
Abstract
There is growing evidence that repeated brain concussion can result in cumulative and long-term behavioral symptoms, neuropathological changes, and neurodegeneration. Little is known about the factors and mechanisms that contribute to these effects. The current study addresses the need to investigate and better understand the effects of repeated concussion through the development of an animal model. Male Long-Evans rats received 1, 3, or 5 mild lateral fluid percussion injuries or sham injuries spaced 5 days apart. After the final injury, rats received either a short (24 h) or long (8 weeks) post-injury recovery period, followed by a detailed behavioral analysis consisting of tests for rodent anxiety-like behavior, cognition, social behavior, sensorimotor function, and depression-like behavior. Brains were examined immunohistochemically to assess neuroinflammation and cortical damage. Rats given 1, 3, or 5 mild percussion injuries displayed significant short-term cognitive impairments. Rats given repeated mild percussion injuries displayed significantly worse short- and long-term cognitive impairments. Rats given 5 mild percussion injuries also displayed increased anxiety- and depression-like behaviors. Neuropathological analysis revealed short-term neuroinflammation in 3-injury rats, and both short- and long-term neuroinflammation in 5-injury rats. There was also evidence that repeated injuries induced short- and long-term cortical damage. These cumulative and long-term changes are consistent with findings in human patients suffering repeated brain concussion, provide support for the use of repeated mild lateral fluid percussion injuries to study repeated concussion in the rat, and suggest that neuroinflammation may be important for understanding the cumulative and chronic effects of repeated concussion.
Introduction
B
Work from our laboratory and others has found that a single mild lateral fluid percussion injury (mLFP) in the rat can model behavioral and pathological effects of a single concussion (DeRoss, 2002; Gurkoff et al., 2006; Shultz et al., 2011; Wu et al., 2010). These findings suggest that repeated mLFP might hold promise as a means to study the effects of multiple concussion and CTE in an animal model. The present study was undertaken to provide useful information about the behavioral impairments and associated pathologies that might result from the cumulative effects of repeated concussion. Here rats received either 1, 3, or 5 mLFPs, or sham treatment, at 5-day intervals. Previous work has shown that the cerebral pathophysiology at 5 days after lateral fluid percussion injury in the rat is equivalent to approximately 2–4 weeks post-concussion in humans (Giza and Hovda, 2001). This sequence of injuries is similar to what might occur in athletes and military personnel under common concussion management guidelines (Maroon et al., 2000). Consistent with emerging human evidence, here we predicted that repeated mLFPs might result in cumulative effects that would manifest as greater enduring behavioral impairments and brain pathology than a single mLFP.
Methods
Subjects
Subjects were 122 young adult male Long-Evans hooded rats (Charles River Labs, St. Constant, Quebec). Prior to surgery rats weighed between 250 and 300 g, were housed in pairs in acrylic cages (26 cm×48 cm×21 cm) at 21±1.0°C, and were naïve to all experimental procedures. After surgery rats were housed individually under a 12-h:12-h light/dark cycle, with lights on from 7:00 to 19:00 hours and ad libitum access to food and water. All procedures complied with guidelines of the Canadian Council on Animal Care and were approved by the University of Western Ontario.
Surgery: Craniotomy and injury cap
Anesthesia was induced in a sealed acrylic glass box using 4% isoflurane±2 L/min oxygen flow. Anesthesia was maintained throughout surgery using a nose cone with a standard stereotaxic device and 2% isoflurane +500 mL/min oxygen flow. Under aseptic conditions rats underwent craniotomy surgery to create a circular window (3 mm diameter) centered at A/P −3.0 mm, M/L 6.0 mm with reference to the bregma (Paxinos and Watson, 1986). A hollow plastic injury cap was sealed over the craniotomy with silicone adhesive, cyanoacrylate, and dental acrylic anchored to stainless steel screws inserted into the skull. Topical antibiotic ointment was applied and a removable plug was inserted into the injury cap to seal the craniotomy. All rats received a subcutaneous injection of analgesic (ketoprofen, 5 mg/kg).
Injury groups and schedule
Rats were randomly assigned to one of four treatment groups: one concussion (1C), three concussion (3C), five concussion (5C), or sham-control (sham; Table 1). Repeated mLFP methods were adapted from work done in our lab and others (DeRoss et al., 2002; Shultz et al., 2011; Thompson et al., 2005). On day 1, 24 h post-surgery, rats were placed in a sealed acrylic glass box into which 4% isoflurane±2 L/min oxygen flow was introduced for anesthesia. The plug was removed from the cap and under aseptic conditions the cap was filled with sterile saline and connected to the fluid percussion device (FP302; AmScien Instruments, Richmond, VA). At the first response of hindlimb withdrawal to a toe pinch, sham, 1C, and 3C rats were removed from the device, which constituted sham treatment, and 5C rats received mLFP. A fluid percussion force of 1.0–1.5 atm was delivered by the fluid percussion device to produce each mLFP. This percussion force was chosen based on previous rodent studies that have validated the use of mLFP in a rat model of concussion (DeRoss et al., 2002; Shultz et al., 2011; Wu et al., 2010). No percussion force was delivered during sham treatments. Acute injury measures consisting of duration of apnea, duration of unconsciousness, and latency to occurrence of the self-righting reflex were all monitored beginning immediately after each sham or mLFP (Griesbach et al., 2009; Gurkoff et al., 2006; Shultz et al., 2011). Apnea duration was the time from injury to the return of spontaneous breathing. Duration of unconsciousness was the time from injury to the return of the hindlimb withdrawal in response to toe pinch. Time to the self-righting reflex was the time from injury to return to an upright position. Rats were again anesthetized and received either a sham or mLFP on days 6, 11, 16, and 21, according to the schedule in Table 1.
Each repeated treatment was separated by a 5-day period. S, sham injury; mLFP, mild lateral fluid percussion injury.
After the last treatment rats were randomly assigned to receive either a short recovery period (SR) of 24 h, or a long recovery period (LR) of 8 weeks before the start of behavioral testing. A total of 7 rats died and 5 rats were removed from the study prior to the onset of behavioral testing because of rupture of the dura, loss of the injury cap, or failure to maintain normal body weight. There were 8 experimental groups, with final group sizes as follows: 1C-SR (n=14); 3C-SR (n=14); 5C-SR (n=14); sham-SR (n=17); 1C-LR (n=12); 3C-LR, (n=14); 5C-LR (n=13); and sham-LR (n=12).
Behavioral test apparatus
Anxiety-like behavior was assessed using an elevated-plus maze with two arms intersecting at a 90° angle, thereby creating 4 individual arms each 55 cm long and 12 cm wide, placed 50 cm above the floor. Two opposing closed arms were shielded by 50-cm-high walls, and two opposing open arms contained no walls (Saucier et al, 2008; Shultz et al., 2011; Walf and Frye, 2007). An overhead video camera recorded all trials. Video was scored for the number of entries into and amount of time spent on each arm by a researcher blind to group membership.
Spatial cognition was assessed using a water maze consisting of a circular pool (1.5 m in diameter, 45 cm deep) filled with tap water at 29±1.0°C. A clear acrylic glass escape platform (9 cm×9 cm) was hidden 2 cm below the water surface in the center of the southeast pool quadrant during acquisition, and in the center of the northwest quadrant during reversal. Polypropylene beads floating on top of the water prevented rats from seeing the hidden platform, and doors, cabinets, and posters on the walls provided distal visual cues (Cain et al., 1993; Shultz et al., 2009,2011). A video camera mounted above the center of the pool fed a computer for the recording and objective analysis of digitized swim trials (Poly-Track; San Diego Instruments, San Diego, CA).
Locomotor and social behaviors were evaluated in a circular open arena (90 cm diameter, 40 cm walls) with Beta Chip bedding covering the floor (Shultz et al., 2008;2011). A CD camera mounted above the center of the arena fed a computer for the recording and objective analysis of digitized behavioral data (EthoVision 3.0.15 Behavioral Monitoring/Analysis System).
Sensorimotor ability was tested using an elevated narrow wooden beam 1 m long and 2 cm wide (Shultz et al., 2011,2009). A video camera recorded all trials, which were scored by a researcher blind to group membership to determine traverse times and the number of slips and falls onto soft padding on the floor. Experience with the water maze does not affect performance on the beam task (Beiko et al., 1997).
Depression-like behaviors were assessed using the forced swim test, in which a rat was placed for 5 min in a clear glass cylinder (20 cm diameter) filled with 25±1.0°C water to a depth of 30 cm (Jones et al., 2008; Porsolt et al., 1977; Shultz et al., 2011). Behavior was recorded using a side-view video camera and video was later scored by a researcher blind to group membership.
Experimental procedure: Day 1
Behavioral testing began either 24 h (SR) or 8 weeks (LR) after the last injury treatment. Rats were placed in the center of the elevated-plus maze facing an open arm and allowed to explore the maze freely for 5 min (Walf and Frye, 2007). Shortly after elevated-plus maze testing, 10 trials of water maze acquisition training were given. Rats were run in squads of 5, and were given approximately 5–6 min between each of the 10 trials. A trial began when a rat was placed in the pool adjacent to and facing the pool wall, and ended when the rat stood on the hidden platform or 60 sec elapsed, at which point the rat was placed on the platform by the experimenter (Shultz et al., 2011). Trials began at one of four start locations (north, south, east, or west), and were ordered pseudo-randomly to prevent sequential starts from the same location. After 15 sec on the platform rats were placed in a drying chamber heated by an infrared lamp.
Experimental procedure: Day 2
Approximately 24 h after the completion of water maze acquisition training, individual rats were placed in the center of the open field apparatus and explored the arena for 10 min (Shultz et al., 2011). Testing in the open field was followed by 10 trials of water maze reversal training. Water maze reversal procedures were identical to acquisition except that the hidden platform was now located in the opposite quadrant of the pool (Shultz et al., 2011,2009).
Experimental procedure: Day 3
Two rats that had received the same number of injuries or control treatments were placed in the open field and social behavior data were collected for 30 min. On the day before testing, the dorsal surface of one rat from each pair was colored black using hair dye, allowing EthoVision to distinguish and track each rat separately (Shultz et al, 2011,2008).
To acclimate rats to the beam task, rats received 5 training trials with a 4-cm-wide beam, followed by 5 trials with a 2-cm-wide beam (Shultz et al., 2011,2009). Rats then underwent forced swim training consisting of 15 min of swimming in the forced swim apparatus (Jones et al., 2008; Porsolt et al., 1977; Shultz et al., 2011).
Experimental procedure: Day 4
Beam testing took place in a darkened room and consisted of 10 trials with the 2-cm-wide beam (Shultz et al., 2011,2009). A trial began when a rat was placed on the illuminated start end of the beam, and ended when the animal reached the darkened goal platform. A maximum of 60 sec was allowed for each trial. Rats were run in squads of 5, with inter-trial intervals of approximately 5 min.
Shortly after beam testing each rat was placed in the forced swim apparatus for a 5-min test session (Jones et al., 2008; Porsolt et al., 1977; Shultz et al., 2011).
Behavioral analyses
For the elevated-plus maze, entries and time spent in the open and closed arms were summed separately. All four of the rats' paws had to enter an arm for it to be considered an entry (Walf and Frye, 2007). As time spent in the open arm is decreased in rats that exhibit anxiety-like behaviors, the following percentage score was calculated for the time spent in the open arm: time in the open arm/[time in the open arm±time in the closed arm] (Saucier et al., 2008; Shultz et al., 2011; Walf and Frye, 2007). Entries into the closed arm were calculated as a measure of locomotion (Hogg, 1996; Walf and Frye, 2007).
Search time and direct and circle swims in the water maze were used as measures of spatial place memory (Morris, 1989; Shultz et al., 2011; Whishaw and Jarrard, 1995). Search time was the time in seconds from release until the rat climbed onto the hidden platform (maximum 60 sec). Direct and circle swims were measured because they represent efficient swim paths that are normally generated by control rats swimming to a fixed visible or hidden platform, they provide data from each trial, and they are not confounded by changes in swim speed (Beiko et al., 2004; Cain and Boon, 2003; Cain et al., 2006; Whishaw and Jarrard, 1995). A direct swim was one that remained entirely within an 18-cm-wide virtual alley from the start point to the hidden platform without crossing over itself. A circle swim was one that approximated an arc of a circle without exceeding 360° or crossing over itself. Direct and circle swims were summed and calculated as a percentage of the total swims for each test session. Swim speed was used as a measure of motor ability and was objectively calculated in cm/sec by Poly-track.
For open field and social behavior analysis, EthoVision automatically calculated the total distance traveled (cm) by each rat (open field and social), and the mean distance apart (cm) between the rats in each tested pair (social; Shultz et al., 2011,2008).
For beam task analysis, traverse time and the number of slips and falls were scored as measures of sensorimotor function (Kolb and Whishaw, 1985; Shultz et al., 2011,2009). Traverse time was defined as the time required to traverse the beam (maximum 60 sec). Slips and falls were scored when one or more paws slipped off the beam, or when a rat fell completely off the beam. Rats that fell off the beam were given a maximum time of 60 sec.
The following measures were scored to assess depression-like behaviors: (1) time spent immobile (primary measure), defined as the rat making only the necessary movements to keep its head above water, (2) time spent escaping, defined as the rat actively struggling to escape the cylinder with its forepaws breaking the surface of the water, and (3) time spent swimming, defined as the rat remaining active in the cylinder but not struggling (Jones et al., 2008; Porsolt et al., 1977; Shultz et al., 2011).
Brain tissue preparation and immunohistochemical procedures
After behavioral testing the animals were deeply anaesthetized with sodium pentobarbital (270 mg/mL IP) and transcardially perfused with ice-cold PBS followed by 4% paraformaldehyde in PBS. The brains were removed, placed in 4% paraformaldehyde solution, and stored at 4±1.0°C for 24 h. Following fixation, the brains were placed in an 18% sucrose solution for cryoprotection (Shultz et al., 2011,2009,2008).
Randomly selected brains from 5 or 6 rats per injury group were used for immunohistochemical analysis. Serial 35-μm coronal cross-sections were obtained through the cortex at the level of injury (approximately −3.0 mm posterior to the bregma). This site was chosen because it allowed reliable quantification of mLFP-induced changes in the rat (Aihara et al., 1995; Shultz et al., 2011), in cortical areas that have been shown to be altered in human concussion (Umile et al., 2002). Monoclonal mouse anti-ED1 (1:500; Serotec, Raleigh, NC) and NeuN (1:500, Chemicon, Temecula, CA) antibodies were used for immunohistochemical staining. Randomly selected, representative sections of the injured area from each animal were processed free-floating for staining as described previously (Weaver et al., 2001). Immunoreactivity was revealed with a glucose-diaminobenzidine-nickel solution. The stained sections were rinsed in PBS, mounted on slides, dehydrated through a gradient of ethanol baths, cleared, and cover-slipped with DPX mounting medium. PBS was substituted for the primary antibody on control sections in each reaction.
Immunohistochemical analyses
For assessment of activated microglia/macrophages an experimenter blinded to the treatment groups captured a light photomicrograph at 1.25×magnification from the ED1-stained ipsilateral coronal cross-section closest to the level of injury in each rat. Photomicrographs were placed with the longitudinal fissure oriented vertically (Fig. 1). Using Image Pro Plus software, one line was drawn from the most dorsal point of the longitudinal fissure to the third ventricle (Fig. 1). A second line was drawn from the third ventricle at a 60° angle to the surface of the injured cortex. It was known that this 60° angle would indicate the approximate epicenter of the injury (Paxinos and Watson, 1986), an area known to display a neuroinflammatory response after a single mLFP (Aihara et al., 1995; Shultz et al., 2011). In the event that the epicenter of the injury had shifted from the standard 60° angle or significant cavitation had occurred, an additional 20° was allotted to either side of the 60° line, and the region of greatest immunoreactivity was located within these boundaries (Fig. 1). A photomicrograph at 20×magnification was then obtained for quantification. All photomicrographs were captured under fixed microscope illumination settings and exposure times to ensure objective and consistent image quality across all pictures. To count the total number of activated microglia/macrophages, the Image Pro Plus color detection function was used to identify ED1-positively-stained cells (Shultz et al., 2011,2008). A standard color threshold was set to detect inflammatory cells while excluding the background (Shultz et al., 2011,2008).
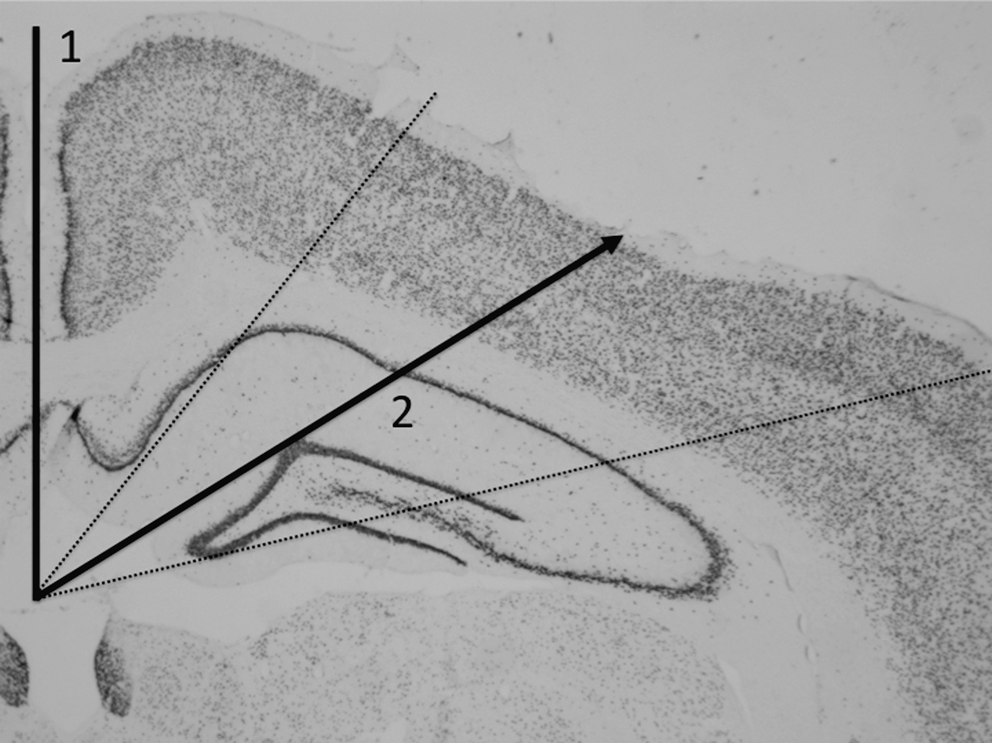
Brain area used for immunohistochemical quantification. Solid line 1 was drawn from the most dorsal point of the longitudinal fissure to the third ventricle. Solid line 2 was drawn from the third ventricle to the surface of the cortex, at a 60° angle to solid line 1, to indicate the approximate epicenter of the injury. The dotted lines indicate an additional 20° allotted on each side of solid line 2 to ensure inclusion of the epicenter of the injury. Images for immunohistochemical quantification were obtained from the cortex between the dotted lines.
An experimenter blinded to the injury groups also completed a semi-qualitative analysis that assessed damage in the injured cortex. A 1.25×magnification photomicrograph of NeuN-stained injured ipsilateral cortex was obtained from each coronal cross-section using a standard light microscope. A score of 1 was given for no or mild (e.g., slight cortical depression) cortical damage. A score of 2 was given for moderate cortical damage (e.g., slight cavitation/cortical loss). A score of 3 was given for severe cortical damage (e.g., obvious cavitation/cortical loss). This rating scale was adapted from previous studies (Braak and Braak, 1991; Jones et al., 2008; Lemstra et al., 2007; Li et al., 2006).
Statistical analyses
Water maze search time and beam traverse times were analyzed by SPSS 18.0 using mixed design analysis of variance (ANOVA), with injury group as the between-subjects factor and trial as the within-subjects factor. Simple effects post-hoc F-tests were carried out when appropriate. One-way ANOVAs, with injury group as the between-subjects factor, were used to analyze acute injury measures, elevated-plus maze measures, direct and circle swims, swim speed, distance traveled, mean distance apart, slips and falls, forced swim measures, and ED1 quantification. Fisher's LSD post-hoc pair-wise comparisons were carried out when appropriate. Statistical significance was set at p<0.05.
Results
Acute injury measures
Statistical analyses were conducted on the acute injury measures taken after each of the group's final assigned mLFP- or sham-treatment. Acute injury measures worsened with repeated injuries, as indicated by significant effects of injury group on apnea (F (3,98)=3.983, p<0.01; 5C>all other groups, all p<0.05; Fig. 2A), unconsciousness (F (3,98)=9.320, p<0.001; 5C > all other groups, 3C > sham, all p<0.05; Fig. 2B), and self-righting reflex times (F (3,98)=24.577, p<0.001; 5C > all other groups, 3C > 1C and sham, all p<0.05; Fig. 2C).
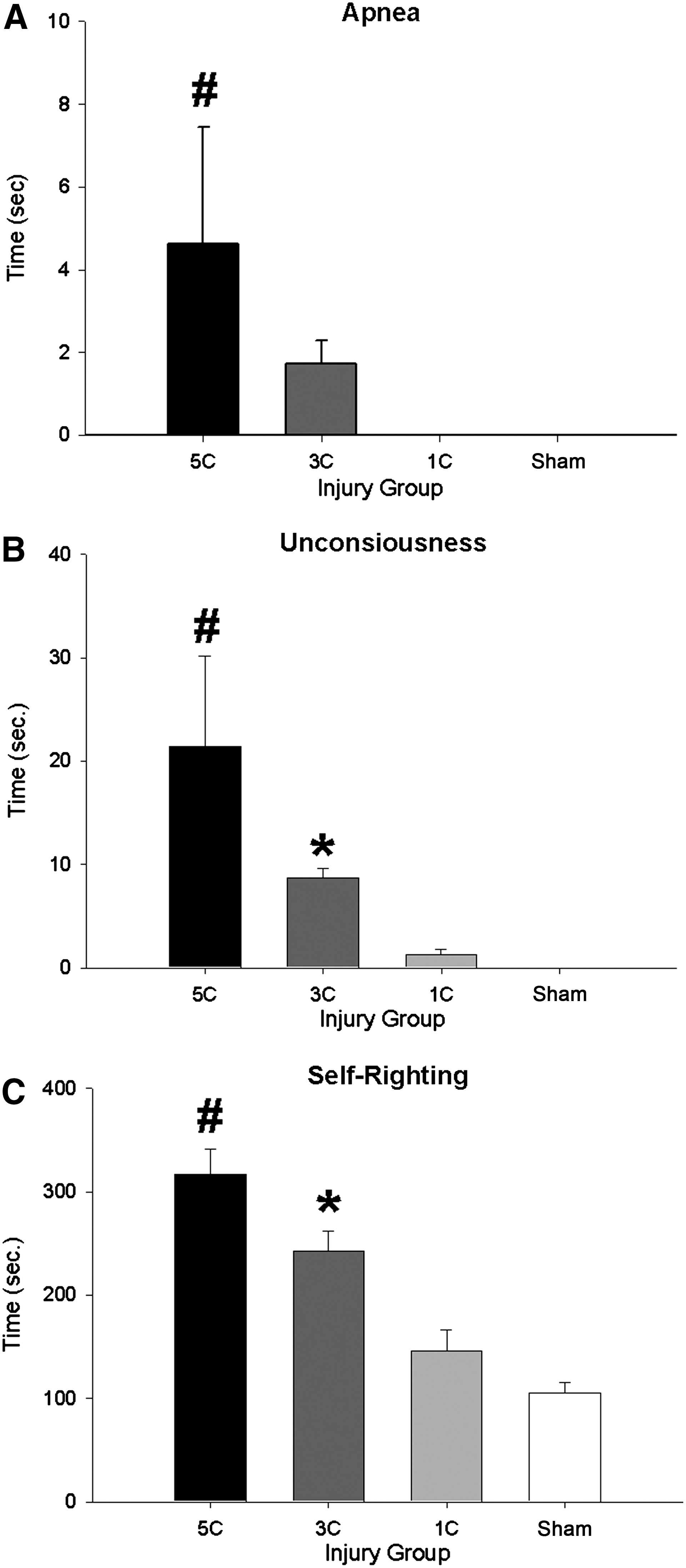
Acute injury measures. (
Elevated-plus maze
The 5C-SR group spent significantly less time in the open arms compared to the sham-SR group, as indicated by a significant effect of injury group (F (3,54)=2.788, p<0.05; 5C-SR < sham-SR rats, p<0.01; Fig. 3A). There were no SR group differences in closed-arm entries (Fig. 3B; p<0.05).
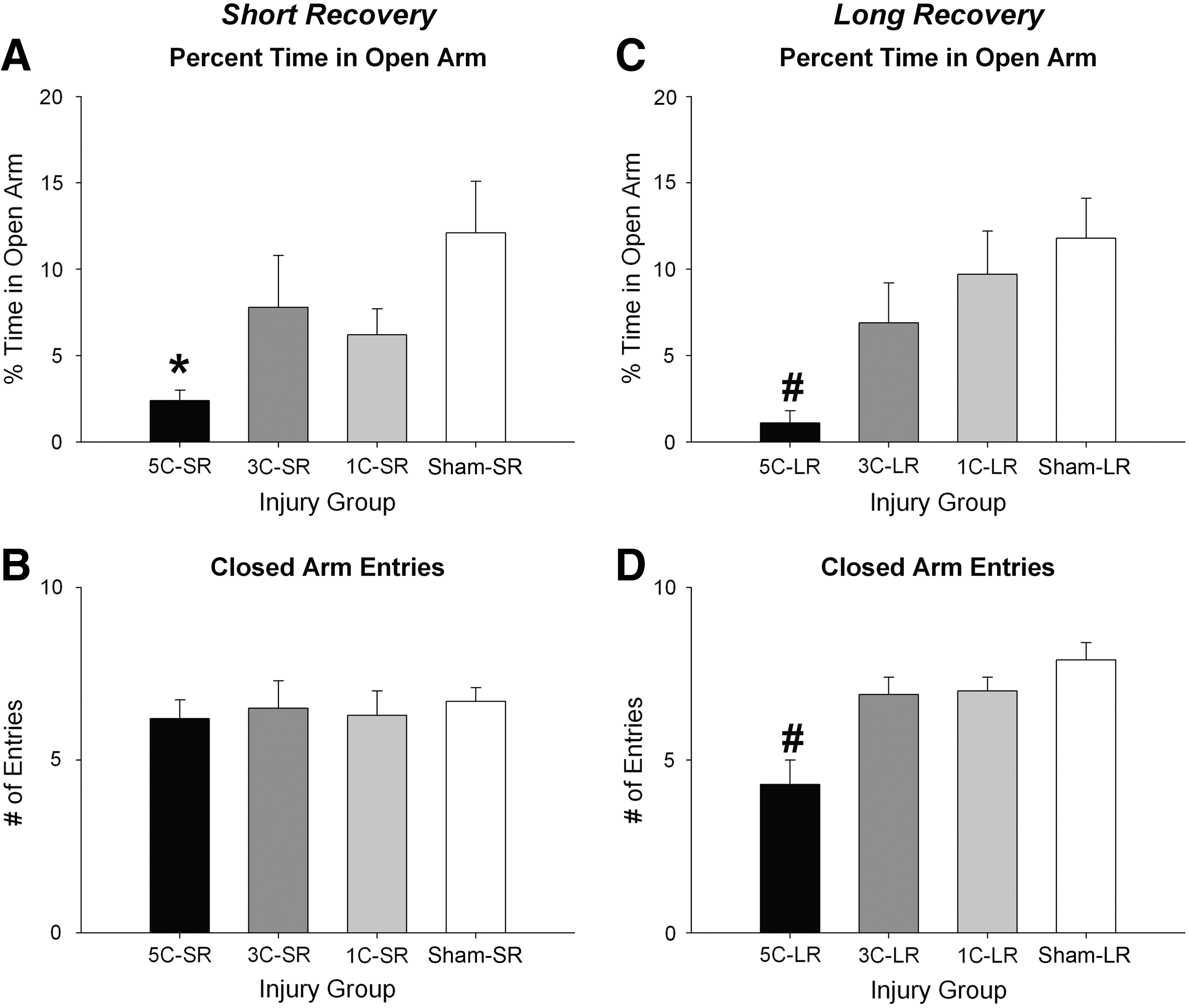
Short and long recovery elevated-plus maze results. (
The 5C-LR group spent significantly less time in the open arms (F (3,47)=5.140, p<0.01; 5C-LR < all other groups, all p<0.05; Fig. 3C), and displayed fewer entries into the closed arms (F (3,47)=10.771, p<0.001; 5C-LR < all other groups, all p<0.001; Fig. 3D) compared to all other groups.
Water maze: Short recovery
During acquisition training search time decreased in all SR groups as testing progressed, as indicated by a significant effect of trial (F (9,486)=11.867, p<0.001). Search time decreased less in all injured groups than in the sham-SR group, as indicated by a significant effect of injury (F (3,54)=8.126, p<0.001; 5C-SR, 3C-SR, and 1C-SR > sham-SR, all p<0.05; Fig. 4A). 5C-SR rats also displayed longer search times than 1C-SR rats (p<0.05; Fig. 4A).
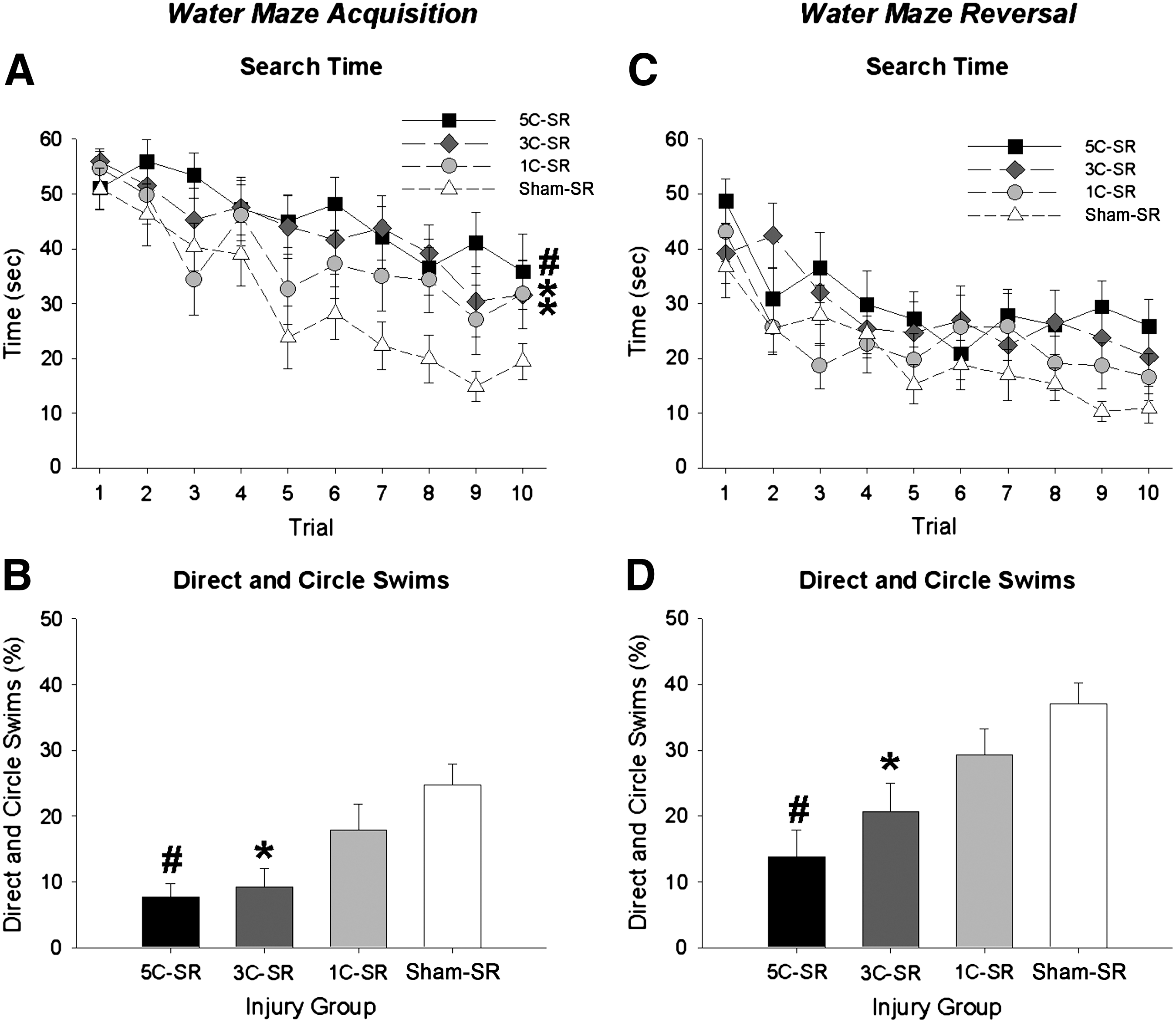
Short recovery water maze results. (
Direct and circle swim data were consistent with the acquisition search time data in revealing fewer direct and circle swims in the 5C-SR and 3C-SR groups during acquisition (F (3,54)=6.769, p<0.001; 5C-SR and 3C-SR < sham-SR, all p<0.01; Fig. 4B), and reversal (F (3,54)=6.786, p<0.001; 5C-SR and 3C-SR < sham-SR group, all p<0.01; Fig. 4D) compared to the sham-SR group. The 5C-SR group also displayed significantly fewer direct and circle swims than the 1C-SR group in acquisition training and reversal training (all p<0.05).
During reversal training search time decreased in all SR groups as testing progressed, as indicated by a significant effect of trial (F (9,486)=10.547, p<0.001; Fig. 4C).
There were no group differences in swim speed during either acquisition or reversal training sessions (all p<0.05; data not shown).
Water maze: Long recovery
Search time decreased in all LR groups as testing progressed during acquisition (F (9,423)=12.135, p<0.001; Fig. 5A), and reversal (F (9,423)=4.595, p<0.001; Fig. 5D). Search time decreased less in the 5C-LR and 3C-LR groups than in the 1C-LR and sham-LR groups during acquisition (F (3,47)=9.793, p<0.001; 5C-LR and 3C-LR > 1C-LR and sham-LR, all p<0.01; Fig. 5A), and reversal (F (3,47)=13.488, p<0.001; 5C-LR and 3C-LR > 1C-LR and sham-LR, all p<0.01; Fig. 5D).
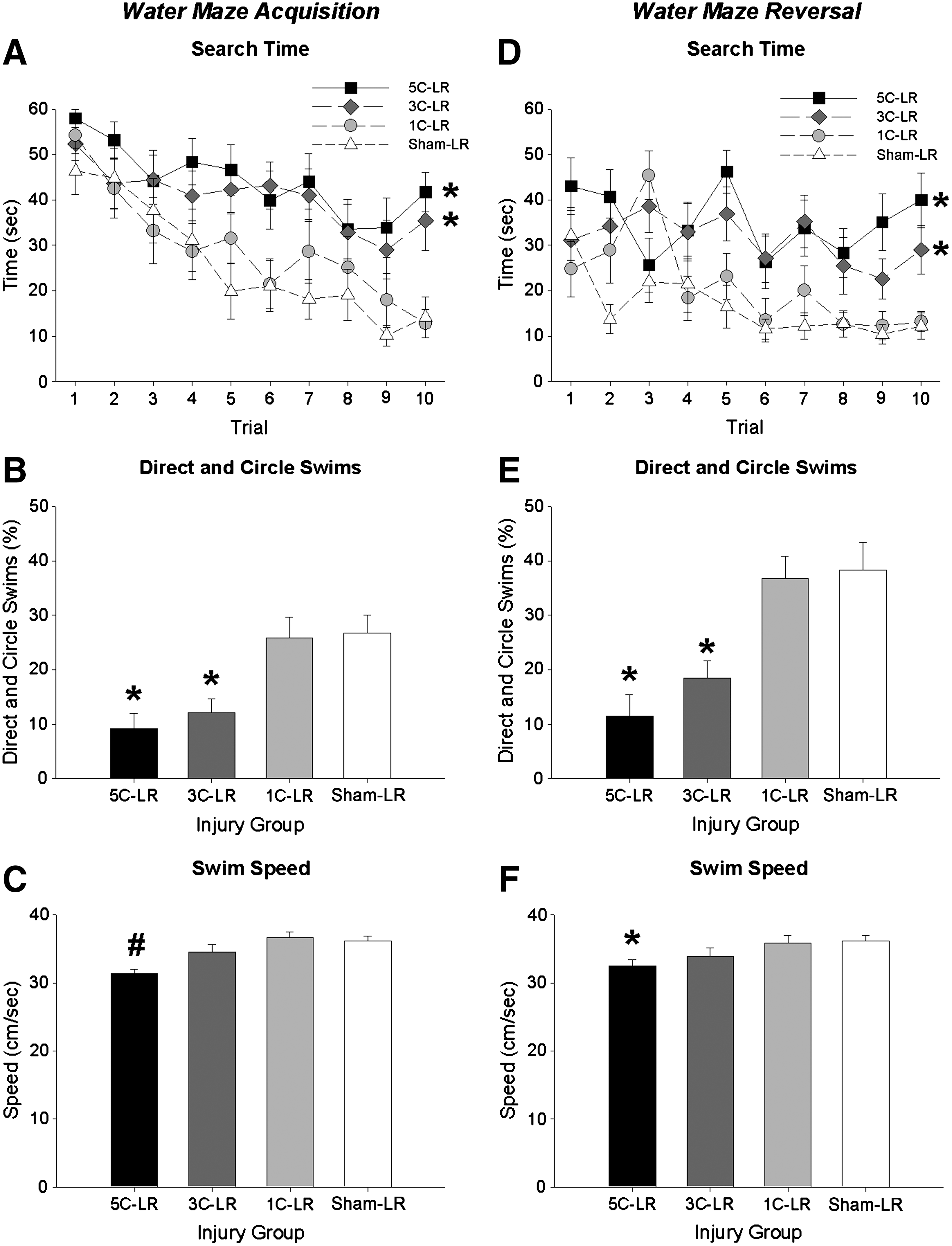
Long recovery water maze results. (
The direct and circle swim data were consistent with the search time data in revealing fewer direct and circle swims in the 5C-LR and 3C-LR groups compared to the 1C-LR and sham-LR groups during acquisition (F (3,47)=8.297, p<0.001; 5C-LR and 3C-LR<1C-LR and sham-LR, all p<0.01; Fig. 5B), and reversal (F (3,47)=10.702, p<0.001; 5C-LR and 3C-LR<1C-LR and sham-LR, all p<0.01; Fig. 5E).
The 5C-LR group displayed slower swim speed during both training sessions, as indicated by a significant effect of injury group during acquisition (F (3,47)=6.998, p<0.01; 5C-LR<all other groups, all p<0.05; Fig. 5C), and reversal (F (3,47)=2.876, p<0.05; 5C-LR<1C-LR and sham-LR, all p<0.05; Fig. 5F).
Forced swim test
The SR groups did not differ on any behavioral measure during forced swim testing (all p>0.05; data not shown).
The 5C-LR group displayed increased time immobile, as indicated by a significant effect of injury group (F (3,47)=2.898, p<0.05; 5C-LR<sham-LR, p<0.01; Fig. 6). There were no group differences on the measures of time spent swimming or escaping (all p>0.05; data not shown).
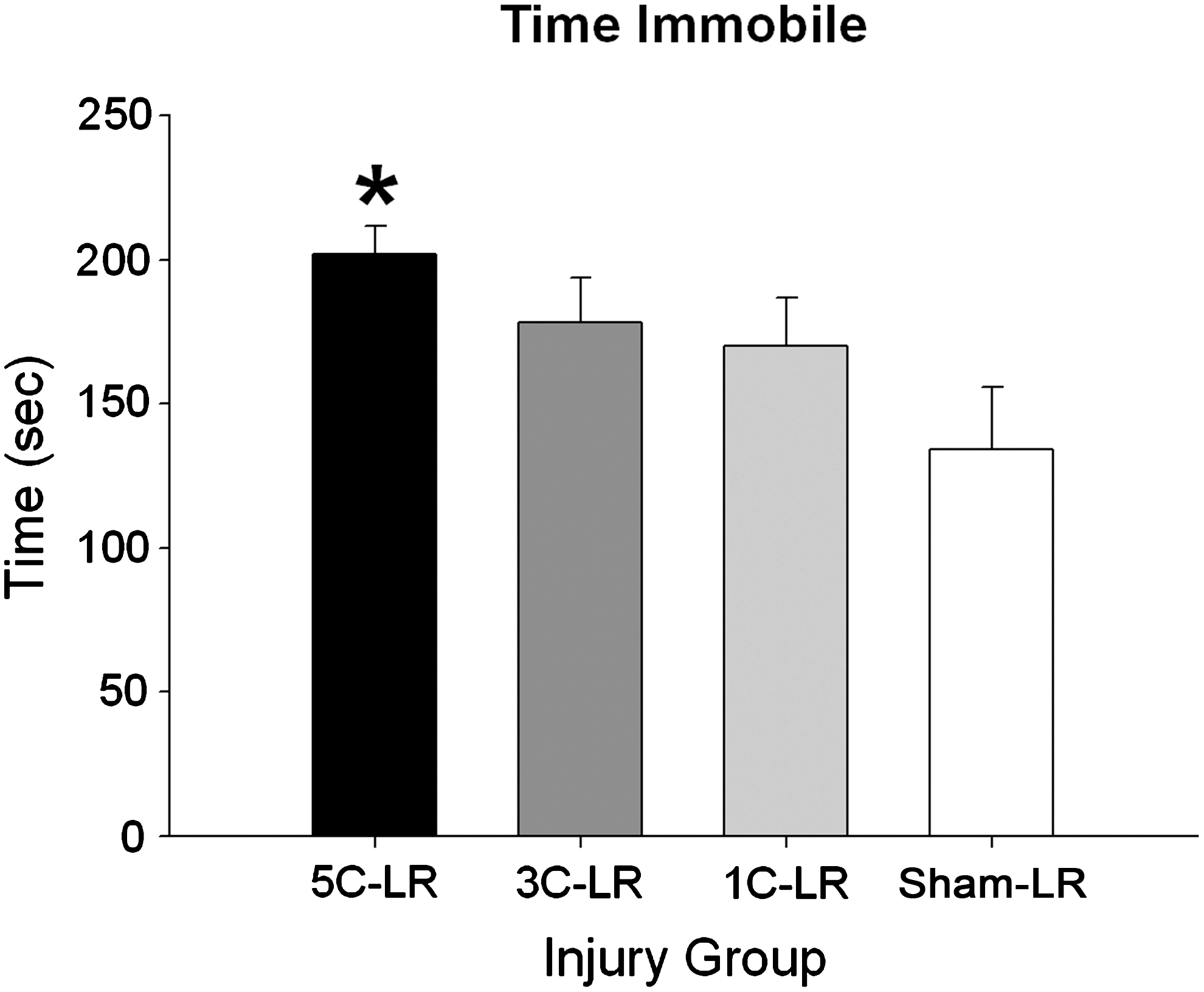
Long recovery forced swim time immobile results. The 5C-LR rats spent significantly more time immobile than the sham-LR group. Histogram bars represent means of data collected during the forced swim test trial (±standard error of the mean; *different from sham-LR animals, p<0.01). For additional statistical detail see the results section (1C-LR, one concussion-long recovery period; 3C-LR, three concussions-long recovery period; 5C-LR, five concussions-long recovery period; sham-LR, sham-long recovery period).
Beam task, open-field, and social behavior
The groups did not differ on any measure in the beam, open-field, or social behavior tasks (all p>0.05; data not shown).
Neuroinflammation
The 5C-SR and 3C-SR groups displayed significantly more ED1-labeled activated microglia/macrophages than both the sham-SR and 1C-SR groups, as indicated by a significant effect of injury group (F (3,23)=13.023, p<0.001; 5C-SR and 3C-SR > sham-SR and 1C-SR, all p<0.01; Fig. 7).
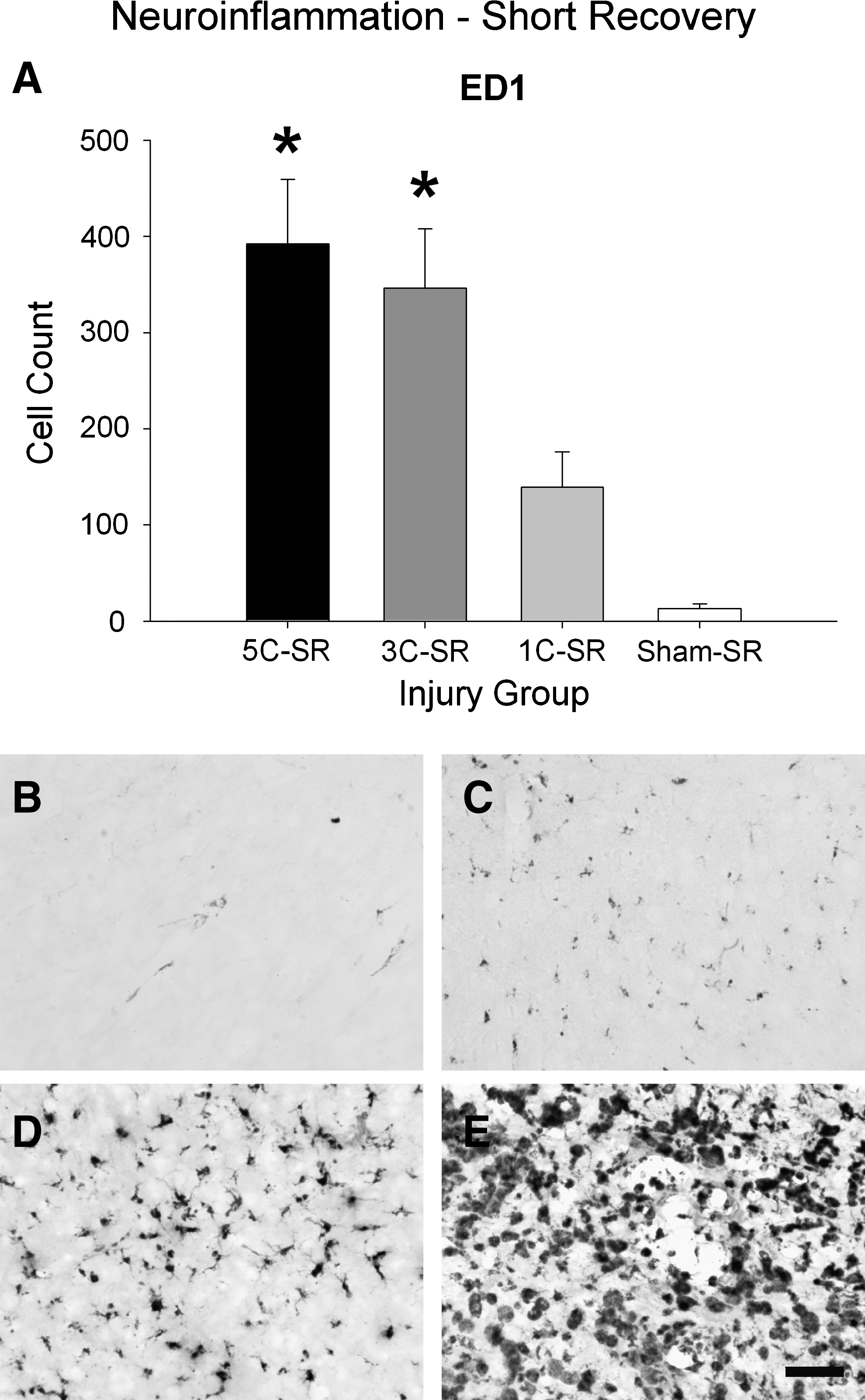
Short recovery ED1 results. (
The 5C-LR group displayed significantly more ED1-labeled activated microglia/macrophages than all the other groups, as indicated by a significant effect of injury group (F (3,22)=7.074, p<0.05; 5C-LR > all other groups, all p<0.05; Fig. 8).
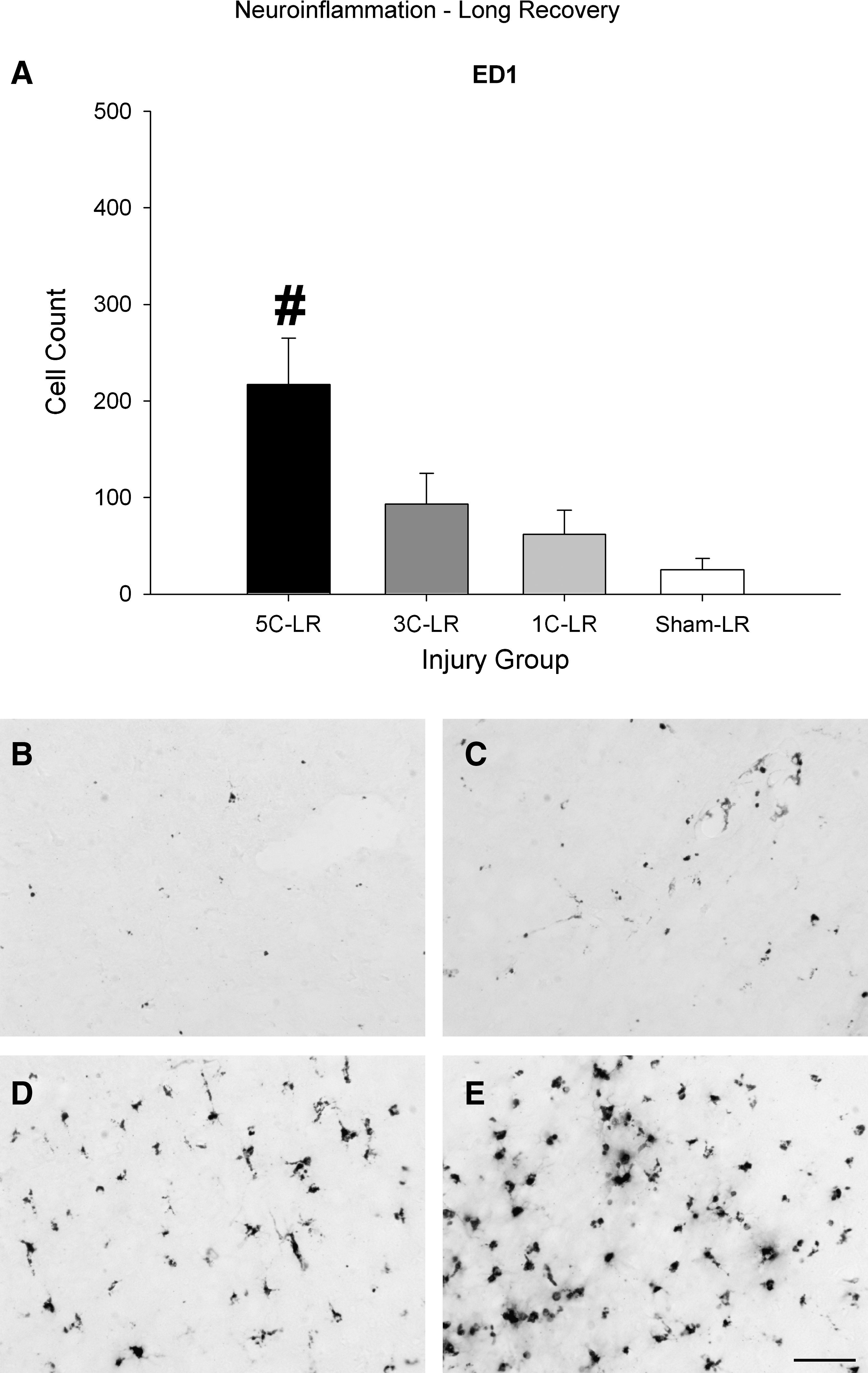
Long recovery ED1 results. (
Cortical damage
Semi-qualitative analysis indicated that approximately 66% of 3C-SR rats displayed moderate cortical damage, and that 50% of 5C-SR rats displayed severe damage (Fig. 9). There was no indication of moderate or severe damage in the 1C-SR and sham-SR groups.

Semi-qualitative analysis of cortical damage. The histogram bars in panels (
Approximately 60% of 3C-LR rats displayed moderate cortical damage and approximately 66% of 5C-LR rats displayed severe cortical damage (Fig. 9). A single 1C-LR rat displayed severe damage. The remaining 1C-LR and sham-LR rats displayed no indication of moderate or severe damage.
Discussion
The present study addressed the question of whether repeated mLFP injuries in the rat result in cumulative effects on anxiety-like behavior, spatial cognition, social behavior, sensorimotor ability, depression-like behavior, neuroinflammation, and brain damage. The findings suggest that repeated mLFP injuries can produce cumulative short- and long-term changes in anxiety-like behavior and spatial cognition, and cumulative long-term changes in depression-like behavior. Both the 5C-SR and 5C-LR groups spent less time in the open arm of the elevated-plus maze, indicating increased anxiety-like behavior. In the water maze task all 5C and 3C groups were impaired on most or all measures relative to sham controls, and the 1C-SR group had scores on certain measures that were intermediate between the 5C-SR and sham-SR groups. The 5C-LR group also displayed significantly more immobility than the sham-LR group in the forced swim task, suggesting increased depression-like behavior. There were no significant between-group differences in locomotion, sensorimotor ability on the beam task, or social behavior. Immunohistochemical examination of brains from rats tested behaviorally found that 5C-SR and 3C-SR rats displayed increased neuroinflammation after short recovery and that increased neuroinflammation was still present after long recovery in 5C-LR rats. Semi-qualitative analysis of cortical damage found that the majority of brains from rats given repeated mLFP displayed moderate or severe cortical damage. Taken together the findings with mLFP in the rat appear to be consistent with many features of repeated concussion observed in humans.
Nature of the behavioral impairments
Past studies that found decreased time spent in the open arm of the elevated-plus maze have interpreted this outcome as an increase in anxiety-like behavior (Jones et al., 2008; Walf and Frye, 2007). The current finding that the 5C groups spent less time on the open arm of the elevated-plus maze is novel in mLFP research, and suggests increased anxiety in these groups. Our use of a specific test for anxiety-like behavior allows us to evaluate the suggestion that cognitive impairment in the water maze might be secondary to anxiety in rats given repeated mLFP (DeRoss et al., 2002). Although the 5C groups displayed both anxiety-like behavior in the elevated-plus maze and impairment in the water maze, both 3C groups displayed water maze impairments in the absence of anxiety-like behavior. This suggests that the repeated mLFP-induced impairment in the water maze task may not be secondary to anxiety, and strengthens the conclusion that repeated concussions can cause cognitive impairment in a rat model.
Cognitive impairments occurred after both single and repeated mLFP treatments. Our finding that 1C-SR, but not 1C-LR, rats were impaired during water maze testing is consistent with similar findings in previous single mLFP water maze studies, and is also consistent with the transient cognitive impairments often observed in humans suffering a single concussion (DeRoss et al., 2002; Gurkoff et al., 2006; McCrory et al., 2009; Shultz et al., 2011). Importantly, the results from both the SR and LR groups show that repeated mLFP can induce cumulative cognitive impairments in the water maze, and the results from the long recovery groups show that the cumulative cognitive impairments persist for at least 8 weeks in the 3C-LR and 5C-LR groups.
Increased time spent immobile in the forced swim task is considered to be indicative of depression-like behavior in the rat (Jones et al., 2008; Porsolt et al., 1977). The current finding that 5C-LR rats spent more time immobile in this task is novel in mLFP research, and suggests an increase in depression-like behavior.
There were no SR group differences in the number of closed arm entries in the elevated-plus maze, swim speed in the water maze, open-field exploration, or beam task measures, suggesting that locomotor abnormalities were not confounding factors in the SR findings. Still, 5C-LR rats displayed fewer closed arm entries in the elevated-plus maze and slower swim speeds during water maze testing. Thus, the possibility that motor deficits contributed to the behavioral deficits observed in the 5C-LR group must be considered. However, Walf and Frye (2007) suggest that arm entries are not an optimal measure of locomotion, and direct measures of sensorimotor ability in the present study found no significant differences between groups in either the open-field task or the beam task. Further, the 5C-SR group spent significantly less time on the open arms of the plus maze, yet made as many closed arm entries as the other SR groups, and the 3C-LR group was impaired to the same extent as the 5C-LR group in the water maze task, yet displayed no indication of locomotor abnormalities. Taken together, the available evidence suggests that it is unlikely that motor-related abnormalities directly accounted for the behavioral changes that occurred in the elevated-plus, water maze, and forced swim task.
Pathology and its relation to behavioral changes
Repeated mLFP produced moderate or severe cortical damage in most rats examined in the current study. Previous studies have found that damage to cortex comparable to that observed here impairs cognitive and emotional function in both humans and rats (Bissiere et al., 2006; Drevets et al., 2008; Jones et al., 2008; Levin et al., 1987; Shamay-Tsoory et al., 2004; Wahl et al., 2000). For example, Wahl and colleagues (2000) found that damage to rat parietal and temporal cortex due to lateral fluid percussion injury resulted in cognitive impairment, and that the impairment varied directly with the amount of damage. In addition, Jones and associates (2008) found that a single severe lateral fluid percussion injury resulted in both cortical damage and increased anxiety-like behaviors. It seems likely that mLFP-induced brain damage contributed to the behavioral changes found in the current study.
Repeated mLFP was also found to induce a neuroinflammatory response consisting of activated microglia/macrophages in the injured cortex. Neuroinflammation has been observed in rat and human traumatic brain injury, with evidence suggesting that these processes might alter normal brain function (Hein and O'Banion, 2009; Lenzlinger et al., 2001; Morganti-Kossmann et al., 2007; Schmidt et al., 2005; Spalletta et al., 2006). For example, previous research from our laboratory and others has revealed a cognitive deficit in the water maze associated with a neuroinflammatory response similar to the one found in the current study (Shultz et al., 2011,2009; Wu et al., 2010). Neuroinflammation also may have contributed to the cortical damage observed in rats given repeated mLFP, as neuroinflammation is capable of inducing secondary brain damage in traumatic brain injury through processes such as apoptosis and the production of reactive oxygen species (Dringen, 2005; Morganti-Kossmann et al., 2007; Schmidt et al., 2005). The finding that 5 mLFPs often resulted in long-term cortical damage in the presence of a neuroinflammatory response may be important for understanding the cumulative and chronic effects of repeated concussion and CTE, as neuroinflammation has been linked to a number of neurodegenerative disorders (Lee et al., 2002; Nandoe et al., 2002; O'Sullivan et al., 2009; Stoll and Jander, 1999; Whitton, 2007; Zilka et al., 2006). Taken together, it seems possible that neuroinflammation could contribute to the behavioral changes and cortical damage found in the current study.
Relation to repeated concussion in humans
Here rats were given repeated mLFPs, with each repeated mLFP separated by a 5-day recovery interval. The cerebral pathophysiology at 5 days after lateral fluid percussion injury in the rat is similar to what occurs at approximately 2–4 weeks post-concussion in humans (Giza and Hovda, 2001). Therefore, this mLFP injury schedule models what might occur in athletes and military personnel under common concussion management guidelines (Maroon et al., 2000). Furthermore, it is not uncommon for athletes to experience multiple concussions and/or hundreds of sub-concussive impacts within a single season, and more than five concussions over the span of a career (Chen et al., 2004; McKee et al., 2009; Pellman et al., 2004). However, it should be noted that it would be rare for any individual to suffer five concussions in a time period similar to the one utilized in the current study while following the more conservative concussion guidelines that are now being implemented (Cantu, 2009).
The current study found that repeated mLFP induced behavioral and neuropathological changes in the rat that are similar to features of repeated concussion in humans. Athletes who experienced three or more concussions displayed more severe acute symptoms, long-term cognitive impairments, and increased incidence of depression compared to athletes who experienced a single concussion, and repeated head trauma can lead to CTE with long-term cognitive impairments, dementia, depression, and anxiety (Cantu, 2007; Guskiewicz et al., 2005; Guskiewicz et al., 2007,2003; McKee et al., 2009). Analysis of post-mortem brain tissue taken from individuals who had sustained repeated concussion and displayed chronic behavioral symptoms have reported that neuronal loss and cortical atrophy are pathologies associated with repeated concussion and CTE (McKee et al., 2009). The current findings of progressively worse acute injury measures, cumulative and long-term cognitive deficits, persistent anxiety-like behavior, long-term depression-like behavior, neuroinflammation, and cortical damage in rats after repeated mLFP are similar to many of the above symptoms and pathologies seen in humans. Taken together, the current findings provide support for the use of repeated mLFP as a model of repeated concussion, and possibly CTE. Future studies might utilize this model to better understand the factors involved in the cumulative and chronic nature of these injuries, and provide insight into their management and treatment through the development and application of new guidelines and therapies.
Conclusions
Repeated mLFP induced cumulative and long-term cognitive deficits, anxiety, and depression in adult male Long-Evans rats. As these injuries also resulted in neuroinflammation and cortical damage, it seems possible that these pathologies may be involved in the behavioral changes observed. These behavioral and pathological changes in rats suffering repeated mLFP are consistent with symptoms and pathologies seen in humans that have experienced repeated concussion, and provide support for the further use of repeated mLFP in the rat to investigate repeated concussion.
Footnotes
Acknowledgments
This research was supported by a grant from the Natural Science and Engineering Research Council (NSERC) to Dr. Donald P. Cain, a scholarship from NSERC to Dr. Sandy R. Shultz, and a grant from CIHR to Dr. Arthur Brown. We thank Drs. R. Sutton and D. Hovda, UCLA Brain Injury Research Center, for training. Additional thanks go to Drs. K.-P. Ossenkopp and D. MacFabe, and to F. Boon, L. Tichenoff, and R. Taylor for technical assistance. Dr. Sandy R. Shultz is now at the Department of Medicine (Royal Melbourne Hospital), Melbourne Brain Centre, University of Melbourne, Parkville, Victoria, Australia.
Author Disclosure Statement
No competing financial interests exist.