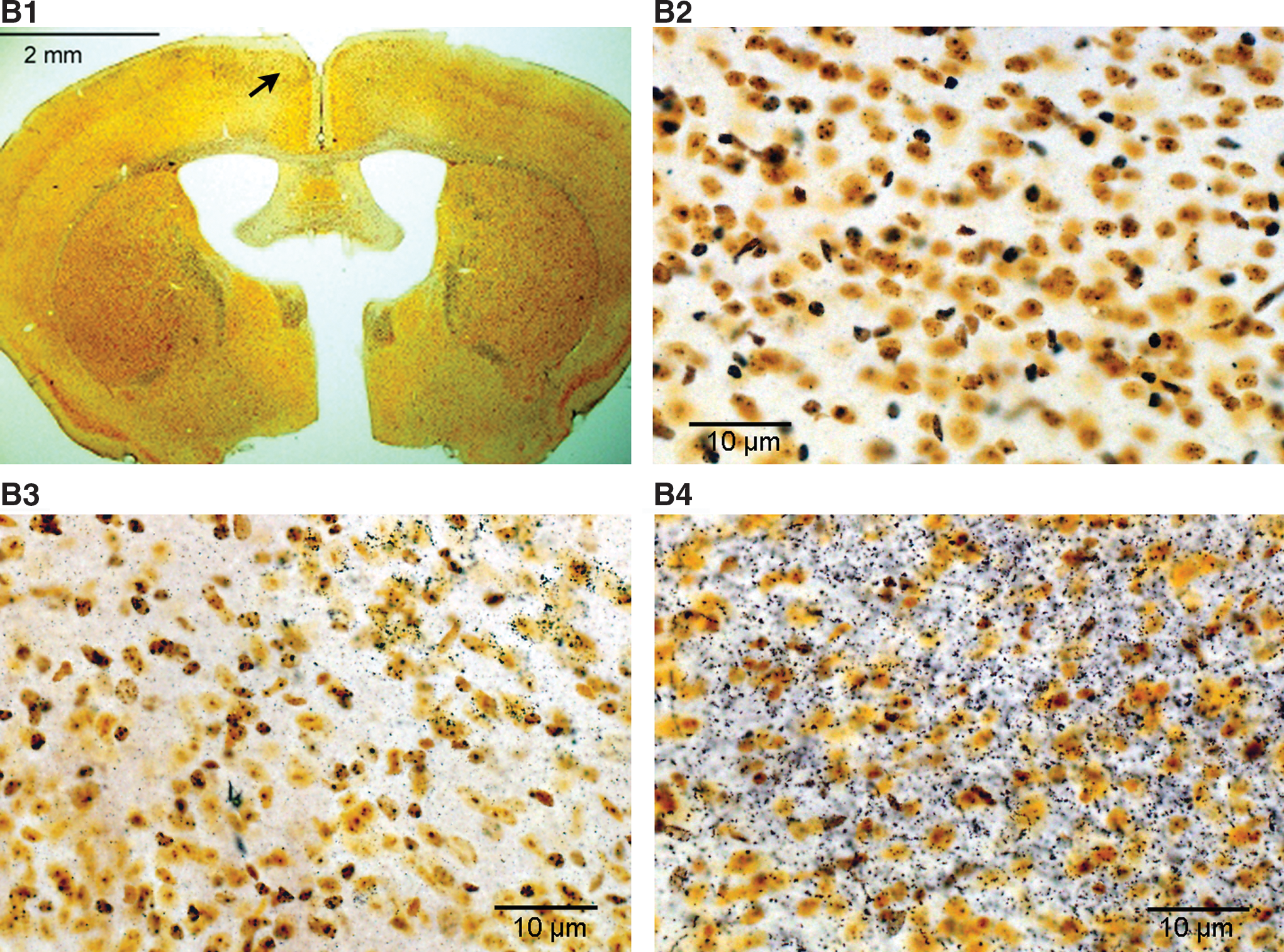

Fluoro-Jade B labeled neurons were absent in the brain sections of sham controls (Fig. 7C1, C2) and single blast exposed mice at 20.6 psi, at 24 h (data not shown). Significantly increased numbers of Fluoro-Jade B positive neurons (150±79 dead cells/mm2, n=3) were observed in animals that received BOP3 at 24 h (Fig. 7C3, C4). The observed neuropathology was primarily in the contralateral cerebral hemisphere including the second and third layer of the cortex, the cortex–amygdala transition zone, and the third ventricle. A few degenerated Purkinje cells in the cerebellum were also observed (data not shown).
Motor function behavioral test in repeated blast exposed animals
Mice received daily Rota Rod training in both a constant speed mode (4 rpm), and an accelerating speed mode (from 1 to 10 and 20 rpm, respectively) with three trials of 3 min maximum duration. After 3 days of training, the latency to fall was 97.8±7.4 sec at 10 rpm and 62.9±6.9 sec at 20 rpm (Fig. 8A). Latency to fall at baseline, 2 h, and 1, 3, and 5 days after single or three repeated BOP exposures for each mouse were compared. Blast-exposed mice showed a decrease in motor performance following BOP1 or BOP3 (Fig. 8B). The decrease in performance was significant at 2 and 24 h after blast exposure at 10 rpm. At 72 and 120 h after BOP, the performance was improved on the Rota Rod. At 20 rpm, both single and repeated BOP exposed animals showed significantly lower latency time to fall compared to sham controls up to 72 h after injury. With 20 rpm, at 72 or 120 h post-blast, there was no significant performance recovery in animals that had received BOP3. A marginal reduction in performance decrement at 20 rpm was observed between BOP1 and BOP3 animals in the Rota Rod test, but it was not significant.

Neuromotor functional analysis of single and repeated blast exposed mice. (
Discussion
The data presented here show that single blast exposure in mice leads to overpressure-dependent brain injury, and the severity of injury is elevated with an increased number of blast exposures. In single blast exposed mice, blast peak overpressure dependent increase in the severity of brain injury characterized as mild, moderate or severe has been described earlier (Cernak et al., 2011; Koliatsos et al., 2011). In our studies with single blast exposure, we observed significant animal mortality at 25 psi overpressure and it is consistent with the development of severe blast TBI. Necropsy of these animals showed subdural hemorrhage, and significant injury to lungs and internal organs indicating severe polytrauma (data not shown). A few animals exposed to 25 psi BOP also showed hunched back posture and reduced eating and overnight death after blast exposure. Mortality after single blast exposure at 13.9 and 20.6 psi was minimal, and occurred immediately after blast exposure, probably because of apnea or injury to lungs and other organs. A BOP of 20.6 psi was chosen to further study the effects of repeated blast exposures, as the mortality rate at this peak overpressure was minimal after first blast exposure.
In the repeated blast exposed group, second and third blast exposures further increased the overall mortality rate of the animals. Increased mortality suggests cumulative injury in repeated blast exposed animals or possibly a decrease in BOP threshold for injury after the first blast exposure was reported earlier, using different model systems (Saljo et al., 2009, 2011). In repeated blast exposed groups, 8% of the surviving animals showed subdural hemorrhage and 6% of the animals showed lung and abdominal hemorrhage indicating moderate to severe brain injury with polytrauma. The animals were restrained using a net inside the animal holder to minimize the movements caused by blast, and any potential impact with the holder resulting in brain injury. However, it is hard to completely rule out the possibility of minor head acceleration under these conditions. Restraining the animal in the holder and minimizing the mobility caused by blast is important to reduce other types of injuries, and it has been reported to reduce subdural and subarachnoid hemorrhage (Cernak et al., 2011).
Animals exposed to two or three repeated BOP at 20.6 psi showed more prolonged unconsciousness and severe brain injury compared to those exposed to single blast exposure. The righting reflex time after last blast exposure was significantly prolonged following blast exposure, and was further increased after three blast exposures. This indicates that repeated blast exposures of animals increases the duration of unconsciousness. Gross examination of the animals exposed to blast showed no physical injury, indicating that extremity injury was not the reason for delayed righting reflex time. Unlike our studies with mice, single blast exposure of rats at 120 kPa did not result in any delays in the righting reflex time (Readnower et al., 2010). However, it has been reported that single blast exposure led to an increase in paw flexion and eye reflex times in rats (Readnower et al., 2010). Unconsciousness related to bTBI has been reported in war victims (Matthews et al., 2011a,b; Wilk et al., 2010). Preliminary studies reveal that the increase in righting reflex time correlates to the extent of neuropathology in repeated blast exposed animals (Wang and Nambiar, unpublished data).
A BOP-dependent body-weight loss was observed in mice after single blast exposure. A similar type of body-weight change after mild and moderate brain injury spanning >30 days in mice has been reported earlier (Cernak et al., 2011). The recovery of body-weight loss was slower with higher blast overpressure exposure. Repeated blast exposures at 20.6 psi increased the body-weight loss compared to single blast exposure, and the increased body-weight loss remained for 3 days after blast exposure. Body-weight loss could be caused by changes in appetite or metabolism, olfaction, or other injury effects of blast, which needs to be studied further. Changes in olfaction and taste have been reported after explosive blast injury in rats and pigs (Bauman et al., 1997, 2009).
Increase in intracranial pressure after blast exposure has been reported earlier (Saljo et al., 2008, 2009,2010, 2011). The increase in intracranial pressure has been suggested to be the consequence of increased edema, hemorrhage, blood–brain barrier damage, and CSF flow obstruction (Hoane et al., 2006; Saljo et al., 2011; Unterberg et al., 2004; Vink et al., 2003). Brain edema was significantly increased in the three repeated blast exposed mice at early time points, indicating that edema is a component of increased severity of repeated bTBI. The modest increase in brain water content could also be the result of an increase in cerebral blood volume, and needs to be investigated further. Brain water content was reduced to normal levels in animals at 24 h, suggesting that it is a transient acute effect of repeated blast exposures. Early increase in intracranial pressure that reached to peak at 10 h and gradually returned back to normal levels at 24 h has been reported with repeated low-level blast exposure in pigs (Saljo et al., 2011).
Oxidative stress is one of the major contributors of bTBI and increased brain oxidative stress after blast exposure has been reported in experimental animal models as early as 3 h post-exposure (Cernak et al., 2001b; Readnower et al., 2010). In our mouse repeated blast exposure model, ROS, a measure of oxidative stress, was increased as early as 6 h post-blast. It has been reported that upregulation of 4-hydroxynonenal and 3-nitrotyrosine in the brain, as a measure of increased oxidative stress, was returned to normal by 24 h post-blast exposure in rats (Readnower et al., 2010). Our results indicate that, compared to 6 h, the ROS levels were further increased at 24 h post-exposure, suggesting ongoing oxidative stress. Our results agree with the earlier report showing maximum increase in malondialdehyde at 24 h in the hippocampus of rats exposed to single blast (Cernak et al., 2001b). Increase in ROS formation after blast exposure was similar between single and three repeated blast exposures, showing that multiple blasts do not further increase the level of ROS formation. Although there is an increase in the level of ROS at 24 h, the magnitude of responses to single and three repeated BOPs were similar. Similar levels of lipid peroxidation after two, three, and four blast exposures in the lungs and increased lung injury with time has been reported earlier (Elsayed and Gorbunov, 2007; Guy et al., 1998).
Limitations of our study include use of single marker for oxidative stress injury. It has been reported that the measure of dichlorofluorescein fluorescence does not necessarily reflect the levels of ROS directly but rather an overall index of oxidative stress in cells (Wang and Joseph, 1999b). To better understand, additional markers of oxidative stress are required to quantify the role of oxidative injury in this model. Physiological observation before and after the third blast exposure has not been performed in this study. Because body armor was not used, lung, heart, vasculature, or the autonomic nervous system may be similarly affected by repeated exposures. Also, animals may become hypoxic or hypo- or hypertensive and that could contribute to brain injury.
Hematoxylin and eosin staining of the brain at 24 h showed non-significant changes between sham and repeated blast exposed animals (data not shown). In contrast, there was greater silver staining in the cortex and cerebellum of repeated blast exposed animals compared to sham controls at 24 h. It has been reported that the cerebellum showed more axonal injury after blast exposure with body shielding (Garman et al., 2011). Silver staining was more intense in the cerebellum compared to the cortex in repeated blast exposed animals. However, the cerebellum did not show any neuronal death, and the injury seemed to be predominantly an axonal injury. It is possible that the frontal cortex and the cerebellum may have different type/extent of brain injury after blast exposure. Another possibility is that cerebellar injury involves possible acceleration-mediated injury caused by blast, as it is closely associated with the midbrain and spinal cord at the neck region. Increased silver staining of different brain regions after single blast exposure in mice has been described recently (Koliatsos et al., 2011).
Neurons damaged by blast exposures may die gradually and are evident with Fluoro-Jade B staining. Animals that exhibited longer righting reflex time showed degenerating neurons in dorsal and piriform cortex regions. This line of evidence suggests that the righting reflex time may be an indicator of the extent of the severity of brain injury. Limited evidence of neuronal death was reported after blast exposure (Garman et al., 2011). Mild repeated closed head injury also showed no substantial evidence of neuronal death (Shitaka et al., 2011). Recently, Pun and associates showed neuronal death in rats after high intensity blast at 24 h that abated after 4 days (Pun et al., 2011). They also found white matter tunnel positive cells after both low- and high- intensity explosive blast. Although blast exposure has been reported to show very limited evidence of neuronal death, the observed neuronal death in our studies at 24 h is probably related to the severity of the repeated blast exposures, and needs to be studied for a longer time to see whether they subside and regenerate. In the cerebral cortex after repeated blast exposures, Fluoro-Jade B staining was higher on the medial contralateral side than on the ipsilateral side. Probably, BOP from the ipsilateral side caused reflection of the shockwaves or compression on the contralateral side, causing more injury. In contrast, Fluoro-Jade B staining did not show any neuropathology after single blast exposure in mice (data not shown), which was similar to previous studies (Koliatsos et al., 2011).
Analysis of motor function after blast exposure showed a significant difference in performance following single blast exposure in animals starting from 2 h. The difference in neuromotor performance was maintained between sham controls and blast-exposed animals for 5 days, which was gradually improved with time. A similar profile in neuromotor function with mild and moderate BOP-induced TBI has been previously reported in mice (Cernak et al., 2011). At higher acceleration, the decreased neuromotor function did not change even after 5 days, indicating that repeated blast exposed animals are behaviorally impaired for a prolonged time after exposure. At higher acceleration, there was a marginal but not significant decrease in the neuromotor function in multiple blast exposed animals compared to single blast exposed animals. This observation needs to be investigated further to correlate with the neuropathology.
Conclusion
In summary, repeated blast exposures showed increase in mortality, body-weight loss, and righting reflex time compared to single blast exposure in mice. Also, brain edema and neuropathology by silver staining and Fluoro-Jade B were significantly higher in repeated blast exposed animals compared to single blast exposed animals. The time interval of 1 min and 30 min for the second and the third blast exposure is to mimic serial blast exposures in a short time period and is different from repeated blast exposures that occur within a few days. It is not clear what makes the brain more vulnerable to second and third blast exposures when they are closely juxtaposed, compared to when they occur after longer intervals. The vulnerability mechanism might be different when the second and the third blast exposures are close compared to when there are significant time intervals, and needs to be studied further. On the other hand, there was no significant difference in ROS and neuromotor performance in repeated blast exposed animals compared to single blast exposed animals. A trend of decreased neuromotor performance was observed in repeated blast exposed animals compared to single blast exposed animals, although it was not significantly different. A more comprehensive neurobehavioral battery of tests is required to discuss functional impairments resulting from single and repeated blast exposures.
Footnotes
Acknowledgments
We thank Dr. Richard Shoge, Irene Gist, and Andrea Edwards for technical help in blast experiments, coordination, and discussion.
Author Disclosure Statement
No competing financial interests exist.
The contents, opinions and assertions contained herein are private views of the authors and are not to be construed as official or reflecting the views of the Department of the Army or the Department of Defense.