
Abstract
Gene expression of specific brain biomarkers offers the possibility of shedding light on the difficult molecular pathways of traumatic brain injury (TBI) and may be useful to estimate the age of trauma. Gene expression rates of cerebellar injuries are not yet sufficiently established. In 12 cases (mean age 42 years) of TBI including a pathological change in cerebellum (with known survival times ranging from immediate death to 96 h), brain tissue samples from different brain regions were analyzed with real-time polymerase chain reaction (PCR) for expression of caspase-3, tyrosine kinase receptor B (TrkB), S100B, and glial fibrillary acidic protein (GFAP) mRNA. The pH was measured to gain information about a possible correlation to RNA degradation. For comparison, corresponding brain regions were arranged from control samples of subjects that died from sudden death.
We found a correlation between pH and the degradation of RNA in samples from the contralateral site, where the samples with degraded RNA have a lower pH (p<0.05). For short survival times, the expression changes of caspase-3 (p<0.05) and the expression changes of TrkB (p<0.1) in the cerebellum show a significant increase compared to the controls. The cerebellar gene expression changes seem to occur much faster and stronger compared to the other investigated regions, in particular the cerebral trauma site. These findings could make the cerebellum an important target area to study the expression changes after TBI.
Introduction
P
Methods
Sample collection
Brain tissue samples were collected semi-sterile from 14 individuals during autopsy. Samples included the traumatically injured site of the cerebrum (i.e., the macroscopic or microscopic contusion area), contralateral area (i.e., the gyrus corresponding to the trauma site in the other half of the cerebrum), hippocampus, and a fraction of the cerebellum from the trauma containing cerebral site. It should be mentioned that there were no macroscopic visible contrecoup injuries in the recruited trauma cases. The samples were cut into pieces of about 1 cm3 and stored at −80°C until analysis or in 4% formaldehyde for histological investigations, respectively. The samples from two control individuals included prefrontal cortex, hippocampus, and cerebellum from the same hemisphere and were identically stored.
The trauma group consisted of 12 cases (five males, seven females) with a mean age of 42 years (range 18–81 years). The control group had only male individuals (mean age, 52 years). In all trauma cases, cranial fractures (n=10) and/or brain contusions (n=7) were diagnosed; intracranial hemorrhage (epidural, subdural, and/or subarachnoidal) as a sign of vascular trauma was found in all cases (n=12). Causes of trauma were traffic accidents in nine cases, and three cases of fatal falls. Survival times ranged from immediate death to 96 h, with eight cases surviving the trauma less than 2 h, and four cases surviving up to 4 days. The mean post-mortal delay (i.e., the time between death and beginning of the section) was 88 h (range, 25–312 h). Macroscopic signs of putrefaction were not found in any case. In two individuals increased blood alcohol concentrations were found (c>2% or g/L). None of the individuals showed toxic concentrations of narcotic agents.
Strict criteria were established for the selection of two control cases: only individuals who died from natural death (myocardial infarction, ruptured thoracic aneurysm), without any signs of external injuries, and with a short post-mortal interval (up to 2 days) were recruited. Neither macroscopic nor microscopic changes of brain tissue were found in the control samples. The clinical characteristics of the individuals in respect to their treatment are shown in Table 1.
CA, catecholamines; CPR, cardipulmonary resuscitation; I, intubation; M, man; NST, neurosurgical treatment; OD, osmotic drugs; W, woman.
Measurement of tissue pH
The pH was determined from frozen tissue. In TBI cases, tissue from the cerebral trauma site and the contralateral site was used. About 200 mg of tissue were homogenized using a pestle in 10 volumes of distilled water (adjusted to pH 7.0). The pH values were measured at room temperature with a pH-meter (MP, Mettler-Toledo, Columbus, OH). Measurements were made in duplicate for each sample.
RNA extraction
RNA was extracted from 50–100 mg of frozen tissue using TriFast reagent (Peqlab, Erlangen, Germany) according to the manufacturer's instructions. RNA concentration was measured on NanoDrop and RNA integrity was verified by agarose gel electrophoresis.
Reverse transcription and quantitative PCR
Two μg of RNA were reversely transcribed into cDNA with GoScript Reverse Transcription System (Promega). Quantitative real-time PCR (qPCR) was carried out at the ABI7300 (Applied Biosystems, Foster City, CA) using GoTaq qPCR Master Mix (Promega, Madison, WI). The cDNA was used in 1:10 dilutions for qPCR, which was freshly prepared before each run.
The primers for qPCR were designed using PerlPrimer software (V1.1.19;
The sequences of primers were as follows: HMBS_fw GTAACGGCAATGCGGCTGCA, HMBS_rev GGTACCCACGCGAATCAC (annealing temperature 57°C); SDHA_fw ACGTTCGACCGGGGAATGGT, SDHA_rev TTCCGTGCCTCTGCTCCGTA (60°C); GAPDH_fw GCACCACCAACTGCTTAGC, GAPDH_rev GGCATGGACTGTGGTCATGAG (60°C); Casp_fw AAGCACTGGAATGACATCTC, Casp_rev AAACATCACGCATCAATTCC (59°C); TrkB_fw GCAGGTGATCCGGTTCCTAA, TrkB_rev CCTGTGTGTGGCTTGTTTC (57°C); S100B_fw GGAAGAGGATGTCTGAGCTGGAG, S100B_rev AGCTTGTGCTTGTCTCCCTCC (59°C); GFAP_fw CTGCAGATTCGAGAAACCAG, GFAP_rev TCCTTAATGACCTCTCCATCC (59°C).
To verify accurate amplification, a dissociation step was added after the PCR run.
Data analysis
To determine the efficiency of every PCR run, the program LinRegPCR was used (
Statistical tests (Student's t-test and ANOVA test for correlation) were carried out with the analysis features of Microsoft Excel (2003; Bellevue, WA) and OpenOffice (OpenOffice.org, V3.2), respectively.
Results
pH measurement of brain tissue
The pH was measured on tissue of traumatically injured individuals (cerebral trauma site and contralateral site) as well as on individuals that died from other reasons (external control). In all experimental groups, the pH was about 6.1±0.3, with differences found only in the second decimal column.
Recent studies have shown that there is no clear correlation between pH and RNA integrity. Kingsbury and colleagues 7 found a pH influence on the preservation status of the RNA, and Mexal and colleagues 8 showed that differences in pH lead to altered expression profiles. In contrast to this, Ferrer and colleagues 9 reported that pH is not a useful variable to predict the RNA quality.
We tested a possible correlation between brain pH and RNA degradation. For the trauma sites, we observed no significant differences in pH in samples with or without degraded RNA. Nevertheless, we found a correlation between pH and degradation of RNA in samples from the contralateral site, where the samples with degraded RNA had a lower pH (pH 5.8 compared to pH 6.1 in the trauma site; p<0.05). This finding is consistent with possible physiological regulation mechanisms in the brain in that a secondary tissue ischemia can lead to a light metabolic acidosis. 10 The correlation test (ANOVA) between pH and patient-dependent parameters, such as sex, age, survival time after injury, and post-mortal delay, did not reveal any correlation, which is partially in agreement with a study by Monoranu and colleagues. 11
Real-time PCR
For each sample, the mRNA expression of three reference genes—hydroxymethylbilane synthase (HMBS), succinyl dehydrogenase subunit A (SDHA), and glycerinaldehyde-3-phosphate dehydrogenase (GAPDH)—was measured and a normalization factor was determined. The mRNA expression of the candidate genes was normalized according to this normalization factor. These normalized data were then related to the expression of the respective gene in the control samples. The obtained data represent the relative expression of the target genes in the TBI samples compared to that of the controls (Fig. 1).
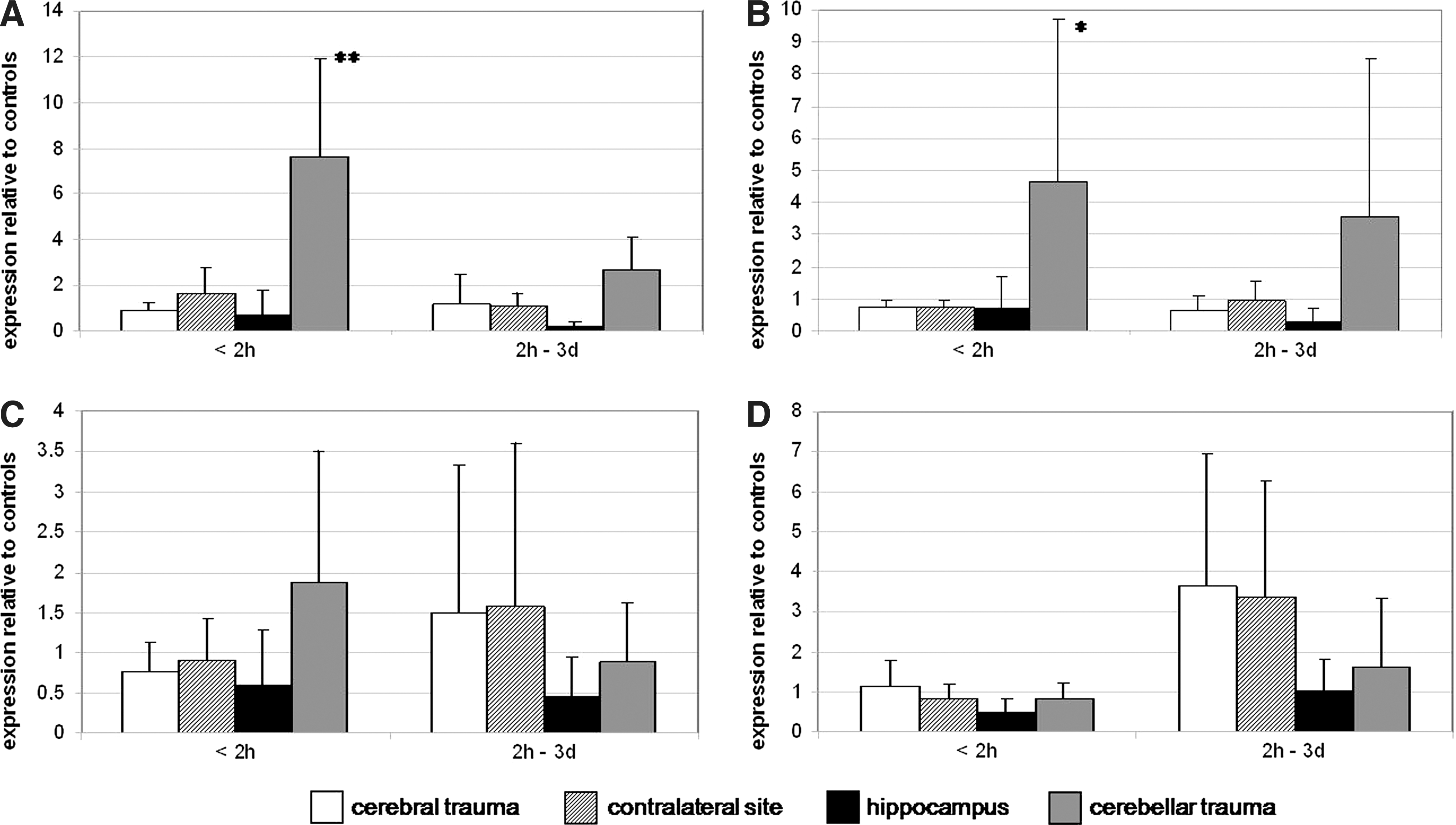
Expression of caspase-3, TrkB, S100B, and GFAP mRNA in the cerebral site, the contralateral site, the hippocampus, and the cerebellar site after traumatic brain injury. (
According to the survival time, the samples were divided into two groups—the first having survival times of up to 2 h, the second comprising samples with a survival time of >2 h to 4 days.
Caspase-3 and TrkB show no or only slight differences in mRNA expression after 2 h survival time, except for in the cerebellum. This brain region shows a higher expression of caspase-3 and TrkB mRNA than in the other investigated brain regions, both for a survival time of less than 2 h and 2 h to 4 days (Fig. 1A,B). For short survival times, the expression changes of caspase-3 in the cerebellum show a significant difference (p<0.05) in comparison to the control samples. The cerebellar expression of TrkB is also significantly higher in the group with short survival times compared to the not-traumatically injured control brains, but with a lower significance level (p<0.1).
The expression of S100B mRNA is about two-fold higher in the cerebral trauma site in cases that survived the traumatic brain injury more than 2 h compared to those with shorter survival times. Also the contralateral site shows a higher expression level of S100B in cases with a longer survival period. A horizontal brain herniation seems not to be the reason for this finding, because only two trauma cases with midline shift of the cerebral hemispheres were detected. The highest expression differences, albeit in the opposite direction, were detected for the cerebellum (i.e., a higher expression of S100B mRNA in the group with short survival times, and a decreased expression level in the cases surviving from >2 h to 4 days). In the hippocampus no significant changes could be observed (Fig. 1C).
The expression differences of GFAP mRNA in the cerebral trauma and contralateral site are nearly similar to those of S100B. An activation of GFAP mRNA seems to occur after a time of 2 h to 4 days. A slighter increase of GFAP mRNA was found in the hippocampus and cerebellum (Fig. 1D). None of these expression changes shows any significance.
Additionally, in two of the trauma cases increased blood alcohol concentrations were detected, but there were no significant gene expression differences in those cases in relation to the other trauma samples.
Discussion
The presented data indicate changes in the expression of the mRNA levels of four marker proteins in the brain after TBI in respect to survival time and trauma region.
Surprisingly, it appears that mRNA expression of caspase-3, TrkB, and S100B is much stronger in the cerebellar trauma site than in the cerebral trauma site up to 2 h after injury. All investigated cerebella showed signs of contusion or tissue lacerations; therefore this activation of mRNA may be a direct reaction due to injury. A fracture line through the posterior part of basal skull could be observed in eight cases. These expression changes seem to occur very quickly, within 2 h after the injury. Longer periods of survival lead to a decrease in the expression level of the biomarkers caspase-3 and TrkB, as well as biomarker S100B in the cerebellar trauma side.
However, significant expression changes could be observed for caspase-3 and TrkB in the cerebellar trauma sites with short survival times in comparison to the control cases. Caspase-3 is involved in apoptotic processes. An increase in expression has been observed via immunohistochemical methods within hours after a traumatic injury in humans 1 and the co-expression on GFAP-positive astrocytes (e.g., in rat traumatic injury). 12 Also a correlation between severity of trauma and the mRNA expression level of caspase-3 was found in the rat brain. 13 However, only the expression of mRNA and protein in the cerebral areas of trauma were studied. The results presented here may give first insights into the processes occurring after TBI with special focus on the expression in the cerebellum.
The TrkB gene encodes for the neurotrophic tyrosine kinase receptor B. This is a membrane-bound receptor for brain-derived neurotrophic factor (BDNF). A study conducted by Benisty and colleagues 4 revealed that TrkB mRNA is constitutively more strongly expressed in the cerebellum compared to other brain regions. The data reported here were normalized to controls, so one can state that the high expression level in the cerebellum is in fact a reaction to the traumatic injury. Benisty and colleagues 4 also described a high mRNA expression of TrkB in the cerebral cortex. No difference seems to occur in the expression level after a TBI in this region in our study (Fig. 1).
The mRNA expression of S100B seems to increase with longer survival times, which is in concordance with the findings of von Gertten and colleagues 14 in the injured rat brain, as well as on protein levels, where an increase in S100B concentration was reported 1–2 days after the injury. 15
GFAP is widely accepted as a specific marker for fibrillary and reactive astrocytes. Astroglial reaction after TBI includes the formation of a glial scar at the side of an acute lesion that may be harmful, inhibiting axonal regeneration, but otherwise beneficial, promoting healing and functional recovery of neuronal pathways in healthy tissue. 16 –18 An increase of GFAP mRNA expression in the cerebral trauma site is therefore not surprising because astrocytes become activated and could possibly migrate into the injured region, as described by Goldberg and colleagues. 19 Even new data show that human astrocyte migration is proceed. 20,21
In respect to the research literature, it should be mentioned that not all astrocytes are GFAP-positive in rat brain 22 and that there is a heterogeneity of reactive astrocytes, depending on the distance to the lesion site. 23
Of special interest are the findings that an increase in GFAP mRNA does not occur immediately but rather after a period of >2 h. A study performed in mice brain after experimentally induced trauma reports the earliest reaction of GFAP mRNA expression showing at 1 day post-injury and peaking after 3 days. 24 An increase of GFAP protein in the human brain was measured immunohistochemically 1 day after the injury. 3
Finally, ramified protoplasmic astrocytes are the predominant form in gray matter in almost all regions of the central nervous system while in the cerebellum another form of elongated radial glia exist (Bergmann glia) that share many features with the protoplasmic astrocytes in respect to their perisynaptic association. 25 Therefore GFAP expression, but no glial demarcation, seems to follow the lesion in the molecular layer of cerebellum. 26
In four cases with survival times of only up to a few minutes, none or unsuccessful, brief medical treatment is documented (see cases 1, 5, 9, and 10 in Table 1). These measured data served as internal control to check for specific results. The changes in expression in these few cases are altogether similar to the other samples in the group with survival times of less than 2 h with a recorded anesthesia chart, except for the changes for S100B in trauma site, where it seems a longer survival period is needed for specific expression changes. In comparison with the data in the literature, the presented changes in mRNA level after TBI appear characteristic, 27,28 but it cannot be completely excluded that additional factors (e.g., cardiopulmonary resuscitation) may influence the level of mRNA expression.
We find high standard deviation in almost all of the investigated sample groups. This fact is certainly due to the heterogeneity of the brain material and the different patterns of injury. The inclusion of cases with a high post-mortal delay may also be a reason for high standard deviation values. We decided to use these cases because the quality of the RNA seemed to be sufficient and survival times like this reflect the width of circumstances in reality.
However, the expression changes on the mRNA level do not necessarily represent the changes of the protein concentration. To verify the present results of mRNA expression changes, it is important to investigate these gene products on the protein level (e.g., via Western blot analysis or immunofluorescence methods).
It is well known that S100B and GFAP are released to the peripheral blood circulation soon after brain injury occurs and are helpful in predicting the clinical outcome of patients. 29,30 Both markers are associated with intracranial as well as intracerebral hematoma. Previous studies showed that GFAP and S100B serum concentrations change with time. The concentrations of GFAP and S100B increase over time in the first days after trauma, 31 corresponding to our results for the GFAP and S100B mRNA expression in cerebral trauma and contralateral site as well as cerebellar trauma site almost completely. Up to now there are no conclusive studies of these biomarkers on post-mortem analysis of human blood serum available.
In conclusion, we present novel findings for the gene expression in the cerebellum following a TBI, which also include the anatomic structures of the posterior part of basal skull. The cerebellar gene expression changes appear to occur much faster and with greater strength compared to those in other investigated regions, in particular the cerebral trauma site. These findings could make the cerebellum an interesting target region to study the expression changes after TBI and the examination of the wound age.
Footnotes
Author Disclosure Statement
The authors declare that they have no financial interests to disclose. The investigations were conducted in accordance with the human and ethical principles of the University of Leipzig (no. 117-12-23012012).