
Abstract
Brain injury and its related increased intracranial pressure (ICP) may lead to increased vagus nerve activity and the subsequent suppression of innate immunity via the cholinergic anti-inflammatory pathway. This may explain the observed increased susceptibility to infection in these patients. In the present study, we investigated the association between brain injury, vagus nerve activity, and innate immunity. We determined heart rate variability (HRV) as a measure of vagus nerve activity, plasma cytokines, and cytokine production of ex vivo lipopolysaccharide-stimulated whole blood in the first 4 days of admission to the neurological intensive care unit (ICU) in 34 patients with various forms of brain damage. HRV, immune parameters, and the correlations between these measures were analyzed in the entire group of patients and in subgroups of patients with conditions associated with high (intracranial hemorrhage [ICH]) and normal ICP (subarachnoid hemorrhage [SAH] with an extraventricular drain alleviating ICP). Healthy volunteers were used for comparison. HRV total spectral power and ex vivo-stimulated cytokine production were severely depressed in patients compared with healthy volunteers (p<0.05). Furthermore, HRV analysis showed that normalized units of high-frequency power (HFnu, corresponding with vagus nerve activity) was higher, and the low-frequency:high-frequency ratio (LF:HF, corresponding with sympathovagal balance) was lower in patients compared to healthy volunteers (p<0.05). HFnu correlated inversely with ex vivo-stimulated tumor necrosis factor-α (TNF-α) production (r=−0.22, p=0.025). The most pronounced suppression of ex vivo-stimulated cytokine production was observed in the ICH group. Furthermore, in ICH patients, HFnu correlated strongly with lower plasma TNF-α levels (r=−0.73, p=0.002). Our data suggest that brain injury, and especially conditions associated with increased ICP, is associated with vagus nerve-mediated immune suppression.
Introduction
T
In our hypothesis, we proposed a critical role for increased intracranial pressure (ICP; Kox et al., 2008). Elevated ICP is a common feature of TBI (Mowery et al., 2008) and SAH (Voldby, 1988). As a result of high ICP, compression of the brainstem may lead to increased firing of the medulla oblongata, where the efferent vagus nerve originates. Direct evidence for this phenomenon was found in cats, in which inflation of a subdural balloon resulted in increased vagus nerve discharges relative to ICP levels (Matsuura et al., 1984). Also, increased parasympathetic predominance has been demonstrated in the acute phase of TBI in patients (Biswas et al., 2000; Winchell and Hoyt, 1997). Furthermore, significant increases in parasympathetic activity were found in patients in the acute phase of SAH (Kawahara et al., 2003), and a recent study reported that parasympathetic predominance is a predictor of mortality in SAH (Chiu et al., 2011). In contrast to the acute phase, the late phase of TBI and SAH (>30 days after the insult) is associated with sympathetic rather than parasympathetic predominance (Baguley et al., 2006; Kawahara et al., 2003; Keren et al., 2005; King et al., 1997), possibly related to the normalization of ICP.
All of the clinical studies mentioned above have used heart rate variability (HRV) as a measure of autonomic nervous system activity, as this is the only non-invasive tool available to monitor both branches of the autonomic nervous system in humans. Power spectral analysis of short-term (5-min recordings) HRV provides a measure of autonomic modulation of the heart rate, and is widely used to assess autonomic nervous system activity (Rajendra et al., 2006; Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology, 1996).
The aim of this study was to assess the association between brain injury, vagus nerve activity, and innate immunity. To the best of our knowledge, this has never been studied in humans before. We determined HRV and measures of innate immunity in patients admitted to the neurological intensive care unit (ICU) with various forms of brain damage. Concentrations of circulating cytokines were analyzed, and in light of the spleen-mediated effects of the vagus nerve on circulating leukocytes, we also assessed the inflammatory response of circulating leukocytes in ex vivo stimulation experiments. HRV and immune parameters as well as the correlations between these parameters were analyzed in the entire group of patients, as well as in subgroups of patients, to compare brain injury associated with high and normal ICP.
Methods
Study population
We performed a prospective observational study in 34 adult patients admitted to the neurological ICU of a tertiary care university hospital in Nijmegen, the Netherlands in 2010 and 2011. Patient inclusion was aided by the help of nurses of the research-ICU department, which made rounds along the wards twice daily. The study protocol was approved by the Ethics Committee of the Radboud University Nijmegen Medical Centre, and complied with the Declaration of Helsinki, including current revisions and good clinical practice guidelines. The Ethics Committee waived the need for informed consent, because of the study's observational nature. All patients ≥18 years of age with traumatic and non-traumatic brain injury were eligible for the study. Patients were excluded in cases of known arrhythmias or a pre-existent immune-compromised status. One of the objectives of this study was to investigate the relationship between conditions associated with increased ICP, vagus nerve activity, and the innate immune response. Therefore we aimed to analyze three separate subgroups: SAH, intracranial hemorrhage (ICH), and ICU control patients that suffered from trauma without brain damage (Trauma). In our institute, most SAHs are treated with extraventricular drains, thereby alleviating increased ICP. Therefore this group of patients represents isolated brain injury without high ICP. Brain injury diagnoses were based on history, clinical examination, computed tomography, and computed tomography angiography. Baseline HRV data obtained in a group of 40 healthy male volunteers that participated in a human endotoxemia experiment (previously published in Kox et al., 2011b) were used for comparison. In addition, for comparison of ex vivo stimulation data, we performed whole blood stimulation experiments identical to those performed in patients in 4 healthy male volunteers on 4 different days, with each measurement separated by 5 days.
Data collection
The first measurement was performed within 24 h of the initial event. Patients were measured every 24 h for the first 4 days after admission to the ICU (days 0, 1, 2, and 3) unless they died or were discharged from the ICU. At these time points, blood was drawn for cytokine analysis and ex vivo stimulation and HRV were measured. Relevant medications, including antibiotics, sedatives, analgesics, and inotropic drugs were recorded on a daily basis. Furthermore, demographic and clinical data as well as in-hospital mortality were recorded.
Heart rate variability analysis
A 5-min 3-lead ECG recording was obtained using a Medilog AR12 recorder (Huntleigh Healthcare, Cardiff, U.K.). To ensure consistency, patients were not disturbed during recordings (no nursing or medical interventions were done), and no medication was altered or started. R-peak position was determined at a sample rate of 4096 Hz. HRV was analyzed using dedicated software (Medilog Darwin HRV; Huntleigh Healthcare). In each 5-min recording, QRS complexes were detected and only normal-to-normal beat (NN) intervals were tabulated, yielding an interval tachogram. In each 5-min recording, artifacts such as premature (supra)ventricular beats or other arrhythmias were detected by the software, and each recording was also visually inspected to ensure good quality of the recordings. Recordings with artifacts such as premature (supra)ventricular beats or other arrhythmias comprising more than 5% of the total epoch were discarded. After linear detrending, power spectral density was determined by fast Fourier transformation (FFT) of interval tachograms using the Welch method and a FFT width of 1024. Very-low-frequency (VLF) power (0.0033–0.04 Hz) cannot be reliably obtained from 5-min recordings and was therefore not analyzed (Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology, 1996). With regard to the expected differences in total power between patients and healthy volunteers, and large variations in spectral power between patients, only normalized units or ratios are meaningful for comparison. Therefore, normalized units of high-frequency power (HFnu), a measure of cardiac vagus nerve activity (Malliani et al., 1991,2005; Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology, 1996), and the low-frequency:high-frequency ratio (LF:HF), which is thought to reflect either sympathovagal balance or sympathetic nervous system activity (Godin et al., 1996; Pelosi et al., 1999; Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology, 1996; Yang et al., 2000), were used to assess autonomic activity. The physiological correlate of LF:HF is debated, because the LF component has been reported to be jointly contributed to by both sympathetic and parasympathetic activity (Berger et al., 1989). In the remainder of this article, we will refer to it as an index of sympathovagal balance, because it is mostly used as such. HRV indices, their frequency ranges, physiological correlates, and methods of calculation are listed in Table 1.
ms, milliseconds; AU, arbitrary units.
Ex vivo whole blood stimulation
Ex vivo blood stimulation was performed using a custom method we developed in house. This system consists of tubes pre-filled with 2 mL of culture medium or 2 mL of culture medium containing lipopolysaccharide (LPS). The pre-filled tubes were stored at −80°C and brought to room temperature before use. Then 0.5 mL of lithium-heparin anticoagulated blood (Vacutainer System; BD Biosciences, Plymouth, U.K.) was added to the pre-filled tubes and the tubes were gently inverted to ensure proper mixing. Subsequently, the tubes were incubated for 24 h at 37°C, centrifuged (2500g at 4°C for 10 min), and the supernatant was stored at −80°C until analysis. In non-stimulated samples, no cytokine production was observed in any of the samples from patients or healthy volunteers, ruling out contamination.
Reagents
Culture medium (RPMI 1640 Dutch modification) was obtained from ICN Biomedicals (Costa Mesa, CA). Culture medium was supplemented with gentamicin 10 μg/mL, L-glutamine 10 mM, and pyruvate 10 mM. Escherichia coli LPS (serotype O55:B5) was purchased from Sigma-Aldrich (St. Louis, MO). LPS was further purified as described previously (Hirschfeld et al., 2000), and used at a final concentration of 10 ng/mL.
Cytokine measurements
For plasma cytokine determination, EDTA-anticoagulated blood was collected from the patient's arterial catheter and immediately centrifuged at 2000g for 10 min at 4°C. Plasma was stored at −80°C until analysis. Plasma concentrations of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and IL-10 were measured using a simultaneous Luminex Assay according to the manufacturer's instructions (Milliplex; Millipore, Billerica, MA). Cytokine concentrations in ex vivo-stimulated whole blood supernatants were determined by enzyme-linked immunosorbent assay (ELISA) according to the manufacturer's instructions (IL-6 and IL-10: Sanquin, Amsterdam, The Netherlands; TNF-α: R&D Systems, Minneapolis, MN).
Statistical analysis
None of the measured parameters was normally distributed (calculated using the Shapiro-Wilk test). Therefore, data are represented as median (range). The statistical tests used are indicated in the text and figure legends. Differences between two groups were tested with Mann-Whitney U tests for continuous data, or Fisher's exact tests for binominal data. For differences between three or more groups/days, Kruskal-Wallis tests with Dunn's post-hoc test (corrected for multiple comparisons) were used. Spearman's correlation was used unless specified otherwise. p Values of correlation analyses in subgroups were corrected for multiple testing. p Values of 0.05 or lower were considered significant. Statistical analyses was performed using GraphPad Prism 5 (GraphPad Software, San Diego, CA) and SPSS 16.0 (SPSS Inc., Chicago, IL).
Results
Study population
Demographic characteristics of the patient population and healthy volunteers are shown in Table 2. Physiological parameters and relevant medications are listed in Supplementary Table 1 (see online supplementary material at
Data are presented as median (range).
The miscellaneous group consisted of multi-trauma patients with neurological damage (n=3), post-anoxic encephalopathy (n=5), transverse myelitis (n=2), cerebellar infarction (n=1), and intoxication (n=1).
p Values for all patients versus healthy volunteers: age p<0.0001; sex p<0.0001. p Values between patient groups: age p<0.05; Trauma versus SAH, p<0.01; Trauma versus ICH; APACHE-II p<0.05; Trauma versus SAH and ICH; SAPS p=NS; GCS p<0.05; Trauma versus ICH; sex p<0.05; SAH versus miscellaneous; in-hospital mortality p=NS.
SAH, subarachnoid hemorrhage; ICH, intracranial hemorrhage (left hemisphere n=2, right hemisphere n=1, bilateral n=1, central n=2); APACHE-II, Acute Physiology, Age, and Chronic Health Evaluation-II score.
HRV and inflammatory markers in the entire study population
In the entire patient population (n=34), there were no differences in any of the HRV parameters between days. Therefore, data from all 4 days were pooled. HRV total spectral power (an overall measure of variability) was diminished in patients compared to healthy volunteers (Fig. 1A). As depicted in Figure 1B and C, HFnu (cardiac vagus activity) was significantly higher, and LF:HF (sympathovagal balance) was significantly lower in patients compared to healthy volunteers, indicating increased cardiac vagus nerve activity. Similarly to HRV data, there were no differences over time in ex vivo-stimulated cytokine production in both healthy volunteers and patients, and therefore the data were pooled. Ex vivo-stimulated whole blood of patients revealed distinct hyporesponsiveness to LPS compared to healthy volunteers (TNF-α shown in Fig. 1D; IL-6: healthy volunteers 9254 [3548–17670] pg/mL versus patients 1800 [8–16885] pg/mL, p<0.0001; IL-10: healthy volunteers 293 [120–404] pg/mL versus patients 47 [12–366] pg/mL, p<0.0001).

Heart rate variability (HRV) parameters total spectral power (
In all healthy volunteers, plasma cytokine levels were below detection limits. In patients, plasma IL-6 and IL-10 concentrations were significantly higher on the day of admission and decreased afterwards (Fig. 2); this pattern was due to the high plasma concentrations seen in the Trauma group on the day of admission, as described later. Plasma TNF-α levels were relatively low and did not change over time (6 [2–43] pg/mL).
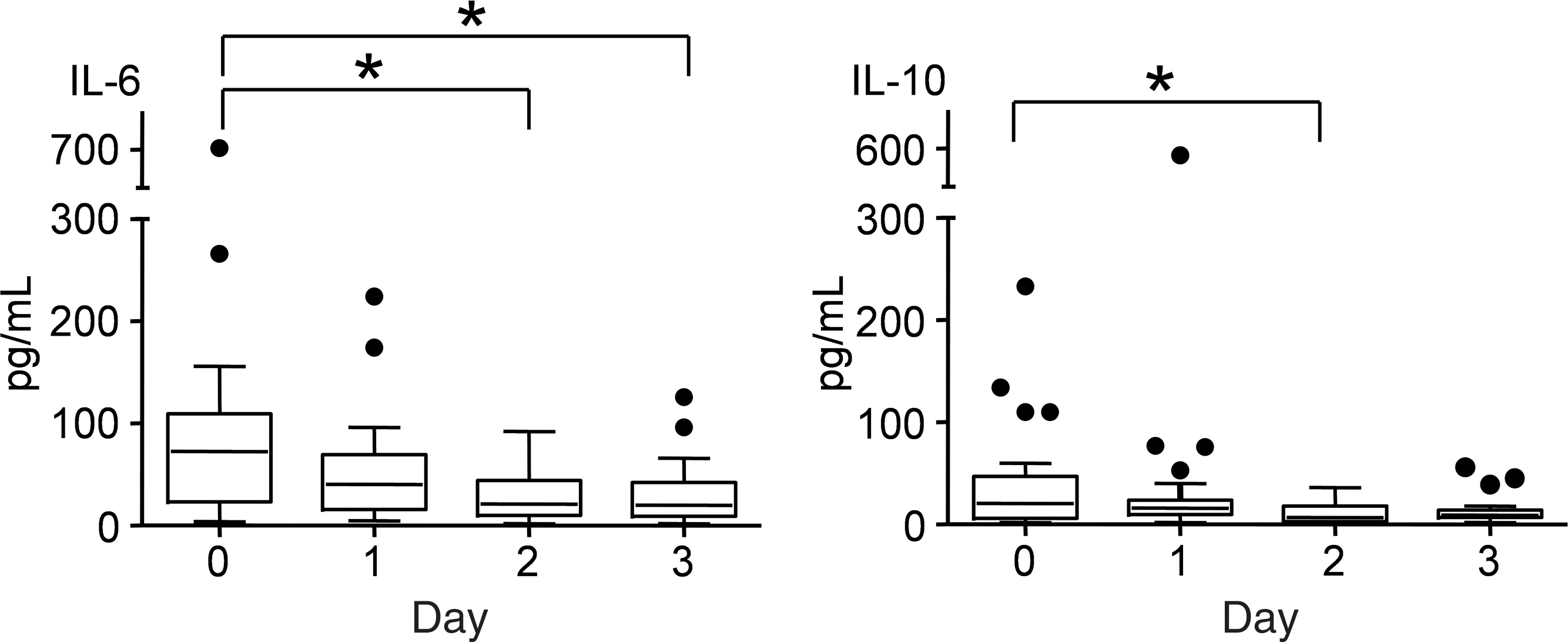
Concentrations of the plasma cytokines interleukin-6 (IL-6) and IL-10 during the study period in the entire patient population. Data are represented as boxes and whiskers (Tukey, *p<0.05).
To investigate the relationship between autonomic activity and ex vivo-stimulated cytokine production or plasma cytokines, we correlated HFnu and LF:HF with these inflammatory parameters, and found a significant inverse correlation between HFnu and ex vivo-stimulated TNF-α production (r=−0.22, p=0.025).
HRV and inflammatory markers in subgroups of patients
No differences in HRV parameters between days 0 and 4 were observed in patients in the SAH, ICH, or Trauma subgroups. Therefore, all HRV data were pooled. No significant differences in HRV parameters were found between the SAH, ICH, or Trauma subgroups. Furthermore, no differences in ex vivo-stimulated cytokine production between days 0 and 4 were observed within subgroups, and the data were pooled. Ex vivo production of both pro-inflammatory (TNF-α and Il-6) and anti-inflammatory (IL-10) cytokines in response to LPS stimulation was significantly lower in patients in the ICH group compared to the other two groups (Fig. 3). Furthermore, stimulated IL-10 levels were lower in the Trauma group compared to the SAH group. Ex vivo cytokine production in all patient subgroups was significantly lower compared with healthy volunteers. The markedly suppressed ex vivo cytokine production in the ICH group compared to the other two subgroups could not be explained by large differences in immunomodulatory covariates, such as anti-inflammatory medication or leukocyte counts (Supplementary Table 1; see online supplementary material at
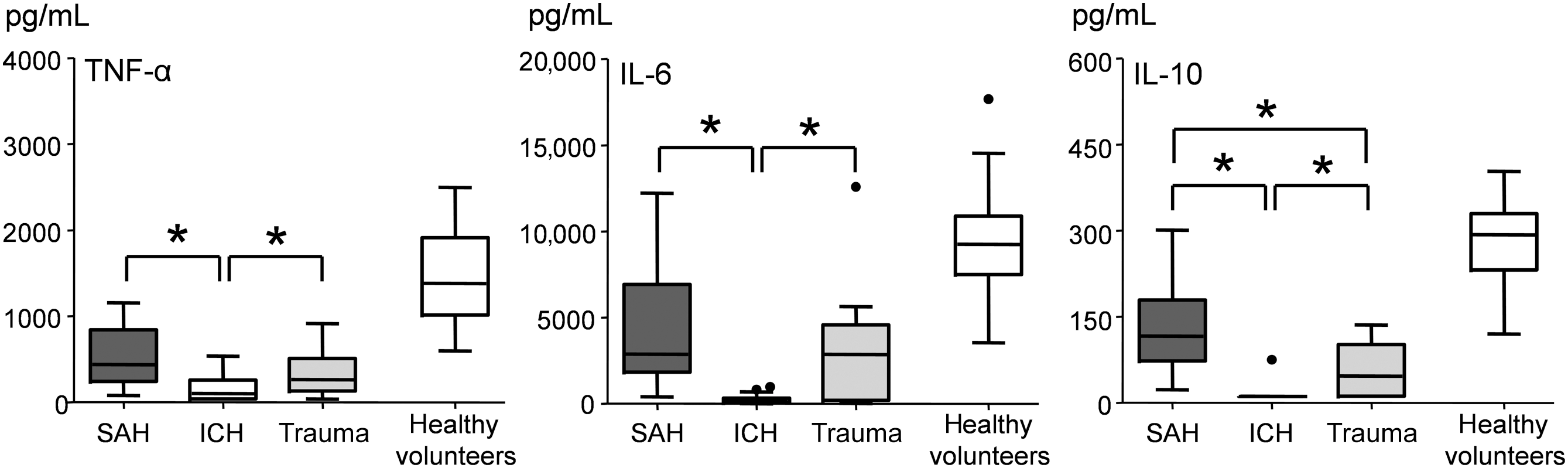
Tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and IL-10 production in whole blood of patients and healthy volunteers ex vivo-stimulated with lipopolysaccharide. Data are represented as boxes and whiskers (Tukey test). Ex vivo stimulation was performed on 4 different days in both patients and healthy volunteers and these data were pooled (*p<0.05). Cytokine production in all patient subgroups was significantly lower compared with healthy volunteers, but for reasons of clarity, this is not indicated in the figure (SAH, subarachnoid hemorrhage; ICH, intracranial hemorrhage).
Within the Trauma group, plasma levels of IL-6 were significantly higher on day 0 (146 [83–702] pg/mL) compared with day 2 (48 [13–82] pg/mL, p<0.05). In the other two groups, no differences in plasma cytokine levels between days were found. Pooling of the data from the different days revealed significantly higher plasma levels of TNF-α and IL-6 in the Trauma group compared to the other two groups (TNF-α: Trauma 8 [3–15], SAH 4 [2–11], ICH 5 [3–19] pg/mL; IL-6: Trauma 83 [13–702], SAH 26 [2–131], ICH 17 [2–79], p<0.05). Plasma IL-10 levels were higher in the Trauma group compared to the SAH group (Trauma: 20 [2–110]; SAH: 6 [2–110] pg/mL; p<0.05). The elevated cytokine levels in the Trauma group, especially on the day of admission, were likely due to the trauma insult itself.
We further examined the relationship between vagus nerve activity and inflammatory parameters in the ICH group, since this group exhibited greatly suppressed ex vivo-stimulated cytokine production. Correlation analysis between HFnu and ex vivo-stimulated cytokine production was not feasible in light of the very low levels of cytokines seen in the stimulated samples (approximately 50% of samples were below the detection limit). However, we did find a strong inverse correlation between HFnu and plasma TNF-α levels, indicating that higher cardiac vagus nerve activity is associated with lower plasma TNF-α levels (Fig. 4). No such correlations were found in the other subgroups, but this could not be explained by large differences between subgroups in cardiorespiratory parameters or medications that could influence HRV (Supplementary Table 1; see online supplementary material at
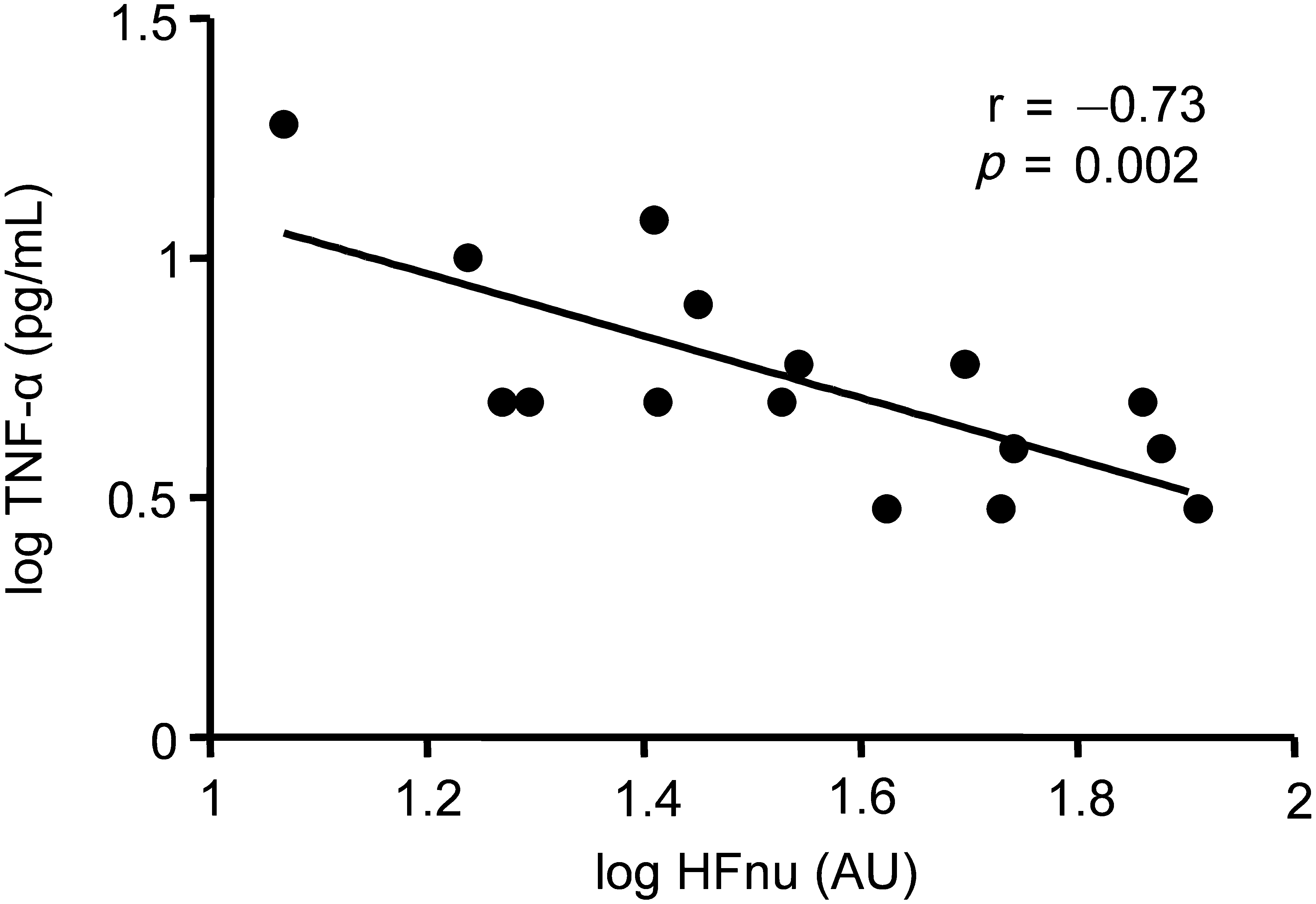
Correlation between normalized units of high-frequency power (HFnu) and plasma tumor necrosis factor-α (TNF-α) in intracranial hemorrhage patients. For visual purposes, data were log-transformed and Pearson r and p values are indicated in the graph. Correlation analysis was also performed on non-log-transformed data: Spearman r: −0.70, p=0.004. (AU, arbitrary units.)
Discussion
In the present study, we report increased HFnu levels, a measure of cardiac vagus nerve activity, in patients with brain damage compared to healthy volunteers. Furthermore, these patients exhibited a distinct immune paralysis, as reflected by low levels of cytokine production in ex vivo-stimulated whole blood. Higher levels of HFnu correlated with lower levels of ex vivo-stimulated TNF-α production. The most pronounced immune paralysis was found in patients with intracranial hemorrhage, a condition associated with high ICP. In these patients, higher levels of HFnu strongly correlated with lower levels of plasma TNF-α. These observations suggest that brain injury, and especially conditions associated with increased ICP, is associated with vagus nerve-mediated immune suppression.
We found markedly lower levels of overall HRV in our patient population compared to healthy volunteers. This is in accordance with previous findings of various investigators, showing that brain injury is associated with diminished HRV, and that low HRV predicts unfavorable outcome (Baguley et al., 2006; Haji-Michael et al., 2000; Papaioannou et al., 2008). The observed decrease in physiological heart rate variability indicates the uncoupling of organ systems generally seen in critically ill patients (Godin and Buchman, 1996; Godin et al., 1996). Sedatives and analgesics have been shown to contribute to the loss of variability (Estafanous et al., 1992; Kanaya et al., 2003); however, as large decreases in HRV are also found during experimental human endotoxemia (LPS administration in healthy volunteers; Godin et al., 1996; Jan et al., 2010; Kox et al., 2011b), other factors must also play a role. Brain injury patients exhibited higher HFnu and lower LF:HF ratios compared to healthy volunteers, suggesting a shift in the balance between cardiac autonomic activity towards increased vagus nerve signaling. These results are in accord with previous findings of parasympathetic predominance in the acute phase following SAH (Kawahara et al., 2003) and severe head injury (Winchell and Hoyt, 1997). Our findings cannot be explained by the considerably higher age of the patient population compared to the healthy volunteers, as increased age has been linked to higher LF:HF ratios (Antelmi et al., 2004).
Ex vivo stimulation of whole blood with LPS revealed that both pro-inflammatory and anti-inflammatory cytokine production was severely impaired in patients compared with healthy volunteers. These findings suggest that following brain injury, monocytes (the main producers of the cytokines studied in 24-h whole blood LPS stimulation experiments; Dinarello, 1991) have a reduced capacity to respond to pathogens. Our findings are therefore indicative of a distinct immune paralysis in brain injury patients. To the best of our knowledge, this has never been studied in brain injury patients before. A few studies have investigated effects of severe head injury on T-lymphocytes and found decreased T-helper cell activation, attenuated lymphokine-activated killer cell cytotoxicity, and depression of PHA-induced pro-inflammatory cytokine production (IL-2 and interferon-γ; Miller et al., 1991; Quattrocchi et al., 1990,1991,1992).
In our patient population, we found a significant inverse correlation between cardiac vagus nerve activity (as reflected by HFnu) and ex vivo TNF-α production in response to LPS. In particular, TNF-α production has been shown to be greatly limited by vagus nerve stimulation in animal models (Borovikova et al., 2000; Wang et al., 2003). Our findings therefore suggest activation of the cholinergic anti-inflammatory pathway, in which increased vagus nerve input to the spleen results in the “education” of passing circulating leukocytes to release lower levels of inflammatory cytokines (Huston et al., 2006,2009). Our findings are in accord with a recent study in healthy volunteers, in which significant inverse correlations of roughly the same magnitude were found between HF power and ex vivo LPS-stimulated TNF-α production (Marsland et al., 2007). Only one study to date has investigated the effects of electrical vagus nerve stimulation on ex vivo cytokine production in humans (De Herdt et al., 2009). This investigation in refractory epilepsy patients with an implanted vagus nerve stimulator demonstrated a significant reduction in LPS-stimulated IL-8 production after 6 months of vagus nerve stimulation. No significant effects were observed after 3 weeks of stimulation, although interestingly, a trend towards lower levels of TNF-α was present.
Our study was strictly observational; therefore standard care was not altered for study purposes. Unfortunately, ICP was only measured in 2 patients on the day of admission. As this was anticipated, we decided prospectively to create subgroups of brain injury that are associated with increased and normal ICPs. A significantly more pronounced immune suppression was found in the ICH group compared with the SAH and Trauma groups. This is supportive of our hypothesis that high ICPs result in more pronounced immune suppression. In the ICH group, high ICPs were suspected based on the space-occupying lesions that were seen on CT imaging of the brain, whereas in the Trauma group without brain damage, and the SAH group in which ICP was alleviated by the placement of extraventricular drains in all but one of the patients, ICP was suspected to be lower. Although HFnu levels in the ICH group were not significantly higher compared to the other groups, we found a strong inverse correlation between HFnu and plasma TNF-α concentrations, indicating anti-inflammatory effects of increased vagus nerve activity. In accordance with this, correlations between higher vagal HRV parameters and lower plasma levels of inflammatory mediators have been found in both healthy volunteers (Sloan et al., 2007), and coronary heart disease patients (Frasure-Smith et al., 2009).
Our study has several limitations. First, HRV only reflects cardiac autonomic activity. At present, it is unclear whether cardiac vagus nerve activity reflects vagal input to other organs that are important in the immune response (e.g., the spleen). Animal and human studies have established that the output of the sympathetic nervous system is highly differentiated (Niijima et al., 1991; Rogausch et al., 1997; Sayk et al., 2008). No such investigations of the parasympathetic nervous system are known to us, but it appears plausible that differentiated output to organs does exist. Unfortunately, besides HRV, no techniques to measure vagus nerve activity are currently available in humans. Second, the effects of ICP on vagus nerve activity and the immune response could not be properly assessed in this study, since ICP probes were placed in only 2 of our patients. Third, the question remains whether our findings in circulating monocytes can be extrapolated to tissue-resident macrophages, which are the predominant cells involved in the initial innate immune response against invading pathogens. We have recently shown that after administration of LPS to healthy volunteers, ex vivo hyporesponsiveness to LPS, reflecting the monocyte compartment, is present in the acute phase but quickly wanes (Kox et al., 2011a). However, in vivo hyporesponsiveness to LPS, reflecting the tissue-resident macrophage compartment, persists for several weeks (Kox et al., 2011a). In this respect, ex vivo hyporesponsiveness might even underestimate the capacity of tissue-resident macrophages to respond to inflammatory stimuli. Finally, demographic characteristics may have influenced our results. For instance, another contributor to the observed low HRV seen in the patient population compared to healthy volunteers could be the significantly higher age of the patient population, as increasing age is associated with decreased HRV (Antelmi et al., 2004; Bigger et al., 1995). Differences in age and other factors that might influence autonomic modulation, such as disease severity, were present between patient groups as well. However, no differences in HRV indices were found between these groups, suggesting that these factors did not have a major impact on the results of this study.
In conclusion, in brain injury patients, higher vagus nerve activity is associated with attenuated ex vivo LPS-stimulated monocytic TNF-α production. Furthermore, in ICH patients, in association with high ICPs, a pronounced ex vivo immune paralysis was observed. In addition, in these patients higher vagus nerve activity was also associated with lower levels of circulating TNF-α. These findings are suggestive of effects of the cholinergic anti-inflammatory pathway, and might contribute to the immune paralysis seen in brain injury patients.
Footnotes
Acknowledgments
The authors would like to thank the research nurses of the intensive care department for patient inclusion, as well as Ebru Cevik and Maarten van den Berg for patient measurements.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.