
Abstract
Mild traumatic brain injury (mTBI) accounts for the majority of head trauma cases. Despite some lasting cognitive, emotional, and behavioral deficits, there are frequently no overt morphological defects, suggesting that changes may result from alterations in the physiology of individual neurons. We investigated hippocampal neural activity in rats during a working memory task to determine the effect of mTBI on cellular physiology. Male Sprague-Dawley rats (300–350 g) underwent mTBI via lateral fluid percussion (1.5 atm), and were compared with sham-operated rats. The rats then underwent bilateral implantation of electrodes into the CA1 and CA3 hippocampal subfields and were trained to perform in a delayed nonmatch-to-place swim T-maze. Single-neuron activity was analyzed during task performance 30–90 days after trauma. There were no histological differences between control and mTBI rats. Stereological analysis demonstrated no neuronal loss. Nevertheless, rats subjected to mTBI demonstrated significantly poorer performance on the task with increasing delay. Examination of single-neuron spiking activity revealed no significant difference in firing rates or spike characteristics, but rats exposed to mTBI were found to have significantly fewer cells with activity spatiotemporally correlated with location in the maze (“task-specific cells,” p<0.05 by Fisher's exact test). Memory deficits, including disorganized patterns of hippocampal neural activity after mTBI, were seen in rats. Because it is seen in the absence of clear morphological defects, these data suggest that functional impairment after mTBI may result from alterations in the activity of individual neurons.
Introduction
A
Previous studies have identified significant electrophysiological changes in the hippocampus after moderate to severe TBI (D'Ambrosio et al., 1998; Witgen et al., 2005). However, mTBI is likely to be associated with more subtle functional abnormalities because obvious histological changes are not seen. It has long been known that a subset of pyramidal cells in the hippocampus and entorhinal cortex display task-specific firing attributes, and multiple investigators have demonstrated that firing patterns are frequently highly tuned to spatial and performance characteristics among rats engaged in many different types of conditioning paradigms (Hampson and Deadwyler, 1996; Hampson et al., 1993,1999; Hollup et al., 2001; Wilson and McNaughton, 1993). This activity requires integration among multiple areas in the mesial temporal lobe, and lesion studies have demonstrated that damage to tracts connecting these structures can lead both to reductions in the information encoded by such cells and to reduced performance on memory tasks (Ahmed and Mehta, 2009). It is possible that disruption of these cells or the pathways connecting them may underlie some of the functional deficits seen after mTBI. In the present study, we use single-unit recording of hippocampal pyramidal cells to determine the effect of mTBI on single-unit cellular activity and task performance to determine whether impaired neural function could be associated with animals showing memory deficits.
Methods
Subjects
Six male Sprague-Dawley rats (Harlan Laboratories, Inc., Indianapolis, IN) 2–3 months of age and weighing 300–350 g were used. The animals were individually housed in vivariums, maintained at 22°C on a 12-h light/dark cycle, and given ad libitum access to food and water. All procedures followed the guidelines established in the Guide for the Care and Use of Laboratory Animals (U.S. Department of Health and Human Services), and were approved by the Case Western Reserve University Institutional Animal Care and Use Committee.
Injury
Animals were randomly assigned to either the mTBI (n=3) or sham (n=3) group. Under isoflurane anesthesia, a 4.8-mm-diameter lateral burr hole was made over the right parietal cortex, midway between the coronal and lambdoid sutures, and a Luer-Loc syringe hub was secured on the skull at the site of the burr hole with cyanoacrylate and dental acrylic. Twenty-four hours after surgical preparation for lateral fluid percussion injury (LFPI), each animal was connected to the injury device, and those assigned to the injured group were subjected to mTBI at 1.4–1.6 atm (Dixon et al., 1987).
Electrode implantation
Electrode implantation surgery took place 2–9 days post-injury and was performed under isoflurane anesthesia with pulse oximetry monitoring. Sixteen-channel Teflon-coated stainless steel microelectrode arrays (Neurolinc, Sandisfield, MA) were stereotactically implanted bilaterally into the hippocampus. Each array consists of two rows of eight electrodes 200 μm apart, separated by 800 μm, with one row 1.2 mm shorter than the other. The following coordinates were used: M/L±2.9 mm, A/P −3.00 mm, D/V −3.2 to −3.5 mm, to the bregma (Paxinos and Watson, 2009), rotated 30° from the midline. This target places the longer (lateral) row of electrodes into CA3, and the shorter row into CA1 (Hampson et al., 1999). Skull screws were used to provide stability for the arrays. Intraoperative physiological monitoring was performed to monitor spontaneous unit activity in order to verify placement in the appropriate hippocampal cell layers, after which the array was fixed permanently to the skull with dental cement.
At the conclusion of the experiment, electrode location was verified by passing 15 μA for 15 sec through each wire to produce iron deposition, and the rats were subsequently transcardially perfused using 4% paraformaldehyde and 3% potassium ferrocyanide, producing Prussian blue at the location of the tip of the electrode. Anatomic verification of the position of the electrodes in CA1 and CA3 was confirmed by microscopic analysis of tissue counter-stained with neutral red.
Stereology
To determine whether cell loss occurred as a result of mTBI, unbiased stereology was performed on the hippocampal CA3 subfield ipsilateral to injury as described previously (West et al., 1991). Cell counts were performed using Stereologer software, 2nd edition. Every sixth 50-μm coronal hippocampal section was used, and 115–180 neurons over 8–10 sections were counted, corresponding to A/P −2.52 mm through −4.44 mm relative to the bregma (Paxinos and Watson, 2007). The sampling areas were outlined under a 2.5×objective, and cell counting was done under a 100×oil-immersion lens. Neurons were identified by their distinct morphological features; only those cells with a clearly defined nucleolus within a defined nuclear membrane were included in the counts.
Cognitive assay
The delayed nonmatch-to-place (DNMP) swim T-maze was constructed and performed as previously described (Fig. 1A; Whiting and Hamm, 2006). Briefly, the T-maze apparatus was made of clear acrylic glass, and consists of a lane 10 cm wide, with a 110-cm stem and two 55-cm arms. A 10×10-cm removable platform was located at the end of each arm, and a removable divider was used to block entry into one of the arms. Each trial consisted of two phases separated by a variable delay: (1) a sample phase, in which the rat is forced to swim in one direction chosen in a pseudorandom fashion, followed by (2) a nonmatch phase, in which the rat was free to choose either side, but must swim into the opposite arm from the sample phase to find the platform and avoid a 15-sec forced swimming period. Animals started training 14 days after electrode implantation until able to perform the task with fewer than 20% errors. At this point, they were required to perform the task with increasing delay (15, 30, 60, and 120 sec).
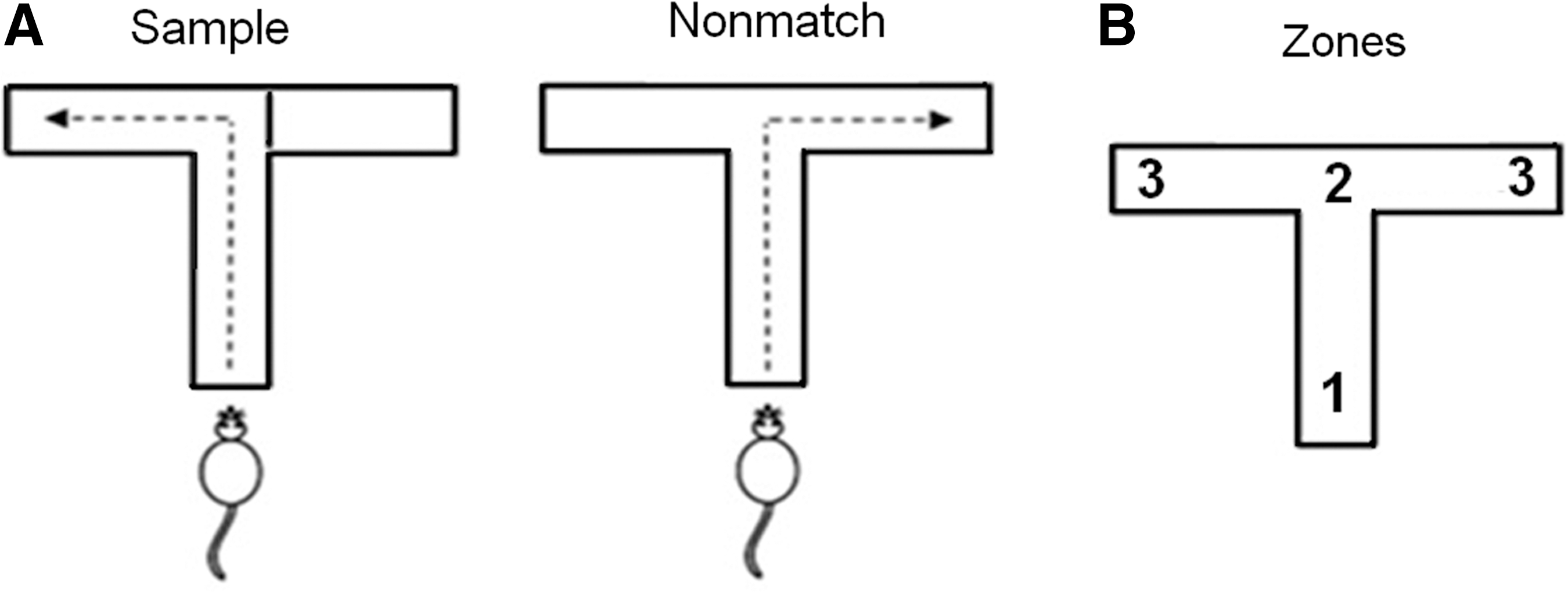
The delayed nonmatch-to-place swim T-maze. (
Electrophysiological recording
Collection of data occurred starting at 30 days after mTBI. A headstage and cable system with a commutator was attached to the electrodes to allow electrophysiological assessment during the final day of training and during all subsequent experiments. A video tracking system (EthoVision; Noldus Inc., Centerville, VA) was used to track the animal during the task, and extracellular action potentials were continuously recorded from the array using a commercial platform (Plexon Inc., Dallas TX). To identify single-unit activity, signals from the electrodes were amplified (5000–10,000×), high-pass filtered (200 Hz), and digitized at 40 kHz. Complex spike discharges characteristic of CA1 and CA3 pyramidal cells were identified using spontaneous rate, waveform, and characteristic phasic bursts as defined on autocorrelelogram and interspike interval histograms (Hampson et al., 1993). Theta cells were excluded from the analysis. SortClient (Plexon Inc.) was used for real-time multiple-unit discrimination using principal component analysis for cluster cutting, with subsequent optimization using Offline Sorter (Plexon Inc.).
Data analysis
NeuroExplorer (Nex Technologies, Littleton, MA) was used to determine spatiotemporal correlations between single-unit spike rates and animal location in the maze. To do this, the point at which the animal passed the decision point at the top of the T was determined, and three 3-sec epochs were defined centered on this point. Assuming unidirectional swimming at a constant speed, this procedure establishes regions of the T-maze corresponding to stem, turning point, and arm, each accounting for one-third of the total path (Fig. 1B). Peri-event histograms and rasters were obtained for each pyramidal cell based on the time stamp of the decision point, and the average firing rate in each epoch was determined. This value was compared with the Poisson spike frequency distribution calculated across the entire experiment. Any unit that consistently fired at a rate higher than the 99% confidence interval for that distribution during one of the three epochs of the maze (entry, turning point, or platform) for a certain condition (sample right, sample left, nonmatch right, or nonmatch left) was defined as a cell with task-specific firing characteristics. These data were collected using a single test day with no delay to establish injury-related differences in hippocampal activity. Subsequently, we examined swim T-maze performance with the introduction of increasing delays. We correlated the behavioral data from the delay trials with the initial observed changes in neuronal specificity.
Statistical analysis
Statistical analyses were performed using the Statistical Package for the Social Sciences (SPSS, Inc., Chicago, IL). Two-tailed Student's t-test and analysis of variance (ANOVA) was used for parametric data, and Fisher's exact test was used for categorical data. A p value of 0.05 or less was defined as statistically significant.
Results
Injury
The average LFPI severity was 1.5 atm (range 1.4–1.6 atm). Average return of the righting reflex was 296 sec for the injured animals, and 90 sec for the sham animals.
Behavioral assessment
A two-way ANOVA revealed that the rate of initial learning during training was comparable between mTBI and sham groups [F (1,4)=0.872, p=0.403; Fig. 2A]. Under the experimental conditions, when there was no delay (0 sec) between sample and nonmatch phases, mTBI and sham animals performed similarly in the swim T-maze task. However, when a delay (15, 30, 60, or 120 sec) was added, the mTBI group made significantly more incorrect decisions than the sham group (p<0.05 by Student's t-test; Fig. 2B). Due to the small sample size, we combined the four delay periods for statistical analysis of the T-maze task performance. Figure 3 shows the average time for each rat at water entry and arrival at the platform, with 0 sec being the point at which the turn was made (either to the right or left). Latency to reach the goal was averaged for all correct trials used in the electrophysiological analysis. These data showed no significant difference in swim times between the sham (M=6.48±SEM=0.236) and mTBI (M=5.95±SEM=0.257) groups [t(4)=1.510, p=0.206). On average, water entry occurred 4.25 sec prior to the decision point (range 3.7–4.7 sec), and arrival at the platform occurred 2.0 sec after the decision point (range 1.7–2.6 sec). As a result, the 3-sec epochs centered on the decision point correspond roughly to entry into the water, making the turn, and having arrived on the platform (Fig. 1B).
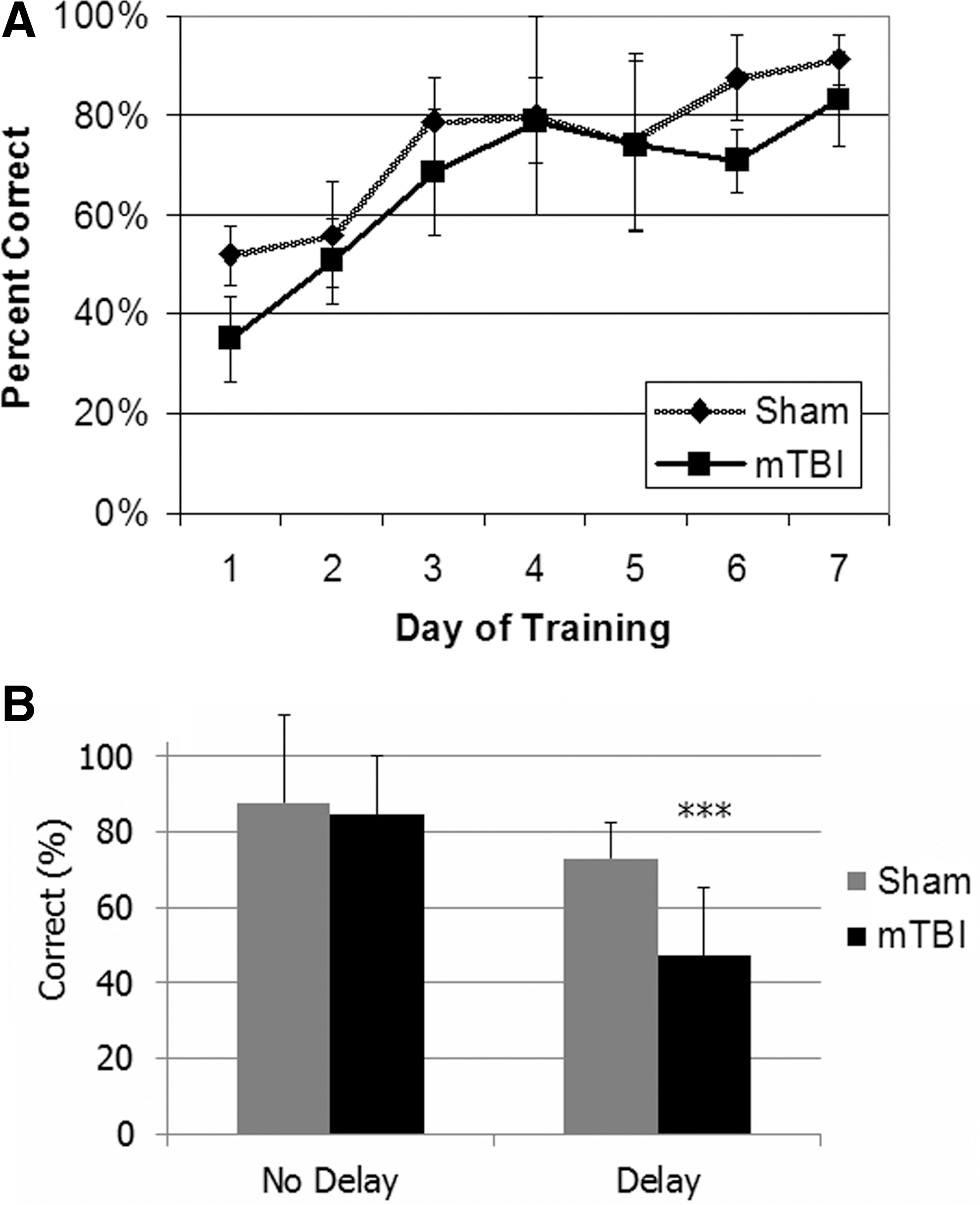
Performance in the delayed nonmatch-to-place maze. (
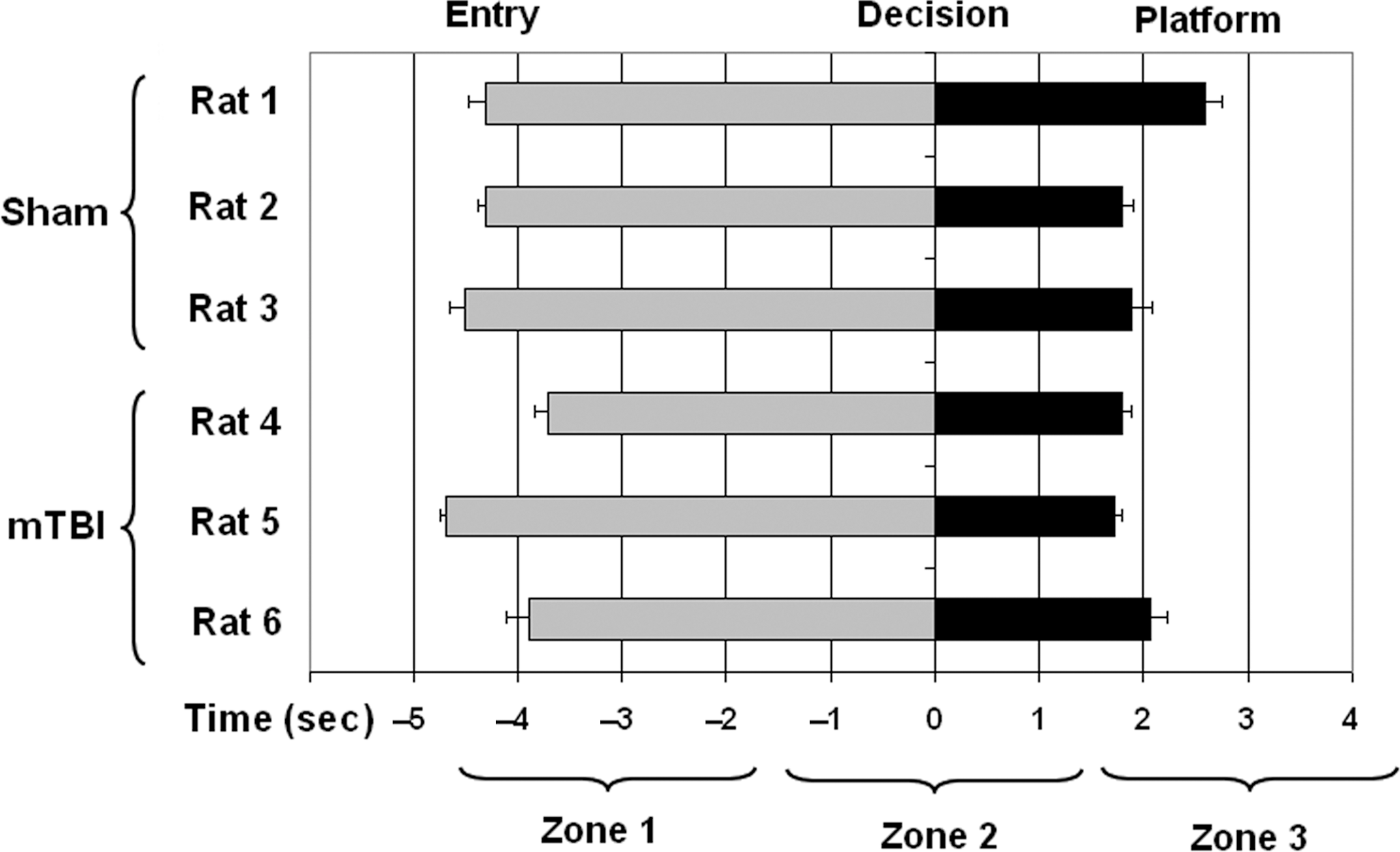
Mean times for entry into the maze and arrival at the platform. We determined that using time rather than place to identify task-specific cells was appropriate by first establishing that all the animals swam at similar rates of speed during task performance on the testing day. There was no significant difference in latency to reach the goal platform between groups (p>0.05). These data include all correct trials for the sham (125) and mild traumatic brain injury (mTBI) (94) groups used in the electrophysiological analysis. Zero seconds corresponds to the point at which the rat turns left or right, so water entry occurs at a negative time point, and arrival at the platform at a positive time point. Rats 1–3 are sham; rats 4–6 are mTBI. Error bars represent within-subject standard error of the mean for latency to reach the goal platform.
Single-unit electrophysiology
We recorded the activity of multiple single units in the CA1 and CA3 subfields of both hippocampi in each rat (Fig. 4A). Autocorrelelograms and interspike interval histograms demonstrated complex spike characteristics that were not significantly different between groups (Fig. 4B and C). There was no difference in average firing frequency (Fig. 4D). A total of 40 pyramidal cells were identified in the sham group, compared with 31 in the mTBI group, but this difference was not statistically significant. A majority of cells in both groups demonstrated no change in firing rate based on position in the maze. In every rat tested, a subset of cells consistently demonstrated a change in firing rate in one or more of the zones of the maze during one or more trial types. The patterns were diverse and sometimes complex, with some cells demonstrating increased firing in certain parts of the maze regardless of trial condition, and others firing under very specific conditions (e.g., only when turning left, only in the nonmatch phase, or only during sample right/nonmatch left trials). Some typical examples are shown in Figure 5. The average firing rate across the entire experiment was higher for these cells than for cells that did not change firing rates during the task because of increased rate during the task, but there was no net statistical difference in firing frequency between the sham and mTBI groups (Fig. 4C).
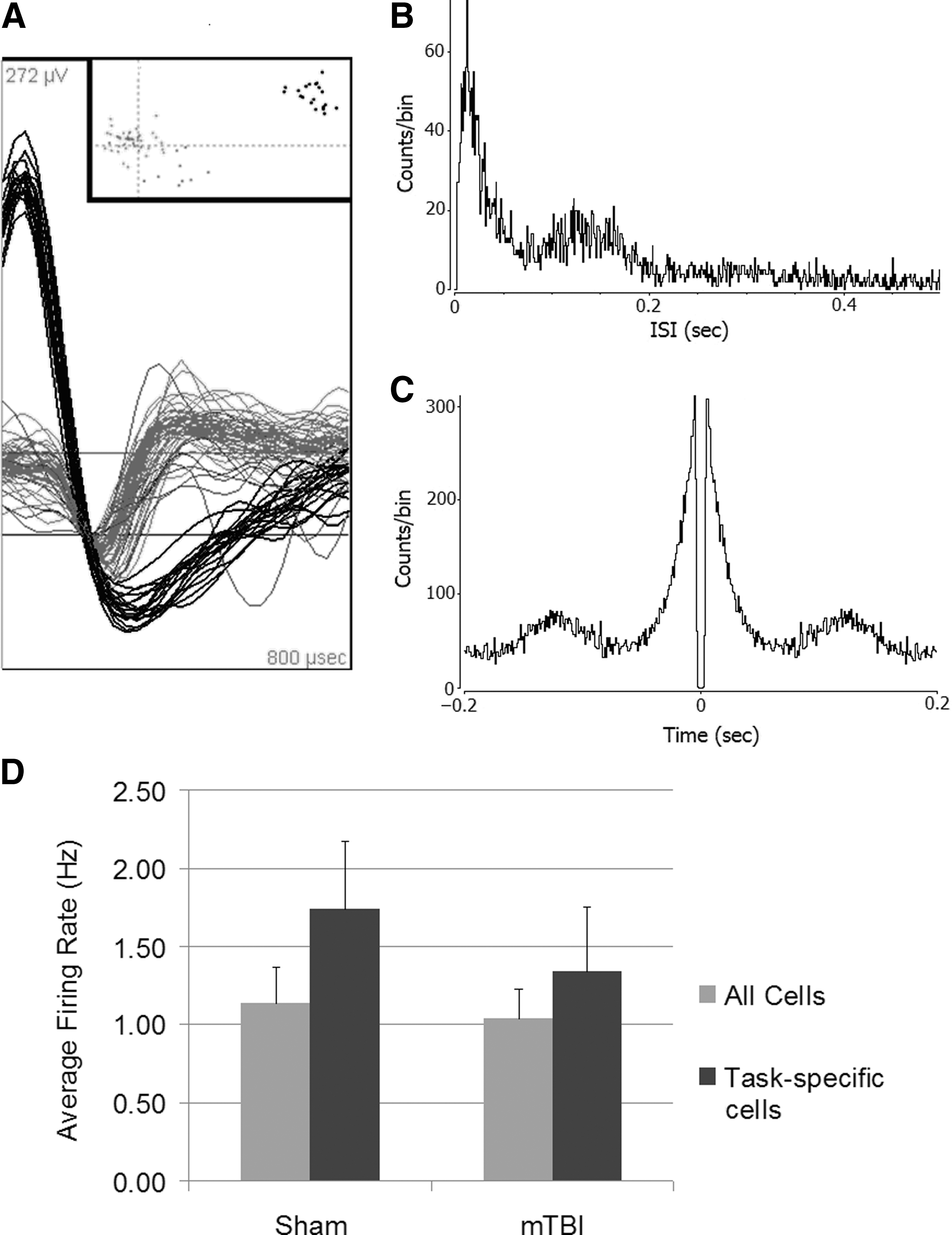
Neuronal recordings from hippocampal pyramidal cells. (
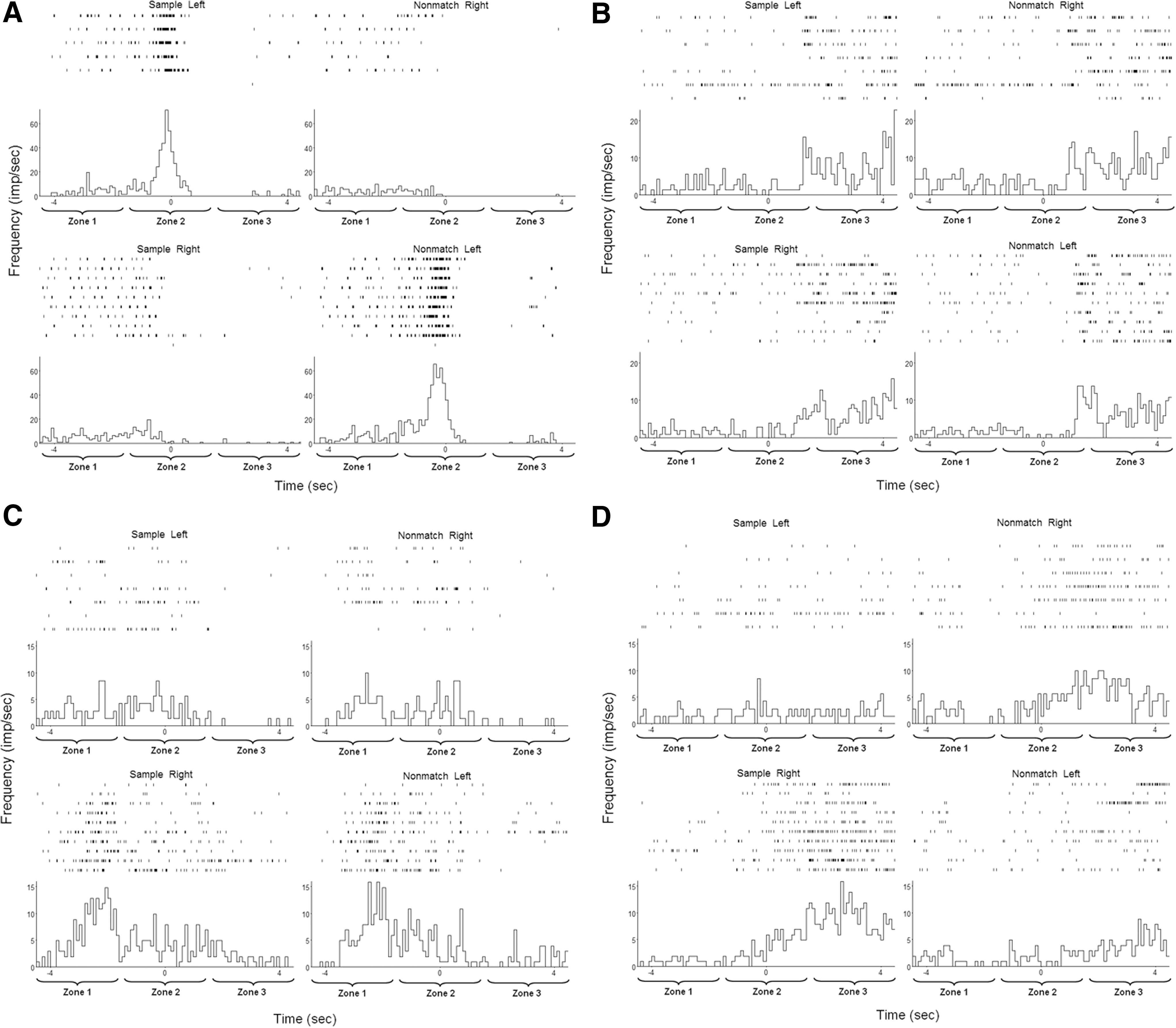
Examples of “place” cells. Four cells from a single animal (rat 2) are shown demonstrating firing characteristics during 17 correct completions of the maze (7 sample left-nonmatch right and 10 sample right/nonmatch left). Individual peri-event rasters are shown along with peri-event histograms (100 msec bins), centered on the decision point. Differences were considered significant if the mean firing rate across the entire 3-sec epoch lay outside the 99% confidence interval based on the Poisson distribution of spikes for that cell across the entire experiment. All four of these cells met this criterion. (
The proportion of cells with and without task-specific physiology for each rat is shown in Figure 6. Altogether, there were 6 such cells among the mTBI group, compared with 18 in the sham group, which represents a significant difference (p<0.05 by Fisher's exact test). There was no significant difference in the proportion of total or task-specific cells per CA1 or CA3 hippocampal subfield.
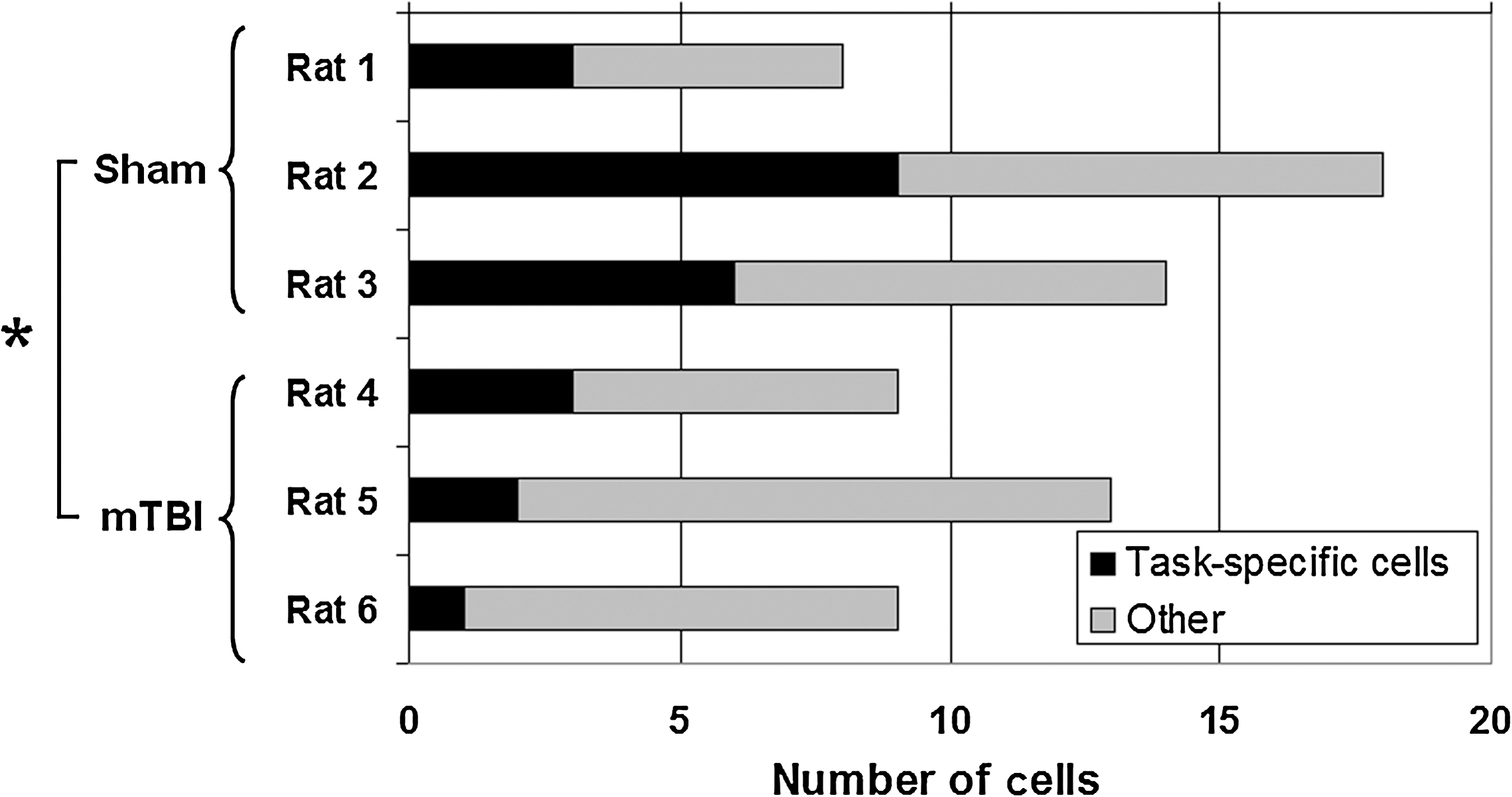
Proportion of task-specific and non-task-specific pyramidal cells. There were significantly fewer cells with consistent changes in firing rate with the task after mild traumatic brain injury (mTBI) (6 of 31 total cells), compared with sham rats (18 of 40 total cells; *p<0.05 by Fisher's exact test).
Histology and stereology
Histological analysis was performed using neutral red and analyzed by visual inspection. No differences were seen between the sham and mTBI groups. Stereological analysis of CA3 ipsilateral to injury demonstrated no significant difference in the average number of cells per unit volume between the mTBI (M=296.33±SEM=34.35) and sham (M=283.67±SEM=9.84) groups [t(4)=0.354, p=0.741; Fig. 7].
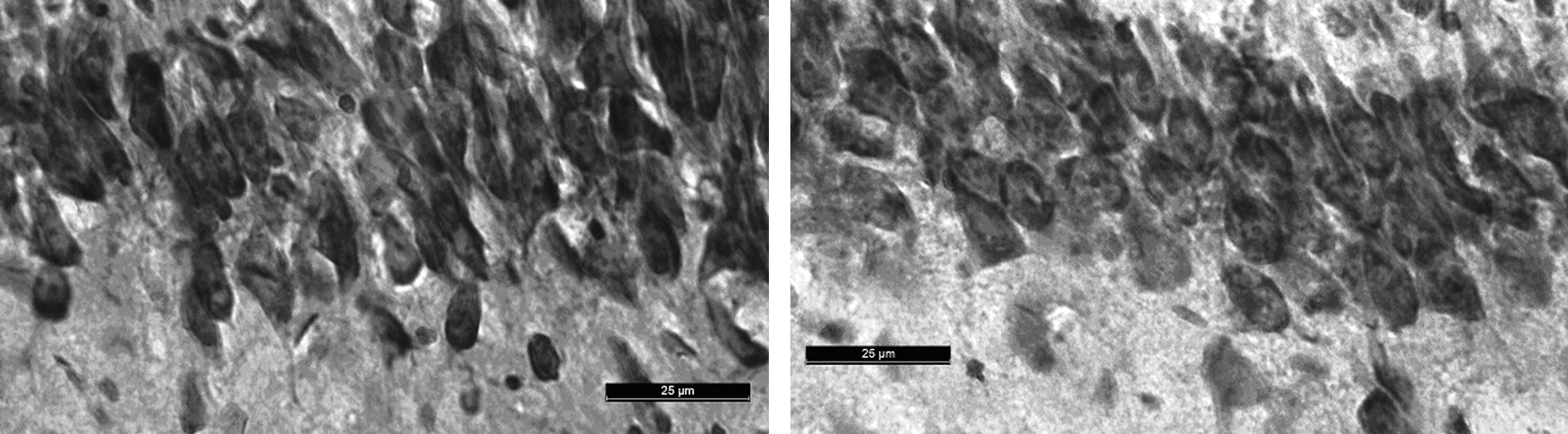
Representative images of similar areas of CA3 from (
Discussion
Much is known about cognitive and behavioral dysfunction after mTBI, but there is scant literature about electrophysiological abnormalities that may contribute to these changes. Previous studies using hippocampal slices have documented differential alterations in synaptic efficiency and excitatory post-synaptic potential-spike activity in various hippocampal subregions (Cohen et al., 2007; D'Ambrosio et al., 1998; Witgen et al., 2005), but these observations were made after moderate LFPI (2.0 atm), and were associated with loss of up to 40% of neurons in all subregions of the hippocampus. Analysis of local field potentials from the hippocampus in awake rats demonstrated that moderate TBI (2.15 atm) is associated with smaller increases in theta band activity when comparing quiet behavior to active rearing, although without a significant difference during performance in a Barnes maze (Fedor et al., 2010). To date, there has been no attempt to analyze the effect of mTBI on hippocampal single-unit activity during performance of a complex task.
In the present study, we demonstrated that mTBI is associated with less organized firing patterns in the hippocampus during swimming in a DNMP T-maze, possibly accounting for the difference in performance seen in the same animals. We chose the DNMP swim T-maze task because it requires integration of both spatial (left/right) and non-spatial (match/nonmatch) information. The task does not require food or water deprivation and can be run repeatedly (unlike the Morris water maze), and increasing the delay can be used to identify subtle differences in working memory. Previous research using the DNMP swim T-maze has established that moderate TBI (2.0 atm) is associated with decreased performance with increasing delay, which suggests that TBI is associated with impaired ability to handle increased demand on working memory systems (Whiting and Hamm, 2006). We have confirmed that this effect is seen with mTBI (1.5 atm) in the absence of significant neuronal loss by stereological analysis.
It has been shown by multiple investigators that pyramidal cells in the hippocampus often demonstrate firing patterns that are spatially and temporally correlated with task context. Cells with action potential frequency tuned to the location of the animal in an open field during foraging (“place cells”) have been observed in CA1, and this process is known to be dynamic with exploration of a new environment (Wilson and McNaughton, 1993). Such correlation of cellular activity with position is also seen during swimming in a water maze (Hollup et al., 2001). Using a forced alteration task, it has been established that differential place- and context-associated firing patterns occur in CA1 and the medial entorhinal cortex, respectively, and that these structures encode complementary aspects of spatial information (Lipton et al., 2007). In a delayed nonmatch-to-sample task, it has been shown that that some CA1 and CA3 cells are capable of encoding nonspatial information (e.g., sample versus nonmatch phase), as well as position (Hampson et al., 1993), and cells with different characteristic patterns are located in discrete places along the axes of the hippocampus (Hampson et al., 1999). There is evidence that these cells are part of the primary mechanism mediating task performance, since task errors are associated with loss of these patterns (Hampson and Deadwyler, 1996), and artificial manipulation of their activity can degrade or enhance performance (Berger et al., 2011). While the physiology of hippocampal cells has been extensively studied, there has been little investigation of single-unit hippocampal physiology in disease states. One study of the effects of aging on hippocampal activity demonstrated similar numbers of cells with place-specific firing patterns, but with more consistency of place fields over time for young rats (12 months old) compared with older rats (24 months old; Barnes et al., 1997). By contrast, in the present study, we have shown that mTBI is associated with a smaller proportion of cells that fire in consistent patterns during task performance.
Our data suggest a relative paucity of task-specific patterns of cellular activity after mTBI in the absence of a change in the rate of firing between groups. The association of such findings in animals with reduced working memory suggests that the deficits seen after mTBI may be related, in part, to disordered firing of individual neurons that mediate task performance. Because the stereological analysis did not reveal significant cell death in CA3, and visual inspection of the hippocampal sections did not reveal overt histological abnormalities, it is unlikely that these changes are due to direct cellular damage. Previous research into the physiology of so-called “place cells” within CA1 has indicated that disconnection of entorhinal or CA3 input can lead to more diffuse and less discrete position-associated firing fields within CA1 (Ahmed et al., 2009). Likewise, it is known that lesions of either the fornix or entorhinal cortex reduce the total number of hippocampal pyramidal cells with firing rates related to place, without any change in the distribution of such cells within hippocampal subfields (Miller and Best, 1980). Ibotenic acid lesions to cortical regions immediately dorsal to the hippocampus and radio-frequency lesions to the fornix and fimbria are able to impair T-maze performance in a spatial memory task (Clark et al., 2000). Therefore, although we did not directly investigate the effect of mTBI on pathways associated with the hippocampus, our observation of a reduction in the relationship between firing and place after mTBI may suggest network dysfunction on a fairly large scale. We found that mTBI was associated with a decreased number of neurons with task-related specificity without any overt cell loss, so nonlethal disruption of neuronal function may relate to memory impairment during a cognitively demanding memory task. Specifically, changes in overall firing rate, burst activity, and cell survival may not be prerequisite for impaired memory function in cognitively-demanding tasks. Future studies are needed to determine the relationship between global network dysfunction, hippocampal connectivity, and single-unit changes in a TBI model.
There are several important limitations to this study. First, analysis of electrophysiological data was performed only from correct trials, because there were too few incorrect decisions to allow analysis of the difference between correct and wrong choices. Previous reports have demonstrated that incorrect trials are associated with decreased task-specific physiology in intact animals (Hampson and Deadwyler, 1996), and the effect of mTBI on such differential effects in the same cell population is unclear. Second, despite evidence that mTBI affects the substrate for development of cells that underlie task-specific physiology, there was no difference in the rate of acquisition of the task between sham and mTBI rats, suggesting that the dynamic process by which task-specific cells develop may be relatively unaffected despite large differences in the final number of cells showing task specificity. Third, although we demonstrated that swim speed was approximately the same across animals, the definition of maze zones by temporal rather than spatial criteria does introduce uncertainty as to the exact location of the animal during different firing patterns. Finally, because of the small number of animals in each experimental group, it is possible that incidental factors other than exposure to mTBI may have influenced the results, although it is unlikely because the groups differed only in terms of their prior exposure to mTBI.
Conclusions
We have demonstrated that mTBI is associated with a significantly smaller subset of hippocampal pyramidal cells that demonstrate consistent context-specific firing patterns during performance of a memory task. Because this event occurs without any appreciable cell loss or other electrophysiological change, nonlethal disruption of neuronal physiology may be related to cognitive deficits seen after mTBI.
Footnotes
Acknowledgments
We thank Osmond Wu, B.A., and Thomas Ostergard, B.S., for assistance with the experiments. Financial support has been provided by Case Western Reserve School of Medicine and the Neurosurgery Research and Education Foundation of the American Association of Neurological Surgeons.
Author Disclosure Statement
No competing financial interests exist.