
Abstract
A penetrating traumatic brain injury (pTBI) occurs when an object impacts the head with sufficient force to penetrate the skin, skull, and meninges, and inflict injury directly to the brain parenchyma. This type of injury has been notoriously difficult to model in small laboratory animals such as rats or mice. To this end, we have established a novel non-fatal model for pTBI based on a modified air rifle that accelerates a pellet, which in turn impacts a small probe that then causes the injury to the experimental animal's brain. In the present study, we have focused on the acute phase and characterized the tissue destruction, including increasing cavity formation, white matter degeneration, hemorrhage, edema, and gliosis. We also used a battery of behavioral models to examine the neurological outcome, with the most noteworthy finding being impairment of reference memory function. In conclusion, we have described a number of events taking place after pTBI in our model. We expect this model will prove useful in our efforts to unravel the biological events underlying injury and regeneration after pTBI and possibly serve as a useful animal model in the development of novel therapeutic and diagnostic approaches.
Introduction
P
Methods
Animals and surgery
Sprague-Dawley rats weighing between 250 and 300 g were deeply anesthetized by a 2.4-mL/kg intra-abdominal injection of a mixture of 1 ml midazolam (5 mg/mL), 1 mL Hypnorm® (VetaPharma Ltd., Sherburn Enterprise Park, Sherburn-in-Elmet, Leeds, U.K.), and 2 mL of distilled water. All experiments were approved by the local ethics committee. A midline incision was made through the skin and periosteum, and a burr hole 2.7 mm in diameter was drilled with its center 3 mm lateral and 3 mm posterior to the bregma. The rat was thereafter placed in a stereotactic frame and positioned so that the impactor probe was positioned directly above the dura exposed by the burr hole. A lead pellet (Accupell; Crossman, Bloomfield, NY) was accelerated by air pressure from a specially designed accelerator made from a modified air rifle (CNC-Process AB, Hova, Sweden). This projectile then impacted a probe consisting of a metal cylinder with an attached carbon fiber pin (length 30 mm, diameter 2 mm) with a tip radius of 1 mm (Fig. 1B). This probe was fitted into a holder and guided by a narrow tube. Depth of penetration was limited by use of a brass ferrule (Swagelok, Solon, OH), fitted around the carbon pin, that is deformed by the metal cylinder at the end of the impactor probe, ensuring that penetration depth was limited to 5 mm. In the current article we used a pressure level in the accelerator chamber that produces a speed of the impactor probe of about 90 m/sec at the time it starts to enter the brain. The numbers of animals used are listed in Table 1.

The penetration rig. (
RAM, radial arm maze; 5CSRTT, five-choice serial reaction time task; EPM, elevated plus maze.
Tissue procurement and staining procedures
The rats were sacrificed by an overdose of pentobarbital and transcardially perfused with Tyrode's solution, followed by fixative containing 4% formaldehyde in phosphate buffer (pH 7.0; APL; Kungens Kurva, Stockholm, Sweden). The brains were dissected out, post-fixed for 2–3 h at 4°C, rinsed in 0.01 M PBS, and transferred to 0.01 M PBS solution containing 10% sucrose and stored overnight at 4°C, and then cut into coronal sections using a cryostat. The sections were thaw-mounted onto gelatin-coated slides and stored at −20°C until sectioning.
Hematoxylin and eosin (H&E) staining
For general brain pathology, sections were stained in hematoxylin (Histolab, Göteborg, Sweden) for 15 min, and rinsed in water (15 min), followed by staining with eosin (2 min), and thereafter were dehydrated through a series of increasing ethanol concentrations, and finally immersed in xylene for 15 min and cover-slipped with Entellan (Merck, Darmstadt, Germany).
Fluoro-Jade
For detection of the degeneration of cortical neurons. Sections were stained with Fluoro-Jade C (Millipore, Billerica, MA) according to the manufacturer's instructions. Briefly, the slides were immersed in 80% EtOH with 1% NaOH for 5 min, followed by 2 min in 70% EtOH, and 2 min in distilled water, and incubated in 0.06% potassium permanganate solution for 10 min. The slides were subsequently rinsed in water, and transferred to a 0.0001% solution of Fluoro-Jade C in 0.1% acetic acid. The slides were then rinsed in distilled water, air dried, and then cleared in xylene and cover-slipped with Entellan.
Immunohistochemistry
The slides were incubated with primary antibodies overnight at 4°C. The following antibodies were used: anti-Iba1 to label macrophages/microglia (1:400; Wako Pure Chemical Industries, Ltd., Osaka, Japan), and anti-glial fibrillary acidic protein (GFAP; 1:200; Sigma-Aldrich, St. Louis, MO). After three 5-min rinses in 0.01 M PBS, the sections were incubated for 1 h at room temperature with Cy2-secondary antibodies (1:200; Jackson ImmunoResearch, West Grove, PA), and/or Cy3-secondary antibodies (1:400; Jackson ImmunoResearch). The sections were examined in a Nikon E600 microscope (Nikon, Shinjuku, Japan) using appropriate filter settings. Images were captured with a Nikon Digital Sight DS-U1 (5 megapixel) camera, controlled with Nikon EclipseNet software. For image analysis, the pictures were imported into Image J software (National Institutes of Health,
Silver staining
Brains were sectioned in 40-μm sections and processed free-floating using the FD Neurosilver Kit (PK301; FD Neurotechnologies Inc., Baltimore, MD) according to the manufacturer's instructions.
Electron microscopy
For electron microscopic analysis, rats were perfused with Tyrode's solution, followed by 4% glutaraldehyde in 300 mOsm phosphate buffer. The cortex was dissected out, post-fixed, and rinsed. Following this, the tissue was osmicated, dehydrated in acetone, and embedded in Vestopal W. Ultrathin sections were cut on a microtome, and contrasted with uranyl acetate and lead citrate. Finally, the sections were examined in a Philips EM 201 electron microscope.
Determination of brain water content
Following sacrifice, the brains were removed and placed in rat brain blocker (David Kopf Instruments, Tujunga, CA). Using a razor blade, two coronal cuts were made through the brain and underlying structures; one 1 mm posterior to the bregma and one 5 mm posterior to the bregma. All parts of the brain not within these boundaries were discarded. The remaining part was divided along the midline and the left (uninjured) side was discarded. The remaining piece of injured cortex with underlying structures was weighed (wet weight). Thereafter the tissue samples were incubated for 72 h at 75°C, and thereafter weighed (dry weight). The brain water percentage was calculated using the formula: [(wet weight – dry weight)/wet weight] * 100. In the current study, examination was limited to one time point: 72 h post-injury.
Beam-walking test
The beam-walking task (BWT) was used to assess balance and coordination.
The set-up consisted of a rigid 2.5-cm-wide flat beam 1.5 m in length leading to a brightly decorated goal box. The beam was placed 50 cm above the floor and foam cushions were placed underneath the beam. Three days before habituation, the animals were deprived of food until their body weight was reduced to 85% of the initial free-feeding weight (∼3 days). In the habituation phase (2 days) the rat was first placed in the goal box to feed on the reward pellets provided (45 mg, raspberry flavored). Subsequently, progressive beam training was initiated, starting from the beam end closest to the goal box and progressing gradually to the opposite end. At testing, the time taken for the animals to transverse the 1-m length of the beam was recorded with a stopwatch. The animals were finally tested 1 day before surgery, and thereafter at 3 through 7 days after surgery. Animals that were unable to perform the test were designated “immobile,” and no time was entered. However, the animals were kept and tested on the following days.
Elevated plus maze
The elevated plus maze (EPM) is one of the most widely used behavioral paradigms, which presumably measures fear-motivated avoidance behavior. The EPM used in the current study consists of an elevated cross-shaped platform with two open and two closed arms. The apparatus was custom made with powder-coated metal according to specific dimensions for rats: the platform stands 52 cm above the ground; each arm measures 50 cm from the central platform and is 10 cm wide; the side walls of the closed arm measure 30 cm high. A railing along the open arms, measuring 2 cm high, was added in-house to prevent rats from falling off and to increase open arm entries. The experimental procedure was initiated by the placement of the rat on the central platform with its head facing a closed arm. The rat was then allowed to roam the maze without any visual or audio distraction for 5 min. The whole EPM exposure was video recorded, and various behavioral patterns were subsequently counted and timed, including the following: (1) time spent in open arms; (2) time spent in closed arms; (3) number of rearings; and (4) number of central platform crossings. The rats were tested on the EPM at 3 days post-injury.
Radial arm maze
To measure working and reference memory, we used the radial arm maze (RAM). The maze consisted of an octagonal central platform with eight automated sliding guillotine doors giving access to eight radiating arms of equal length (measurements of the maze: width 1690 mm, length 1250 mm, and height 1450 mm). Each arm contains two pairs of photoelectric cells mounted on the proximal and distal end of the arm to differentiate between arm entries and visits. In addition at the end of each arm there is a food site, the contents of which are not visible from the central platform. Prominent extra-maze visual cues are present to allow spatial recognition of arm position. During the habituation phase (2–3 days) the food-restricted rats were familiarized with the maze. All doors were open and food rewards were scattered around the maze to entice the rats to explore. After the rats were allowed to freely explore the maze and had consumed the food rewards at the food disc at the distal arm, the training phase was begun (a span of 10 or more days). For measuring reference memory, the rats were trained to retrieve four food pellets from each selected baited arm only once. To train the rats to do so, they were first placed in the center arena with all doors closed. After 5 sec, all eight doors to the radial arms were opened and the rats were allowed to explore the RAM until the entrance into one arm was detected. Then all doors closed, except to the arm being visited. After the animal returned to the central area, the open door would close. This was followed by a confinement period (5 sec), and then the doors reopened for a new round of choice. This cycle was repeated automatically until the rats visited all four baited arms or after 10 min had elapsed. For each trial, the arm choice, latency to obtain all the pellets (i.e., response latency), and the number of visits to each arm was automatically recorded by the MazeSoft software (Panlab, Barcelona, Spain). After each run, the maze was cleaned with absorbent paper to prevent a bias due to olfactory cues. Over time, the rats would also learn that certain arms were not baited and avoid them accordingly. The animals were trained twice each day until a stable baseline performance (>75% accuracy) was reached. After surgery and exposure to trauma or sham exposure the rats were tested daily, starting at 72 h post-injury, up to 7 days.
Five-choice serial reaction time task
The five-choice serial reaction time task (5CSRTT) was used to examine changes in attention. The 5CSRTT (Med Associates Inc, St. Albans, VT) consists of an operant chamber placed within a sound-attenuating box. Within the chamber, five niches are found on one side, and each of these niches contains a light emitting diode (i.e., for stimulus presentation), and an infrared motion sensor. On the opposite side of the chamber, a food magazine linked to a food pellet dispenser is installed for implementation of positive reinforcement to correct responses. A food restriction protocol (as previously described) was used to ensure motivation on the task. First, the animals were habituated to the context and food pellets, and subsequently the nose-poke and pellet retrieval procedure. This was followed by a training period in which the subjects learned to detect and nose-poke the appropriate niche for food reward with a shortening time window (gradually 60 sec to 15 sec to 5 sec, and then 2 sec) over several sessions. Upon satisfaction of the prescribed performance criteria (>80% correct; less than 20% missed; over 3 consecutive days), at stipulated test parameters (2-sec stimulus duration; 5-sec limited hold), the animals were then subjected to penetration injury and then evaluated from 3 days after injury to 7 days for changes in performance. Animals that were unresponsive were designated “immobile,” and no test values were recorded or assigned for that particular occasion. However, the animals were kept and tested on the following days.
Statistical analysis
Statistical analyses were carried out using GraphPad Prism 5.0 (Graphpad Software, La Jolla, CA). Comparisons between two groups were made using Student's t-test, and multiple group comparisons were performed using one-way analysis of variance (ANOVA), followed by Dunnett's test. Contingency tables (used in the beam walk and 5CSRTT analyses) were analyzed using the chi-square test for trend. Data are presented as mean±standard deviation (SD).
Results
Injury severity
An air pressure of 50 bars (5 MPa) causes the lead pellet to accelerate to a speed of 238±1 m/sec, which in turn causes the probe to accelerate to a speed of 87±3 m/sec, and penetrate the brain to a depth of 4.45±0.18 mm. At this injury severity survival was high (96.7%), and the animals were able to groom and did not display overt symptoms of pain or discomfort. Preliminary histological examination revealed extensive tissue destruction; we therefore chose to limit our analyses to this level of injury.
Histopathology of tissue destruction
The injury caused severe damage to the lateral and medial parietal cortices, corpus callosum, hippocampus, and several parts of the posterior thalamus (Fig. 2A–D). At 14 days post-injury, (the longest survival time studied) a large cavity was visible (Fig. 2D). Erythrocytes were visible in the brain parenchyma at 24 and 72 h post-injury (Fig. 2A, B, G, and H). At 24 h rouleaux stacks were observed in the cortex (Fig. 2F). Occasional erythrocytes were observed among numerous macrophage-like cells at 72 hours post-injury (Fig. 2I). At 14 days, a dense rim of cells was observed surrounding the cavity (Fig. 2J). The dorsal striatum was also affected, as evident by a lighter staining of the tissue on the lesioned side (Fig. 2K–M). Also, severe enlargement of the lateral ventricle on the ipsilateral side was noted (Fig. 2K).

General pathology. (
Silver staining could not be detected at 24 or 72 h (data not shown), but was very pronounced in white matter tracts such as the internal capsule and fimbria of the hippocampus at 7 days post-injury (Fig. 3B). High magnification revealed classic end bulbs in the aforementioned areas (Fig. 3D and E). Further, the fornix on the injured side was heavily stained with silver (Fig. 3G) and Fluoro-Jade (Fig. 3I). Degeneration of cortical neurons was assessed by Fluoro-Jade staining. There was a significant increase in the number of labeled neurons at 24 h post-injury (Fig. 4), and although the number remained elevated, it was not significant for the other time points examined (72 h, 7 days, and 14 days post-injury; Fig. 4C).
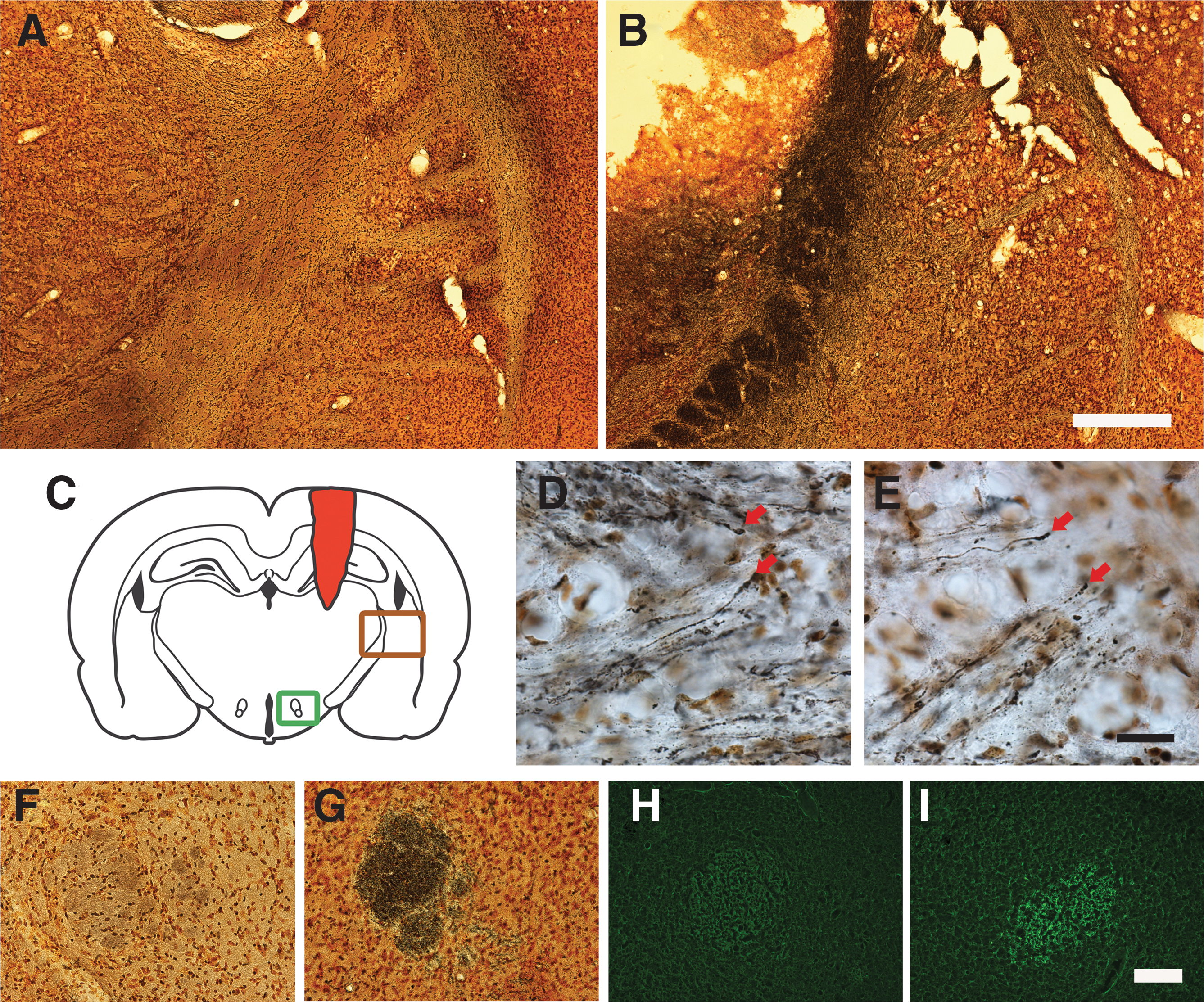
White matter degeneration post-TBI. Silver staining of uninjured (
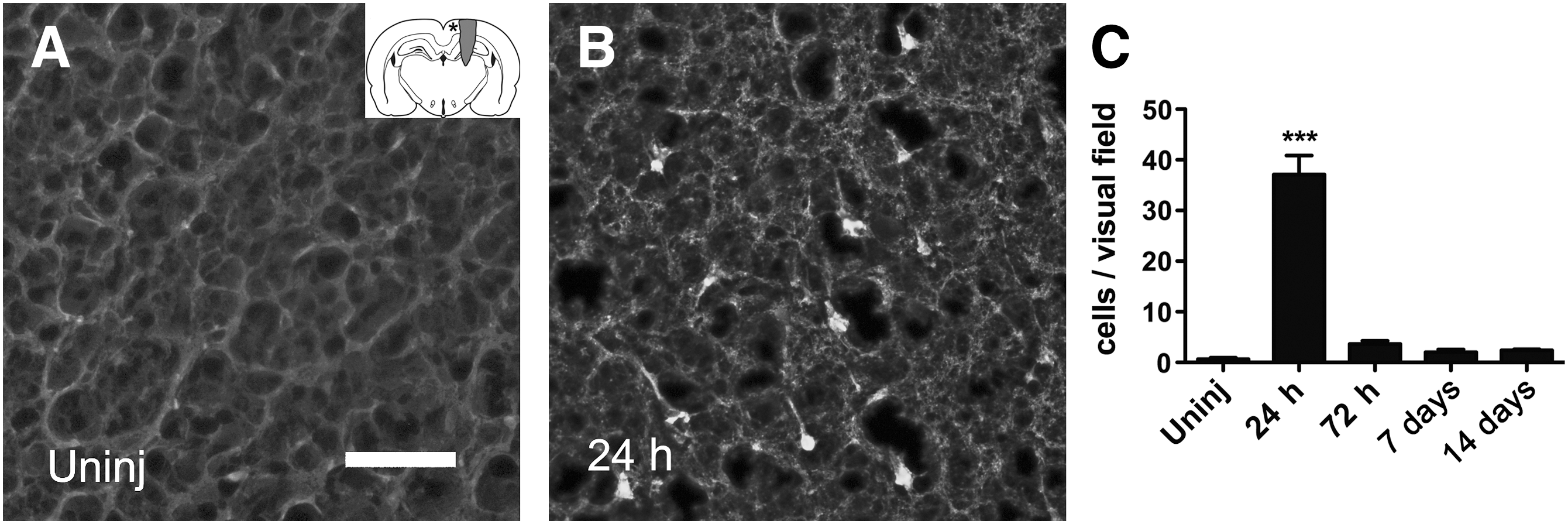
Cortical neuron degeneration. Fluoro-Jade staining in uninjured (
Blood–brain barrier defects, edema, and brain water content
Edema and blood–brain barrier (BBB) leakage was examined using light and electron microscopy and determination of brain water content. BBB defects were analyzed histologically using anti-rat IgG antibodies (Fig. 5A). The IgG staining was confined to the lesion area, and no staining was observed on the contralateral side of the brain or in the ventral hypothalamus on the injured side. The staining intensity was very high at 24 and 72 h post-injury, and thereafter began to diminish. We also examined the injury using electron microscopy of the injured cortex at 7 days after injury. Our results showed pronounced extracellular edema surrounding the blood vessels (Fig. 5B). Finally, quantification of brain water content was determined by examining the dry:wet ratio. The percentage of water in control animals was 78.03±0.27% (n=5), whereas in the injured animals the water content was 79.71±0.23% (n=5), indicating the accumulation of water in the tissue (Fig. 5C).
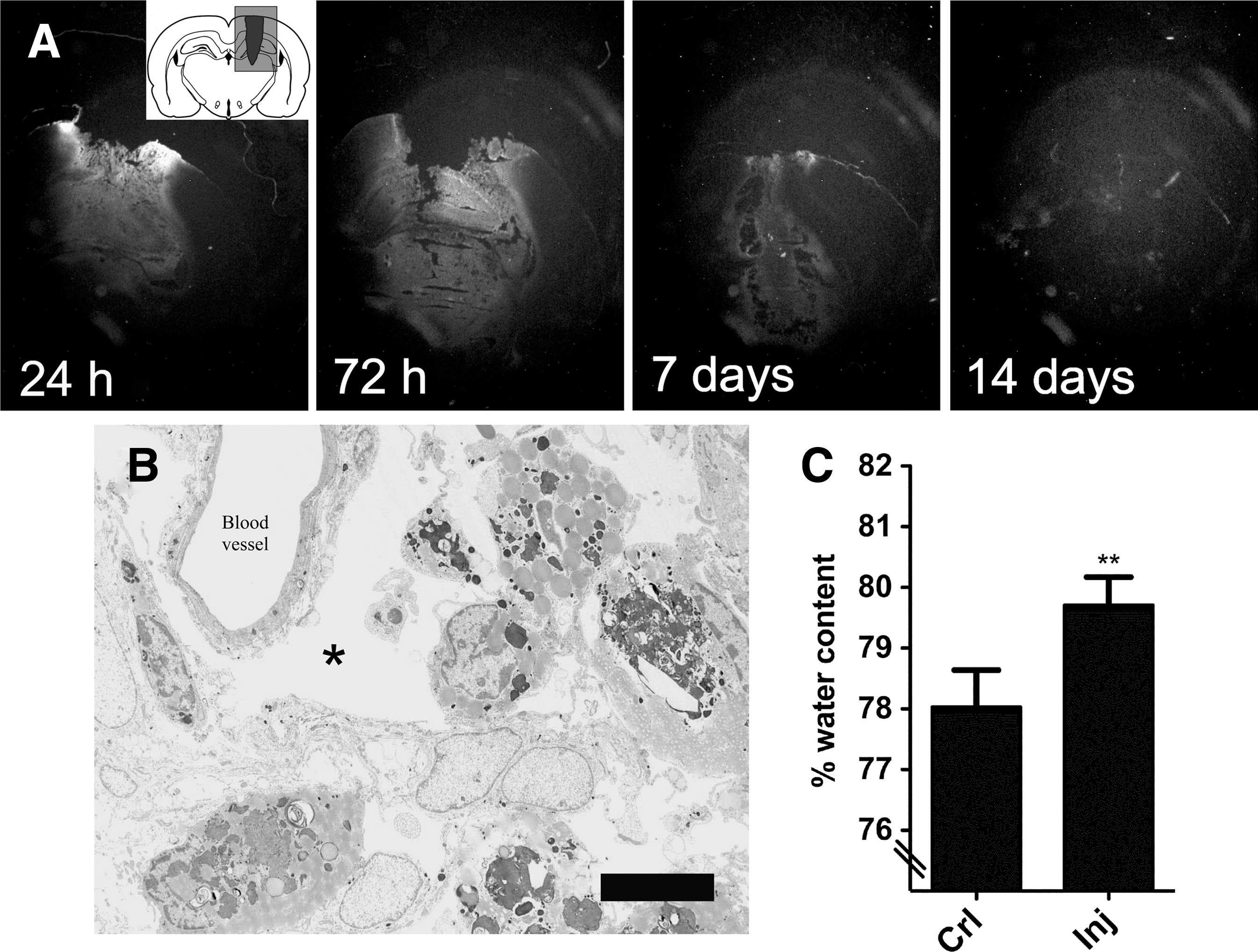
Blood–brain barrier defects after TBI. (
Reactive gliosis
Reactive astrocytes (as indicated by GFAP staining) were confined to a region surrounding the lesion core, intermingled with Iba1-positive cells (macrophages/microglia; Fig. 6B). In contrast, Iba1-positive cells were also present in the lesion core (Fig. 6B). Cortical gliosis was quantified in a region approximately 3 mm medial to the injury by image densitometry of GFAP and Iba1 staining intensity. A slight increase in GFAP staining could be detected after 24 h, but was not statistically significant, whereas after 72 h, staining was increased further, and remained so up to 14 days, the latest time point studied (Fig. 6E). A similar time course of activation was found for macrophages/microglia (Fig. 6H).
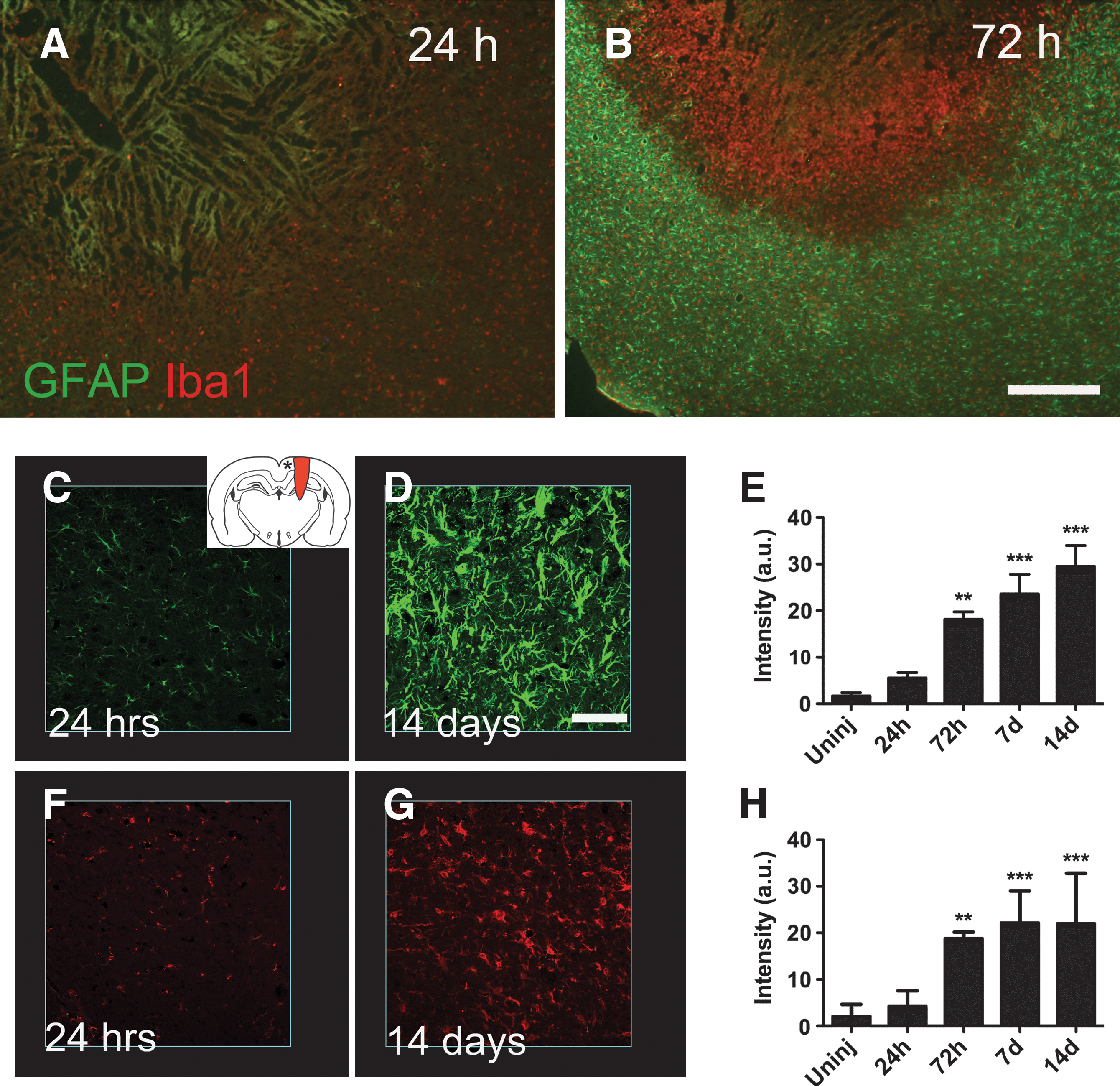
Gliosis post-TBI. GFAP (green) and Iba1 (red) immunoreactivity in horizontal sections at 24 (
Body weight post-injury
After an initial weight loss of 4.4±2.9% (n=11) for the first 24 h injury, injured animals gradually regained weight and were statistically significantly heavier than before injury at 8 days, compared to nine 9 days for control animals (Fig. 7). At the end of the experiment (15 days post-injury), injured rats had gained 5.8±4.2% compared to 3.7±3.0% (n=11) for the control group (Fig. 7). Data are expressed as mean±SD (n=11 in each group; *p<0.05, **p<0.01, ***p<0.001 by repeated-measures ANOVA followed by Dunnett's test, compared to before injury T 0).
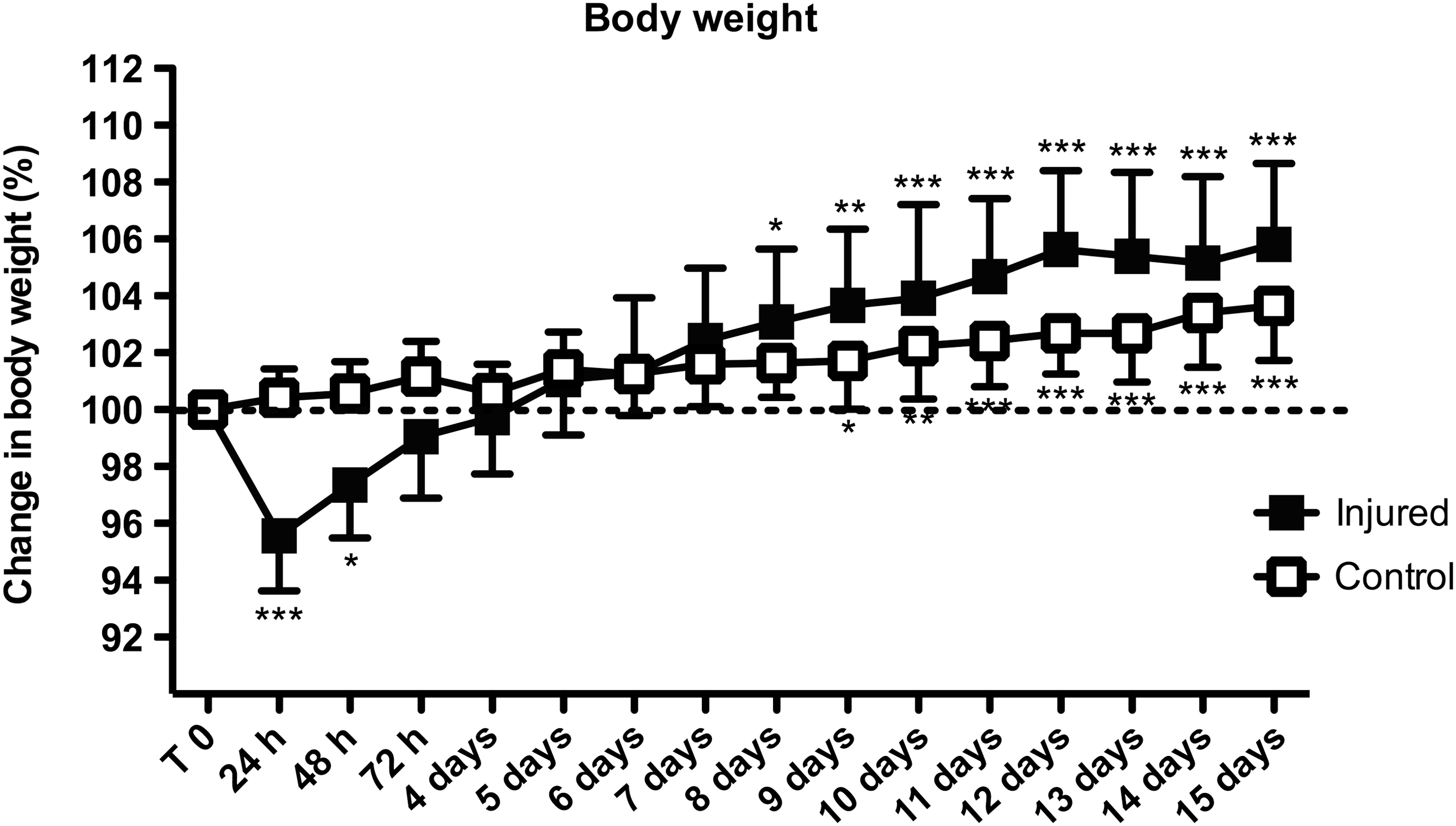
Weight gain post-TBI. After an initial weight drop, injured animals gained weight rapidly, and reached a significant increase in body weight (compared to pre-injury weight) earlier than control animals. Data are expressed as mean±standard deviation; n=11 in each group; *p<0.05, **p<0.01, ***p<0.001 by repeated-measures analysis of variance, followed by Dunnett's test, compared to uninjured T 0; TBI, traumatic brain injury).
Behavioral outcome
In the present study, we investigated the functional outcome of pTBI by the use of well-established models of memory (radial arm maze), anxiety (elevated plus maze), sensorimotor function (beam walking test), and attention (five-choice serial reaction time test).
Beam walk
In the beam walk test; initial trials were performed 3 days post-injury. At this time the majority (11 of 15) of injured animals were unable to balance or move on the beam (Fig. 8B). This gradually declined such that on the fourth day 9 of 15 were unable to move, and on the fifth day only 3 of 15 showed this impediment. On the sixth day, two animals that were able to do the test at 5 days failed to perform, but on the final day of testing (7 days post-injury) only one animal still had impairments. There was a significant (p<0.001) influence of time as determined by chi-square testing for trend in the injured animals (Fig. 8B). The latency increased after injury from 2.3±1.1 sec to 8.4±4.3 sec at 3 days, and 10.6±8.1 sec at 5 days. After 7 days the latency for the injured animals was 4.3±1.8 sec, compared to 1.58±0.8 sec for the sham animals (Fig. 8C).
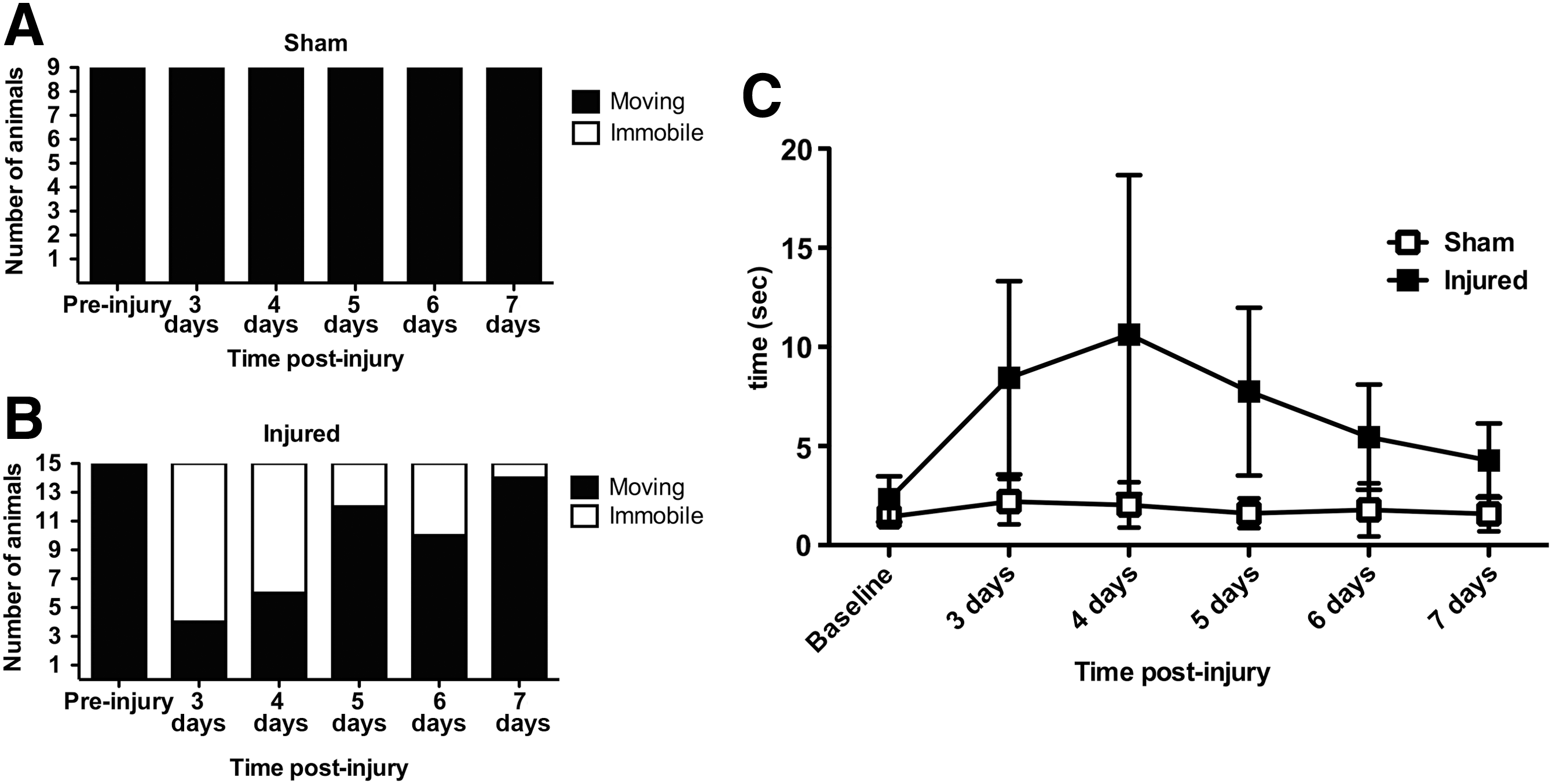
Time course analysis of performance in the beam walk task. All animals in the control group were able to complete the task (
Elevated plus maze
In the elevated plus maze, injured rats did show a reduced time spent in closed arms (p<0.05 by Student's t-test; n=9 sham, n=19 injured), whereas time spent in open arms and at the center platform increased slightly (Fig. 9A). However, the number of total arm entries (Fig. 9B), as well as both rearing activity and the number of crossings, was lower (Fig. 9C), all pointing towards a general reduction in activity.

Lower level of anxiety behavior in injured animals. Injured animals spent less time in the closed arms of the elevated plus maze compared to control animals (
Radial arm maze
Visit response latency (the time taken to correctly find all four baited arms) did not change significantly after injury (data not shown), and neither did the number of working memory errors (Fig. 10A). However, we did observe a significant (p<0.001 by repeated-measures ANOVA followed by Dunnett's test compared to pre-injury baseline; n=10) increase in reference memory errors that persisted throughout the testing period of 7 days (Fig. 10B).
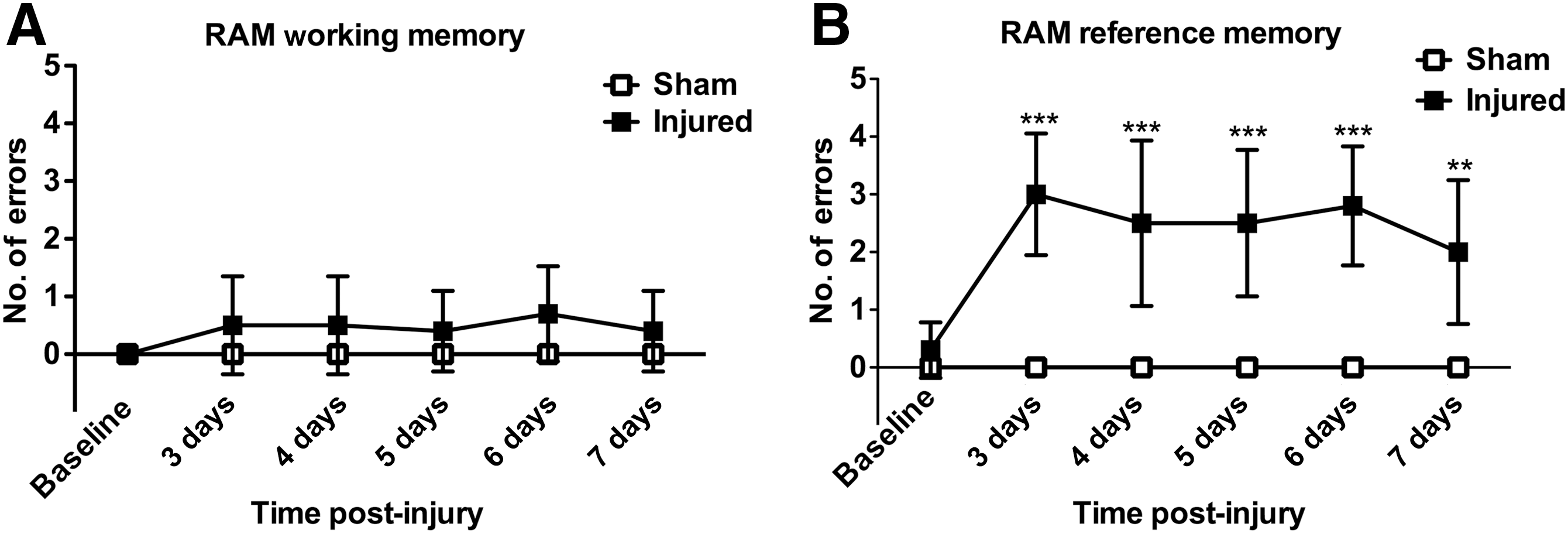
Reference memory deficit post-TBI. No significant difference in the number of errors in the working memory paradigm could be detected (
Five-choice serial reaction time test
At 3 and 4 days post-injury, 4 of 10 injured animals were unable to complete the task (Fig. 11B), whereas all animals in the control group were able to do so (Fig. 11A). This gradually changed so that on the fifth day the corresponding number was 3 of 10, and on the final day of testing (7 days post-injury) 1 animal was still unable to respond (Fig. 11B). However, these results were not significant in this series of experiment (p=0.075 by chi-square testing for trend). There was a transient reduction in the number of correct choices in the injured group, but this was followed by a gradual increase (Fig. 11C). Further, we did not observe any clear trend concerning choice latencies or premature responses (data not shown).

Transient deficiency in the five-choice serial reaction time task (5CSRTT) post-injury. (
Discussion
A new neurotrauma model
Although it has been argued that further refinement or development of new animal models for TBI is unnecessary, and that research instead should be focused on refining and facilitating comparison between existing models (Morales et al., 2005). We and others (e.g., Cernak, 2005; Statler et al., 2001) feel that penetration injury is not accurately modeled in current standardized TBI models, and that a new model is therefore justified. As mentioned in the introduction, rodent models for penetrating TBI are scarce. One of the few models currently in use was developed to primarily recreate the temporary cavity formed around a penetrating bullet injury (Williams et al., 2005). The force in this model is directed perpendicular to the injury tract, and is therefore less well suited to model entry wounds and their effects on meninges, blood vessels and the skull. Since a speed of 60 m/sec is sufficient for a projectile to break bone (Lee et al., 1997; Ordog et al., 1984), the 90 m/sec we were able to achieve in our model led us to argue that our work has a reasonable resemblance to the clinical setting with regard to low-velocity pTBI. In addition to the velocity, the shape of the projectile and its behavior upon impact are of great importance for injury severity (Fackler, 1998; Kluger, 2003; Ordog et al., 1984). The relationship between velocity and the shape of the missile can be studied in our model, but not as easily in the PBBI model.
An important factor contributing to the severity of pTBI is the presence of skull fragments in the wound (Karger, 1995; Solmaz et al., 2009). Although the kinetic energy in our model is sufficient to penetrate an intact skull, thus making it possible to better mimic the clinical scenario, in order to increase reproducibility of the trauma, we did not perform this in the present set of experiments. Of note, recent experiments suggest that insults to bone tissue such as the skull (e.g., craniotomy; Cole et al., 2011), as well as a peripheral fracture in combined models (TBI plus tibial fracture; Maegele et al., 2007) both affect the production of inflammatory cytokines that may contribute to the progression of TBI, making this a highly relevant field for future experiments.
Neuronal and axonal injury
We observed a progressive expansion of the injured area over the observation period, at the end of which a large cavity was observed. This progressive nature of the injury will be examined in future studies and could form a platform for the investigation of novel neuroprotective treatments. In addition to the H&E staining we also examined neuronal injury/degeneration by use of Fluoro-Jade and silver staining. The advantage of using a combination of these techniques, whereby the full extent of injury can be captured, has been shown by Hall and co-workers (Hall et al., 2008). In the present study, Fluoro-Jade staining of neurons in the cerebral cortex was pronounced at 24 h (the earliest time point studied), and rapidly declined thereafter. This time course is in line with previously published results using the PBBI model (Williams et al., 2006b). This could potentially be a signature of the pTBI type of injury, since after CCI Fluoro-Jade staining seems to persist beyond 24 h (Hall et al., 2008; Yang et al., 2010). In the CCI model axonal injury is detected (by silver staining) as early as 6 h post-exposure in the cortex, but deeper white matter structures such as the internal capsule are unaffected (Hall et al., 2008), in contrast to the findings of our model and those of Williams and colleagues (2005). However, assessing the injury zone using silver staining has its limitations. As pointed out by Hall and associates (2008), silver staining tends to stain the entire degenerating axon (or dendrite), which makes it difficult to identify the sub-cellular lesion site (unless retraction bulbs are identifiable). One way to circumvent this problem could be by including immunohistochemical staining of accumulated β-amyloid precursor protein (β-APP). We have not included this analysis in the current study, but have previously reported β-APP staining in the cortex as early as 3 h post-injury in our pTBI model (Risling et al., 2011).
Blood–brain barrier defects and hemorrhage
A BBB defect was noted in the lesion areas by IgG immunohistochemistry as well as by an increase in water content (as determined by the dry:wet ratio) after injury. Moreover, using electron microscopy, we detected an extracellular perivascular edema, in contrast to the intracellular edema observed after rotation-induced TBI (Risling et al., 2011), or CCI (Unterberg et al., 2004). Edema is a common feature after pTBI (Carey et al., 1990), and although the molecular mechanisms underlying this phenomenon are not completely understood, several studies implicate an increase in the expression or modulation of aquaporin-4 as a key event (Gunnarson et al., 2008; Manley et al., 2000; Neal et al., 2007). After injury, intracerebral hemorrhage was pronounced, with the presence of erythrocytes at 24 and 72 h. Hemorrhage is a serious consequence of TBI, in that it raises intracranial pressure (ICP) and causes the leakage of hemoglobin and iron from lysed erythrocytes, which has been linked to unfavorable outcomes due to oxidative injury (Davalos et al., 2000; Wu et al., 2002; Xi et al., 2006). Future studies will be needed determine the exact relationship between hemorrhage, edema, and ICP after pTBI (Carey et al., 1989; Xi et al., 2006). Finally, BBB disruption causes the influx of several blood-borne molecules not normally present in the parenchyma that affect the progression of injury and recovery. Leakage of blood-borne fibrinogen into the central nervous system (CNS) is known to activate microglia (Adams et al., 2007), interact with neuronal growth cones to inhibit outgrowth (Schachtrup et al., 2007), and to stimulate the production of growth inhibitory neurocan from reactive astrocytes (Schachtrup et al., 2010). In contrast, leakage of plasma-derived fibronectin into the brain after TBI has a neuroprotective effect (Tate et al., 2007).
Gliosis
In the current study, we examined the time course of glial activation post-pTBI.
A commonly used marker of activated astrocytes is an upregulation of GFAP expression. We previously reported an increase in GFAP activity surrounding the lesion using in situ hybridization histochemistry (Risling et al., 2011). In this study we used semi-quantitative immunohistochemistry to describe the time course of astrocyte activation in the injured cortex. A statistically significant increase was observed at 72 h and persisted up to 2 weeks (the latest time point studied).
In the PBBI model, Williams and co-workers described a more rapid activation of astrocytes, but also with a shorter duration (Williams et al., 2007). The reason for this difference is currently unclear, but the methodological differences of inducing the injury could be a main reason. Astrocytes are known to carry out several functions after CNS injury that are of relevance to TBI, including removal of excessive amounts of extracellular glutamate (Rothstein et al., 1996), resealing the BBB (Bush et al., 1999), and facilitating axonal regeneration, under some circumstances (Frisen et al., 1995; Tom et al., 2004). However, reactive astrocytes also contribute to edema formation (Sofroniew and Vinters, 2010), and form fibrotic scar tissue that inhibits axonal regrowth (Asher et al., 2000; Fawcett, 1994) and produces proinflammatory cytokines (Eddleston and Mucke, 1993).
In addition to astrocytes, we also analyzed the microglia/macrophage response by use of Iba1 immunohistochemistry. In our model, cortical microglia remained activated up to 14 days post-injury, longer than the reactive response described in the PBBI model (Williams et al., 2007). However, this could be due to differences in how penetration was achieved and/or in the different marker used (Iba1 in our study versus OX-18). Similarly to astrocytes, activated microglia can perform both protective and harmful tasks, such as scavenging, phagocytosis, antigen presentation, and synaptic stripping, as well as secretion of both proinflammatory and anti-inflammatory mediators (Cullheim and Thams, 2007; Streit et al., 1999). Currently, no mutually exclusive marker exists to differentiate activated microglia from blood-borne macrophages. The BBB defects observed in our study make it possible that the Iba1-postitive profiles observed in and around the lesion consist of a mixture of these two types of cells. Although recent genetic tracing experiments have revealed that their embryonic origin is different (Ginhoux et al., 2010), the contribution of these two cell populations to injury and repair after TBI is currently unknown. Given the large amount of detrimental and beneficial processes mediated by astrocytes and macrophages/microglia, elucidation of these processes might be a key to understanding the pathological response of pTBI.
Body weight
Initial loss of body weight is observed after CCI (Dixon et al., 1991), and blast-induced TBI (Cernak et al., 2011), with a recovery rate inversely related to the magnitude of injury. Although few studies have specifically addressed the mechanisms at play, decompressive craniotomy has been shown to accelerate regain of body weight after CCI (Zweckberger et al., 2003), as has treatment with progesterone of male rats after CCI (Djebaili et al., 2004). In contrast, the PBBI model does not cause weight loss (Williams et al., 2005). In our model, weight loss was significant at 24 and 48 h post-injury, and thereafter reached a statistically significant weight gain (compared to pre-injury weight) faster than control rats. In the present study, survival was limited to 2 weeks, and it is not clear how age, sex, or the genetic make-up (breed) of the injured animals affected the rate of weight loss or gain in our model. Brain-injured patients often experience metabolic disturbances (Kaufman et al., 1987; Wilson and Tyburski, 1998), and problems with weight regulation in the chronic phase can in some cases manifest itself as obesity (Castano and Capdevila, 2010; Patradoon-Ho et al., 2005). Possible explanations could be either direct injury to the thalamic circuits regulating satiety, eating behavior, or pituitary dysfunction, or growth hormone deficiency (Norwood et al., 2010; Popovic, 2005). Given the complex interaction between neuronal, endocrine, and social influences on eating, resolving this issue will require extensive research efforts in the years to come.
Behavior analysis
Sensory and motor coordination were assessed with the beam walk test (Hamm et al., 1994). Injured rats displayed a clear impairment in this task, with a gradual return of function to levels nearly comparable to pre-injury levels after 7 days. Spontaneous recovery is commonly seen after experimental TBI (Hamm et al., 1994; Williams et al., 2005), but which of the several possible mechanisms in our lesion paradigm are most important remains to be determined. In the elevated plus maze, we observed that injured rats spent less time in closed arms compared to control rats. Similar results have been presented for weight-drop injury (Pandey et al., 2009), but these results are in contrast to the increased anxiety-like behavior observed by rats (in the EPM) in a study of the long-term effects of fluid percussion injury (Jones et al., 2008). However, in the current study, the number of total arm entries, as well as both rearing activity and the number of crossings was lower, all pointing towards a general reduction in activity. Given that much of the motor function returns after 7 days (Fig. 8), future exploration of the effects of anxiety will focus on long-term effects. Concerning memory function, the rats used in this study showed deficits in reference memory, but not in working memory after TBI, in contrast to what has been reported after fluid percussion injury (Lyeth et al., 1990). A possible explanation could be the extensive damage to the thalamus seen in our model (Fig. 2), since thalamic injury can cause reference memory deficits (Mitchell and Dalrymple-Alford, 2005). Finally, the attention deficit observed could possibly be due to lesions of the prefrontal cortex, since this affects performance in the 5CSRTT (Maddux et al., 2007), whereas lesions of the parietal cortex do not (Muir et al., 1996). However, performance in the 5CSRTT can also be modulated by subcortical regions, since bilateral lesions of the habenula affect performance (Lecourtier and Kelly, 2005).
Conclusion
In conclusion, we have developed a novel method for pTBI that permits non-fatal rapid penetration injury to the rodent brain. We have characterized the injury and described the time course of several pathological reactions, such as cavitation, hemorrhage, neurodegeneration, and gliosis. In addition to the histological findings, we observed transient changes in most behavior tests, but the most striking feature was a persistent deficiency of reference memory, likely due to injuries of the cortex and the hippocampus. In our model, the lesion site can easily be changed to other parts of the brain, which could be of interest since even small changes in location can affect lesion development and outcome (Vink et al., 2001). In its present design, the rig only allows us to make injuries at a straight angle in the coronal plane. Thus we are currently not able to replicate all the variations of injury trajectories (e.g., bilateral hemispheric and unilateral caudal) described by Tortella and co-workers using the PBBI model (Williams et al., 2006a). Finally, our model can be used to address external factors of importance on the outcome of pTBI, such as the speed of penetration or the shape of the penetrating object (akin to the situation after CCI; Pleasant et al., 2011), or internal factors such as the animal's age, sex, or the genetic regulation of TBI secondary injury components such as the inflammatory reaction (Bellander et al., 2010).
Footnotes
Acknowledgments
We thank Mrs. Maria Angeria for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.