
Abstract
End-to-side neurorrhaphy is widely used in the peripheral nervous system for nerve repair; however, the application of this technique has been limited to somatic nerves. The feasibility of nerve regeneration through end-to-side neurorrhaphy between autonomic and somatic nerves with different characteristics in the peripheral nervous system is still undetermined. In this study, rats were divided into three groups for different treatments (n=10 per group). In the end-to-side neurorrhaphy group, left L6 and S1 were transected in the dura, and the distal stump of L6 ventral root was sutured to the lateral face of L4 ventral root through end-to-side coaptation. In the no repair group, the rats did not undergo neurorrhaphy. In the control group, the left L6 dorsal root and S1 roots were transected, respectively, but the L6 ventral root was kept intact. After 16 weeks, the origin and mechanism of nerve regeneration was evaluated by retrograde double labeling technique as well as histological examination and intravesical pressure measurement. Retrograde double labeling indicated that the reconstructed reflex pathway was successfully established and the primary regeneration mechanism involved axon collateral sprouting. Morphological examination and intravesical pressure measurement indicated prominent nerve regeneration and successful re-innervation of the bladder in the neurorrhaphy group, compared with the “no repair” group (p<0.05). No significant changes were observed in the histology of the donor nerve and the bilateral extensor digitorum longus muscles in the neurorrhaphy group. Nerve regeneration may be achievable for nerve repair through end-to-side neurorrhaphy between autonomic and somatic nerves without apparent impairment of donor somatic nerve.
Introduction
N
End-to-side neurorrhaphy is a surgical technique that involves suturing the distal end of a transected nerve to the lateral face of an adjacent healthy nerve to induce collateral sprouting from the healthy donor nerve and minimize the donor nerve morbidity, while promoting recipient nerve function recovery simultaneously (Guena et al., 2006). The concept of end-to-side neurorrhaphy was proposed first by Létiévant in 1873. Many relevant reports have been published to describe the success of nerve repair as end-to-side neurorrhaphy since Viterbo (Lundborg et al., 1994; Viterbo et al., 1992, 1994a, 1995; Zhao et al., 1997). The potential benefits of this approach are considerable. First, the detrimental effects to the donor nerve could be minimized. Second, the problems associated with end-to-end neurorrhaphy and nerve grafting can be avoided. Third, the distance required for re-innervation can be shortened thus decreasing the denervation time and the degree of target organ atrophy.
However, in most of the previous experimental and clinical studies, the end-to-side neurorrhaphy technique was limited to somatic nerves of the peripheral nerve system (Battiston et al., 2009; Beris et al., 2007; Bontioti et al., 2005; Jaberi et al., 2003; Mennen et al., 1999; Ozbek and Kurt, 2006). Is it possible to use this technique for nerve repair between autonomic nerve and somatic nerve with different characteristic in the peripheral nerve system? If this end-to-side neurorrhaphy technique is feasible between autonomic nerve and somatic nerve, then the detrimental effects to the donor somatic nerve may be avoided. We sought to answer these questions and report our results of a preliminary experiment with end-to-side neurorrhaphy of autonomic nerve and somatic nerve in rats.
Methods
Animal preparation and surgical procedure
Thirty healthy mature male Sprague–Dawley rats with an average weight of 250±30 g were used in this study. All the experiments were approved by our institute's Animal Care and Ethics Committee, and the animals were treated in accordance with the Animal Care Guidelines of the National Bureau of Health. All efforts were made to minimize the number of animals used and their suffering. The rats were randomly divided into three groups for different treatments, with 10 rats in each group.
All of the surgery was performed with the rats under deep anesthesia, using 10% chloral hydrate in deionized water solution (300 mg/kg body weight; Sinopharm Chemical Reagent, Shanghai, China) administered intraperitoneally. A schematic diagram outlines the surgical procedures performed within the three groups (Fig. 1). In group I, the end-to-side neurorrhaphy group (n=10), a transverse laminectomy from L3 to S1 with a midline incision on the back was performed under sterile conditions with the rats lying prone. The dura was opened by a paramedian incision, and then the dorsal roots (DRs) and the VRs from L3 to S1 on the left side were exposed and dissected. L6 VR, L6 DR, S1 VR, and S1 DR were transected. The distal stump of the L6 VR (recipient nerve) was trimmed diagonally using straight microscissors to create a 2-mm oblique surface, and the longitudinal axis of L4 VR (donor nerve) was wrapped in a helicoid fashion at a 40 degree angle. Then, the trimmed oblique end of L6 VR was coapted to the lateral face of the intact L4 VR with 10-0 nylon sutures (Ethicon 10/0, ETHICON, Shanghai, China) visualized under a microscope with 12.5×magnification (OPMI 9, Carl Zeiss, Germany). To prevent spontaneous nerve regeneration into the remaining stump, the proximal end of L6 VR was ligated and buried into the adjacent muscle. In group II, the “no repair” group (n=10), L6 VR, L6 DR, S1 VR and S1 DR were transected. However, the distal stump of L6 VR was not coapted to the lateral face of the intact L4 VR. In group III, the control group (n=10), L6 DR, S1 VR and S1 DR were transected, but L6 VR was kept intact. The most important significance of the design of group III was to provide the number of normal axons in the preganglionic parasympathetic nerve (PPN) and also to compare the number of regenerated axons with group I. The wound was closed in the layers in all rats. Penicillin (200,000 U/day) was administered intramuscularly for 3 consecutive days after operation. The rats were caged separately with free access to water and standard food at suitable temperature with a 12:12 h light:dark cycle. We selected 4 months as the survival period of model rats, to ensure that nerve regeneration could be achieved (Xiao and Godec, 1994). During the first month post-operation, it was necessary to squeeze the abdomen of the rats in groups I and II to promote micturition, thus avoiding urinary retention.
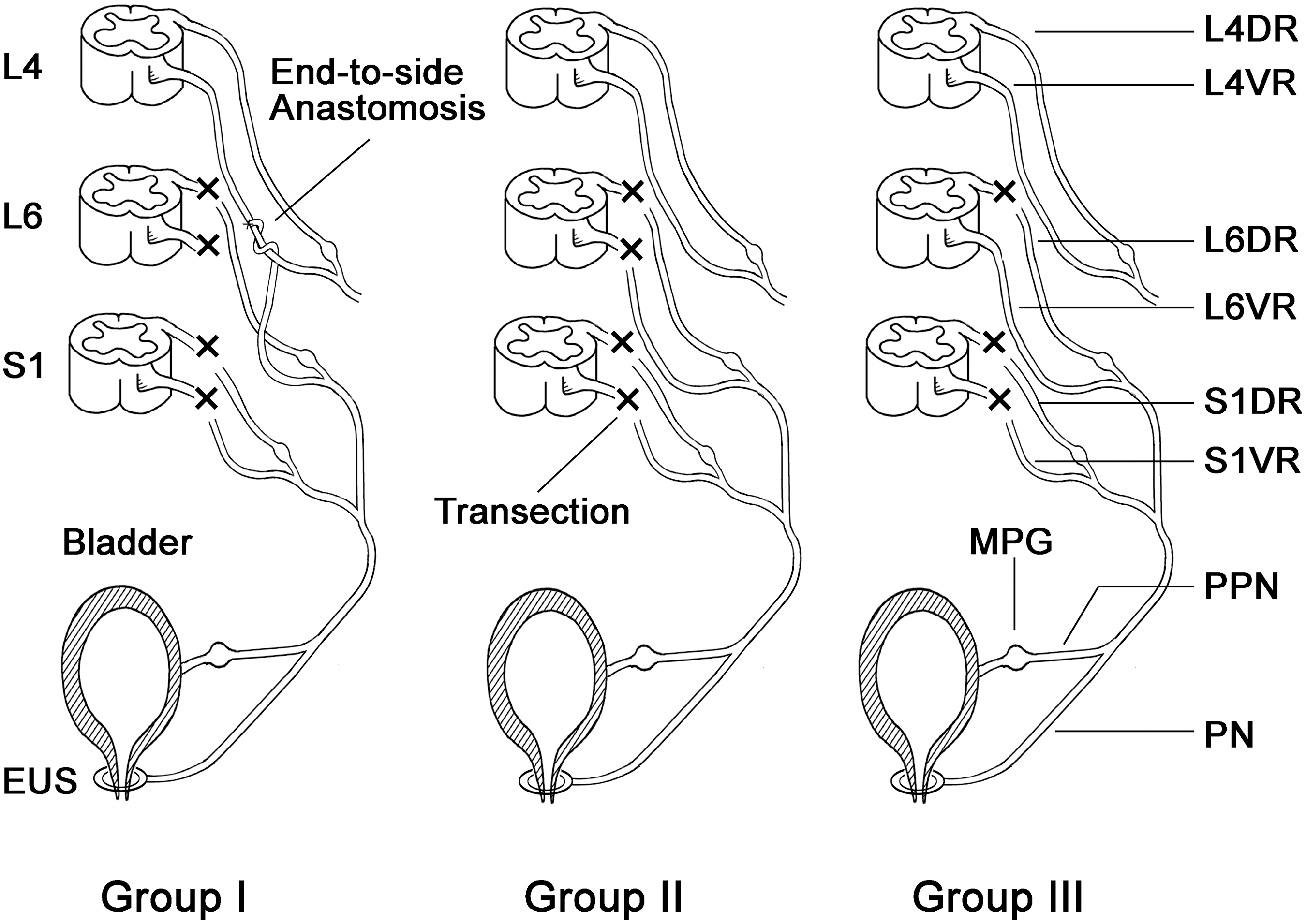
Schematic diagram outlining the surgical procedures performed in all the groups. In group I, L6 VR, L6 DR, S1 VR, and S1 DR were transected. Then, the distal stump of L6 VR (recipient nerve) was wrapped over the longitudinal axis of L4 VR (donor nerve) and coapted to the lateral face of the intact L4 VR. In group II, L6 VR, L6 DR, S1 VR, and S1 DR were transected. However, the distal stump of L6 VR was not coapted to the lateral face of the intact L4 VR. In group III, L6 DR, S1 VR, and S1 DR were transected, but L6 VR was kept intact. (Group I, end-to-side neurorrhaphy group; Group II, “no repair” group; Group III, control group; VR, ventral root; DR, dorsal root; PPN, preganglionic parasympathetic nerve; PN, pudendal nerve; MPG, major pelvic ganglion; EUS, external urethral sphincter).
Retrograde double labeling
After 16 weeks, all the rats in each group were re-anesthetized for retrograde double labeling. Two fluorescent retrograde axonal tracers, Fast Blue (FB, Polysciences, USA) and Fluorogold (FG, Biotium, USA) were used in the experiment to determine the origin of regenerative axons in the recipient nerve. First, 4% FG 0.5 μL was injected into the left major pelvic ganglia (MPG) of the rats in each group via a 1-μL syringe (Micro-injector, Shanghai, China) and allowed to stand for 10 min before leaving. To avoid contamination of the adjacent tissues or leakage of the FG, the injection site was covered with glycerin. After a transit time of 3 days, the other retrograde tracer, 3% FB 1 μL was injected in the same way into the left sciatic nerve (SN), that is, the peripheral nerve of L4 VR. During another 7 days of transit, intravesical pressure (IVP) measurement for each group and weighing of the extensor digitorum longus (EDL) muscle in group I were performed. After finishing the above mentioned procedure in addition to the measurement, the rats were perfused with 250 mL of 0.9% saline through ascending aorta followed by 500 mL of ice-cold 4% paraformaldehyde in a 0.1 M phosphate buffer saline (PBS, pH 7.4). After perfusion, the spinal cord segments from L3 to S1 were harvested, and postfixation was performed by placing 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS) for 4 h and they were then removed into a 30% phosphate-buffered sucrose solution overnight at 4°C for dehydration, and frozen in over the counter (OTC) resin. Transverse serial sections (20 μm thick) of the spinal cord were cut and obtained using a cryostat (Leica, CM8500, Germany) and mounted on gelatinized slides. Every fourth section was viewed and the FB or/and FG labeling neurons were counted using ultraviolet light under a fluorescence microscope (Olympus, IX71, Japan) equipped with a CCD camera (Nikon, DXm 1200, Japan).
Morphological examination of the recipient nerve and nerve fibers counting
After wet weight measurement of bilateral EDL muscles, the following 10 mm long nerve segments were harvested and prepared. In group I, first, the L6 VR and PPN ∼45–55 mm distal to the coaptation site were harvested without perfusion for semi-thin cross-section and ultrastructure preparation. The rats were perfused, then two segments of L4 VR that were 5 mm proximal and 5 mm distal to the coaptation site were also harvested. Finally, the corresponding PPNs of other rats in groups I, II, and III were also harvested to prepare paraffin section for comparison (Fig. 2). The obtained nerve specimens for paraffin sections were fixed in 4% paraformaldehyde overnight in 0.1 M PBS, dehydrated with graded ethanol, embedded in paraffin and cross-sectioned at a thickness of 5 μm, and stained with 1% toluidine blue (TB, Sigma-Aldrich, Shanghai, China) for comparison. The regenerative PPNs distal to coaptation were prepared for semi-thin cross-sections with a thickness of 0.5–1 μm and were cut using an ultramicrotome (Ultracut UCT, Leica, Germany) from the middle of the harvested PPNs, and stained with 1% TB for morphological examination. The microscopic images were digitized on a gray scale using automated digital analysis software (Image-Pro Plus 6.0, Media Cybernetics, USA) to calculate the axon amount in all fields of the sections (Jotz et al., 2011; Liao et al., 2009; Vaitkeviciene et al., 2006). The axons' number of PPNs originating from donor somatic nerve were also calculated by axon counting of the semi-thin section.
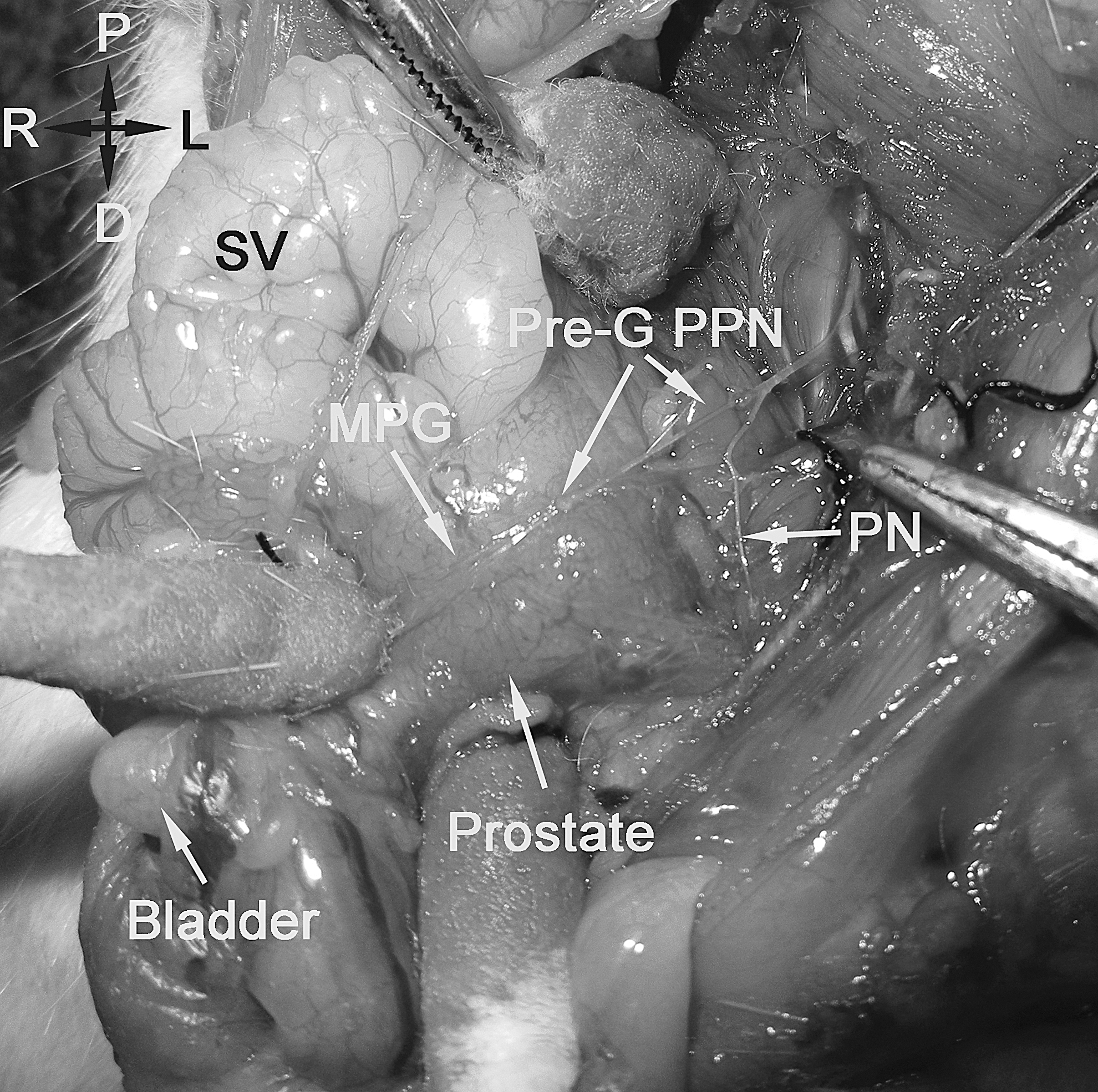
Macroscopic picture showing the anatomical location of the PPN after dissecting the adjacent nerves in group I. (SV, seminal vesicle; Pre-G PPN, preganglionic pelvic parasympathetic nerve; PN, pudendal nerve; MPG, major pelvic ganglion).
Ultrastructure of the regenerative axons
The harvested L6 VR distal to coaptation and PPN in group I were fixed in a solution of phosphate-buffered 2.5% glutaraldehyde, pH 7.4 for 12 h at 4°C and postfixed with 2% osmium tetroxide for 1 h, dehydrated, and embedded in Epon for ultrastructure observation. The morphology of the myelinated axons and unmyelinated axons in each cross-section was evaluated under transmission electron microscope (Tecnai G2, The Netherlands).
Measurement of IVP
On the seventh day after injecting fluorescent tracer FB, the rats were anesthetized and a vertical lower abdominal incision was made to identify the bladder along with the ureter on the both sides. The ureters were ligated and cut bilaterally. A polyethylene catheter (5-F, Shanghai, China) was inserted into the dome of the bladder through a small incision and was held in place with a purse-string suture. The catheter was tunneled subcutaneously and anchored to the skin on the back with a silk ligature so that no manipulation of the catheter could be done by the rats. The abdomen was closed and the bladder was emptied by gently pressing on the hypogastrium. After the rats were placed in the prone position, the left L4 VR proximal to the coaptation in group I, the left comparable segment of L4 VR in group II, and L6 VR in group III were dissected and exposed. After this procedure before the cystometry, the rats were gently restrained for 4 h to ensure that they were conscious from the anesthesia. IVP was measured on conscious rats by infusing the bladder with saline solution at room temperature, continuously using a microinfusion pump (WZ-50C, Hangzhou, China) at the rate of 2.4 mL/h. The urinary catheter was connected via a T-shaped tub to a microinfusion pump and a pressure transducer, which is a biological signal converter system (BL-410, Chengdu, China) to record the increased value of bladder pressure (mm Hg). A bipolar stimulating electrode was placed on the left L4 VR proximal to the coaptation in group I and the left comparable segment of L4 VR in group II. In group III, the left L6 VR was stimulated as normal control. The serial electrical stimulation was delivered at a frequency of 10 Hz for 1 ms with an intensity of 0.5 mA. The peak amplitude of the pressures was calculated by BL-410 software for analysis. Each rat was stimulated using the same parameters three times.
Wet weight and histological examination of the bilateral EDL muscles
As the EDL muscle of the rat is mainly dominated by L4 VR (Balice-Gordon and Thompson, 1988), we regarded it as an index to evaluate the function of L4 VR. After the IVP measurement, the rats were deeply anesthetized and EDL muscles were harvested bilaterally for wet weight measurement, and the right EDL muscle was used as an internal control for each rat in group I. The wet weight of the EDL muscle was recorded by harvesting the muscle without the tendon, gently blotting the surface with absorbent paper to remove any blood or serum and instantly weighing every muscle using an electronic balance (Mettler Toledo, AL 104, Switzerland). Following the wet weight measurement of EDL muscle, the muscle tissues were stored overnight in 4% paraformaldehyde in a 0.1 M PBS (pH 7.4) at 4°C, and the specimens were dehydrated with graded ethanol, and finally embedded in paraffin for hematoxylin and eosin staining (10 μm) to examine under light microscope.
Statistical analysis
All the data were expressed as mean±standard deviation (SD). The analysis of wet weight of bilateral EDL muscles in group I was computed using paired samples t test, and the possible differences between different groups were evaluated by one-way ANOVA using SPSS 15.0 for windows software. Statistical significance was set at p<0.05.
Results
Retrograde double labeling
The distribution and numbers of retrogradely fluorescent labeled neurons are presented in Table 1. In group I, FG single-labeled neurons and FG-FB double- labeled neurons were localized dominantly in the left ventral horn of L4, and FB single-labeled neurons were located in the left ventral and dorsal horns of L3–L5. The total number of FB-FG double-labeled neurons and FG single-labeled neurons were 7.8±1.8 and 1.1±0.9, respectively. The total number of FB-FG double-labeled neurons was significantly higher than that of FG single-labeled neurons in L4 (p<0.05) (Fig. 3A, Table 1). In group II, only FB single-labeled neurons were observed in the left ventral horn and dorsal horn of L3-L5; FG single-labeled neurons and FG-FB double-labeled neurons were not found in the L4 or L6 segments (Fig. 3B, Table 1). In group III, FB single-labeled neurons were observed in the left ventral and dorsal horns of L3–L5, and FG single-labeled neurons were observed in the left sacral parasympathetic nucleus of L6, but no FG-FB double-labeled neurons were found (Fig. 3C, D, Table 1). The total number of FB single-labeled neurons in groups II and III were significantly higher than that in group I (p<0.05; Table 1). No labeled neurons were observed on the contralateral side, indicating that no leakage of fluorescent tracers had occurred.
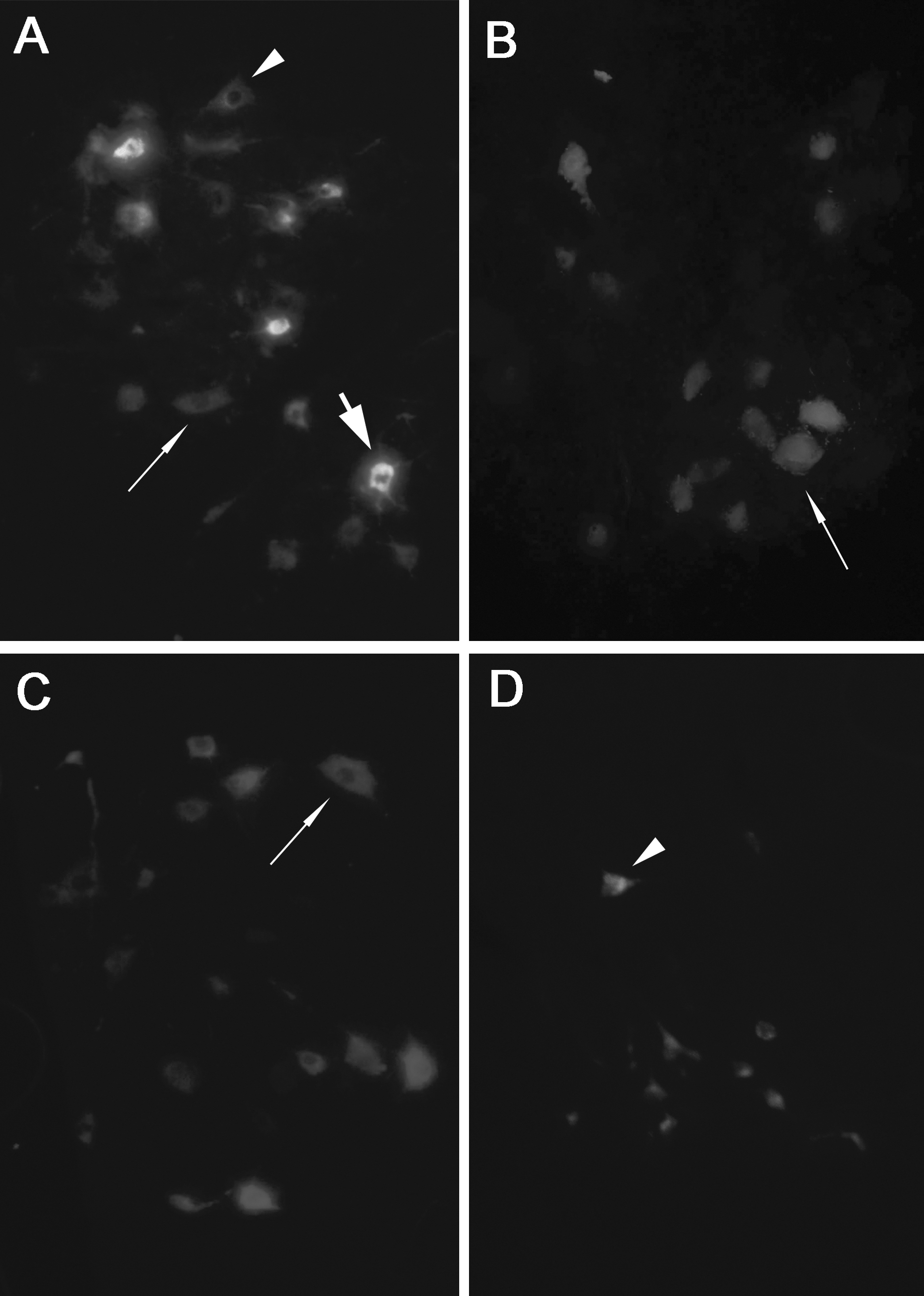
A fluorescence photomicrograph of retrogradely labeled neurons in different groups after injection of FG into the left MPG, and injection of FB into the left sciatic nerve.
p<0.05 as compared with the number of FG single labeled neurons in group I. † p<0.05 as compared with the number of FB single labeled neurons in groups II and III.
Group I, end-to-side neurorrhaphy group; Group II, “no repair group”; Group III, control group; FB, Fast Blue single-labeled neurons; FG, Fluorogold single- labeled neurons; DL, FB-FG double-labeled neurons.
Morphological examination of the recipient nerve and nerve fiber counting
When the neurorrhaphy site was exposed from adjacent tissues along the previous incision from the back, it was observed that the distal end of the L6 VR was already attached close to the lateral face of the L4 VR (Fig. 4). The morphological examination of PPN showed prominent axon regeneration in group I (Fig. 5 A, D). However, obvious myelin swelling and degeneration were observed in PPN in group II (Fig. 5B). The number of normal axons in the PPN was observed but no sign of degeneration was seen in group III (Fig. 5C). The mean number of myelinated axons of PPN in groups I, II, and III were 326.5±57.2, 2.1±1.6, and 533.6±80.0, respectively. There were statistically significant differences among the three groups (p<0.05; Fig. 5G). The mean number of myelinated axons in the proximal and distal coaptation site of L4 VR in group I were 872.9±14.8 and 870.1±18.1, respectively, and there were no statistically significant differences between the proximal and distal coaptation sites of L4 VR (p>0.05). Moreover, the mean number of myelinated axons was 882.2±10.3 in the comparable segments of L4 VR in group III, and there were no statistically significant differences with that of group I (p>0.05; Fig. 5H).

Macroscopic appearance of the end-to-side neurorrhaphy site between L6 VR and L4 VR 16 weeks after operation in group I (C, coaptation site; VR, ventral root distal to coaptation).
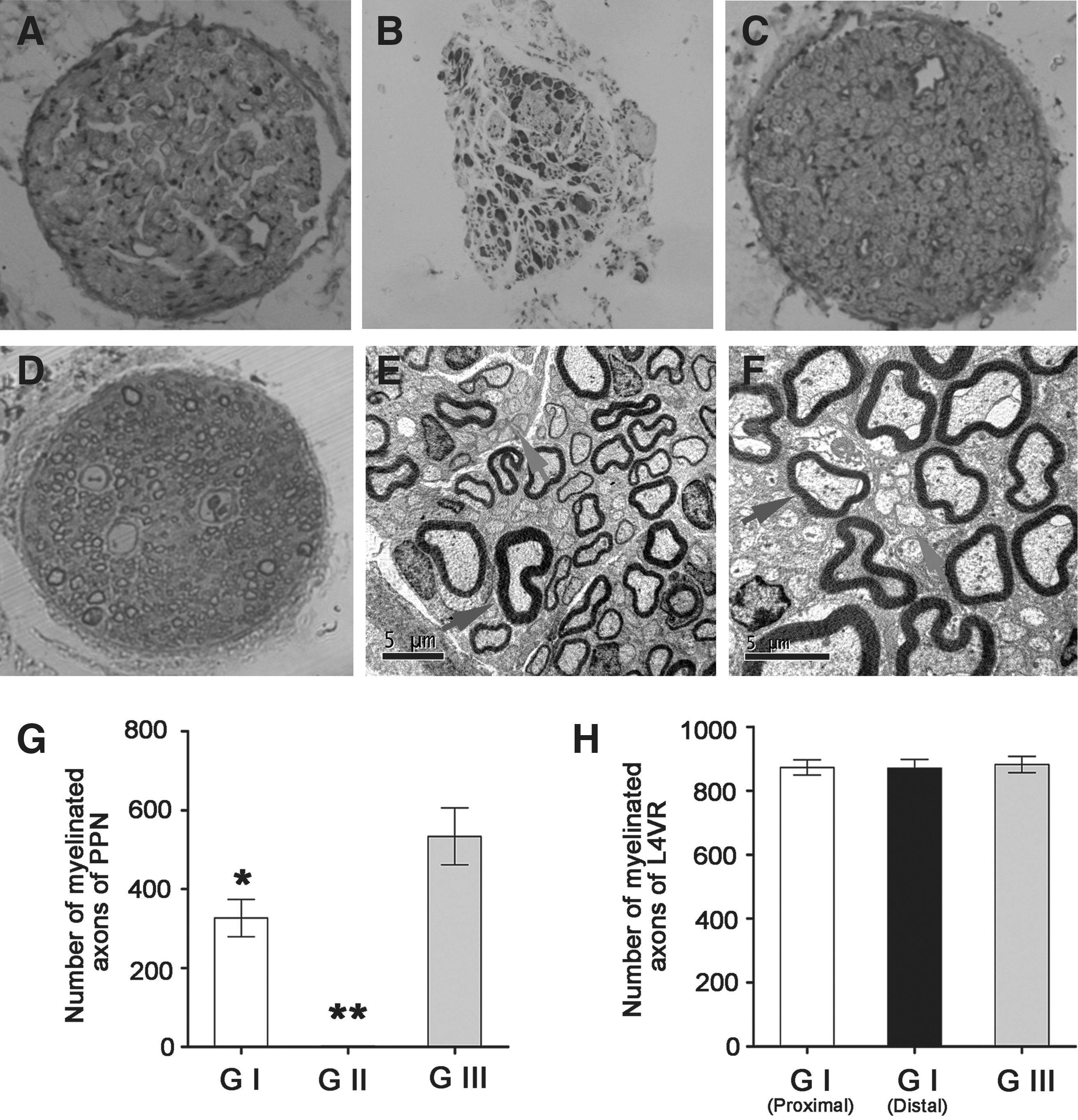
Morphology and quantification of nerve fibers in groups I, II, and III after 16 weeks. (
Ultrastructure of the regenerative axons
Many myelinated nerve fibers and unmyelinated nerve fibers were observed in the cross-sections of the PPN and distal L6 VR in group I (Fig. 5 E, F). Unmyelinated axons were usually shown in clusters. There were no obvious sign of Wallerian degeneration in the distal stump of L6 VR and PPN.
IVP measurement
When the left L4 VR proximal to coaptation in group I was stimulated, the IVP began to increase gradually with the stimulation. The mean maximum pressure elicited by the reflex pathway was 11.9±1.0 mm Hg, then subsequently the pressure dropped to baseline. The micturition of rats was also observed simultaneously. No obvious change of IVP was observed when the left L4 VR was stimulated using the same parameters in group II. The mean maximum IVP in group III was 22.5±1.3 mm Hg by the same stimulation on the left L6 VR (Fig. 6). There was a significant difference in IVP between group I and group II (p<0.05) and also between the group I and group III (p<0.05). However, the mean IVP in group I could reach 53% of that in group III.
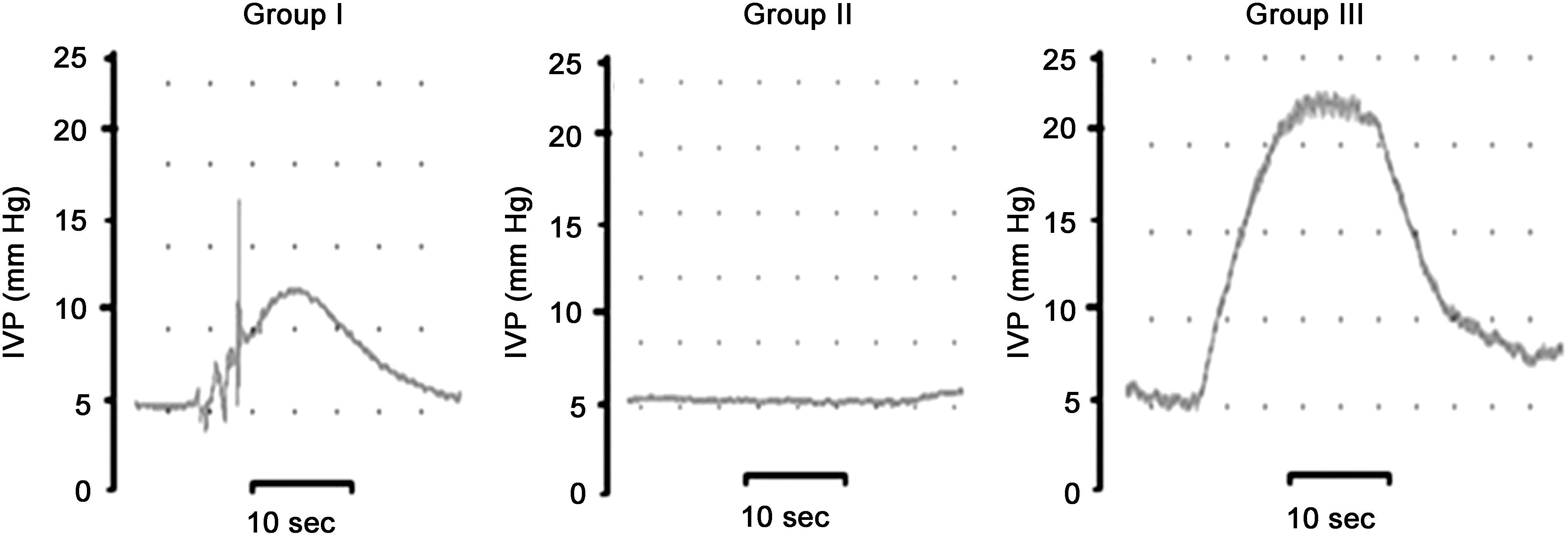
The graph of intravesical pressure measurement in three different groups. The intravesical pressures were recorded by using the same electrical stimulus (10 Hz, 0.5 mA) to excite the L4 VR proximal to coaptation in group I and the corresponding L4 VR in group II. In group III, the intravesical pressure was recorded by stimulating the left L6 VR. The recorded intravesical pressure in group I was more than half of that in group III (Group I, end-to-side neurorrhaphy group; Group II, “no repair” group; Group III, control group; IVP, intravesical pressure; VR, ventral root).
Wet weight and histological examination of the bilateral EDL muscles
There were no statistically significant differences between the wet weight of the left EDL muscle (0.1682±0.0036) and that of the right EDL muscle (0.1691±0.0036) in group I, whereas the right side of each rat in group I served as an internal control (Fig. 7A, p>0.05). The appearance of the muscle sections could reveal the degree of muscle atrophy where fibrous connective tissue increased instead of normal muscle fibers. The morphology of the muscle fibers on the left side appeared to be similar to that on the right side. No obvious fibrous connective tissue was found on either side in group I (Fig. 7B,C), which indicated that atrophy in the EDL muscle could be avoided.
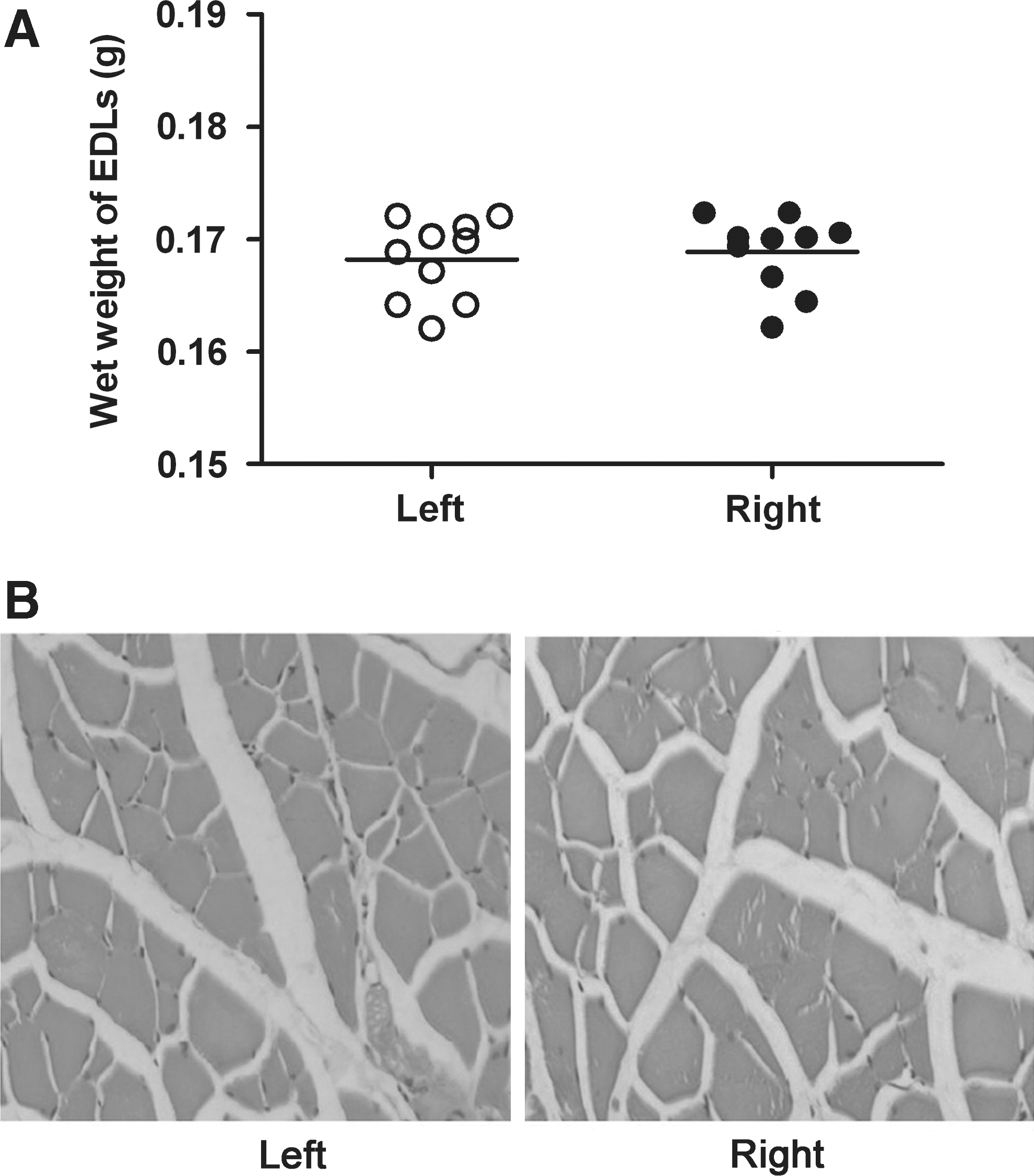
Comparisons of the wet weights of the bilateral extensor digitorum longus (EDL) muscles of rats in group I.
Discussion
Neurogenic bladder with voiding dysfunction caused by myelomeningocele and SCI is a major medical and social problem. Approximately >500,000 patients have neurogenic bladder after SCI (Lin et al., 2010), and 2500 babies are born with myelomeningocele with neurogenic voiding dysfunction each year in the United States (Tanagho and Schmidt, 1988). In the past decades, many efforts were made to treat bladder dysfunction. Xiao and Godec (1994) established a new somatic-autonomic reflex pathway for inducing micturition by performing intradural end-to-end neurorrhaphy of the left L4 VR to the left L6 VR, which innervated the bladder and external urethral sphincter in rats. Zheng and associates (2009) and Lin and associates (2010) reconstructed bladder function by intradural end-to-end microanastomosis of L5 VR or T12 VR to S2 VR with autogenous nerve grafting. However, this coaptation was performed inevitably at the expense of donor function, and sacrificed another nerve segment. Recently, Peters and Xiao (Peters et al., 2010; Xiao et al., 1999; Xiao et al., 2003; Xiao et al., 2005) demonstrated that the somatic-CNS-autonomic reflex pathway through end-to-end neurorrhaphy of L5 VR to S2/S3 VR could restore controllable voiding function of the bladder. However, this approach also inevitably sacrificed donor function and also aggravated the morbidity of the lower limbs in some patients to some extent.
End-to-side neurorrhaphy between autonomic nerve and somatic nerve for nerve repair in the peripheral nervous system is a novel and original concept that would attract much attention from neuroscientists and reconstructive surgeons. Our results first demonstrate that nerve regeneration is possible through end-to-side neurorrhaphy between autonomic nerve and somatic nerve.
For rats, the innervations of bladder and the external urethral sphincter are illustrated in Figure 1. PPN consisting of pure autonomic nerve fiber arises from L6 and S1 and synapses with postganglionic neurons in the MPG that innervate the bladder. In the present study, to exclude the interference of L6 DR, S1 VR, and S1 DR joining the PPN, we transected L6 DR, S1 VR, and S1 DR before performing the end-to-side neurorrhaphy between the distal stump of L6 VR (recipient nerve) and the intact L4 VR (donor nerve) in group I. Therefore, the PPN of group I only consists of regenerating nerve fibers. In group II, the “no repair” group, L6 VR, L6 DR, S1 VR, and S1 DR were transected. However, the distal stump of L6 VR was not coapted to the lateral face of the intact L4 VR. In group III, the control group, L6 DR, S1 VR and S1 DR were transected, but L6VR was kept intact. Therefore, the PPN of group III only consisted of autonomic nerve fibers deriving from L6VR. The most important significance of the design of group III was to provide the number of normal axons in the PPN, and to compare the number of regenerated axons with group I.
Our coaptation method not only differed markedly from nerve grafting but also was distinguished from the traditional end-to-side coaptation (Bontioti et al., 2005; Mennen et al., 2003; Viterbo et al., 1994b; Zhang and Fischer, 2002). We used autonomic nerve as recipient nerve by end-to-side neurorrhaphy in a helicoid fashion around the longitudinal axis of the donor somatic nerve. In this study, we established a new pathway by intradural end-to-side neurorrhaphy of the left L6 VR to the left L4 VR with the L4 VR intact along with the recipient nerve regeneration and reconstruction of bladder function by morphological examination of nerve fibers and IVP measurement. In the retrograde nerve tracing study, we injected the tracers into the SN and MPG respectively to verify that the connected reflex pathway was established between L4 VR and the L6 VR. SN is mainly composed of L3–L5 spinal nerves and MPG is the site where the autonomic nerve fibers of L6 nerve synapse with the parasympathetic postganglionic neuron that innervates the bladder. Each tracer should have obviously different modalities such as extraction wavelength, color, and different cellular distribution. FG is yellow and labels the cytoplasm and nucleolus whereas FB is blue and labels only the cytoplasm; the combination of FG and FB is yellow-blue under the ultraviolet filter set. Because of the abovementioned different features, FG or FB single-labeled neurons and FG-FB double-labeled neurons could be easily detected. We found that FG-FB single-labeled neurons and FG-FB double-labeled neurons had both appeared in the left ventral horn of L4 in group I, but that none was observed in L6 segment. The distribution and quantitation of all the retrogradely labeled neurons in the three groups are shown in Table 1. In our study, we just calculated the numbers of fluorescent labeled neurons in each segment of L3 to S1 using ultraviolet light under a fluorescence microscope. The more details of fluorescent labeled neurons including both the quantification of cell numbers and spatial distributions of three dimensions would be provided with the application of non-biased stereological approach (Burke et al., 2009). Current theories regarding the re-innervation mechanism after end-to-side neurorrhaphy are broadly categorized into two explanations (Jung et al., 2009). One is that the regenerated axons could originate from axons collaterally sprouting from intact donor nerve when double-labeled neurons are detected (Yamauchi et al., 2000), and the other is that newly formed axons also originate from terminal sprouting (Matsuda et al., 2005) because of slight injury to the donor nerve during the nerve suturing process when single-labeled neurons could be observed. The appearance of more double- labeled neurons in group I of our study implied that the main regeneration mechanism was collateral axons sprouting (Zhang et al., 1999).
The present study using IVP measurement method provided the functional evidence that the nervous signals could transmit to the bladder through the neurorrhaphy site and the regenerative nerve to initiate contraction, which could induce micturition in rats in group I. The IVP could reach more than half of normal value when the left L4 VR proximal to the coaptation was activated by electronic stimulation in group I. The results of retrograde nerve tracing and nerve fiber stimulation both demonstrated that the nerve fibers of L4 VR could regenerate and grow into the distal stump of L6VR through axons collaterally sprouting, and the regenerated nerve could re-innervate the target organ.
In our experiment, the results of morphological examination and ultrastructure of regenerative nerve indicated that the recipient nerve distal to coaptation could achieve prominent regeneration of axons after end-to-side neurorrhaphy. The results of semi-thin cross-sections and electronic micrograph of PPN were consistent. In group III, the number of myelinated axons of PPN only originated from normal L6 VR. Although the number of regenerative myelinated axons of PPN in group I was obvious less compared with that of PPN in group III, no obvious swelling of axons and nerve degeneration were observed in the PPN in group I. Also, the numbers of myelinated axons of L4 VR proximal to coaptation appeared to be similar to those of the distal stump of L4 VR, which indicated that there was no obvious injury to the donor nerve. And the results of wet weight and histological examination of EDL muscles also verified that there was no adverse effect to the target organ (EDL muscle) of donor nerve, which also indicated that the regeneration mechanism of axons in the donor L4 VR was collateral axonal sprouting.
Conclusion
In summary, from the abovementioned results in our experiment, we could reconstruct the bladder function without apparent injury to the donor nerve by performing intradural end-to-side neurorrhaphy of L6 VR (recipient nerve) to L4 VR (donor nerve). The obtained positive results of nerve regeneration may be based on the following elements, to our knowledge. First, the minimal injury to donor nerve during the nerve suturing process promoted the releasing of some nerve growth factors, and therefore regeneration occurred (Lutz et al., 2000; Yamauchi et al., 2000). Second, the angle at the coaptation site may have facilitated axons' growth into the recipient nerve (Yan et al., 2002). Third, the surface area of coaptation between the donor nerve and the recipient nerve was increased greatly by trimming the end of the distal stump of L6 VR diagonally, and using the helicoid neurorrhaphy (Yan et al., 2002). In addition to those elements described previously, other factors and variables that could also affect the regeneration of axons included neurotrophic factors, the number of Schwann cells, the timing of coaptation, and the type of re-innervated muscle.
Our neurorrhaphy method has the following advantages over traditional methods of bladder function reestablishment: 1) It does not sacrifice the donor nerve that is used to replace a nerve gap during nerve grafting. 2) Suturing the distal end of the recipient nerve to an intact healthy donor nerve in an end-to-side fashion promotes the recipient nerve function recovery and prevents target organ atrophy that could occur during the re-innervation period when a long nerve segment defect is present. 3) This procedure does not require rhizotomy of the donor nerve, which is important for protecting the lower limb function of the patients with myelomeningocele or partial SCI. The most important significance of our study is that it not only provides an alternative approach for bladder re-innervation without impairment of donor nerve function, but also initiates an original end-to-side neurorrhaphy method between autonomic and somatic nerves.
Therefore, the idea of achieving re-innervation of autonomic nerve and its target organs without sacrificing the donor somatic nerve function appears extremely attractive in peripheral nerve surgery. In this study, we focused only on the construction of end-to-side neurorrhaphy between autonomic nerve and somatic nerve and also evaluated the feasibility of this surgical strategy in re-innervation and functional recovery of the urinary bladder. Following this study, we will further investigate the effects of the end-to-side neurorrhaphy between autonomic nerve and somatic nerve in the treatment of bladder dysfunction after SCI in a rodent model. Furthermore, because of the different regeneration abilities of nerve fibers in humans and rats (Yamauchi et al., 2000) the epineurium of humans is obvious thicker than that of rats (Yamauchi et al., 2000; Zhao et al., 1997). The effects of nerve regeneration in humans after end-to-side neurorrhaphy needs to be further investigated. End-to-side neurorrhaphy in combination with some nerve growth factors to speed up axons' sprouting of donor nerves and their perforation should be explored further in the future.
Footnotes
Acknowledgments
We thank Professor Yang-Guan from the Department of Pathology, Tongji Medical College, Huazhong University of Science and Technology, for his support during ultrastructure sections preparation and analysis in the study. This study was supported by grants from the National Natural Science Foundation of China (NSFC) (30500535, and 30872567) and a grant from The National Basic Research Program of China (2003CB515304).
Author Disclosure Statement
No competing financial interests exist.