
Abstract
Traumatic brain injury (TBI) is a leading cause of mortality and morbidity worldwide. Currently, there is no effective strategy to treat the functional sequelae associated with TBI. Experimental evidence shows that the intravenous administration of bone marrow stromal cells (BMSC) during the first week after TBI prevents neurological deficits, but no experimental studies have shown evidence of the effect of intravenous BMSC on chronic brain injury sequelae. Here we studied the effect of intravenous administration of BMSC on functional outcomes 2 months after experimental TBI in rats. Adult Wistar rats were subjected to weight-drop impact causing severe brain injury, and 2 months later BMSC in saline, or saline alone, was intravenously injected. All experimental groups were evaluated by means of the modified Neurological Severity Score (mNSS), and internal zone Permanence Time (izPT) after video-tracking box (VTB) analysis, over the following 2 months to test the efficacy of BMSC therapy. At the end of the study period the animals were sacrificed and their brains were studied to evaluate possible differences between groups. Two months after BMSC administration no significant differences were detected in the motor and sensory evaluation between animals treated with BMSC and controls, and no differences were detected after histological study of the brains. Our present results suggest that intravenous administration of BMSC after TBI, when the neurological deficits are well established, has no beneficial effect in neurological outcomes or on brain tissue.
Introduction
T
At present, stem cells represent a therapeutic option to treat traumatic lesions of the central nervous system (CNS). This cell therapy may help restore neurological function. Adult bone marrow is a source of stem cells, and among these are bone marrow stromal cells (BMSC) (Friedenstein et al., 1976). These cells can differentiate not only into cells of the embryonic layer, such as chondroblasts, osteoblasts, or myoblasts (Prockop, 1997), but under certain conditions they can differentiate into endothelial or neuroectodermic cells (Benayahu et al., 2007; Dewaza et al., 2004; Kotobuki et al., 2004; Parr et al., 2007; Sánchez-Ramos et al., 2000; Woodbury et al., 2000; Zurita et al., 2007,2008a). BMSC are easily to obtain and culture, and they can be used in autologous treatments. Thus BMSC may represent a suitable therapeutic alternative to treat TBI. BMSC have been reported to play a significant role in experimental models of TBI, promoting functional recovery (Li and Chopp, 2009), and they can differentiate into neural cells (Bonilla et al., 2009; Li and Chopp, 2009; Qu et al., 2008; Zurita et al., 2008b).
A number of studies have been conducted to determine the best time and means to administer the cells, mainly during the first week after TBI. Intravenous administration has been most commonly used during the acute stage (Chen et al., 2001; Lu et al., 2001; Mahmood et al., 2001,2003,2004; Qu et al., 2008), and improvements in functional outcome have been reported at least 3 months after treatment. In these studies, BMSC were distributed throughout the body, and some were localized in the lesion zone, where a small percentage express neuronal and glial markers (Lu et al., 2006; Mahmood et al., 2004).
The aim of the present study is to determine if the efficacy of intravenous administration of BMSC during the acute or subacute stage of TBI can also be obtained in the chronic phase, after established neurological sequelae.
Methods
Animal model
All animal experiments described in this study were approved by the experimentation committee of Puerta de Hierro-Majadahonda Hospital. For TBI, 20 female adult Wistar rats were premedicated with intravenous phenytoin 24 h before surgery to prevent epileptic seizures. To induce the brain injury, the rats were placed in an induction chamber, where 8% sevofluorane was delivered in a continuous oxygen flow at 3 L/min. Once the animals were anesthetized they were connected to a small face mask adapted to a stereotactic frame, where fresh gas flow was adjusted to 2.5–3%, and then they were medicated with subcutaneous morphine (2.5 mg/kg) and meloxicam (2 mg/kg). During surgery, under aseptic conditions a 1-cm incision was made along the mid-line of the scalp; the animals were subjected to a 10-mm-diameter craniotomy on the right hemisphere, adjacent to the central suture, midway between the lambda and the bregma. The dura was cut with microsurgery scissors to expose the brain.
Traumatic injury was induced using the technique described by Allen (Allen, 1911), and currently used by us to induce spinal cord injury (Zurita and Vaquero, 2006) and TBI (Bonilla et al., 2009). The brain injury was induced with a 12 mm2-thick cylindrical steel bar weighing 25 g, which was allowed to fall vertically onto the brain surface from a height of 20 cm, guided by a hollow cylinder adapted to the area of the craniotomy. After surgery, the incision was closed with 5-0 absorbable surgical suture. The rats were placed overnight in a temperature- and humidity-controlled chamber.
Neurological evaluation
All rats were tested on the modified Neurological Severity Score (mNSS) and video-tracking box (VTB) tests daily, 3 days before TBI, and weekly during the 4 months of follow-up (2 months after TBI, and 2 months after intravenous BMSC or saline administration).
mNSS testing was performed as previously described, with some modifications (Bonilla et al., 2009). The test is suitable for evaluating long-term neurological function after unilateral TBI, and it is a composite of motor (muscle status and abnormal movement), sensory (visual, tactile, and propioceptive), and reflex tests. Neurological function was graded on a scale of 0–19 (normal score 0; maximal deficit score 19). In the severity scores of injury, one point is awarded for the exhibition of a specific abnormal behavior or for a lack of reflex; thus the higher the score, the more severe the injury. mNSS data are presented as the maximal deficit compared with the internal baseline control.
For a more objective evaluation we used a computer program, Smart, linked to a VTB test, to study the possible neurological deficits and outcome in the animals. This new method adequately quantifies parameters related to locomotor activity and orientation in brain-injured rats. This method has been used in our laboratory to measure behavioral outcome after brain injury caused by intracerebral hemorrhage (ICH) in adult Wistar rats (Otero et al., 2010). VTB tests analyze the images captured by a video camera in a 35×45-cm closed box containing the animal. To study the animal's behavior, we determined different parameters: we captured images for 90 sec, and the maximum speed was established at about 10 cm/sec. We defined different areas to determine differences in the movement and orientation of the animals in the box: an internal zone and an external zone. Rats without TBI preferred the external zone, while animals with brain injury used both zones. We studied the time that the animals spent in the internal zone (internal zone Permanence Time, izPT). Higher scores on the mNSS test, or more time spent in the internal zone of the VTB test, indicated greater neurological deficits.
BMSC isolation
BMSC were obtained from adult male Wistar rats weighing 200–250 g, and they were expanded in vitro for 4 weeks. Using a 1-mL syringe and a 21-gauge needle, fresh complete bone marrow was harvested aseptically from tibias and femurs. Both ends of the bones were opened and the marrow was extruded with 5 mL of alpha-MEM medium (Lonza Group Ltd., Basel, Switzerland). Bone marrow was mechanically dissociated to obtain a homogeneous cell suspension. The cell suspension was filtered through a 70-μm mesh nylon strainer and placed in a 75-cm2 flask for tissue culture with 12 mL alpha-MEM medium containing 20% fetal bovine serum (FBS; Lonza Group Ltd.), 2 mM L-glutamine, 100 U/mL penicillin, 100 mg/mL streptomycin, and 25 ng/mL amphotericin B. The cells were incubated at 37°C in 5% CO2 for 3 days. At this time, non-adherent cells were removed by replacing the medium. The culture medium was replaced three times a week. After the primary cultures reached confluence, they were rinsed three times with phosphate-buffered saline (PBS), and dissociated with 0.25% trypsin solution and 1 mM EDTA for 5 min at 37°C. The cells were washed two times with alpha-MEM medium/2.5% FBS at 1000 rpm for 5 min, and the BMSC obtained were subcultured at 37°C in 5% CO2.
Preparation of transplants and characterization of donor BMSC
To obtain the BMSC for the transplants, cells corresponding to a P1 were rinsed three times with PBS and dissociated with 0.25% trypsin solution and 1 mM EDTA for 5 min, at 37°C. The obtained BMSC, which showed a viability of more than 96%, were injected into the traumatic brain cavity. For flow cytometry analysis, cells were detached with trypsin and labeled with fluorescein isothiocyanate (FITC), phycoerythrin (PE), or Alexa-Fluor 647-conjugated primary antibodies (Ab). The following cell-surface markers were characterized at each passage: CD11b, CD29, CD31, CD45, and CD90, with a suitable fluorocrome-conjugated isotype control (all from AbD Serotec, Oxford, U.K.). Each Ab is specific to rat markers. Standard labeling protocols and the manufacturers suggestions were followed. Cells were suspended at 106 cells/mL in running buffer (RB; Miltenyi Biotec GmbH, Gladbach, Germany), and incubated with primary conjugated Abs (1:10 v/v) for 15 min at 4°C. Then the cells were washed in RB, centrifuged for 7 min at 4°C, and resuspended in 0.5 mL RB. Then 5 μL of propidium iodide (Southern Biotechnology, Birmingham, AL) was added in all conditions to remove dead cells, and they were analyzed by flow cytometry using a Cytomics FC500 MPL cytometer and CFC 500MXP software, version 2.2 (Beckman Coulter Inc., Miami, FL). For cell treatment, only cells from cultures less than 1 month old that expressed mesenchymal phenotype criteria were used: expression of CD29 and CD90 (≥95% positive), and a lack of expression of CD11b, CD45, or CD31 (<5% positive). The collected data were further analyzed and graphs created using CXP software (version 2.1; Beckman Coulter Inc.).
Intravenous transplantation and experimental groups
Two months after TBI all rats were anesthetized and medicated as described above. The injured rats were randomly divided into two experimental groups. Rats in the BMSC-transplanted group (n=10) received 15×106 BMSC in 1 mL of saline injected intravenously into the tail vein 2 months after TBI. Rats in the control group (n=10) received 1 mL of saline alone.
Histological studies
Two months after BMSC or saline administration, the rats were sacrificed for morphological studies. For sacrifice, the animals were anesthetized with 8% sevoflurane in a continuous oxygen flow of 3 L/min, and perfused transcardially with 20 mL heparinized saline, followed by 120 mL of 4% paraformaldehyde in 0.1 M PBS (pH 7.4). The brains were removed and stored in 4% paraformaldehyde in 0.1 M PBS (pH 7.4) for 24 h. The brains were then dissected by separating the hemispheres, and tissue blocks containing each hemisphere were processed for paraffin sectioning. A series of 5-mm-thick sections were cut with a microtome through each block, and one section out of 10 from each block was mounted on glass slides for histological examination after hematoxylin and eosin (H&E) staining. In each case, the volume of the lesion was calculated on histological slices according to the equation: V(mm3)=d2(mm2) – D(mm)/2, where d and D are the smallest and the largest lesion diameters, respectively.
Immunohistochemical and in situ hybridization studies
Brain sections were initially immunostained with diaminobenzidine (DAB) for detection of neurofilament-200 kDa (NF-200, a neuronal marker), glial fibrillary acidic protein (GFAP, an astroglial marker), Ki-67 (a proliferation marker), nestin (a neural stem cell marker), and double-cortin (Dcx, a migratory neuroblast marker). For identification of neurons and astrocytic and neural stem cells, adjacent slides were hydrated and rinsed in PBS, the sections were exposed to 3% H2O2 for 30 min to quench endogenous peroxidase activity, followed by blocking in normal serum for 2 h. Then they were treated with monoclonal antibody NF-200 (dilution 1:100; AbD Serotec), monoclonal antibody GFAP (dilution 1 mg/mL; Chemicon, Temecula, CA), or mouse anti-nestin monoclonal antibody (1 mg/mL; Chemicon) at 4°C overnight. Subsequently, the sections were incubated with biotin-conjugated anti-mouse IgG (dilution 5 μg/mL; Vector Laboratories, Burlingame, CA). Additionally, for Dcx-positive cells the sections were incubated with goat anti-double-cortin polyclonal antibody (1:200; Santa Cruz Biotechnology), and with secondary antibody biotin-conjugated anti-goat IgG (1:200; Vector Laboratories). For proliferating cell detection, adjacent slides were pretreated in boiled citrate buffer (pH 6) in a microwave (650–720 W). A primary rabbit anti-Ki-67 protein monoclonal antibody (1:200; LabVision Westinghouse, Fremont, CA), and a secondary antibody biotin-conjugated anti-rabbit IgG (1:500; Jackson ImmunoResearch Laboratories, Inc., Baltimore Pike, MD) were used.
In all cases the sections were washed in PBS, and incubated with avidin-biotin-horseradish peroxidase complex (Vector Laboratories). DAB was used as a chromogen. Control slices lacking primary or secondary antibodies were analyzed with each series. For possible identification of male donor cells, in situ hybridization studies were performed. Tissue sections (5 mm) were dewaxed and rehydrated with xylene and graded ethanol, and subsequently digested with pronase K (50 mg/mL) for 10 min at 37°C. We used a dioxygenin-DNA probe for sequence specificity for the murine Sry gene, the sex-determining region of the Y chromosome. Hybridization was performed in a hybridization mixture consisting of 50% deionized formamide, 200 mg/mL salmon test DNA, 10% dextran sulfate, 10% 50× Denhardt's solution, 10% 20× standard saline citrate, and 400 ng biotinylated-labeled probe, at 50°C overnight. The dioxygenin-labeled probe was visualized by immunofluorescence using a primary anti-dioxygenin mouse monoclonal IgG antibody (1:100; Jackson ImmunoResearch Laboratories), and a secondary antibody, rhodamine (TRICT)-conjugated anti-mouse IgG (1:200; Jackson ImmunoResearch Laboratories). The sections were stained with 4,6-diamino-2-phenylindole (DAPI) and mounted with glycerol. Negative control sections from each animal received identical staining preparation except that the probe was omitted, and in positive controls the hybridizations were performed in male rat brains. Some sections were subjected to double labeling to visualize the possible co-localization of SrY together with the neuronal marker Neu-N or the astroglial marker GFAP. Each coronal brain section was first treated for hybridization as previously described, and thereafter the primary antibodies mouse anti-Neu-N (1:500; Chemicon) or mouse anti-GFAP (1 mg/mL; Chemicon) were used to detect Neu-N or GFAP, respectively. Cy™-2-conjugated anti-mouse IgG (1:200; Jackson ImmunoResearch Laboratories) was used as a secondary antibody. The sections were stained with DAPI and mounted with glycerol. Negative and positive controls were performed.
Statistical analysis of cell therapy effectiveness
Statistical analysis of the data from control and grafted animals was performed with SPSS software, using the parametric analysis of variance (ANOVA) test. In these studies, p<0.05 was considered significant.
Results
Neurological function evolution
Results of both neurological tests were obtained 2 months after treatment (4 months after TBI). When the mNSS test was studied (Fig. 1), the evolution of rats with intravenous administration of BMSC was similar to the rats in the saline group, with no significant differences seen (p≥0.05). The same results were obtained on the izPT test (VTB image analysis, Fig. 2), and the evolution of BMSC-treated animals was similar to that of saline-treated animals (p≥0.05); no significant differences were seen.
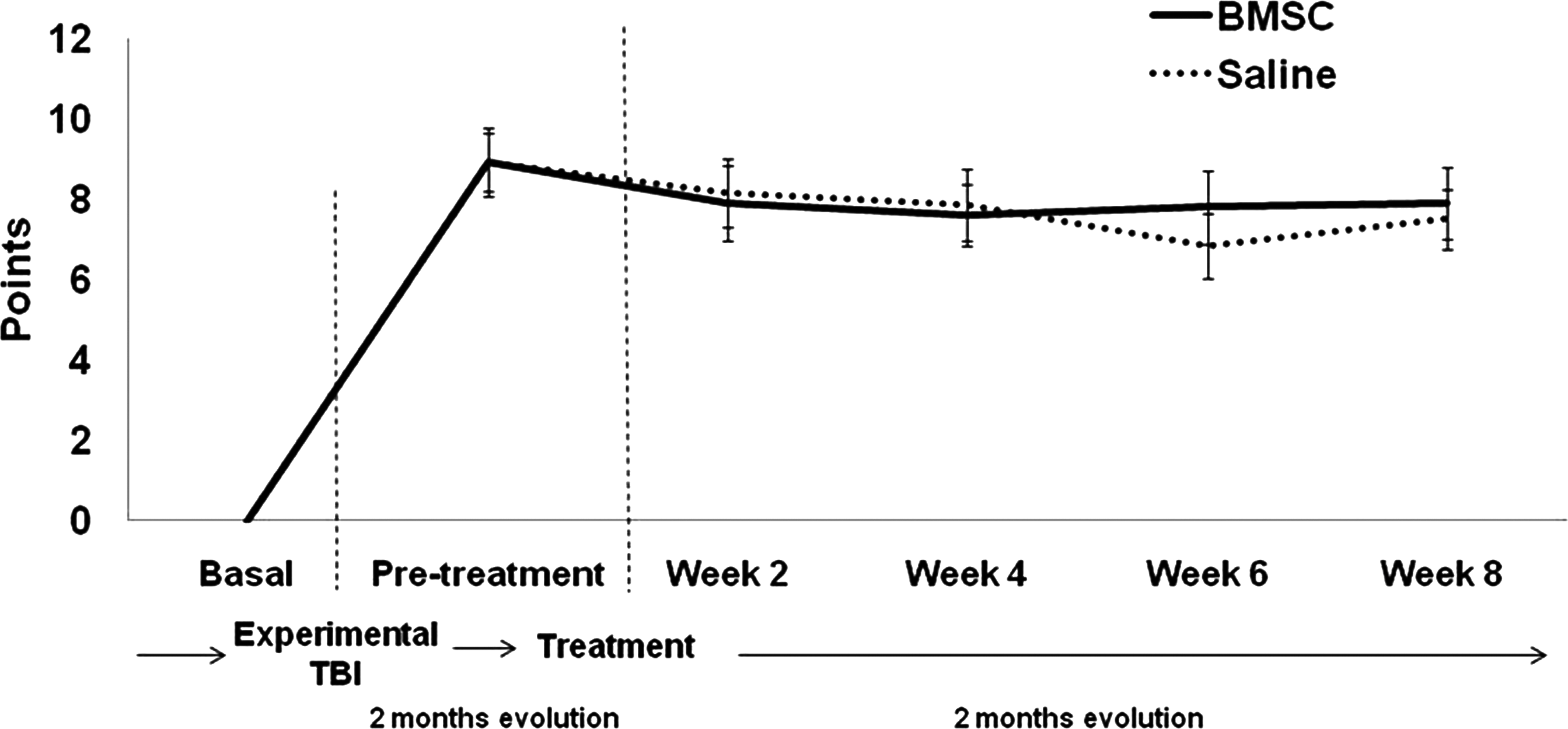
Line graph showing the temporal profile of functional recovery on the mNSS test after the two treatments (saline or BMSC; mNSS, modified Neurological Severity Score; BMSC, bone marrow stromal cell).
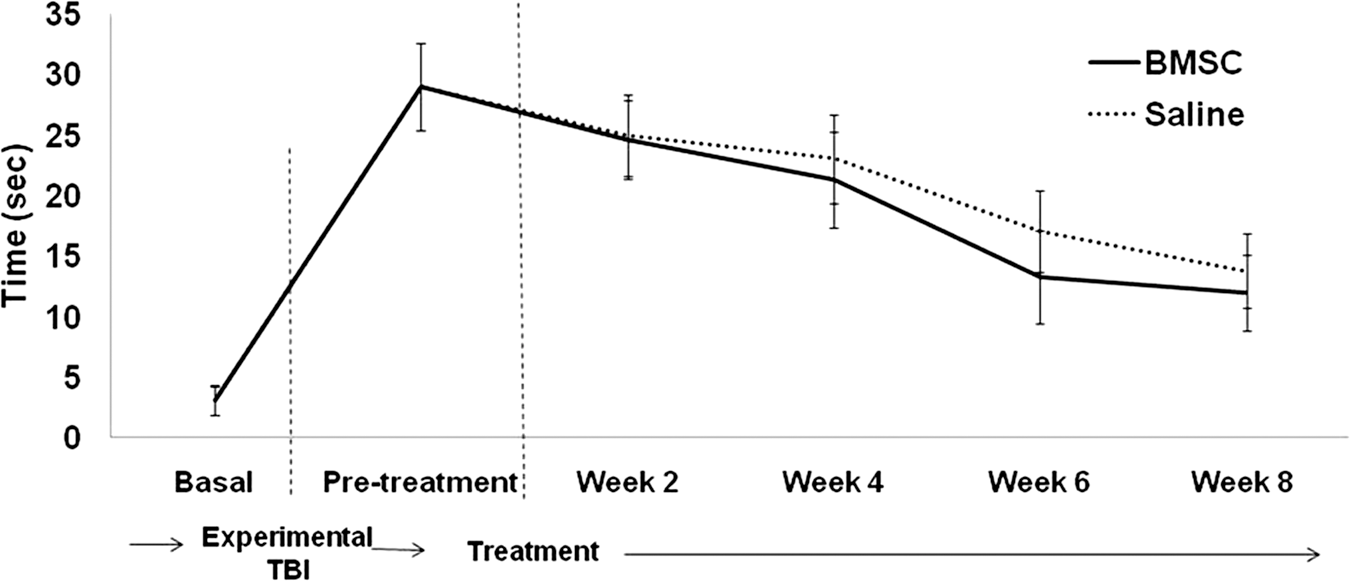
Line graph showing the temporal profile of functional recovery on the VTB test after the two treatments (saline or BMSC; BMSC, bone marrow stromal cell; VTB, video-tracking box).
Morphological study of the lesion zone
At the end of the study, all animals were sacrificed to allow macroscopic study of the lesion zone. To determine if the cell therapy had any influence on tissue morphology, the lesioned brain area and the tissue changes were analyzed with H&E staining. On the qualitative histological analysis, extensive tissue damage with loss of normal cerebral cortical architecture was observed in the rats in both experimental groups. Morphological studies identified the lesion zone in all animals as a central cavity surrounded by a scar tissue wall partially filled with macrophages.
The size of the lesion was variable, but it was not significantly different when the two experimental groups were compared (Fig. 3). In animals with BMSC treatment (50.75±15.66 mm3) a slightly smaller lesion volume was detected than that of the animals in the saline control group (63.45±13.18 mm3) 2 months after treatment, and 4 months after TBI, but the reduction was not statistically significant (p≥0.05).

Bar graph showing the lesion volume calculated from hematoxylin and eosin-stained histological slices in BMSC- and saline-treated animals (BMSC, bone marrow stromal cell).
Histological studies
When the neuronal and astrocytic markers were studied, no significant differences in the tissue architecture were detected between rats that received BMSC and those receiving saline alone. For the proliferative study we used Ki-67 protein, which is present during all active phases of the cell cycle and is absent from resting cells. When the proliferative activity of the cells was compared in the two experimental groups (Fig. 4), an increase in proliferative cells in the BMSC-transplanted rats could be observed, but no significant differences were detected. In the BMSC-transplanted group, the mean (±SD) of cells expressing Ki-67 was 102.10±58.26, while in the control group the mean number of Ki-67-positive cells was 59.56±30.95.
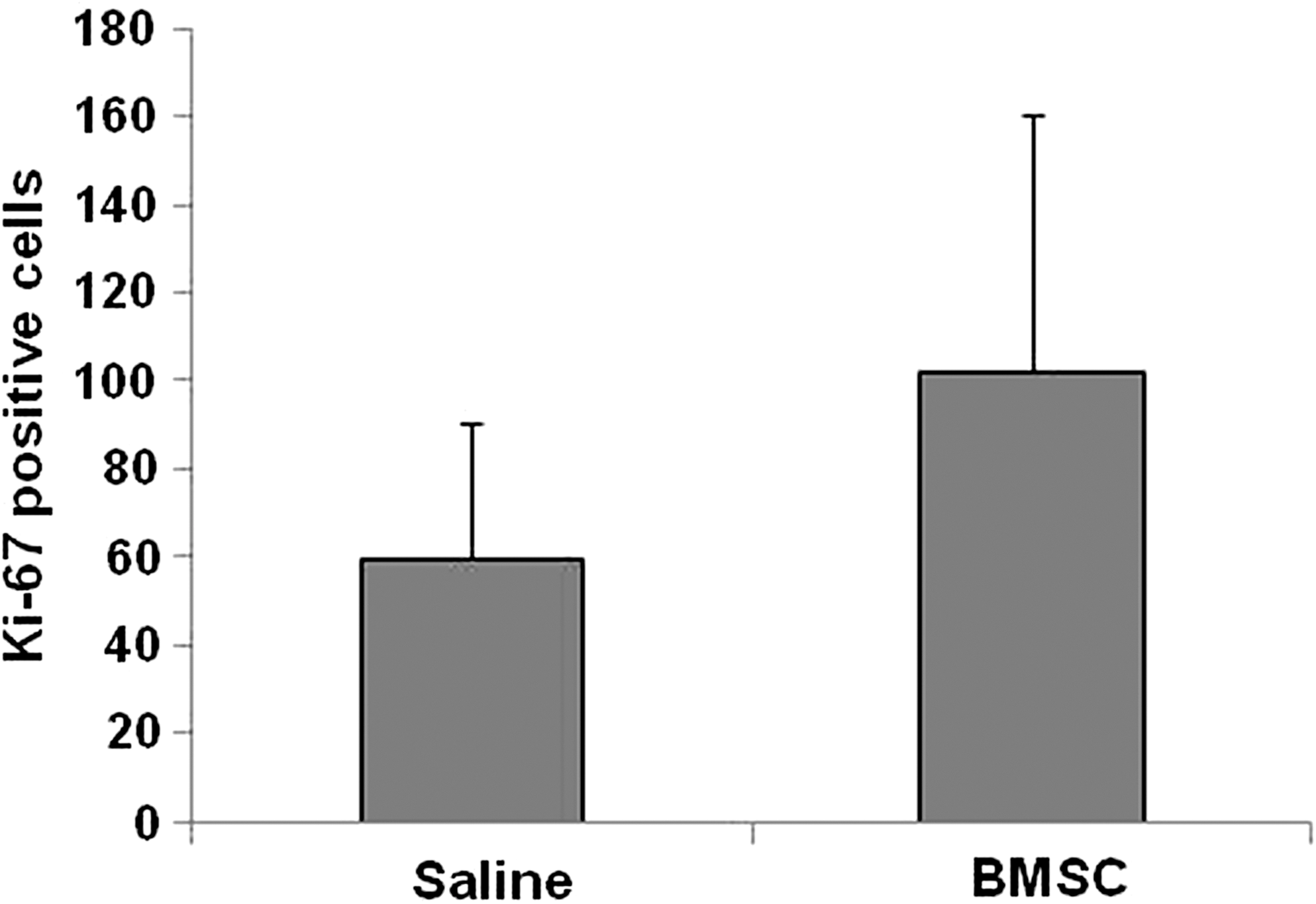
Bar graph showing the number of Ki-67-positive cells after BMSC and saline treatment (BMSC, bone marrow stromal cell).
For neurogenesis detection we studied the nestin and double-cortin expression in samples of animals from both experimental groups. The presence of variable numbers of immunostained cells showing nestin and double-cortin expression was a constant finding in the subventricular zone of the animals belonging to both groups, and occasionally nestin- and double-cortin-positive cells were identified in clusters of undifferentiated cells near the lesion zone. On the other hand, in the animals from the control group, the presence of cells expressing nestin and double-cortin was lower than in the BMSC-transplanted group, but these differences were not statistically significant. No transplanted cells (CR-Y-positive) were detected in the brains of the animals treated with BMSC. Consequently, no cells with co-expression of SrY and Neu-N or GFAP markers were detected.
Discussion
In recent years, the development of cell therapies using adult stem cells has opened a new avenue for treating CNS lesions. Although there is evidence suggesting that cell therapy improves neurological deficits after TBI (Bonilla et al., 2009), at present there is no agreement about which is the optimal time for cell therapy or what is the best route of administration.
The efficacy of intracerebral administration in the acute and chronic stages of brain damage has been reported (Bonilla et al., 2009; Mahmood et al., 2002), but this may require complex surgery. Intravenous cell infusions have many advantages over other routes, as they do not require complex processes to permit administration of large numbers of cells (Qu et al., 2008). Intravenous administration has obvious clinical advantages, and in the literature there are many studies that support the efficacy of this route of delivering BMSC in the first week after TBI (Jackson et al, 2010; Lu et al., 2001; Mahmood et al., 2001,2005). In the chronic stages of brain damage only one experimental study aimed to investigate the optimal time window for IV BMSC therapy, and cell transplantation was done 1 month after stroke (Shen et al., 2007). These researchers found that delayed IV administration of BMSC induced significantly improved functional outcomes. They postulated that the beneficial effects evoked by BMSC treatment may be related to the induction of neurogenesis and reduction of glial scar formation.
Although IV BMSC administration is a promising strategy to improve recovery after TBI, the optimal time to transplant the cells remains unknown. In an effort to better understand the efficacy of different routes of administration, and to study the best and safest means of cell administration to treat TBI, we investigated the efficacy of intravenous BMSC administration at the chronic stage post-TBI. Our data revealed that a single dose of 15×106 BMSC given intravenously 2 months after an experimental TBI did not result in substantial improvement of neurological function or differences in the tissue characteristics when the brains of the animals treated with BMSC and saline alone were compared.
In the present study, functional deficits were evident in all animals after TBI, and decreased during the next 2 months after surgery, at a time when functional deficits were constant and the chronic stage of the brain damage was reached. At this stage, motor and sensory tests are inadequate to quantify cell efficacy after treatment. Thus, we also used VTB testing to complement the mNSS results, because this test is more objective in measuring the functional evolution post-injury. With the mNSS test, when the animals reached the chronic stage post-injury, the test results over the next 2 months were similar, regardless of the treatment received. On the VTB test, the animals decreased the time spent in the study zone, but the evolution of all animals was similar in both experimental groups. There was no motor or cognitive functional recovery after IV administration of BMSC 2 months after TBI. This finding is consistent with recently published data showing that a single dose of BMSC delivered 24 h after a moderate TBI did not result in substantial cell engraftment or improvement in functional recovery, possibly because the majority of cells were localized to pulmonary tissue, with only a small subset (0.0005%) of the total number of cells reaching the brain (Harting et al., 2009). Other findings that support our results were seen in a model of spinal cord injury in the rat, showing the utility of intralesional BMSC administration to obtain clear and progressive functional recovery of established paraplegia, and suggesting that although some functional recovery can be obtained after established paraplegia via systemic administration of BMSC, this recovery is much less than that seen when intralesional administration of BMSC is performed (Vaquero et al., 2006; Zurita and Vaquero, 2004).
Our data clearly show that no beneficial response is associated with IV BMSC therapy at 2 months post-transplantation. The findings obtained with IV administration during the first week post-injury showed that the functional improvement seen with BMSC treatment is not only due to the BMSC replacing neural cells, but is also due to the ability of BMSC to induce growth factor production and promote intrinsic neurorestorative functioning of the brain (Mahmood et al., 2004,2006). During the chronic stage of brain injury, BMSC increase proliferation and endogenous neurogenesis when the treatment is given intracerebrally (Bonilla et al., 2009), but in our present study, no increase in proliferation (no difference in Ki-67-positive cells was seen between the two study groups), or endogenous neurogenesis (no differences in double-cortin and nestin expression were seen between animals in the two groups) was observed with the IV administration during the same period post-TBI.
In previous studies, following intravenous cell transplantation, cells migrated into the brain and gave rise to cells expressing neuronal and astrocyte markers. These cells were found at the site of injury and in some areas associated with blood vessels (Lu et al., 2006). In these works, acute BMSC therapy after TBI was mediated by the integration of the intravenously transplanted cells and differentiation into neural cells. With chronic IV BMSC administration no intracerebral transplanted cells were found, although the number of cells administered was the largest described in such therapy to treat brain damage. Moreover, no neuronal or glial transplanted cells were detected in the lesioned brain, which could be related to a lack of any increase in proliferative and endogenous neurogenesis after cell treatment, and no improvement in neurological function 2 months after transplantation.
Our finding in the present study was that a single intravenous dose of BMSC 2 months after an experimental TBI did not lead to cell integration into damaged brain tissue or improvement in functional recovery. Comparisons of cell delivery routes and assessment of the optimal time for cell therapy administration is necessary as a first step before considering the application of these new techniques in humans.
Footnotes
Acknowledgments
This study was made possible by grants received from the Mapfre Foundation, the MM Foundation, the Rafael del Pino Chair for Neuroscience Research, and the Institute of Health Carlos III (FIS PI060650).
Author Disclosure Statement
No competing financial interests exist.