
Abstract
Permanent olfactory dysfunction can often arise after traumatic brain injury (TBI) and while one of the main causes is the immediate loss of neurons in the olfactory bulb (OB), the emergent neuroinflammatory environment following TBI may further promote OB deterioration. Therefore, we examined the effects of acute anti-inflammatory treatment with minocycline on post-TBI olfactory behavior and on OB surface. The mouse model of closed-head injury by mechanical percussion was applied to anesthetized Swiss mice. The treatment protocol included three injections of minocycline (i.p.) at 5 min (90 mg/kg), 3 h, and 9 h (45 mg/kg) post-TBI. An olfactory avoidance test was run up to 12 weeks post-TBI. The mice were then sacrificed and their OB surface was measured. Our results demonstrated a post-TBI olfactory behavior deficit that was significant up to at least 12 weeks post-TBI. Additionally, substantial post-TBI OB atrophy was observed that was strongly correlated with the behavioral impairment. Minocycline was able to attenuate both the olfactory lesions and corresponding functional deficit in the short and long term. These results emphasize the potential role of minocycline as a promising neuroprotective agent for the treatment of TBI-related olfactory bulb lesions and deficits.
Introduction
T
Olfaction involves various processing stages spanning from sensory neuron input to the olfactory bulb (OB), to ultimately downstream neurons in different brain compartments, such as the olfactory cortex, amygdala, hippocampus, and hypothalamus (Wachowiak and Shipley, 2006; Wilson and Mainen, 2006). It is generally assumed that the primary causes of post-TBI olfactory loss include overextension, distortion, or tearing of the olfactory nerves due to the abrupt movement of the brain, as well as contusion, hemorrhage, or edema in the OB and/or the orbitofrontal regions related to olfaction (Ikeda et al., 1995). Due to the localization of the olfactory system, olfactory dysfunction can easily arise following frontal, temporal, or occipital impacts (Doty et al., 1997; Fujii et al., 2002) of severe, moderate, or even mild severity (Fortin et al., 2010). Yet the possibility of olfactory impairment is significantly increased in the case of severe TBI compared to milder cases (Callahan and Hinkebein, 2002). While the olfactory system has a remarkable capacity for neural regeneration, the prognosis for recovery following head injury is relatively poor (Jimenez et al., 1997; London et al., 2008).
Despite rapidly progressing research, treatment of olfactory dysfunction remains limited, with systemic steroids being the only treatment of therapeutic potential to date (Jiang et al., 2010). Yet even the latter does not appear to apply adequately in the case of post-traumatic anosmia (Duncan and Seiden, 1995). Hence a better understanding of the mechanisms of injury and opportunities for intervention is required before progress can be made in improving olfactory outcome. Recent studies have associated olfactory dysfunction with the presence of a local inflammatory response (Kobayashi et al., 2009).
TBI gives rise to a pronounced inflammatory reaction that represents a coordinated effort by resident microglia and peripheral blood leukocytes to protect the brain after trauma (Kelley et al., 2007). In fact, it has been recently demonstrated that severe injury to the olfactory nerves and the OB produces a rapid and pronounced neuroinflammatory reaction, with ensuing glial scar formation, that is strongly correlated with poor functional outcome (Kobayashi et al., 2009). Moreover, there is an abundance of experimental evidence indicating that intense neuroinflammation, as in the case of TBI, is detrimental for adult neurogenesis both in the intact and injured brain (for a review see Ekdahl et al., 2009).
Therefore, it seems that the degree of post-TBI neuroinflammation may play a critical role in predicting olfactory outcome, and that its early and efficient inhibition is likely to allow the sparing of olfactory bulb tissue and preserve olfactory function. One well-tolerated compound whose anti-inflammatory properties have been widely demonstrated in different models of TBI and of various neurodegenerative diseases is minocycline, a highly lipophilic, second-generation tetracycline. According to the literature, minocycline reduces post-TBI apoptosis and neuroinflammation, while improving neurological outcome (Bye et al., 2007; Sanchez-Mejia et al., 2001). Moreover, our laboratory has recently shown that a short-term treatment protocol with minocycline is able to suppress neuroinflammation and microglial activation, while reducing edema, brain lesion volume, and locomotor hyperactivity in a mouse model of frontal closed-head injury (Homsi et al., 2009, 2010; Siopi et al., 2011). Since one of the main triggers of hyperactivity in mice is the presence of olfactory bulb lesions, we wondered whether the same treatment protocol with minocycline is equally able to preserve olfactory bulb tissue and its function post-TBI.
In an attempt to answer this question, we used the mouse model of frontal closed-head injury by mechanical percussion, as previously described (Homsi et al., 2009,2010) to study the effects of minocycline on post-TBI olfactory behavior and OB integrity. The results obtained in this study yield considerable proof that frontal closed-head injury deteriorates OB tissue and affects olfactory behavior in mice, and that a short-term treatment with minocycline is able to reduce the level of tissue damage and improve recovery in a lasting manner.
Methods
Animals
All experiments were performed on male Swiss mice (Janvier, Le Genet St. Isle, France) weighing 28–30 g, housed in a controlled temperature environment (22±2°C), under a 12-h light/dark cycle, with access to food and water ad libitum. Animal care and experiments were approved by the University Paris Descartes Animal Ethics Committee (P2.MJT.049.08), French regulations, and the European Communities Council Directive of November 24, 1986 (86/609/EEC) on the protection of animals for experimental use.
Mouse model of closed-head injury by mechanical percussion
The mouse model of closed-head injury was performed as previously described (Hellal et al., 2003; Homsi et al., 2009). The mice were anesthetized with 2% halothane before being subjected to TBI. Closed-head trauma was induced by a 50-g weight dropped from 36 cm onto the right side of the head. This model closely mimics closed-head injury in motor vehicle accidents or falls, although skull integrity is generally preserved with no sign of fracture. It generates an intense neuroinflammatory response associated with edema formation, contusion, hemorrhage, and diffuse axonal injury, and is generally associated with a 5–15% mortality rate within the first 5 min following the impact (Homsi et al., 2009,2010).
Treatment protocol with minocycline
The treatment protocol used in this study has been previously shown to produce anti-inflammatory and anti-edematous effects and promote functional recovery in the same model of TBI (Homsi et al., 2009,2010). Minocycline was administered i.p. at 5 min post-TBI (90 mg/kg), then at 3 h (45 mg/kg), and at 9 h (45 mg/kg) post-TBI. The animals were divided into three groups (n=12 per group): a group of naive mice, and two groups of mice that underwent TBI and received either minocycline or its vehicle (phosphate-buffered saline, PBS; 0.01 M, pH 7.4).
Olfactory avoidance test procedure
This test was designed to identify the ability to sense an aversive scent. While rodents demonstrate variable attractive behaviors to food smells (Nyby et al., 1985), they generally show avoidance behaviors towards spoiled smells, for example aliphatic acids (Hebb et al., 2002). Acetic acid is a short aliphatic acid that exerts a pungent, non-predatory odor (Ai et al., 2010); hence it is perceived as sharp or irritating for mice as well as for humans.
The test arena used was a clean transparent cage (14 cm H×19 cm W×35 cm L) without bedding. A black posterboard was placed on the bottom and back side of the cage. The top of the cage was covered with a filtered lid to allow normal air circulation inside the compartment. An appropriate number of filter paper squares (5 cm×5 cm) were prepared in advance. After habituation of the mouse in the test arena for 10 min, two filter papers, one impregnated with water and the other with acetic acid diluted in water to 2.5%, were introduced in successive 3-min trials with 1-min intervals, starting with water as an odorless control. The behavior of the mice was recorded using a video recording device placed near the far end of the cage. Arena cages were cleaned with water before each trial. Testing took place between 10
Preparation of brain samples and quantification of the olfactory bulb surface
At 12 weeks post-TBI, all three groups of mice were anesthetized with sodium pentobarbital (60 mg/kg i.p.) and perfused transcardially with heparinized saline (30 mL/mouse), followed by PBS containing 4% paraformaldehyde (PFA; 50 mL/mouse). The brains were removed cautiously and kept overnight in 4% PFA. In order to measure OB tissue loss, a photo of the upper view of each brain was taken with a digital camera. The ipsilateral and contralateral olfactory bulb surfaces were measured using ImageJ software (National Institutes of Health). The total olfactory bulb surface was then measured for each animal by addition of the ipsilateral and contralateral olfactory bulb surfaces.
Statistical analysis
Data were expressed as mean±standard error of the mean of n observations, where n represents the number of animals used. The data collected from our behavioral and histological studies were analyzed using Statview® statistical software (Statview 5.0; Abacus Concepts Inc., Berkeley, CA), A two-way (group and time post-injury) repeated-measures analysis of variance (ANOVA) was used, and intergroup differences were analyzed by ANOVA, followed by Bonferroni's multiple comparison test at each corresponding time point. Differences with a p value < 0.05 were considered statistically significant. In order to examine whether there is an association between OB volume and olfactory outcome, the corresponding data from all samples (n=34) were used in a Pearson's correlation test.
Results
TBI alters olfactory avoidance behavior in mice
As shown in Figure 1, naïve and TBI mice demonstrate similar olfactory behavior towards water (15.9±4.7 versus 16.0±4.4 sec). However, in the presence of acetic acid, naive mice spent significantly less time investigating the scented paper (3±0.7 sec) than TBI mice (15.6±6 sec, p<0.05), thus demonstrating a more significant aversive behavior than injured mice.
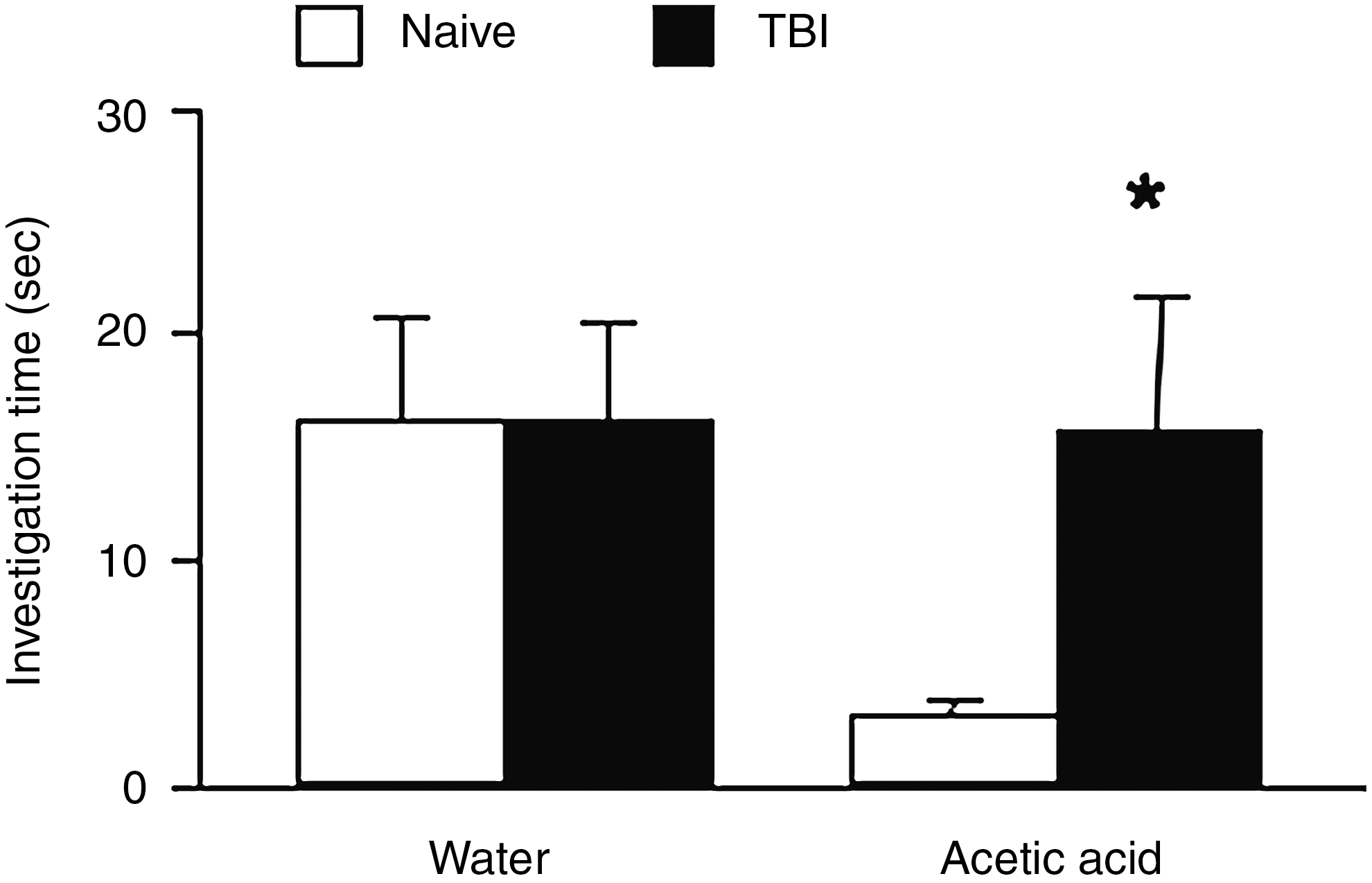
The olfactory avoidance test was carried out at 10 days post-TBI. The duration of the time mice spent investigating the scented filter paper was measured for water and acetic acid (2.5%). Mean investigation times (sec±standard error of the mean) are shown for each odorant during the 3-min testing period with naive and traumatic brain injury (TBI) mice (n=10/group; *p<0.05 versus naive).
Minocycline improves olfactory avoidance behavior from 2 to at least 12 weeks post-TBI
Since our olfactory test exposed a failure of TBI mice to demonstrate an aversive behavior to acetic acid, we used another set of animals to perform an olfactory avoidance test from 1 to 12 weeks post-TBI. Both investigation time and number of investigations of the scented filter papers were measured. Naïve and TBI mice investigated the odorless water-impregnated paper a similar number of times at all time points of the test (Fig. 2). However, in the presence of acetic acid, vehicle-treated mice investigated the filter paper a greater number of times than naive mice, demonstrating a deficit in olfactory avoidance behavior. The differences between naive and TBI mice were most prominent from 2 weeks of the test (p<0.05) up to at least 12 weeks post-TBI (p<0.001). Interestingly, contrary to untreated TBI mice, injured mice treated with minocycline showed greater aversive behavior towards acetic acid, that was prominent from 8 weeks (p<0.05 versus vehicle) up to at least 12 weeks post-TBI (p<0.01 versus vehicle). Moreover, as shown in Fig. 2, while water investigation time was not significantly different between naïve and TBI mice, investigation of the acid-scented paper was divergent in most time points of the test, with vehicle-treated mice exploring the acetic acid for longer periods of time than naïve mice (p<0.05). Likewise, minocycline-treated mice showed greater aversion towards acetic acid than vehicle-treated mice (p<0.05). However, due to the great variability associated with this parameter, the differences between the three groups of mice were not statistically significant at each time point.

The long-term effects of minocycline on the number of investigations of water (
Minocycline reduces olfactory bulb lesion surface at 12 weeks post-TBI
At 12 weeks post-TBI, mice were sacrificed and their OB surface was measured. As shown in Figure 3, TBI caused marked tissue loss in both the ipsilateral and contralateral OB (ipsilateral OB: 2.7±0.6 versus 5.8±0.2 mm2, p<0.001; contralateral OB: 3.7±0.6 versus 5.8±0.2 mm2, p<0.01; total OB surface: 6.5±0.7 mm2 versus 11.6±0.4, p<0.001 versus naive mice). Remarkably, the OB of mice treated with minocycline were considerably preserved compared to vehicle-treated TBI mice (ipsilateral OB: 5.3±1.9 mm2 versus 2.7±0.6, p<0.01; contralateral OB: 4.9±0.3 mm2 versus 3.7±0.6, p=0.06; total OB surface: 10.2±0.8 versus 6.5±0.7 mm2, p<0.01).
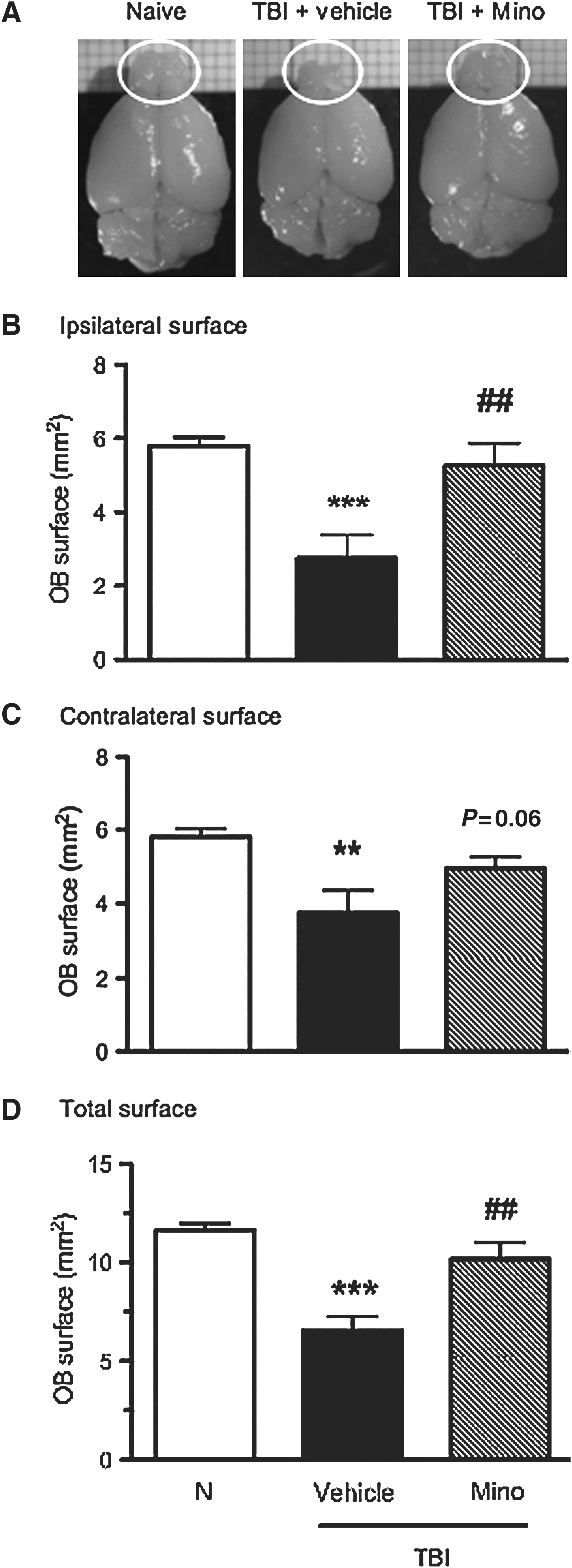
Representative upper views of the olfactory bulbs of naive, vehicle- and minocycline-treated traumatic brain injury (TBI) mice at 12 weeks following trauma (
Olfactory avoidance behavior is strongly correlated with OB surface at 12 weeks post-TBI
In order to verify the validity of our olfactory test, we evaluated whether olfactory performance was correlated with OB surface. As shown in Figure 4, there was no correlation between OB surface and olfactory behavior towards water. Besides, since water is an odorless molecule, it is reasonable to presume that the behavior of mice towards water was not influenced by olfactory acuity. However, in the case of acetic acid, OB surface and olfactory behavior were significantly correlated, both for investigation time (r=−0.49; p=0.0042), and the number of investigations (r=−0.62; p=0.0001), showing that mice with greater OB tissue loss exhibited diminished aversive behavior towards an unpleasant odor.

Correlation study between total olfactory bulb surface and olfactory behavior (number and time of investigations) with water (
Discussion
The findings of this study provide robust evidence that frontal closed-head injury in the mouse results in the formation of substantial OB lesions that persist up to at least 12 weeks post-TBI, and that are strongly correlated with an olfactory deficit, namely a decreased ability to sense an aversive odor. Interestingly, an acute treatment with minocycline was able to considerably reduce OB tissue loss and attenuate the ensuing olfactory impairment in the short and long term following injury.
Primarily, our findings provide proof that our experimental TBI model is eligible to study olfactory dysfunction, reflected by a diminished aversion to acetic acid, not only in the early time points, but also in a late TBI stage. While no study hitherto has evaluated olfactory behavior in experimental TBI conditions, various studies have examined olfactory acuity and function in different models of neurodegenerative diseases or region-specific ablation mice (Fleming et al., 2008; Kobayakawa et al., 2007; Prediger et al., 2011; Wesson et al., 2010). On the detection of a novel odor in their environment, mice reliably display an arousal-related orienting response involving sniffing and whisker movements in the direction of the odor, mostly used for the acquisition of odor information (Wachowiak et al., 2009). Since a single sniff of an odor is sufficient to yield even complex odor detection and discrimination (Kepecs et al., 2006), it can be argued that rapid and prolonged sniffing is a sign of an odor discrimination and/or detection deficit. This test can be highly advantageous in testing the efficacy of a specific treatment against TBI-induced olfactory dysfunction. Interestingly, unlike the odorless water-impregnated paper, both the duration and number of investigations of the acid-scented paper were strongly correlated with OB surface. These results emphasize the validity of our olfactory test, and are in accordance with various clinical studies demonstrating that post-traumatic OB volume seems to decline parallel to olfactory function (Buschhüter et al., 2008; Collet et al., 2009; Haehner et al., 2008; Yousem et al., 1996).
Furthermore, the essential findings of this study are in accordance with numerous clinical data suggesting that OB lesions and subsequent olfactory dysfunction are frequent following frontal and occipital brain injuries of different degrees of severity (Callahan et al., 2002; Fortin et al., 2010; Fuji et al., 2002; Haxel et al., 2008; Joung et al., 2007; Wu and Davidson, 2008). This high prevalence seems reasonable, considering that localization of the OB and olfactory cortex in the rostral part of the brain renders them highly vulnerable to frontal impacts (Doty, 2001). Since our experimental TBI model generates moderate to severe frontal closed-head injury with a pronounced contre-coup lesion (Hellal et al., 2003), we can logically assume that the force and site of the mechanical impact were mainly responsible for the significant OB tissue loss observed in this study. It is also worth noting that in our experimental conditions, TBI provokes a significant increase in locomotor activity that is discernible within the first days following trauma up to at least 12 weeks post-TBI (Homsi et al., 2010). Interestingly, various experimental studies have demonstrated that olfactory bulb lesions provoke significant hyperactive responses in mice and rats (Kang et al., 2010; Mucignat-Caretta et al., 2006; Viggiano, 2008; Zueger et al., 2005). Yet, there are some contradictory data showing that OB ablation does not alter activity levels in the open field, and that olfactory input disruption provokes hypoactive responses instead (Mucignat-Caretta et al., 2004; Slotnick and Gutman, 1977).
In our experimental model, neuroinflammation is very much present shortly after TBI, and is characterized by amplified microglial activation, excessive release of interleukin-1β and edema formation (Homsi et al., 2009,2010). Apart from promoting neuronal death (Glass et al., 2010), brain inflammation can be also detrimental to adult neurogenesis, since excessive microglial activation promotes the death of newly-formed neurons (Ekdahl et al., 2003,2009). Hence, inhibition of post-TBI neuroinflammation, with respect to the timing and extent of the neuroinflammatory cascade, can exert neuroprotective effects by interfering with neuron apoptosis, and by enhancing the production of neurotrophic factors. Interestingly, we have demonstrated that acute treatment with minocycline exerts not only anti-inflammatory and anti-edematous, but also neuroprotective effects in this TBI model, by reducing brain lesion volume and promoting functional recovery, namely a reduction of hyperactivity (Homsi et al., 2010; Siopi et al., 2011).
In the present study, the same acute treatment with minocycline was able to significantly attenuate the trauma-induced OB lesions and improve olfactory function, suggesting that inhibition of the early neuroinflammatory response contributed to some extent to the sparing of OB tissue and consequent olfactory recovery. Besides, experimental studies have demonstrated the efficacy of steroid treatments on OB tissue integrity following olfactory nerve and OB injury (Kobayashi et al., 2009). Moreover, there is clinical evidence showing that oral steroid treatment improved olfactory acuity in some patients with post-traumatic anosmia, and that early administration of the treatment was associated with improved outcome (Fuji et al., 2002; Jiang et al., 2010). Hence, it can be suggested that the anti-inflammatory efficacy of steroids may have a therapeutic role in the treatment of olfactory impairment. Nonetheless, their partial efficacy, as well as their non-negligible side effects when used among clinical populations, renders their use problematic, and highlights the need for therapeutic alternatives (Holbrook and Leopold, 2006). Minocycline's anti-inflammatory action in TBI and various neurodegenerative diseases has been well documented, with microglial inhibition being its most common pathway of neuroprotection (Bye et al., 2007; Familian et al., 2006; Kim and Suh, 2009). However, since minocycline's potent anti-inflammatory activity operates through multiple molecular mechanisms, it is difficult to ascertain herein the precise mechanisms responsible for the protective effect observed on OB tissue.
Taken together, the findings in this article demonstrate a pertinent behavioral test for the evaluation of post-TBI olfactory function in mice, and shows the neuroprotective efficacy of an early post-TBI intervention with minocycline on both OB integrity and olfactory outcome. These findings suggest that the anti-inflammatory efficacy of minocycline may have a therapeutic function in the treatment of post-traumatic olfactory impairment; nevertheless more studies are required to reveal the precise underlying mechanisms of the significant neuroprotective effect observed.
Footnotes
Acknowledgments
This work was supported by the nonprofit organization Fondation des Gueules Cassées (grants to E.S. and M.J.T.).
Author Disclosure Statement
No competing financial interests exist.