
Abstract
Spinal cord injury (SCI) is a devastating condition that currently lacks clinically-relevant and effective neuroprotective therapeutic options. Optimal therapeutic agents for clinical translation should show efficacy in a cervical compression/contusion model using a clinically-relevant post-injury therapeutic time window. To date, few compounds have met that rigorous standard. The objective of this work was to evaluate the efficacy of delayed post-injury administration of soluble Fas receptor (sFasR) via intrathecal catheter following acute cervical SCI in a clinically-relevant contusion/compression model. Female Wistar rats were given a C7–T1 moderately severe clip compression injury, followed by either 8-h or 24-h delayed treatment initiation. Long-term neurobehavioral analysis of motor recovery and neuropathic pain development was undertaken. The extent of oligodendrocyte and neuron survival was assessed in peri-lesional cord sections 8 weeks post-SCI. This was complemented by an evaluation of the level of tissue preservation at and adjacent to the site of injury. In animals treated with sFasR delayed 8 h post-injury, significant behavioral effects were observed, coinciding with enhanced cell survival, peri-lesional tissue sparing, and enhanced integrity of descending fiber tracts compared to control treatments. Animals treated with sFasR delayed by 24 h showed more modest improvements in behavioral recovery, and had consistent improvements in cell survival and tissue preservation. This work has shown for the first time that the Fas-mediated apoptotic pathway can be therapeutically targeted in a clinically-relevant time window post-SCI.
Introduction
S
One of the major cellular events contributing to secondary damage after SCI is the initiation of apoptotic cell death in neurons and oligodendrocytes (Crowe et al., 1997; D'Souza et al., 1996; Katoh et al., 1996; Li et al., 1996,1997,1999a,1999b; Matsushita et al., 2000; Springer et al., 1999; Zurita et al., 2001). Fas-mediated apoptosis of oligodendrocytes is becoming well known as a key player in secondary injury mechanisms following SCI (Ackery et al., 2006; Casha et al., 2001,2005; Demjen et al., 2004; Yoshino et al., 2004).
Activation of the Fas-mediated apoptotic cascade within the receptor cell is triggered by ligand binding, ultimately leading to a cellular cascade of caspase activation and subsequent commitment to cell death (Cascino et al., 1995; Nagata 1997; Nagata and Golstein, 1995; Smith et al., 1990; Suda and Nagata 1994). Following SCI, Fas expression is increased predominantly in the oligodendrocyte population, and a correlation between Fas expression, caspase-8 activation, and apoptotic cell death has been well established in a variety of injury models (Ackery et al., 2006; Cashaet et al., 2001; Choi and Benveniste, 2004; Crowe et al., 1997; Davis et al., 2007; Demjen et al., 2004; Emery et al., 1998; Matsushita et al., 2000; Shuman et al., 1997; Zurita et al., 2001). Given the temporal delay in activation of Fas-mediated apoptotic signaling after SCI, this element of the secondary injury cascade represents an attractive therapeutic target. Several studies have highlighted that a dysfunctional Fas receptor is beneficial following SCI using the lpr mutant mouse (Casha et al., 2005; Martin-Villalba et al., 1999; Yoshino et al., 2004;). Furthermore, the neutralization or disruption of the Fas-mediated pathway using CD95-specific antibody or soluble Fas receptor (sFasR) has shown therapeutic success following SCI, resulting in increased cell survival, axonal sparing, and improved behavioral recovery, and enhanced regenerative responses (Ackery et al., 2006; Dasari et al., 2008; Demjen et al., 2004;). To date, these findings have been demonstrated following the administration of therapeutics at a very early time point post-injury, or at the time of injury itself. While these results are promising, it is important to establish that targeting Fas-mediated apoptosis in a clinically-relevant post-injury time window is associated with neurobehavioral recovery and objective evidence of enhanced peri-lesional tissue sparing.
In this article, we show for the first time that Fas-mediated injury mechanisms can be targeted in a therapeutically-relevant time window following spinal cord compression injury, using intrathecal administration of sFasR delayed by 8 or 24 h post-injury. sFasR is a human fusion protein consisting of the extracellular ligand-binding domain of the Fas receptor fused to a 234-amino acid domain of human IgG1 Fc for stability (Fig. 1A). This protein has the ability to bind with FasL in a competitive fashion with cell-bound Fas receptor, reducing the interaction potential of cell-bound Fas and FasL (Fig. 1B; Brunner et al., 1995; Ju et al., 1995). As a result of this competitive inhibition, it is hypothesized that endogenous Fas activation is limited, resulting in a reduction in post-traumatic Fas-mediated apoptosis. Treatment delayed by 8 h is effective in enhancing cell survival and preserving spinal cord tissue integrity, while allowing for a strong improvement in behavioral recovery. Delaying treatment initiation to 24 h post-SCI maintains a modest level of protective effects, with enhanced cell survival and tissue preservation. Being able to delay therapeutic intervention by such large time windows is advantageous in the translation of findings into a clinical setting, and it is hoped that this work can begin to set the stage for the development of clinically-effective treatment regimens for other molecules already showing promise pre-clinically.
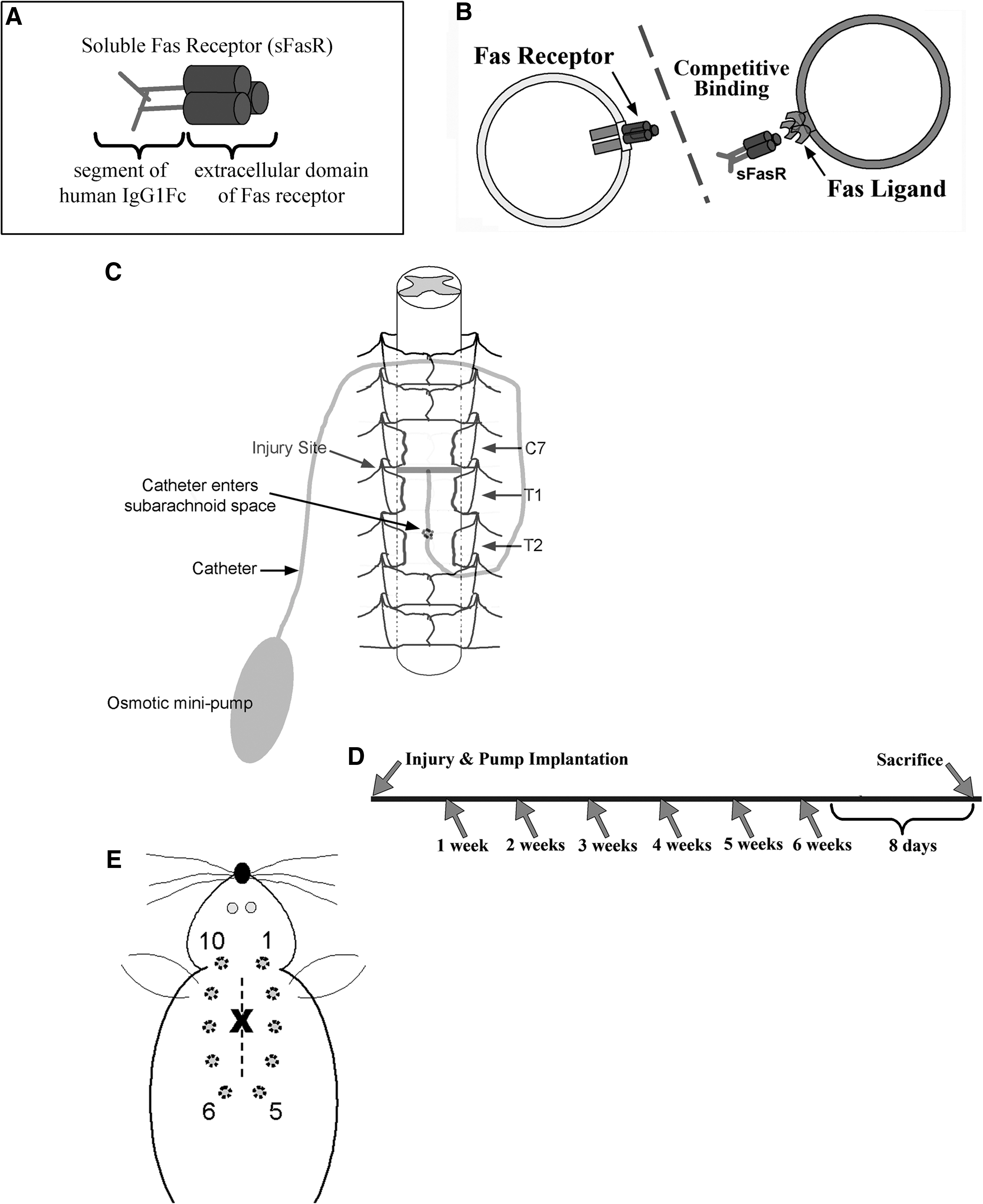
Soluble Fas receptor (sFasR) acts via competitive inhibition of cellular Fas-FasL binding. sFasR is a human fusion protein composed of the Fas receptor extracellular domain, stabilized by a 234-amino acid domain of human IgG1 Fc (
Methods
Spinal cord injury model and drug delivery
All animal use, handling, and care were carried out according to protocols approved by the Animal Care Committee of the University Health Network in accordance with guidelines established by the Canadian Council on Animal Care.
Eighty-seven adult female Wistar rats (weight ∼ 300 g) were anesthetized using 2% halothane in a 2:1 oxygen:nitrous oxide mix, while body temperature was maintained using an underlying thermal pad. A dorsal three-level laminectomy was performed to remove the T2, T1, and C7 vertebral lamina. For sham-operated animals, this marked the end of the procedure. To induce a clinically-relevant experimental spinal cord injury, a 35-g extra-dural clip was closed around the spinal cord at the vertebral level of C7–T1 for 1 min. Following removal of the clip, a subarachnoid catheter was inserted into the area of injury of animals in the saline-treatment group, attached to a mini-osmotic pump containing a saline solution (both catheter and pump from Alzet; Durect Corp., Cupertino, CA). The pump was inserted under the animal's skin along the left side of the body, with the catheter held in place by intramuscular sutures placed along the midline at the level of injury (Fig. 1C). For all other treatment groups (Table 1), the wound was closed using Michel clips and the animals were left under heat lamps to recover after injury. At either 8 or 24 h post-injury, the animals were re-anesthetized as previously described, the wound was re-opened, and a subarachnoid catheter and mini-osmotic pump were inserted into the area of injury as described above for the saline group. Here the mini-osmotic pumps contained either sFasR (Ancell, Bayport, MN) or a solution of bovine serum albumin (0.5 μg/μL BSA; Sigma-Aldrich, St. Louis, MO) in saline, giving a delivery rate of ∼0.6 μg/h. Soluble Fas receptor was considered the experimental treatment, and due to the need for a protein control, BSA was chosen to replicate all of the treatment paradigms using sFasR. In this case, saline acted as a secondary control and it was not felt that a delay in administration was necessary to establish this fundamental control. Figure 1D illustrates the injury model and treatment paradigm in a time-line format.
In the column labeled “SCI?,” the animals were given an experimental clip compression SCI (Yes) or were sham-injured using a laminectomy of C7, T1, and T2 only (No). The “Treatment type” column indicates the contents of the mini-osmotic pump implanted into the animals, with sFasR as the main treatment and BSA or saline as the control solutions. Note that both sFasR and BSA were diluted (10 μg/200 μL) in saline. The “Time of treatment initiation” indicates the time post-SCI of mini-pump and catheter implantation. The starting and final number of animals in each group is indicated in the final two columns.
sFasR, soluble Fas receptor; SCI, spinal cord injury; BSA, bovine serum albumin; N/A, not applicable.
Animal care
Following SCI and/or pump implantation surgeries, the animals were given 5 mL of saline subcutaneously, along with 0.4 mL of buprenorphine analgesic. They were allowed to recover under heat lamps until stable, and were brought to the animal facility for long-term care, where room temperatures were maintained at 26°C. For 2 days prior to, and 5 days following surgery, all animals received oral doses of Clavamox® in their water to prevent infections. Animals receiving the 35-g clip injury were given additional subcutaneous doses of 5% dextrose in a lactated Ringer's solution (5 mL three times per day) until their weight stabilized. Furthermore, any sign of post-operative hematuria was treated with Clavamox and subcutaneous saline injections until the infection cleared. The urinary bladders were manually expressed three times daily for the duration of the experiments.
Immunohistochemistry
At ∼7 weeks post-SCI, the animals were anesthetized using isoflurane and were transcardially perfused using a chilled 4% paraformaldehyde (PFA) solution. Spinal cord segments ∼0.5 cm in size centered on the injury epicenter were removed and post-fixed in 10% sucrose in PFA for 1 day, followed by 20% sucrose in phosphate buffered saline (PBS) for 2–3 days. Following post-fixation, the cords were embedded in OCT medium and snap frozen using dry ice; the cord sections were stored at −80°C until ready for cryosectioning. Using a cryostat, the cords were cut into 14-μm cross-sections and collected serially on glass slides. Sequential slides were stained using hematoxylin and eosin (H&E) and Luxol fast blue (LFB) in order to establish the location of the injury epicenter using tissue morphology and integrity as indicators. The slides containing the injury site and the rostral and caudal cord segments of interest were noted for cell quantification and tissue preservation analysis.
Cell quantification
Tissue sections from ∼ 800 μm rostral to the injury epicenter were taken from the saline, sFasR-8 h, BSA-8h, sFasR-24h, and BSA-24h animal groups (n=8/group). Two adjacent slides from each animal were used to quantify oligodendrocyte and neuron populations surviving at 7 weeks post-SCI, using stereologically-based techniques as described below. Briefly, tissue sections were removed from the freezer and allowed to acclimate to room temperature in PBS for 5 min. Individual sections were then traced using an Immedge pen, and a blocking solution (5% milk in PBS+1% BSA) was applied for 1 h at room temperature. Blocking solution was aspirated from the sections and the primary antibody solution (antibody+blocking solution) was applied to sections and left overnight at 4°C. Primary antibodies for oligodendrocytes (CC1, APC, 1:40; Calbiochem, San Diego, CA), and for neurons (NeuN 1:300; Chemicon, Billerica, MA) were used. Following primary antibody incubation, the tissue sections were washed three times for 10 min each with PBS. Secondary antibody solution (1:400 Alexa-Fluor 488 fluorescent anti-mouse in blocking solution) was applied to the sections and left for 1 h at room temperature. The slides were subsequently washed as previously described, and were cover-slipped with mounting medium containing 4′-6-diamidino-2-phenylindole (DAPI) for nuclear localization. To evaluate non-specific labeling, control staining was carried out on serial sections, for which primary antibody incubation was replaced with blocking solution, and then followed as the others with secondary antibody incubation.
The imaging of tissue sections was carried out with a Leica 510 fluorescence microscope using Stereo Investigator Software to create full section images. Quantification of double-labeled CC1+DAPI-positive cells and NeuN+DAPI-positive cells was carried out in a blinded fashion using Adobe Photoshop CS3 counting software. For the oligodendrocyte counts, the entire section white matter areas were counted, whereas for the neurons the entire section grey matter areas were counted, as determined by tissue morphology and previous H&E- and LFB-stained sections.
Treatment group averages of total cell counts were calculated, and one-way analysis of variance (ANOVA) tests were applied to the 8-h and 24-h delayed administration treatment groups separately, using the saline-treated group as the base control for each. Post-hoc analysis was carried out using the Student-Neuman-Keuls method.
It was elected to sample rostral cord sections for this analysis to provide a representative sample of the penumbra region surrounding the injury epicenter. The most consistent protective effects of sFasR administration were previously observed in the rostral cord (Ackery et al., 2006), and the analysis presented here was complemented by stereological histological morphometric analysis of tissue preservation. Furthermore, the extent of white matter degeneration and demyelination are heightened in the rostral spinal cord following SCI, giving merit to the selection of this area for cell survival analysis.
Tissue preservation
Tissue sections from all treatment groups listed above (except sham animals) at the injury epicenter and at ∼1200 μm and ∼700 μm rostral and caudal to the injury epicenter (as determined above) were stained with H&E and LFB to evaluate tissue morphology and histopathology. Individual tissue sections were imaged using a Leica 510 light microscope and Stereo Investigator software, and areas of interest (spared/intact grey matter and cavity) were measured in relation to the total area of each section using Image J software. Spared/intact grey matter was characterized by a consistent eosinophilic staining pattern, spotted with hematoxylin-positive neurons, and a lack of necrotic and inflammatory infiltrate. The cavity was characterized as any non-staining areas within the boundaries of the tissue section, not created through tissue artifact. Percent grey matter preservation and percent cavity formation averages for each of the five spinal cord locations within each treatment group were calculated (n=5/group/location). Two-way ANOVA examining the effects of time and treatment were carried out, with the 8-h and 24-h delayed administration groups analyzed separately. The saline group acted as the base control for each delayed treatment group. Where appropriate, post-hoc analysis was carried out using the Holm-Sidak test.
Behavioral testing
Following injury and pump implantation, all animals were observed weekly for 6 weeks post-injury and were evaluated using the Basso-Beattie-Bresnahan (BBB) locomotor recovery scoring system. The BBB score is a non-linear scale of 0–21 points used to assess the motor abilities of three joints (hip, knee, and ankle) of the hindlimbs. Sham-injured animals all scored 21 points immediately after the laminectomy surgery, and were therefore left out of formal recovery observations. For all SCI animals, each week the animal was placed in a large open field and was observed and video recorded for 3 min to note the movements of its hip, knee, and ankle joints of both legs. Each limb was given a separate score and then the two were averaged to yield one score for that animal for that week.
Treatment group averages for each of the six observation weeks were calculated, and two-way ANOVA was performed on all of the treatment groups together (five groups in total), and then on the 8-h and 24-h delayed treatment groups independently of each other, using the saline-treated group as a control for each. To further illustrate specific treatment effects, post-hoc analysis was carried using the Holm-Sidak method.
The sizes of the experimental groups used to evaluate the recovery of locomotor function in these experiments were as follows: 16 animals (sFasR 8-h), 8 animals (BSA 8-h), 18 animals (sFasR 24-h), 10 animals (BSA 24-h), and 14 animals (saline).
Neuropathic pain assessment
Following injury and pump implantation, all treatment groups and an additional sham-injured group (n=5) were assessed for neuropathic pain development every other week for 6 weeks post-injury. The manifestation and progression of mechanical allodynia was assessed using von Frey filaments, with a 2-g and 4-g filament used at each time point. Briefly, each animal was allowed to acclimate to its surroundings for approximately 20 min prior to each pain testing period. Starting with the 2-g filament, the animal was touched 10 times around the level of injury (Fig. 1E). The number of withdrawal responses out of 10 were recorded, and the withdrawal responses counted were vocalization, adverse movement away from the stimulus, flinching, shivering, or scratching. The animal then went through the same procedure with the 4-g filament. The average number of withdrawal responses for each treatment group were calculated, and the 4-g stimulus data were analyzed using logistic regression, and Wald confidence intervals for odds ratios were determined. In this analysis, the 8-h delayed treatment groups were separated from the 24-h delayed treatment groups, with the sham animals and saline-treated animals included for reference in each set of analysis.
The sizes of the experimental groups used to evaluate neuropathic pain development were as follows: 16 animals (sFasR 8-h), 8 animals (BSA 8-h), 18 animals (sFasR 24-h), 10 animals (BSA 24-h), 14 animals (saline), and 5 animals (sham-injured).
Results
Delayed soluble Fas receptor administration enhances the survival of oligodendrocytes following SCI
When treatment administration was delayed by 8 h or 24 h post-injury, animals receiving sFasR showed enhanced oligodendrocyte survival in areas adjacent to the injury epicenter at 7 weeks post-SCI, compared to animals receiving BSA or saline control treatments (Fig. 2A). White matter cell counts of APC-positive oligodendrocytes were significantly higher in animals treated with sFasR delayed by 8 h post-injury (589±70) than in animals treated with BSA delayed by 8 h post-injury (221±40; Fig. 2B and C). The secondary control saline-treated group had an average of 195±37 oligodendrocytes per section. The efficacy of sFasR treatment extended to the group with a treatment delay of 24 h post-injury, with sFasR animals having 377±56 APC-positive oligodendrocytes per section, and BSA animals averaging 234±47 (Fig. 2D and E). Of note, sFasR-treated animals receiving 8 h delayed administration had greater survival of oligodendrocytes in the peri-lesional penumbra zone than the 24-h delayed sFasR group (p<0.05).
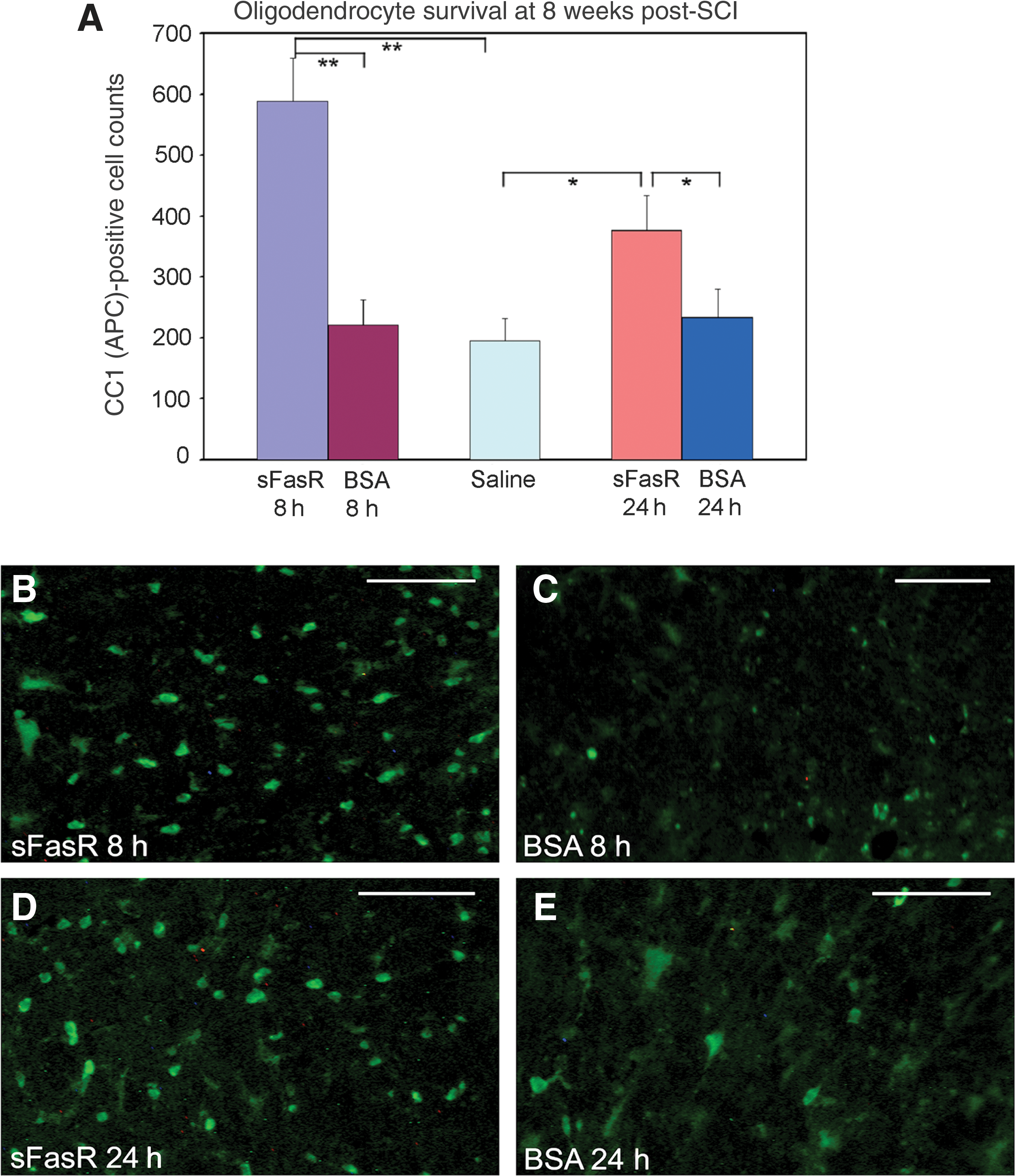
Oligodendrocyte survival. (
Neuronal survival in chronic spinal cord injury is improved with post-injury administration of sFasR delayed by up to 24 hours
Both 8-h and 24-h delays in the initiation of sFasR administration post-SCI were able to promote significant improvement in neuron survival at 7 weeks post-injury in rostral spinal cord grey matter compared to BSA and saline controls (Fig. 3A). NeuN-positive cells were quantified in the grey matter of spinal cord sections ∼ 800 μm rostral to the injury epicenter to reveal that sFasR-treated animals with administration delayed by 8 h had a significant improvement in neuron survival (462±40), compared to BSA treatment delayed by 8 h (266±41; Fig. 3B and C). Saline-treated controls had an average of 295±33 NeuN-positive neurons per section. Moreover, the sFasR treatment could be delayed by 24 h and the significance of neuronal survival was not lost. Animals treated with sFasR delayed by 24 h post-injury had an average of 431±41 neurons per section (Fig. 3D), while the BSA-treated controls also delayed by 24 h had only 222±32 neurons per section surviving at 7 weeks post-injury (Fig. 3E). Interestingly, in contrast to the impact on oligodendrocyte survival, similar neuronal survival was seen in the perilesional region of the cord in the groups receiving sFasR at 8 or 24 h post-injury.
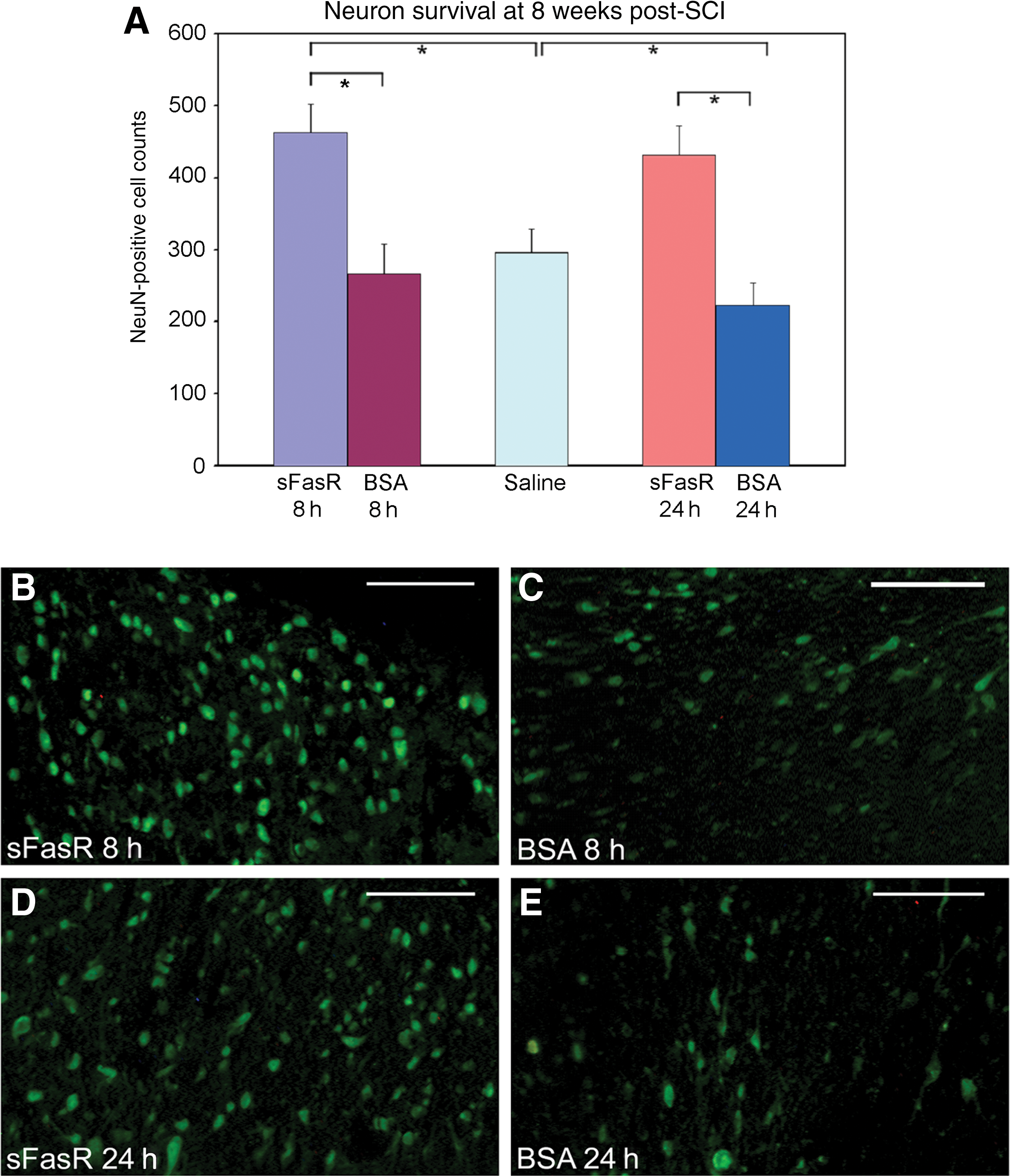
Neuron survival. (
An 8-hour delay in sFasR administration maintains grey matter integrity and reduces the cavity size at 7 weeks following SCI
Tissue sections from animals treated with sFasR delayed by 8 h, BSA delayed by 8 h, and saline-treated controls were analyzed. Rostral, caudal, and injury epicenter spinal cord tissue sections were assessed for tissue morphology, integrity, and cavity size. Animals treated with sFasR showed an overall improvement in grey matter tissue preservation compared to BSA and saline-treated controls, with further significant improvements seen at areas 700 μm caudal and 1200 μm rostral to the injury epicenter (Fig. 4A, B, and C). Furthermore, the overall cavity size was reduced by sFasR treatment, with specific tissue sparing observed at 1200 μm rostral to the injury epicenter, compared to both BSA-treated and saline-treated controls (Fig. 4D).
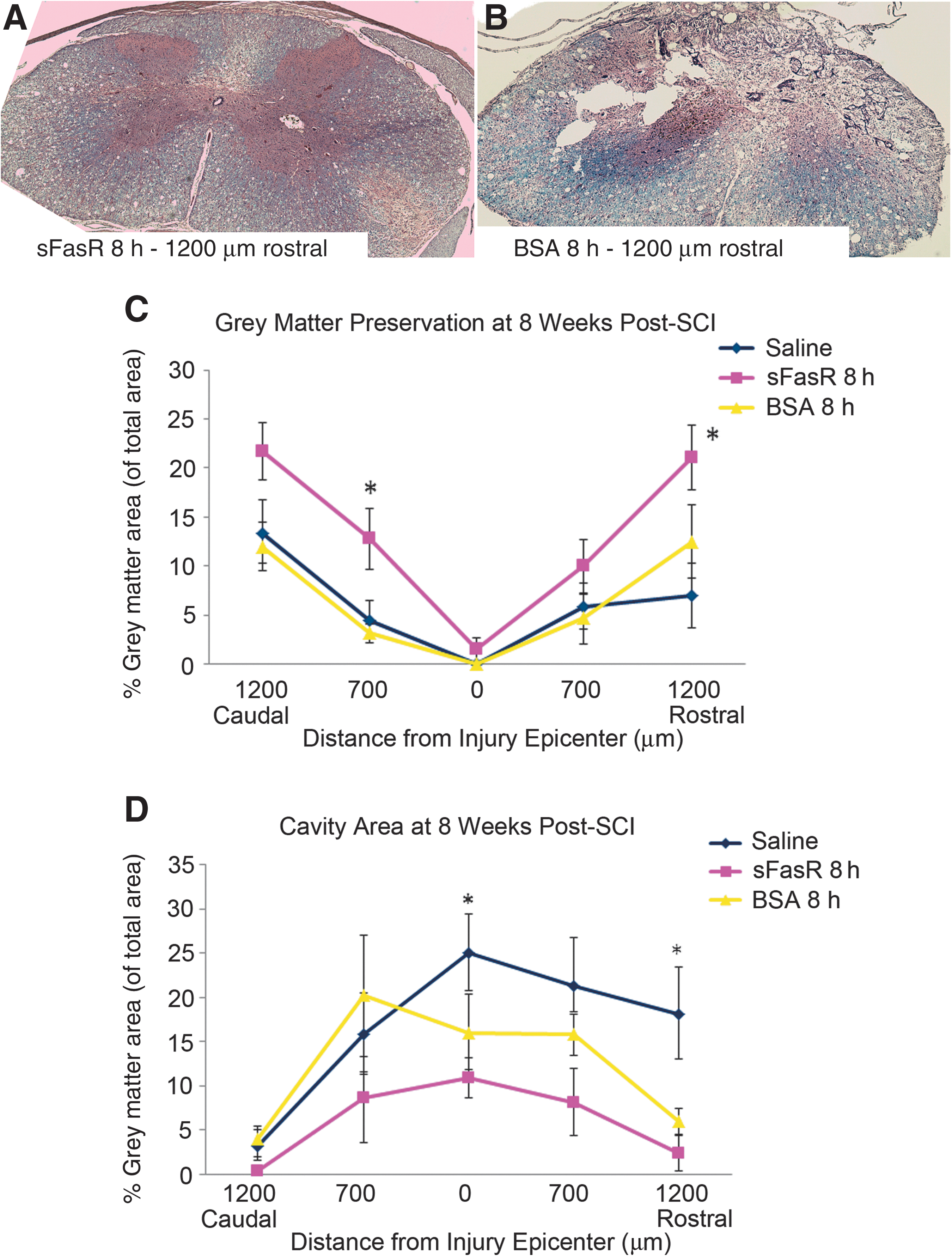
Grey matter preservation and cavity formation. Tissue sections taken from ∼1200 μm rostral and caudal to the injury epicenter, ∼700 μm rostral and caudal to the injury epicenter, and at the injury epicenter, 7 weeks post-SCI were stained with hematoxylin and eosin and Luxol fast blue to show tissue morphology and neuroanatomical structure (n=5 per treatment per location). Representative tissue sections from ∼1200 μm rostral to the injury epicenter, treated with either sFasR (
24-Hour post-injury delay in sFasR administration improves grey matter preservation while modestly reducing overall cavity formation at 7 weeks post-SCI
Tissue sections from animals treated with sFasR and BSA, both with initiation of treatment delayed by 24 h post-injury, were collected from rostral, caudal, and injury epicenter areas of the cord. Stained sections were assessed for the extent of grey matter survival and the area of cavity formation at 7 weeks post-SCI. It was observed that sFasR treatment could be delayed by 24 h with the beneficial effects of grey matter preservation remaining significant compared to the BSA and saline control groups (Fig. 5A, B, and C). The size of the cavity formed at this time point post-injury was also evaluated, and 24-h delayed sFasR treatment gave an overall reduction in cavity size, but this effect was only slight, showing the most benefit in the rostral cord sections (Fig. 5D).
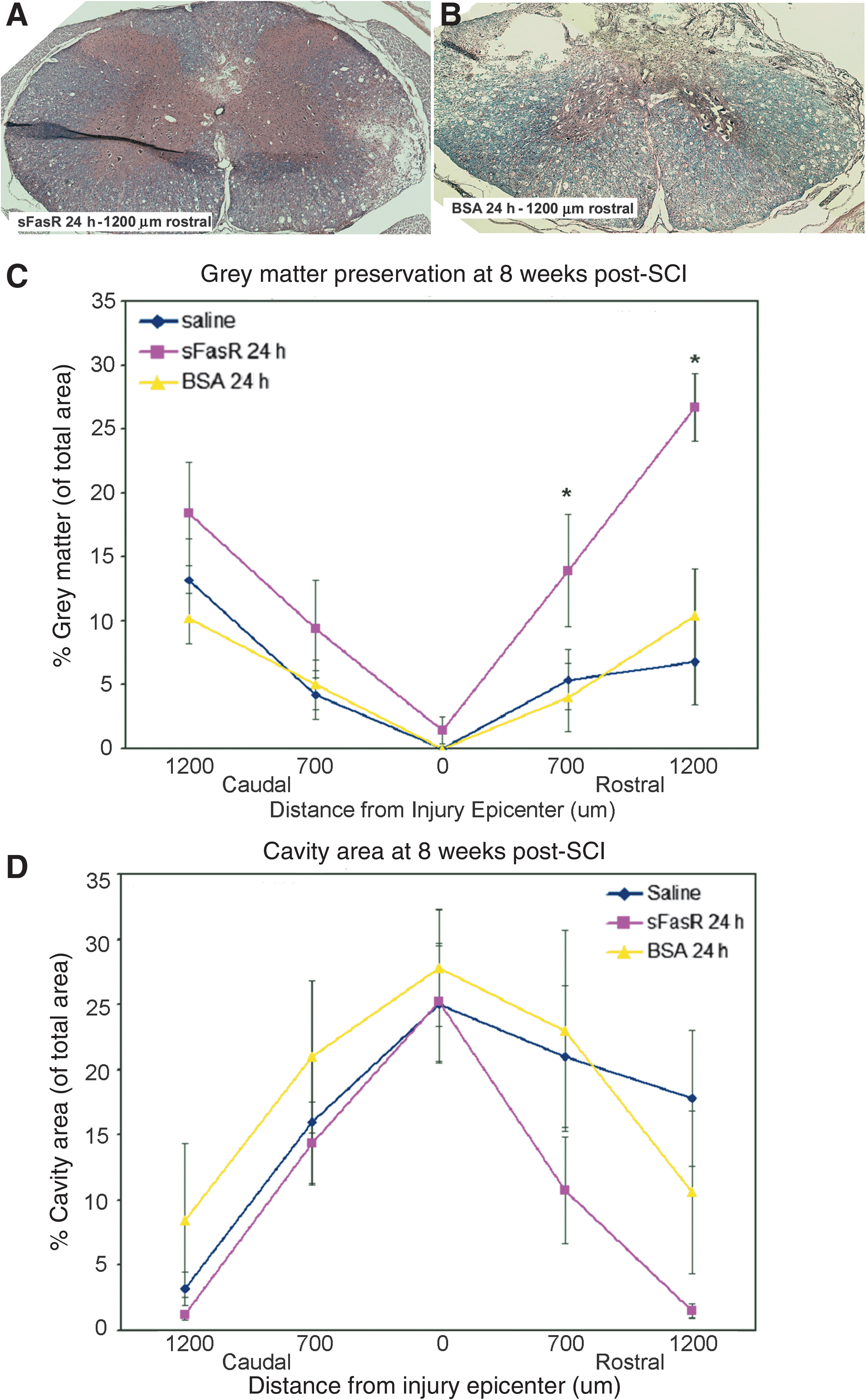
Grey matter preservation and cavity formation. Tissue sections taken from ∼1200 μm rostral and caudal to the injury epicenter, ∼700 μm rostral and caudal to the injury epicenter, and at the injury epicenter, 7 weeks post-SCI were stained with hematoxylin and eosin and Luxol fast blue to show tissue morphology and neuroanatomical structure (n=5 per treatment per location). Representative tissue sections from ∼1200 μm rostral to the injury epicenter, treated with either sFasR (
Intrathecal sFasR treatment shows efficacious effects on behavioral recovery following SCI when administration is delayed by up to 24 hours post-SCI
Long-term behavioral recovery was assessed using the BBB hindlimb locomotor rating scale in spinal cord injured animals treated with sFasR, BSA, or saline, with the sFasR and BSA treatments delayed by either 8 h or 24 h post-injury. The 8-h delayed groups were considered independently from the 24-h delayed groups for treatment significance as assessed by a two-way ANOVA of all five treatment groups and six weekly time points. In both independent analyses, the saline-treated animals acted as a control group.
In the 8-h delayed group, sFasR administration offered a significant improvement in motor recovery of the hindlimbs, with BBB scores between 1.5 and 2 points above the BSA and saline controls (Fig. 6A). Two way ANOVA revealed an overall treatment effect showing that sFasR was better able to enhance functional recovery as compared to both saline and BSA (p<0.001). It should be noted that with a non-linear scale such as the BBB, point differences have altered biological meanings at various levels of the scale. At this level of the BBB, scores ranging from an 8–10 show strong biological differences. An animal with a score of 8 will have no weight-bearing at all, though it is able to have full motion of all three leg joints, while an animal with a score of 9 or 10 is able to bear weight and an attempt at actual stepping is seen.
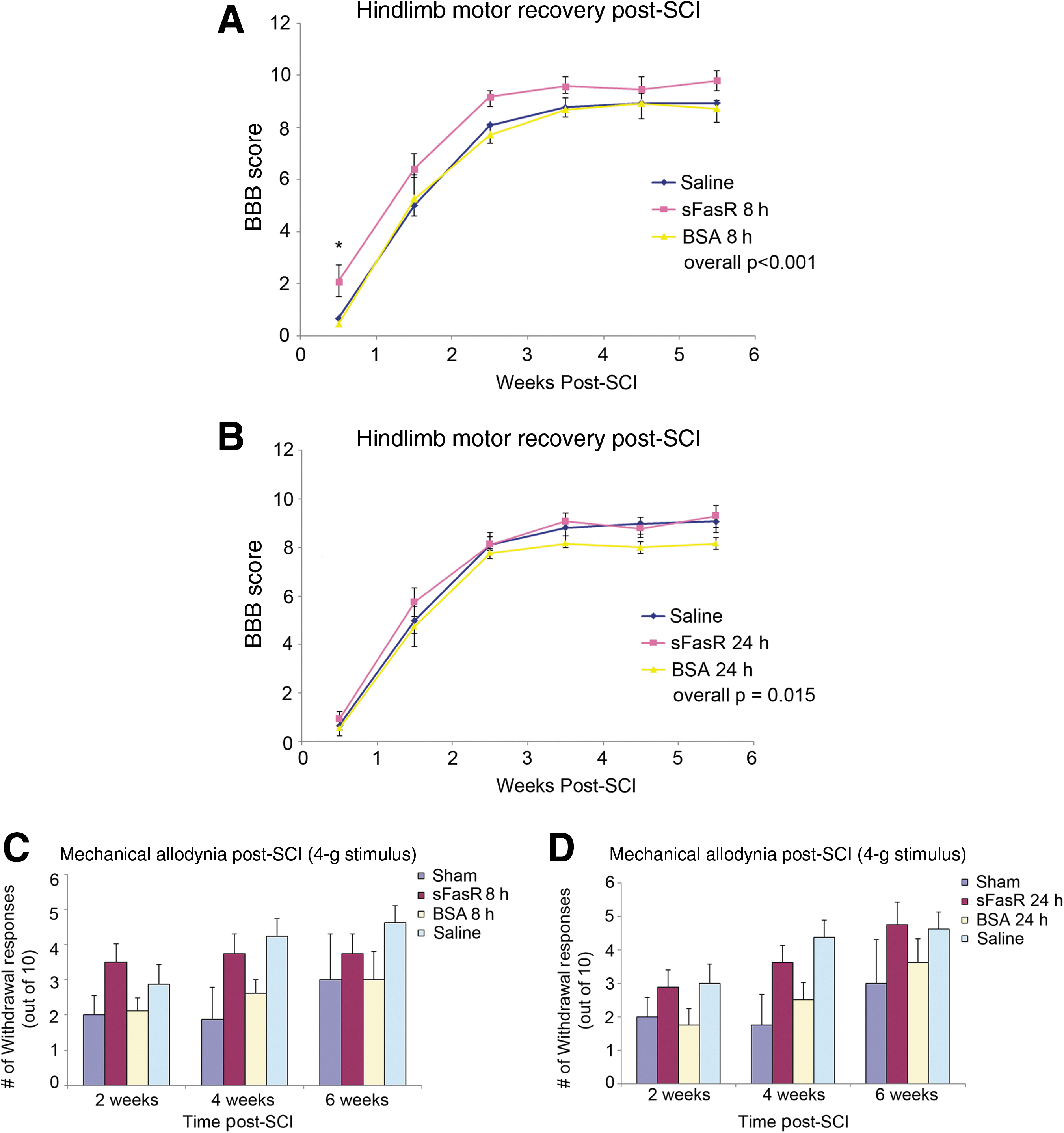
Behavioral recovery and neuropathic pain analysis. (
In the 24-h delayed group, sFasR administration offered a modest improvement in hindlimb motor recovery, but this effect was not as striking as that observed with an 8-h delay in administration (overall two-way ANOVA p=0.015; Fig. 6B).
The development of neuropathic pain was evaluated with mechanical allodynia as the pain response, using 2-g and 4-g von Frey filaments. The 4-g stimulus is considered to be the most appropriate for gauging mechanical allodynia in this kind of SCI model (Bruce et al., 2002). The average number of withdrawal responses from the 4-g filament is shown in Figure 6C and D. Logistic regression using Wald confidence interval for odds ratios revealed that while the level of neuropathic pain was consistently greater in all of the 8-h injured groups (p=0.002), and the 24-h injured groups (p=0.0019) compared to sham animals, the 8-h and 24-h delay in sFasR administration caused no adverse progression of neuropathic pain.
Discussion
Summary of findings
These data provide a clinically-relevant and therapeutically-effective treatment strategy for targeting Fas-mediated cell death following SCI for the first time. We show that sFasR treatment can be delayed by as much as 24 h post-injury and still maintain modest therapeutic effects, with administration after 8 h offering a greater overall benefit. Functional improvements observed in animals treated with sFasR at 8 h post-SCI were reflected neuroanatomically as long-term increases in both oligodendrocyte and neuron survival.
Rationale
There is currently a significant lack of pharmacological therapies available to patients suffering from acute SCI. Methylprednisolone is the only approved drug in clinical use, but it provides limited efficacy in a broad patient population, and may be accompanied by adverse side effects related to the increased risk of infection and wound complications (Baptiste and Fehlings, 2008; Bracken, 1990; Bracken et al., 1992; Geisler et al., 2001; Kaufman et al., 2000; Oudega et al., 1999; Petitjean et al., 1998). Finding a drug that is effective even with a delay between injury and drug administration, as is almost inevitable in real-life clinical cases, is a constant challenge to researchers. Here we have shown a potentially valid, effective, and clinically-relevant therapeutic intervention for patients with acute traumatic SCI that can be successfully delayed by more than 8 h post-injury.
Targeting Fas-mediated apoptosis following SCI
The therapeutic targeting of Fas-mediated cell death following spinal cord trauma has proven successful in the past, using several injury models and treatment paradigms. First, a dysfunctional Fas receptor in the lpr mutant mouse has been shown to facilitate improved cell survival, axonal preservation, and enhanced motor recovery following a clip compression model of SCI (Casha et al., 2005; Martin-Villalba et al., 1999; Yoshino et al., 2004). By targeting the Fas and FasL interaction pharmacologically, Demjen and colleagues were able to show that neutralization of the ligand using a targeted antibody treatment enhanced regeneration, as well as neurobehavioral recovery, in a mouse dorsal hemisection model of SCI (Demjen et al., 2004). Following this, our group demonstrated that intrathecal administration of sFasR at the time of injury can enhance cell survival, promote tissue sparing, and improve locomotor recovery in a highly relevant cervical compression model of SCI (Ackery et al., 2006). The relevance of targeting the Fas-mediated apoptotic pathway after traumatic SCI is clear; however, the clinical applicability of these treatments limits translation into the patient population.
In the present article, we evaluate the therapeutic time window for intrathecal sFasR treatment in a clinically-relevant contusion/compression rat model of acute SCI. It is the first time that a treatment delay of 8 or 24 h post-injury has been reported to have beneficial neuroprotective effects. Although the use of a subarachnoid catheter is slightly more invasive than intravenous injections, it is believed that the proper placement of the catheter in the cervical dural space does not adversely affect the recovery of the animals. Furthermore, the targeted delivery of sFasR to the injury site is expected to reduce the possibility of systemic reactions to the therapeutic agent; currently, adverse systemic effects are known to be problematic in SCI patients receiving steroid and inflammatory drugs intravenously (Bracken et al., 1990,1992).
Cell survival is enhanced following delayed sFasR administration
The significance of enhanced cell survival following moderately severe SCI is great. Not only does an increase in the number of surviving cells indicate a reduction in cell death, but it also suggests the possibility that a more supportive environment was established and maintained throughout the secondary injury process. In this work, sFasR treatment increased long-term survival of both oligodendrocytes and neurons in areas adjacent to the injury epicenter.
By evaluating cell survival at 8 weeks post-injury, an initial challenge is to assess the extent of an anti-apoptotic influence on cell survival as opposed to other possible positive environmental influences of sFasR. Previous work has shown convincing evidence that sFasR reduces the occurrence of post-traumatic apoptosis, suggesting that the extent of cell survival observed in these experiments is likely due in major part to the blockage of Fas-mediated cell death (Ackery et al., 2006). Furthermore, recent work in our lab has shown that the oligodendrocyte population expresses Fas and undergoes Fas-mediated apoptosis in the first week following SCI (Robins-Steele et al., in preparation for submission). These findings further strengthen the case for delayed sFasR administration for enhanced oligodendrocyte survival.
The observed reduction in neuronal death may be secondary to the disruption of the Fas pathway, as neurons are not known to express high levels of Fas, and there are internal inhibitory proteins expressed by these cells that inhibit Fas activation (Fernandez et al., 2007; Segura et al., 2007). However, enhancing the survival of the supportive glia, more specifically the oligodendrocyte population, would ultimately provide a more supportive environment for neurons and their axons normally negatively affected by SCI.
It is important to note that while 24-h delayed sFasR treatment significantly increased oligodendrocyte cell counts compared to controls, the 8-h delay demonstrated more striking beneficial effects. The same effect was not apparent in the evaluation of neuron survival, however, for which the benefits with regard to cell survival were similar in the 8-h and 24-h groups.
This raises the question as to how earlier initiation of sFasR treatment has a stronger impact on long-term oligodendrocyte survival than on neuronal survival. The previously proposed anti-apoptotic effects of sFasR on oligodendrocytes provide some explanation. Here, an early attenuation of oligodendrocyte apoptosis might account for a larger increase in the number of preserved cells than if the anti-apoptotic actions were delayed even slightly. In other words, the 8-h treatment had more time to prevent oligodendrocyte cell death than did the 24-h delayed treatment, resulting in a greater number of surviving cells at 8 weeks post-injury.
Considering the effect observed on neuron survival, it is possible that the preservation of oligodendrocytes contributes to the protection of neurons that might otherwise have died following SCI. If even a small amount of oligodendrocyte survival can impact the survival of neurons, then it is possible that both the 8-h and 24-h delayed treatments achieved this threshold.
Tissue morphology is preserved with delayed sFasR treatment
Correlating with improvements in rostral cord neuron survival, it was also observed that both 8-h and 24-h sFasR administration promoted the preservation and enhanced the integrity of grey matter tissue, particularly in rostral cord regions. This is not the first report in which stereological measurement of the lesion site following an experimental therapeutic showed greater effects in the rostral cord segments following SCI (McTigue et al., 2007). It is possible that by providing a supportive environment around the injury epicenter, there is a positive effect on retrograde degeneration, thus further contributing to less tissue and cell damage and loss. In this study, we did not examine the site caudal to the lesion. Any future work should examine areas both rostral and caudal to the lesion, as any differences could be informative with regard to the mechanisms of neuron survival and regeneration.
Retrograde degeneration of white matter tracts and grey matter tissue damage is common in this model of SCI (Ackery et al., 2006; Casha et al., 2001; Fehlings and Tator, 1995). The ability of delayed sFasR treatment to maintain a strong supportive cell population in rostral cord sections, as noted in the oligodendrocyte cell counts, gives credibility to the idea that this protective benefit might be reflected in improved tissue sparing. It was also noted that cavity formation in animals treated with sFasR was reduced compared to control-treated animals. This observation provides further evidence to support the idea that delayed sFasR treatment contributes to a more hospitable environment at and around the injury site, in part due to its anti-apoptotic effects. An 8-h delay in sFasR treatment was more effective in reducing the cavity size than a 24-h delay, which has been a consistent finding throughout this work. Delaying sFasR administration by 8 h post-injury is overall more efficacious than a 24-h treatment delay; however, not all benefit is lost with later administration.
Delayed sFasR administration has positive impacts on behavioral recovery
Behavioral analysis of hindlimb motor recovery using BBB scores revealed that both 8-h and 24-h delays in sFasR administration provided positive neurobehavioral outcomes. Animals treated with 8-h delayed sFasR showed between a 1- and 2-point increase in BBB scores compared to animals treated with 8-h delayed BSA or saline. This difference is indicative of a biologically significant change. The BBB scale is non-linear, ranging from a score of 0, meaning that the animal has no hindlimb movement in any joint in either limb, to a score of 21, indicating normal locomotion in a non-injured animal (Basso et al., 1996). The non-linearity of the scale is especially obvious in the range from 7–10, scores the animals in this study typically attained by 4 weeks post-injury. Here the differences from one point to the next reflect a significant biological effect. For example, an animal that scores 8 will show the ability to move all three lower limb joints, but lacks the ability to attempt stepping and has no visible weight support associated with its movements. An animal scoring 10 on the scale is able to attempt stepping, and can maintain weight support in stance as well as occasionally during the stepping process. In the context of a human patient suffering from SCI, the difference between these two points could translate into notable differences in quality of life, and would greatly reduce the possibility of co-morbidities associated with injury severity.
In choosing outcome measures for the current study we had to take into account that recovery from the level and severity of injury used in the experiments is not measured appropriately by well-known tests such as grid-walking or foot-print analysis. This is in part due to the strong degree of functional deficit and spasticity found in these animals, especially in the first 4–6 weeks post-SCI. Although the IBB (Irvine et al., 2010) was not used in this study to assess forelimb functional recovery, it could potentially be a useful neurobehavioral assessment tool for cervical SCI research, and may be used in future studies.
The behavioral analysis makes it clear that delaying sFasR treatment by 24 h is less effective than treating at 8 h post-injury, but it should be noted that a modest therapeutic effect remains with later treatment. Further examination of stronger doses and later administration time points would be important to explore the clinically-relevant 24-h window of opportunity. Fas-mediated cell death peaks between 5 and 7 days post-injury, suggesting that a 7-day drug delivery program is optimal to exert maximal anti-apoptotic effects.
Relationships between cell survival and behavioral recovery post-SCI
The specific correlation between neuron and oligodendrocyte survival and behavioral recovery following SCI is not well understood. An observed increase in cell survival has been seen to lead to improved functional recovery (Joshi and Fehlings, 2002). However, this correlation is not always obvious, and there can be preservation of cells at and around the injury site, with no observed beneficial effect on functional recovery. In the case of these experiments, both 8- and 24-h delayed sFasR treatment enhanced the survival of oligodendrocytes and neurons, and there was an additional preservation of grey matter noted in rostral cord sections for both delayed treatment groups. It is particularly worth noting that the significant effect of the 8-h treatment on oligodendrocyte and neuron cell populations is mirrored in the animals' improvement in neurobehavioral recovery. The cell counts for 24-h delayed sFasR were less significant, and this was reflected in the modest recovery of the BBB scores of these animals.
Neuropathic pain (mechanical allodynia) post-SCI
Finally, the development of neuropathic pain is a common complication faced by many patients with SCI (Christensen and Hulsebosch, 1997). While the specific pathologies underlying the progression of neuropathic pain are not well understood, there is some evidence to support the concept that aberrant sprouting of neurites and spontaneous attempts at regeneration play a role in its manifestation (Christensen and Hulsebosch, 1997). Researchers developing neuroprotective and regenerative treatment strategies therefore need to determine if the intervention is perpetuating pain responses, reducing pain responses, or maintaining the natural occurrence of pain. Mechanical allodynia is a form of neuropathic pain found in a large percentage of SCI patients, for whom a normally benign stimulus becomes highly noxious (Christensen and Hulsebosch, 1997).
In the comparison of sFasR-treated animals with BSA- and saline-treated controls, there was no obvious increase in allodynia responses across the 6-week post-SCI period of evaluation. By comparing pain responses with those of sham-injured animals, it is clear that while neuropathic pain progresses post-injury, there is not a significant increase with any of the treatments evaluated in this study.
The field of SCI research is growing at a steady rate, and there are many potential therapeutics showing promise in the lab. In order for researchers and clinicians to translate these therapies into clinical use, several goals need be accomplished. First, the efficacy of any treatment needs to be shown in valid and clinically-relevant animal models of SCI. Second, there needs to be a way to effectively translate the therapeutic paradigm into the SCI patient population. In order to accomplish this, suitable modes of administration and appropriate treatment time lines must be demonstrated in the lab, which then can be implemented in clinical trials. Here we have demonstrated that the efficacious treatment strategy of intrathecal administration of sFasR can be delayed from the time of injury to at least 8 h, while maintaining strong neuroprotective effects. Furthermore, the treatment can be delayed by 24 h and still result in substantial improvement. This work will set the stage for translational SCI research that will be effective in the clinical setting. Future studies of acute, traumatic SCI should aim to demonstrate positive effects with treatments initiated at later time points following SCI.
Footnotes
Author Disclosure Statement
No competing financial interests exist.