
Abstract
Interactions between endothelial and neural stem cells are believed to play a critical role in the kinetics of neural stem cells in the central nervous system. Here we demonstrate that endothelial progenitor cells promote the repair of injured spinal cord through the induction of Notch-dependent astrogliosis and vascular regulation. The transplantation of Jagged1+/+ endothelial progenitor cells, but not Jagged1−/− endothelial progenitor cells, increased the number of reactive astrocytes during the acute phase, and improved functional recovery following spinal cord injury. Expression of the Notch effector Hes5 was upregulated in the injured spinal cord after Jagged1+/+ endothelial progenitor cell transplantation. Furthermore, we found that the Notch ligand Delta-like-1 was highly expressed in Jagged1−/− endothelial progenitor cells. Transplantation of Delta-like-1, as well as Jagged1-overexpressing 3T3 cells, revealed that only Jagged1-overexpressing 3T3 stromal cells enhanced astrogliosis following spinal cord injury. In addition, Jagged1+/+ endothelial progenitor cells exhibited not only dramatic pro-angiogenic effects, but also morphologically abnormal vessel stabilization, compared with Jagged1−/−endothelial progenitor cells in injured spinal cord. Thus, transplanted endothelial progenitor cells promote astrogliosis, vascular regulation, and spinal cord regeneration through activation of Jagged1-Notch signaling.
Introduction
A
Endothelial progenitor cells (EPCs) derived from bone marrow and blood have the potential to differentiate into endothelial cells and to promote neovascularization (Asahara et al., 1997; Isner and Asahara, 1999). We recently reported the involvement of the Notch ligand Jagged1 (Jag1) in EPC kinetics using adult Jag1 conditional knockout mice (Kwon et al., 2008). Here we report that transplantation of Jag1+/+ EPCs promotes astrogliosis, vascular regulation, and functional recovery following SCI, via mechanisms related to Jagged1-Notch signaling.
Methods
The Institutional Animal Care and Use Committee of the RIKEN Center for Developmental Biology approved all animal procedures in this study.
Conditional Jagged1-knockout mice
In this study, we used conditional Jag1-knockout mice (loxP/loxP, Mx-Cre), which were generated and reported on previously (Brooker et al., 2006; Kwon et al., 2008). Mx-Cre, Jag1lox/lox mice were bred and maintained at the animal facility of the RIKEN Center for Developmental Biology. For gene targeting, polyinosinic:polycytidylic acid (poly I:C; 200 μg/200 μL) was administered intravenously four times over a period of 12 days (once every 3 days); 4–6 weeks later deletion of Jag1 was confirmed by Southern blot.
EPC culture
EPCs derived from mouse bone marrow were cultured as previously reported (Kwon et al., 2008; Masuda et al., 2007). Bone marrow mononuclear cells from Jag1+/+ as well as Jag1−/− mice were isolated by density gradient centrifugation and cultured on 6-cm temperature-responsive culture dishes (RepCell; CellSeed, Tokyo, Japan), coated with fibronectin in endothelial cell basal medium-2 (EBM-2; Lonza, Basel, Switzerland), supplemented with 5% fetal bovine serum (FBS), vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF), insulin-like growth factor (IGF), heparin, ascorbic acid, and gentamicin/amphotericin B (3×107 cells/dish). The floating cells were removed at day 4 and the adherent cells were cultured for 3 more days.
Notch Ligand-overexpressing stromal cells
In this study, we used 3T3 stromal cells transfected with retroviral vectors containing Notch ligand genes (Jag1 or Delta-like 1 [Dll-1]), or empty vector as reported previously (Kwon et al., 2008).
Spinal cord injury models
All surgical procedures were performed using an operating microscope (Zeiss, Oberkochen, Germany). Female nude mice (BALB/c-nu/nu, 12 weeks old, weighing 22–24 g) were anesthetized with an intraperitoneal injection of 400 mg/kg 2,2,2-tribromoethanol (Avertin; Sigma-Aldrich, St. Louis, MO). After laminectomy at the 10th thoracic spinal vertebra, we exposed the dura mater. Spinal cord crush injury was performed by compressing the cord laterally from both sides with number 5 Dumont forceps (Fine Science Tools, North Vancouver, B.C., Canada) for 10 sec, as previously reported (Kamei et al., 2010).
Cell transplantation
EPCs (1×106 cells in 200 μL of phosphate-buffered saline [PBS]) derived from Jag1+/+ mice or Jag1−/− mice (Jag1+/+ EPC group and Jag1−/− EPC group) were injected intravenously immediately after injury, as was 200 μL of PBS (PBS group). 3T3 stromal cells transfected with Jag1, Dll-1, or empty vectors (2×105 cells in 2 μL of PBS) were injected into the epicenter of the injured spinal cord at a rate of 1 μL/min using a 5-μL Hamilton syringe immediately after SCI (Jag1-3T3 group, Dll-1-3T3 group, and empty-3T3 group).
Behavioral testing
The recovery of hindlimb motor function was assessed using the Basso mouse scale (Basso et al., 2006). The mice in all groups were assessed before SCI, and 1, 4, 7, 14, 21, 28, 35, and 42 days after injury. The mice were evaluated in an open field by two observers blind to the experimental condition. Results were expressed as mean±standard error. Statistical analysis was performed using two-way repeated-measures analysis of variance (ANOVA) for group×time and Scheffe's post-hoc comparisons.
Electrophysiological recording
Signal conduction in the motor pathway was assessed by motor evoked potentials (MEPs) at 6 weeks after injury as previously described (Kamei et al., 2010). The mice were anesthetized with an intraperitoneal injection of 100 mg/kg ketamine hydrochloride, which has little effect on the MEP (Ghaly et al., 1990). The mice were fixed in a stereotaxic apparatus. A pair of needle electrodes was placed subcutaneously 3 mm on each side of the vertex of the skull. The motor cortex was stimulated transcranially with 0.2-msec square wave pulses using a constant current of 50 mA. The electromuscular responses were recorded from both hamstring muscles using a commercially available system (Viking Quest; Nicolet Biomedical, Madison, WI). All signals were filtered (band pass 0.5–2000 Hz). To ensure reproducibility, at least five replicate responses were recorded, and the recording with the highest amplitude from onset to peak of the negative deflection was used for analysis. The results were expressed as mean±standard deviation (SD). Statistical analysis was performed using one-way ANOVA followed by Scheffe's post-hoc comparisons.
Immunohistochemistry
The mice were anesthetized and transcardially perfused with 4% paraformaldehyde in PBS at days 3, 14, and 42 after SCI. Spinal cords were frozen and sagittally sectioned at 16 μm with a cryostat. Spinal cord sections were stained with the following primary antibodies: mouse anti-nestin (1:100; BD Biosciences, San Jose, CA), rabbit anti-glial fibrillary acidic protein (GFAP, 1:500; Dako, Carpinteria, CA), rabbit anti-tyrosine hydroxylase (TH, 1:500; Chemicon, Temecula, CA), rat anti-CD31 (1:100; Santa Cruz Biotechnology, Santa Cruz, CA), rat anti-CD31 (1:100; BD Biosciences), goat anti-GFAP (1:200; Santa Cruz Biotechnology), goat anti-serotonin transporter (5-HT, 1:500; Immunostar, Hudson, WI). The secondary antibodies (1:500) used were: Alexa-Fluor 594 conjugated goat anti-mouse (Molecular Probes, Eugene, OR), Alexa-Fluor 488 conjugated goat anti-rabbit, Alexa-Fluor 594 conjugated donkey anti-rabbit (1:500), Alexa-Fluor 488 conjugated donkey anti-rat, Alexa-Fluor 488 donkey anti-goat, and Alexa-Fluor 594 donkey anti-goat. Finally, the tissues were counterstained with 4′,6-diamidino-2-phenylindole (DAPI), and observed under a fluorescence microscope (BZ8000; Keyence, Osaka, Japan). For the quantitative assessment of axons, the area of TH+ or 5-HT+ axons at three levels (the caudal region adjacent to the epicenter, 5 mm caudal to the epicenter, and 5 mm rostral to the epicenter) was measured using ImageJ software as previously described (Kaneko et al., 2006; Takami et al., 2002). The diameter and the number of CD31+ vessels, the number of nestin+ or cleaved Notch1+ cells, and the GFAP-negative area were also measured by ImageJ for the quantitative assessment of neovascularization and astrogliosis (Hamzah et al., 2008; Okada et al., 2006). The six sagittal sections per group were used for each analysis. Results were expressed as mean±SD. Statistical analysis was performed using one-way ANOVA followed by Scheffe's post-hoc comparisons.
Real-time RT-PCR analysis
Total RNA was obtained from Jag1+/+ EPCs and Jag1−/− EPCs using the RNeasy Mini Kit (QIAGEN KK, Tokyo, Japan) according to the manufacturer's protocol. Total RNA from spinal cord tissues was also obtained as previously described (Longbrake et al., 2007). At day 3 after SCI, mice were anesthetized and transcardially perfused with 15 mL sterile RNase-free PBS. Spinal cords were rapidly dissected and a 5-mm segment centered on the lesion was removed and homogenized in Trizol (Invitrogen, Carlsbad, CA). RNA was isolated and first-strand cDNA synthesized with the use of the PrimeScriptTM RT reagent kit (TaKaRa, Otsu, Japan) according to the manufacturer's protocol. After the first-strand cDNA was synthesized, real-time quantitative PCR was performed with an ABI Prism 7700 (Applied Biosystems, Foster City, CA), using SYBR Green Master Mix reagent (Applied Biosystems), according to the manufacturer's protocol. The mRNA expressions were calculated for each gene by the 2-ΔΔCT method, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) for normalization (Livak and Schmittgen, 2001). Results were expressed as mean±SD. Statistical analysis was performed using one-way ANOVA followed by Scheffe's post-hoc comparisons. Each primer sequence is shown in Table 1.
GAPDH, glyceraldehyde-3-phosphate dehydrogenase; Jag1, Notch ligand Jagged1; Dll-1, Delta-like 1.
In vitro migration assay
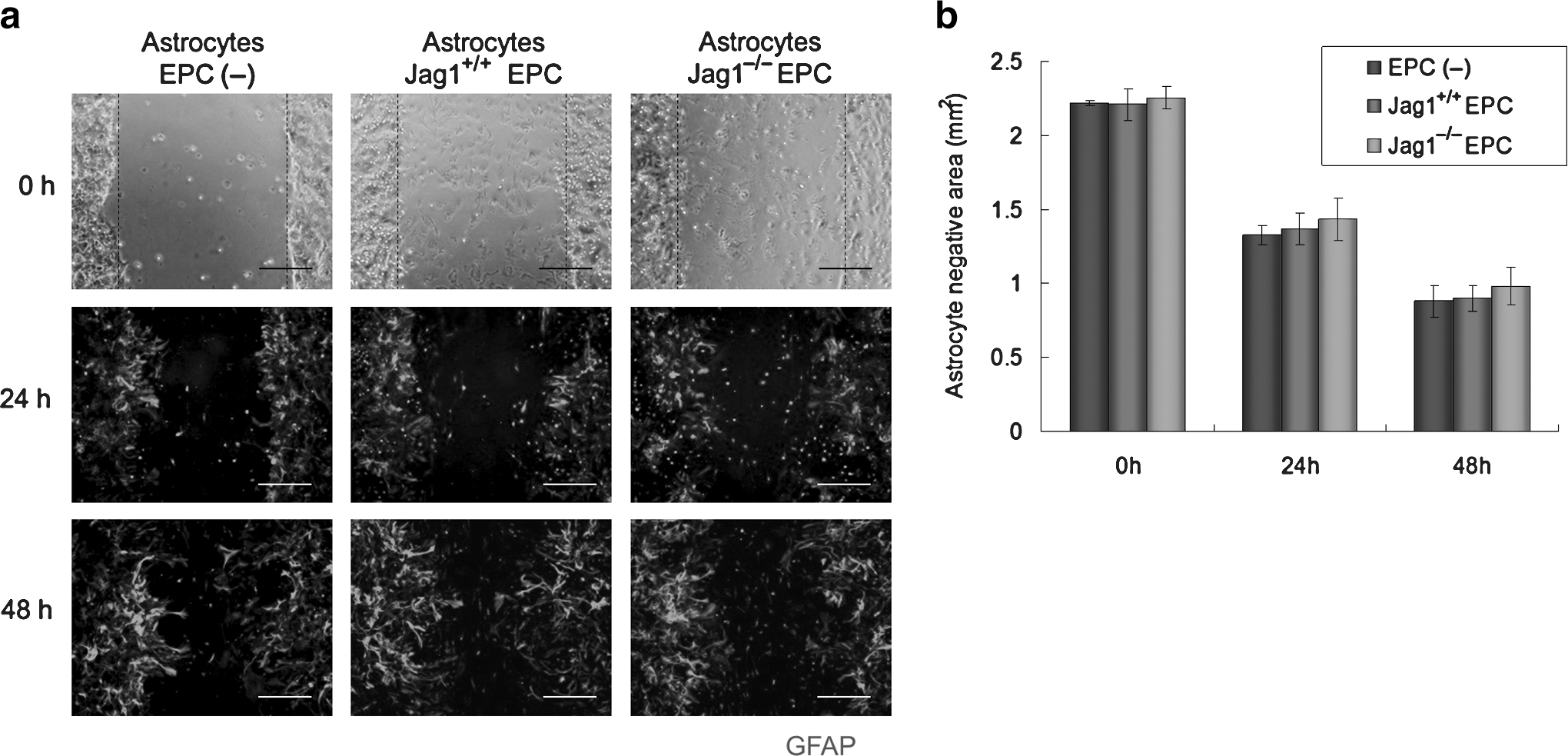
Migration assay of astrocytes was performed by the scratching method as previously reported (Okada et al., 2006). Astrocytes were plated to confluency on cover-slips coated with poly-L-lysine. Astrocytes with Jag1+/+ EPCs and Jag1−/− EPCs were also plated. After immunostaining with GFAP, cell-free areas were evaluated at 24 h and 48 h.
Results
Notch ligands in cultured EPCs
The levels of expression of Notch ligands, including Jag1 and Delta-like 1 (Dll-1), in cultured EPCs were confirmed by quantitative real-time PCR. Jag1−/− EPCs lack expression of Jag1, whereas no significant difference in the expression of Dll-1 could be detected between Jag1−/− and corresponding wild-type EPCs (Fig. 1a).

(
Functional recovery after spinal cord injury
The recovery of hindlimb function was assessed using the Basso mouse scale (BMS; Fig. 1b). All mice had a score of 9 (maximum score) before SCI, with the score being reduced to 0 at 1 day after SCI (n=8/group). The BMS score in the Jag1+/+ EPC group was significantly higher than in the PBS and the Jag1−/− EPC groups at days 14 or later. Although the average score in the Jag1−/− EPC group was slightly higher than in the PBS group at day 14 or later after SCI, there was no significant difference in the BMS score between these two groups. Hindlimb functions while walking at days 1 and 28 after injury are shown in the supplementary movies (Supplementary Movies 1–3; see online supplementary material at
Incorporation of transplanted EPCs
To assess the incorporation of administered EPCs into the injured spinal cord, EPCs were labeled prior to administration by uptake of acetylated low-density lipoprotein conjugated with 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (DiI-acetyl LDL). Spinal cord sections were observed 3 days after SCI under a fluorescence microscope. DiI+ cells could be observed around the injury site in both the Jag1+/+ EPC and Jag1−/− EPC groups; however, no DiI+ cells were detected in the PBS group (Fig. 2a). Almost all of the DiI+ cells were co-labeled with CD31 in both the Jag1+/+ EPC and Jag1−/− EPC groups (Fig. 2b). Further, the number of DiI+ cells was counted per high-power field (HPF) at 200×magnification using Image J software. No significant difference in the number of incorporated DiI+ cells was detected between these two groups (Fig. 2c).
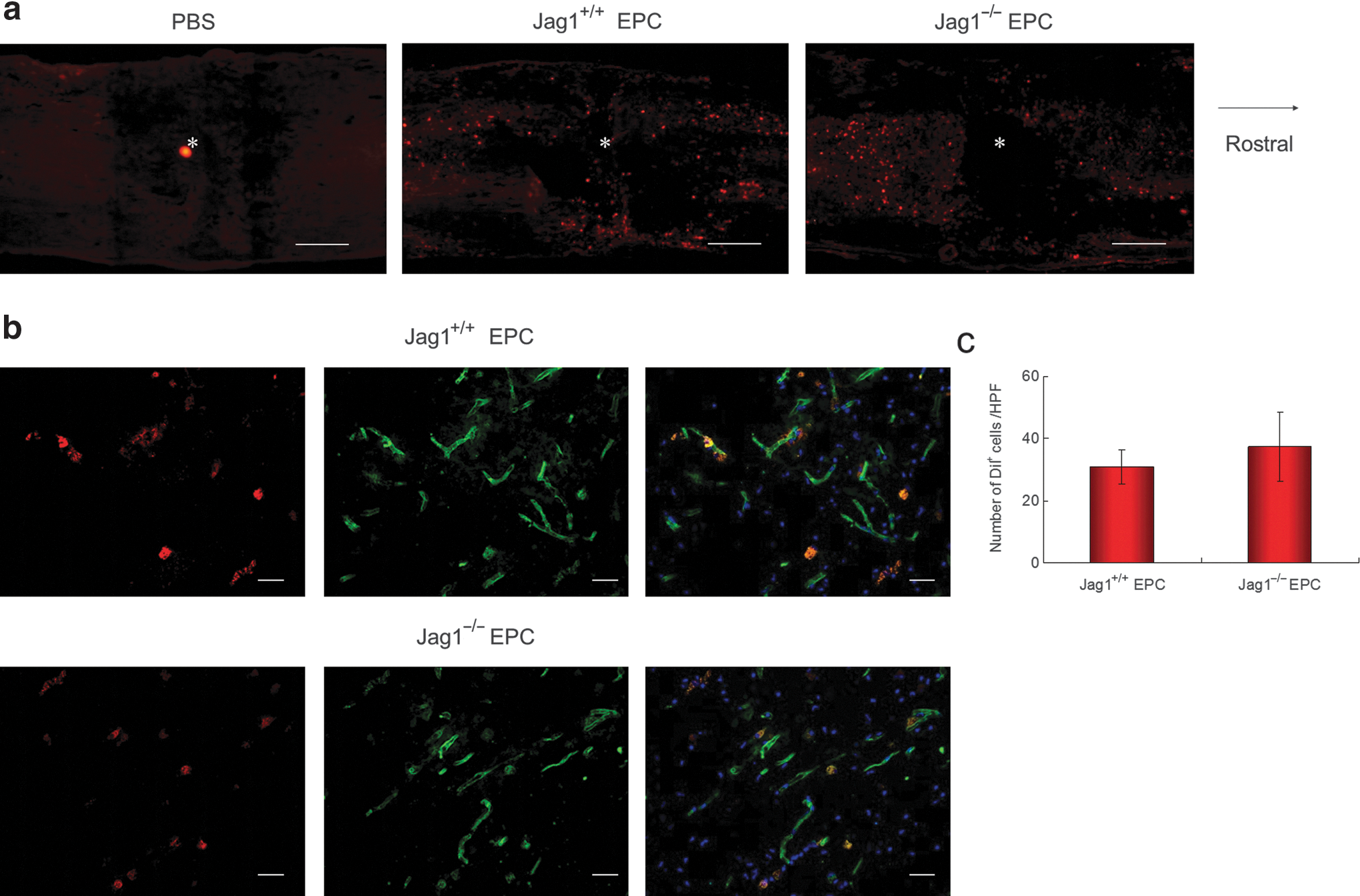
Incorporation of administered DiI+ EPCs (red) into spinal cord. (
Astrogliosis following spinal cord injury
At day 3 after SCI, reactive astrocytes were stained with antibodies against GFAP (green in Fig. 3a) and nestin (red in Fig. 3a). Gliogenesis was assessed around the site of injury (dotted-line box in Fig. 3a). A large number of reactive astrocytes were observed in the Jag1+/+ EPC group, whereas only a few could be detected in the PBS and Jag1−/− EPC groups (Fig. 3a). The number of nestin+ cells in the Jag1+/+ EPC group was significantly greater than in the PBS and the Jag1−/− EPC groups, whereas the number of nestin+ cells in the Jag1−/− EPC group was not significantly different from the PBS group (Fig. 3b). The mRNA expression of the Notch effectors Hes1 and Hes5 in the injured spinal cord were assessed by real-time quantitative PCR. Although the expression level of Hes1 in the Jag1+/+ EPC group was slightly higher than in the PBS and Jag1−/− EPC groups, no significant difference in Hes1 expression could be detected. In contrast, the expression level of Hes5 in the Jag1+/+ EPC group was significantly higher than in the PBS and the Jag1−/− EPC groups, again with no significant difference between the PBS and Jag1−/− EPC group (Fig. 3c). At day 14 after injury, spinal cord tissues were stained with GFAP for the assessment of astrogliosis (Fig. 3d). The GFAP-negative area in the Jag1+/+ EPC group was significantly smaller than in the PBS and Jag1−/− EPC groups, again with the latter showing no significant difference (Fig. 3e). To assess the effect of Jag1/Notch signaling on the migration of astrocytes, an in vitro migration assay was performed in astrocytes only and astrocytes co-cultured with Jag1+/+ EPCs or Jag1−/− EPCs. There was no significant difference in the migration of GFAP+ astrocytes among all groups after 24 and 48 h of culture (Fig. 4).
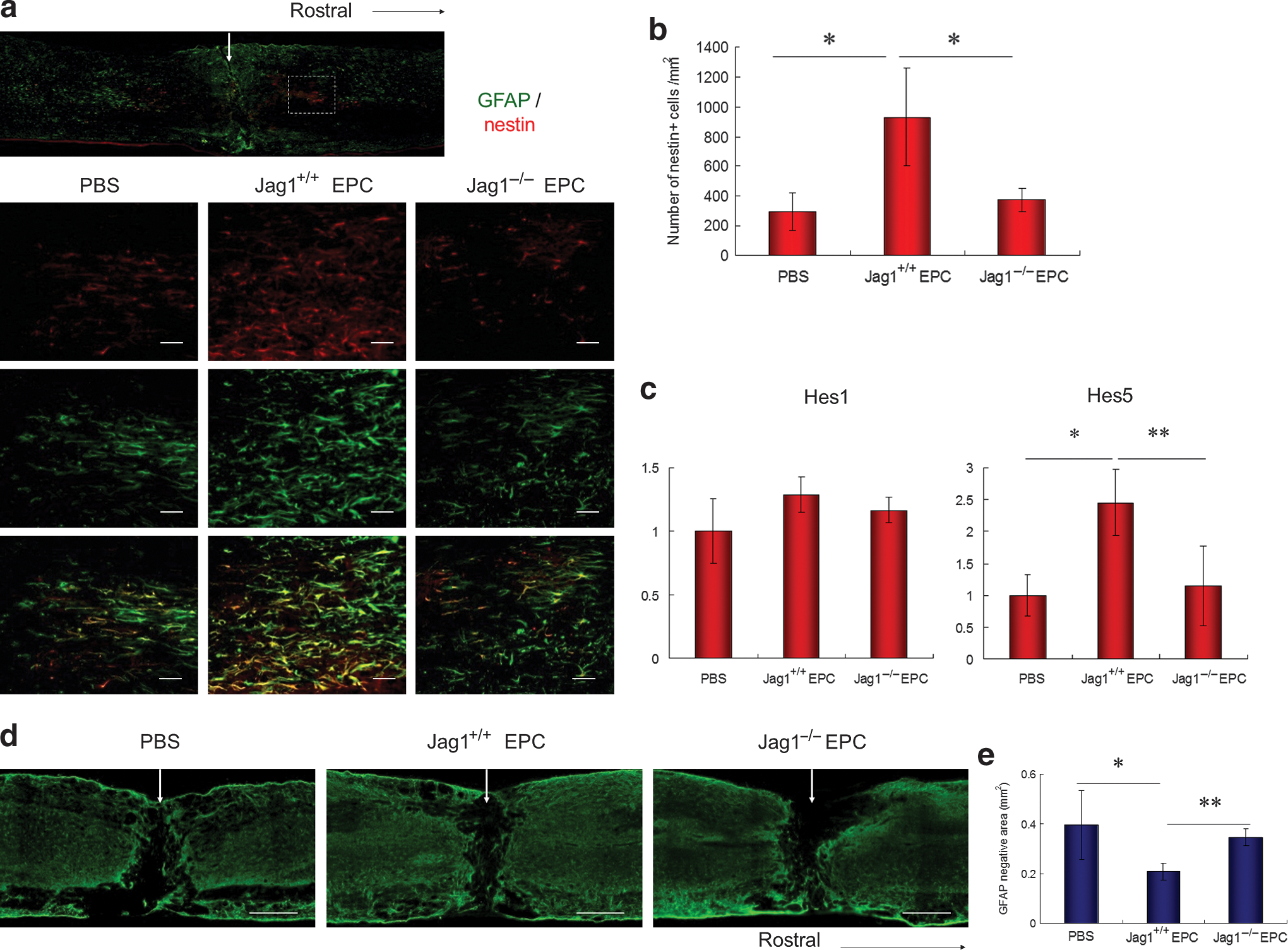
Assessment of astrogliosis. (
(
Neovascularization in the injured spinal cord
Spinal cord sections were immunostained with CD31 at days 3 and 14 after injury in order to assess the degree of neovascularization. CD31+ vessels were assessed at the epicenter of the injury site (epicenter zone; dotted-line boxes in Fig. 5a and c), and the region adjacent to the epicenter zone (border zone; dotted-line boxes in Fig. 5a and b). At day 3 after SCI, although little or no angiogenesis was observed at the epicenter zone in all groups, CD31+ vessels were observed in the border zone. In this area, the mean diameter of the vessels in the Jag1+/+ EPC group was significantly greater than in the PBS and Jag1−/− EPC groups. The number of vessels in the Jag1+/+ and Jag1−/− EPC groups were significantly greater than in the PBS group, whereas no significant difference could be observed between the two EPC groups (Fig. 5b and d). At day 14 after injury, the morphological properties of the CD31+ vessels located in the epicenter zone, which was at the same time devoid of astrocytes, were different from the ones in the border zone, which were covered with astrocytes (Fig. 5a). No obvious morphological differences in the vessels in the border zones could be found among all analyzed groups (Fig. 5f). Blood vessels were therefore assessed at the epicenter zone (dotted-line boxes in Fig. 5a and c). In the PBS and Jag1−/− EPC groups, a chaotic vascular architecture with large vessels adjacent to small vessels was observed, though no morphological abnormalities of the vessels could be observed for the Jag1+/+ EPC group (Fig. 5c). Mean diameters of the vessels in the PBS and Jag1−/− EPC groups were significantly greater than the values found for the gray matter of non-injured spinal cords (Normal in Fig. 5c), and the Jag1+/+ EPC group, with the latter two showing similar values (Fig. 5e). On the other hand, the average number of vessels in the Jag1+/+ EPC group was significantly reduced compared to the PBS group (Fig. 5e). These results suggest that both Jag1+/+ EPCs and Jag1−/− EPCs enhanced angiogenesis at day 3 after SCI, and that Jag1+/+ EPCs but not Jag1−/− EPCs, had normalized vascular morphology at day 14 after SCI.

Assessment of vascularity. (
Immunohistochemical assessment of axons
To analyze defined subsets of descending axons, immunohistochemistry for tyrosine hydroxylase (TH, a marker for descending noradrenergic and dopaminergic axons as well as sympathetic fibers), and 5-HT (a marker for descending serotonergic axons) was performed at day 42 after SCI as previously reported (Biernaskie et al., 2007). To evaluate the axon growth beyond the injury site, quantitative assessment of axons was performed by measuring the area of immunostained axons in the far rostral region (5 mm rostral to the epicenter), the injury site (the caudal region adjacent to the epicenter), and the far caudal region (5 mm caudal to the epicenter; dotted-line boxes in Fig. 6a). In the far rostral region, no significant difference in the area of TH+ and 5-HT+ axons was found among all analyzed groups (Fig. 6b–e). At the injury site, the area of TH+ and 5-HT+ axons in the Jag1+/+ EPC group was significantly larger than in the PBS and Jag1−/− group. In addition, the area of 5-HT+ axons in the Jag1−/− EPC group was significantly larger than in the PBS group. In the far caudal region, TH+ and 5-HT+ axons were clearly observed in the Jag1+/+ EPC group, whereas these axons were rare in the PBS and Jag1−/− EPC group. The area of TH+ or 5-HT+ axons in the Jag1+/+ EPC group was significantly larger than in the PBS and Jag1−/− groups (Fig. 6b–e).

(
Transplantation of Notch ligand-overexpressing 3T3 stromal cells
To clarify the role of ligand-specific Notch activation in the enhancement of astrogliosis following SCI, 3T3 stromal cells transfected with Jag1, Dll-1, or empty vector were transplanted directly into the injury site of spinal cords immediately after SCI (Jag1-3T3 group, Dll-1-3T3 group, and empty-3T3 group). At day 3 after injury, spinal cord sections were immunostained with GFAP and nestin. In the assessment of reactive astrocytes stained with GFAP and nestin (dotted-line box in Fig. 7a), a large number of reactive astrocytes was observed in the Jag1-3T3 group, though these cells were rare in the empty-3T3 and Dll-1-3T3 groups (Fig. 7a). The number of nestin+ cells in the Jag1-3T3 group was significantly greater than in the empty-3T3 and Dll-1-3T3 groups (Fig. 7b). These findings suggest that Jag1/Notch signaling, but not Dll-1/Notch signaling, enhanced astrogliosis after SCI.
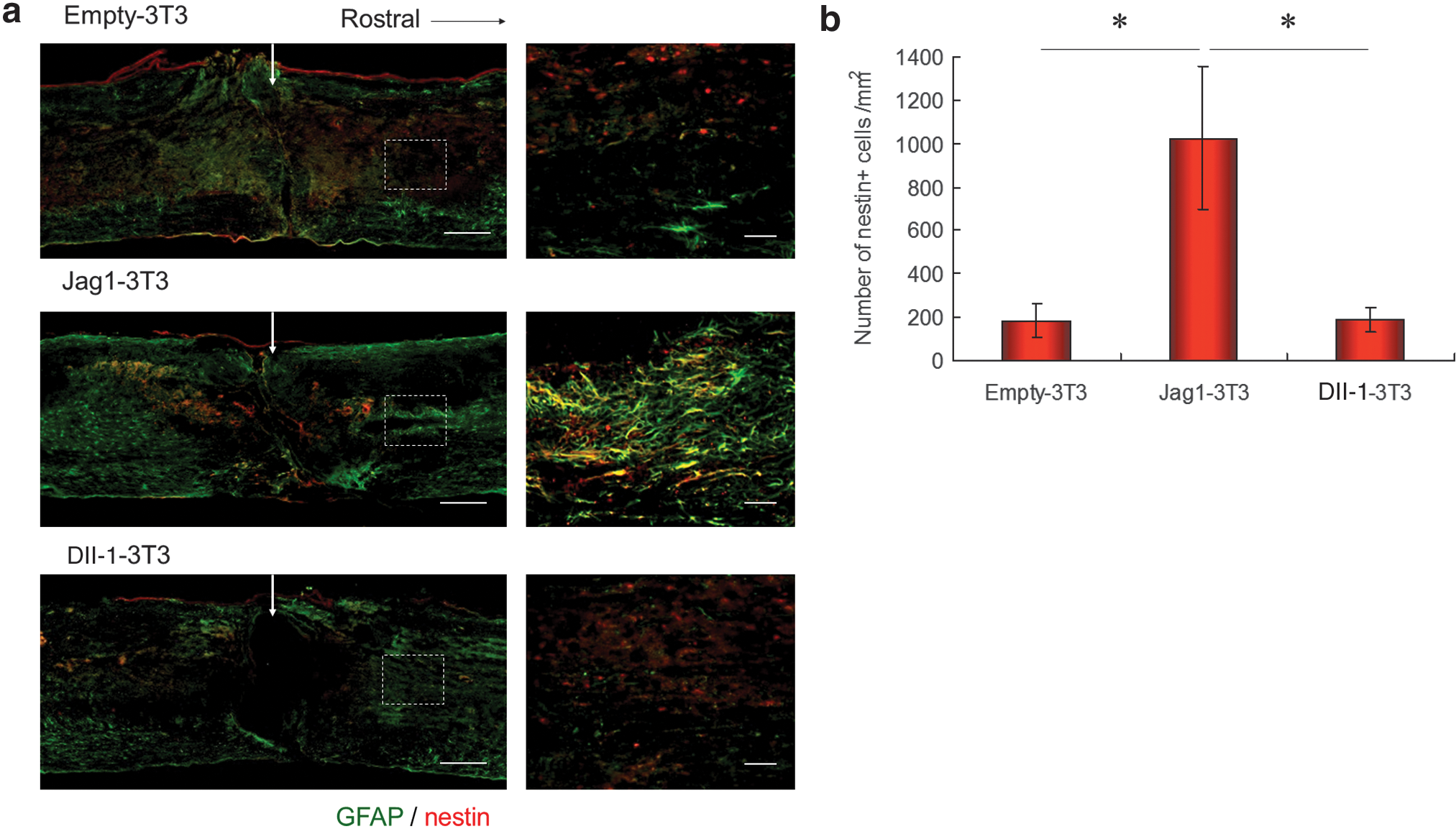
(
Discussion
The current study demonstrates that transplantation of wild-type EPCs promotes astrogliosis and functional recovery following SCI, and that Jag1 deletion in EPCs significantly fails to demonstrate these effects.
Recruitment of EPCs
Intravenously transplanted EPCs were recruited into injured spinal cord tissues in this study. There was no significant difference in the number of incorporated EPCs between the Jag1+/+ EPC group and the Jag1−/− EPC group. These findings suggest that Jag1 expression in EPCs has no effect on cell recruitment. A previous study reported that EPCs were recruited to ischemic or injured tissues by signaling of growth factors and cytokines (Isner and Asahara, 1999). Since stem/progenitor cell mobilization factors, including stem cell factor (SCF), VEGF, and stromal cell derived factor-1 (SDF-1) are upregulated in injured spinal cord tissues in mouse SCI models (Kamei et al., 2010), we speculated that intravenously administered EPCs would be recruited to injured spinal cord tissues by these chemokine signals.
Jagged1-Notch signaling promotes astrogliosis following SCI
Transplantation of Jag1+/+ EPCs, but not Jag1−/− EPCs, enhanced astrogliosis at day 3 after SCI. At the same time, the mRNA expression level of Hes5, which is a well-known critical downstream effector of Notch (Ohtsuka et al., 1999), in the injured spinal cords treated with Jag1+/+ EPCs was significantly higher than in the injured spinal cords treated with Jag1−/− EPCs or PBS alone. No significant difference in the expression of the Notch activator Dll-1 between Jag1−/− EPCs and Jag1+/+ EPCs could be found. From these findings, we speculate that enhanced astrogliosis after transplantation of EPCs might be mediated via Jag1-derived Notch signaling. Notch signaling is believed to inhibit neurogenesis and induce gliogenesis during embryonic development (Yoon and Gaiano, 2005). Previous studies showed that multipotent neural stem cells exist in adult spinal cord (Horner et al., 2000; Johansson et al., 1999), and that Notch signaling inhibits neurogenesis and enhances gliogenesis from adult rat spinal cord-derived neurosphere cells in vitro (Yamamoto et al., 2001). Notch signaling is also important for the maintenance of neural stem cells. Ohtsuka and associates reported that the expression of Hes1 and Hes5 plays an important role in the maintenance of neural stem cells, but is dispensable for gliogenesis in the embryonic telencephalon (Ohtsuka et al., 2001). Some previous studies showed that Hes1 does not promote gliogenesis, but it inhibits both neuronal and glial differentiation (Ishibashi et al., 1994; Sasai et al., 1992). In contrast, Hojo and colleagues reported that Hes5 is specifically expressed by differentiating Müller glial cells in the retina, and plays a critical role in the differentiation of these cells towards a glial cell fate (Hojo et al., 2000). Although ligand-specificity of Notch signal activation in the central nervous system has not been clarified, recent reports suggest that each Notch ligand might independently communicate with the receptor, leading to the activation of a separate signaling cascade even in the same cell, as has shown for hematopoiesis, vasculogenesis, and inner ear regeneration (Brooker et al., 2006; Kwon et al., 2008; Rutz et al., 2005). To confirm the ligand-specific function of Notch signaling for the promotion of astrogliosis in the injured spinal cord, we transplanted specific Notch ligand-overexpressing 3T3 stromal cells directly into the injured spinal cord. Jag1-overexpressing 3T3 cells accelerated astrogliosis, whereas transplantation of 3T3 cells transfected with Dll-1 or empty vector showed no effect. These findings indicate a Jag1-specific effect on the promotion of astrogliosis in the injured spinal cord. At day 14 after injury, the GFAP-negative area in the epicenter of the injury site was significantly reduced after transplantation of Jag1+/+ EPCs, but not Jag1−/− EPCs. However, neither Jag1+/+ EPCs nor Jag1−/− EPCs enhanced the migration of astrocytes in vitro. These findings suggest that Jag1/Notch signaling from EPCs does not enhance migration of reactive astrocytes but that it enhances astrogliosis. On the other hand, previous studies showed that vascular endothelial cells or olfactory ensheathing cells promoted proliferation of neural stem/progenitor cells through activation of Notch signaling (Shen et al., 2004; Zhang et al., 2008). We therefore speculate that transplanted EPCs enhanced proliferation of nestin-positive neural progenitor cells via Jag1-Notch dependent mechanisms in the present study. Glial scars, formed in part by reactive astrocytes after SCI, have long been considered detrimental to the repair of the injured spinal cord, with glial scars and their products acting as physical and chemical barriers to axonal regeneration (Barrett et al., 1984; Bradbury et al., 2002; Menet et al., 2003). However, recent studies have shown that reactive astrocytes are also important in supporting the repair of injured spinal cord (Bush et al., 1999; Faulkner et al., 2004; Okada et al., 2006). Okada and associates showed that limited migration of reactive astrocytes causes widespread infiltration of inflammatory cells and limited functional recovery, whereas rapid migration of reactive astrocytes restricts inflammatory cells and results in notable improvements and functional recovery after SCI (Okada et al., 2006). The enhancement of astrogliosis during the acute or subacute phase of SCI is considered to be beneficial for repair of the injured spinal cord.
Jagged1-Notch signaling by EPCs promotes vascular regulation
Although the transplantation of Jag1+/+ EPCs as well as Jag1−/− EPCs enhanced angiogenesis at day 3 after injury, almost all of the CD31+ vessels were not composed of DiI+ cells, indicating a lack of transplanted EPCs. Therefore, almost all newly formed CD31+ vessels must be composed of endogenous endothelial cells. A previous study reported that EPCs expressed several angiogenic growth factors and promoted angiogenesis (Ii et al., 2005; Rehman et al., 2003; Urbich et al., 2005). We speculate that these growth factors derived from transplanted cells enhanced angiogenesis. The angiogenesis in the Jag1−/− EPC group was less prominent than in the Jag1+/+ EPC group. We previously reported that Jag1 expression in EPCs contributed to the function of EPCs in promoting neovascularization (Kwon et al., 2008). The results regarding angiogenesis after EPC transplantation obtained in the present study are consistent with the findings of our previous study. On the other hand, abnormally-enlarged vessels were observed in the epicenter of the injury site, which was devoid of astrocytes in the PBS and the Jag1−/− EPC groups at day 14 after SCI. These vessels resembled abnormal vessels usually seen in tumors, CNS neurodegenerative diseases, or after VEGF treatment of the injured brain, resulting in vascular hyperpermeability and blood–brain barrier dysfunction (Hamzah et al., 2008; Krum et al., 2002; Zacchigna et al., 2008). In the CNS not only pericytes, but also astrocytes are required for the formation of healthy blood vessels (Zacchigna et al., 2008). Therefore we speculate that abnormally enlarged vessels in the epicenter of the injury site, which was devoid of astrocytes, might be functionally abnormal. Interestingly, the diameter of the vessels in the spinal cord treated with Jag1+/+ EPCs was similar to the values obtained for gray matter of non-injured spinal cords. Notch signaling is essential for normal vascular morphogenesis during embryonic development (Krebs et al., 2000). In particular, Notch activation by Jag1 and Dll-4 is critical for vascular stabilization and maturation (Hellstrom et al., 2007; High et al., 2008). In the present study, Jag1-mediated Notch signaling from transplanted EPCs might contribute to normalization of vascular morphology in the epicenter of the injury site at day 14 after injury. The previous study showed that blood–brain barrier dysfunction allows infiltration of inflammatory cells and inhibits the axon growth in the CNS (Horn et al., 2008). Based on these findings, we speculate that normalization of vascular morphology at the injury site might also contribute to the promotion of functional recovery after SCI.
Neuroprotective effects of EPCs
The previous study reported that EPCs expressed several growth factors, including VEGF, hepatocyte growth factor, granulocyte colony-stimulating factor, and IGF-1 (Ii et al., 2005; Rehman et al., 2003; Urbich et al., 2005). These growth factors were reported to have neuroprotective effects after spinal cord injury (Hung et al., 2007; Kitamura et al., 2007; Nishio et al., 2007; Widenfalk et al., 2003). Therefore neuroprotective effects of transplanted EPCs might also contribute to functional recovery after spinal cord injury.
Footnotes
Acknowledgments
We would like to thank Miki Horii and Ayumi Yokoyama for their technical support. We would like to express our appreciation to the animal facility of the RIKEN Center for Developmental Biology for the use of their facilities. This work was supported by a grant-in-aid for scientific research from the Japan Ministry of Education, Culture, Sports, Science and Technology (no. 21791399 to N.K.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.