
Abstract
Brain-skull relative motion plays a pivotal role in the etiology of traumatic brain injury (TBI). The present study aims to assess brain-skull relative motion in quasistatic circumstances, and to correlate cortical regions with high motion amplitudes with sites prone to cerebral contusions. The study includes 30 healthy volunteers scanned using a clinical 3-T MR scanner in four different head positions. Through image processing and 3D model registration, pairwise comparisons were performed to calculate the brain shift between sagittal and coronal head positional change. Next, local brain deformation was evaluated by comparison between cortical and ventricular amplitudes. Finally, the influence of age, sex, and skull geometry on the cortical and ventricular motion was investigated. The results describe complex brain shift patterns, with high regional and inter-individual variations, outweighing age and sex patterns. Regions with maximum motion amplitudes were identified at the inferolateral aspects of the frontal and temporal lobes, congruent with predilection sites for contusions. No significant influences of age and sex on the cortical shift amplitudes were detected. The 3D cortical deviations varied from −7.86 mm to +5.71 mm for the sagittal head movement, and from −11.46 mm to +7.30 mm for head movement in the coronal plane, for a 95% confidence interval. The present study contributes to a better understanding of the mechanopathogenesis of frontotemporal contusions, and is useful for the optimization of finite-element head models and neurosurgical navigation procedures. Moreover, our results prove that in vivo MRI allows for accurate assessment of brain-skull relative motion in quasistatic conditions.
Introduction
T
The contact forces occurring during a head impact are the cause of most focal injuries (skull fractures and some brain contusions). However, for the development of injuries such as acute subdural hematoma (ASDH), diffuse axonal injury (DAI), and frontotemporal contusions, a specific damaging mechanism linked to inertial loading of the brain-skull complex can be described (Ommaya and Gennarelli, 1974). Significant accelerations and decelerations of the brain inside the skull lead to the development of non-uniform shear, compressive and tensile strains in the brain parenchyma and at the brain-skull interface (Ommaya and Gennarelli, 1974). Typical frontotemporal brain contusions are thought to result from the bouncing of the brain (i.e., compressive forces) on the intracranial surfaces (Kushner, 1998). These frontotemporal contusions were previously described as being even more clinically important and frequent than coup contusions (Hofman et al., 2001; Umile et al., 2002). The frontal and temporal lobes of the brain were previously described as predilection sites for contusion localization (Gurdjian and Gurdjian, 1976). Hence it is reasonable to speculate that higher-motion amplitudes will be observed at the inferolateral aspects of the frontal and temporal lobes. Besides the implications for brain contusion mechanisms, these brain-skull linear and rotational displacements may tear the bridging veins and can lead to ASDH.
Brain shift and deformation have been extensively studied using various techniques and theoretical as well as experimental models. These have included the application of a transparent calvarium to subhuman primates in the attempt to visualize brain motion and distortion directly (Gosch et al., 1969), implanting accelerometers into the brains of post-mortem human subjects (Al-Bsharat et al., 2002; Depreitere, 2004b; Hardy et al., 1997,2001), high-speed x-ray imaging of animals and human cadavers during impacts (Nusholtz et al., 1984), finite-element modelling (FEM; (Horgan and Gilchrist, 2003; Kleiven and Hardy, 2002), and magnetic resonance imaging (MRI) of human subjects undergoing voluntary motion (Bayly et al., 2005; Feng et al., 2010). Despite all these efforts, a direct relationship between brain motion and deformation and the occurrence of frontotemporal contusions, in terms of a clear tolerance criterion, has not yet been identified, mainly due to limitations in directly assessing brain motion and deformation in vivo using human volunteers. Hence, the aim of the present study was to assess the magnitudes of 3D brain shift and accompanying brain deformation in different head positions by applying in vivo MRI techniques, and to relate the resulting deviations to frontotemporal contusion mechanogenesis. The approach used in this study was previously described in our group by Depreitere and colleagues (2004a), and provides useful quantitative and qualitative evaluations of the inter-positional brain shift seen in human volunteers.
Methods
Thirty human volunteers (13 female and 17 male), with no known antecedents of neuropathological changes or head injuries, were recruited for the investigation of relative brain-skull motion in quasistatic conditions using MRI. The term “quasistatic” refers to a situation in which the volunteer is asked to remain motionless during scanning, and only gravitational forces are acting on the brain-skull complex, thus any inertial effect can be ignored. For the data processing, the volunteers were classified in three age groups of equal size: adolescents age <20 years (mean age 19.4 years), young adults 25–40 years old (mean age 31 years), and elderly adults >60 years old (mean age 68.3 years). Informed consent was obtained from each of the volunteers prior to scanning. The study was approved by the Ethical Committee of the University Hospitals Leuven, Belgium.
Magnetic resonance imaging technique
Complete sets of T2-weighted MR scans were obtained for each subject in four consecutive head positions (prone, supine, and left and right lateral), using a clinical 3-T Siemens Magnetom Trio. The optimal scanning protocol had an echo time (TE) of 531 milliseconds and a relaxation time (TR) of 3200 milliseconds, yielding a resolution of 1.1 pixels per millimeter, and a voxel size of ∼0.908×0.908×0.874 mm3. The volumetric sequence was acquired in the sagittal orientation in a 232×290 field of view, and 192 slices with the read-out direction head-feet, left-right, and anterior-posterior. The volunteers were prepared for scanning according to clinical standards. The duration of the MR session was ∼30 min per volunteer, with 4–5 min for each head position.
Image segmentation and volume reconstruction
The 3D volumes of the brain, skull, and lateral ventricles were manually segmented and reconstructed using commercially available software (Mimics® 13.1; Materialise NV, Leuven, Belgium). In order to minimize the errors resulting from manual operation, all segmentations were performed by a single experienced investigator.
After importing the DICOM images into Mimics, the voxels were differentiated between the brain, skull, and lateral ventricles (the bodies of the second and third ventricles) by thresholding the grey values in the native spatial orientation. During this phase several morphology operations (close, dilate, erode, add, or delete voxels) were applied to compensate image intensity inhomogeneities. Three-dimensional reconstructions of the brain, skull, and ventricles were calculated and exported as spatial coordinates of points representing nodes of triangular surfaces. An axial plane was defined to set the inferior limit of the brain by cutting the brainstem just inferior to the cerebellum.
Considering the brain tissue incompressibility experimentally described by previous researchers (Libertiaux et al., 2011), intra-individual inter-positional comparisons of the cerebral volumes were performed in order to evaluate the errors in the segmentation phase.
Rigid registration of the skulls
During this phase the skulls were pairwise aligned and registered in the same reference space (prone versus supine and left lateral versus right lateral) using 3-matic® 6.0 (Materialise NV). An intermediate “N-points” registration step was applied on some anthropometric points (e.g., nasion, glabella, vertex, basion, inion, opisthion, and opisthocranion; Aiello and Dean, 1990), in order to improve the accuracy of the global registration applied in the final step. The global registration procedure was performed using the Iterative Closest Point algorithm (Besl and McKay, 1992) to position and best fit the supine and prone skull orientations in the same reference space. The algorithm alternates the optimization of matches, by matching each model feature to its nearest neighbor. To increase the accuracy of the registration phase, we imposed a maximum allowed distance between matches of 0.05 mm. At the end of this phase, the parts positioned now in the same reference space (skull, brain, and ventricles) were exported as Standard Triangulation Language (STL) files. Using the STL format, the 3D geometry is described by an unstructured triangulated surface, defined using triangular unit normal and vertices in a 3D Cartesian coordinate system.
Calculation and visualization of 3D deviations
To assess the brain shift and deformation, pairwise intra-subject comparisons (prone-supine and left-right lateral) were performed using Focus Inspection® 8.4 (Metris NV, Leuven, Belgium). First, the brain-to-brain relative deviations were calculated (prone relative to supine, and left lateral relative to right lateral), so that regional and global amplitudes of brain shift were obtained. Next, to evaluate whether the global shift of the brain is represented mainly by a rigid body motion of the brain or by a combination of rigid body motion and local deformations, ventricle-to-ventricle pairwise comparisons were performed, following the same methodology. To qualitatively assess brain deformation, the deviations at the level of the ventricles, representative of the less deformable deep brain structures, were compared with the corresponding cortical deviations.
Brain shift was measured between corresponding point pairs from the compared geometries, as normal vectors to the triangulated surface. The large number of measurement points for the cortical (44,866±6252), as well as for the ventricular surfaces (4707±722), ensured a biofidelic representation of the 3D volumes and an accurate calculation of the 3D deviations. The positive values indicate an outward shift, while the negative values indicate an inward shift of the measured part (brain or ventricle) from the position considered as reference, describing a movement in the direction of gravity (e.g., prone relative to supine, and left lateral relative to right lateral). For better visualization, the 3D deviations are presented by means of color codes in Figure 1.
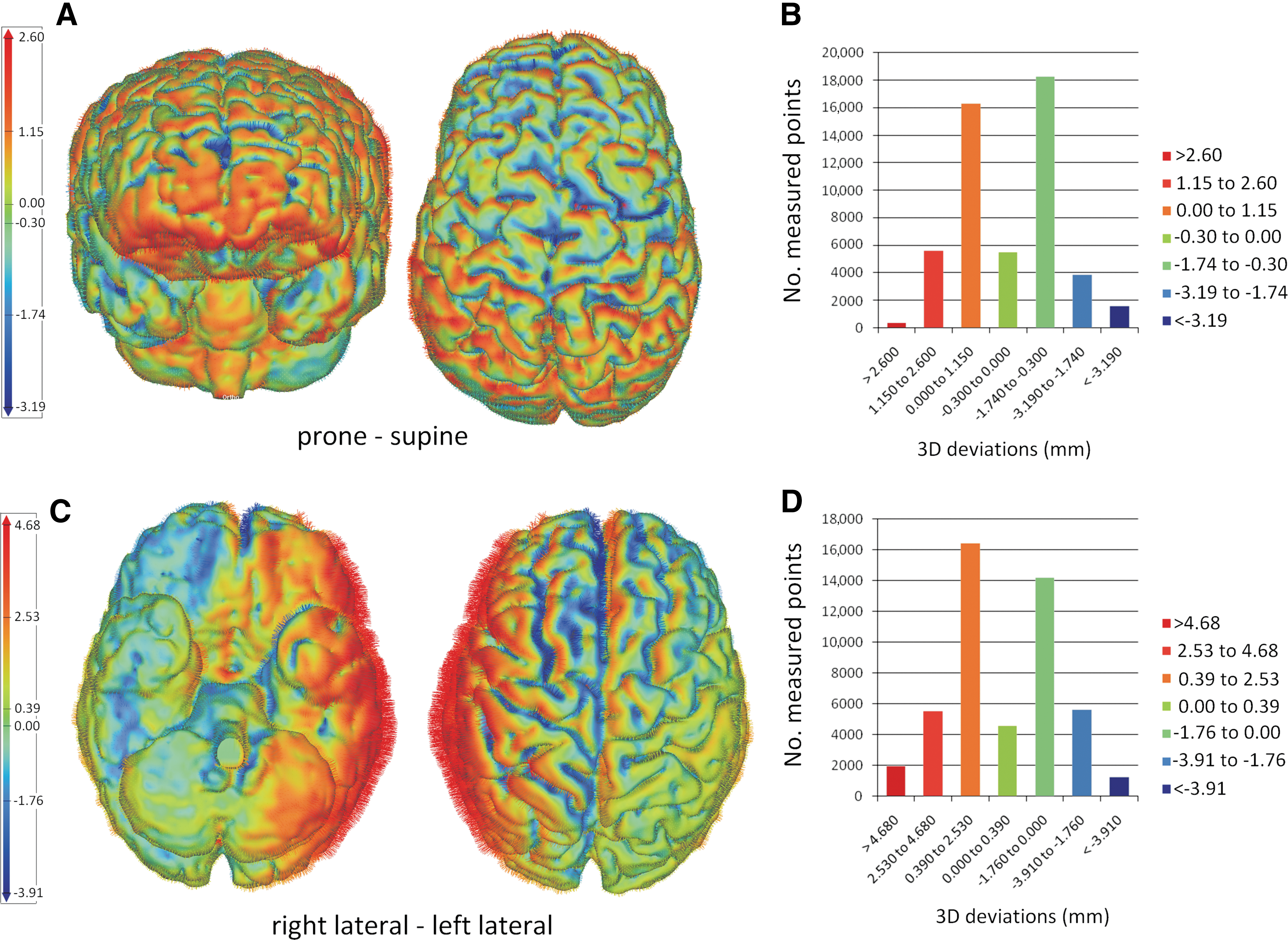
Typical 3D color plots used to visualize the distribution of the cortical displacement amplitudes for a sagittal head movement (frontal and top view of the cerebral hemispheres in
Statistical analysis
The relative effect of potential predictor variables (age, sex, anteroposterior skull diameter, and maximum cranial breadth) on the dependent variables (brain and ventricular shift, brain and ventricular volume) was assessed, controlling for possible interactions. Spearman rank correlation coefficients were calculated using IBM SPSS® 16.0 software. Simple and multiple linear regression analyses were performed to test the significance of the correlation between the cortical and ventricular shift and the predictor variables. The 3D deviations are presented as mean±standard deviation, for a 95% confidence interval (CI), and a significance level defined as p≤0.05. The study methodology is schematically illustrated in Figure 2.
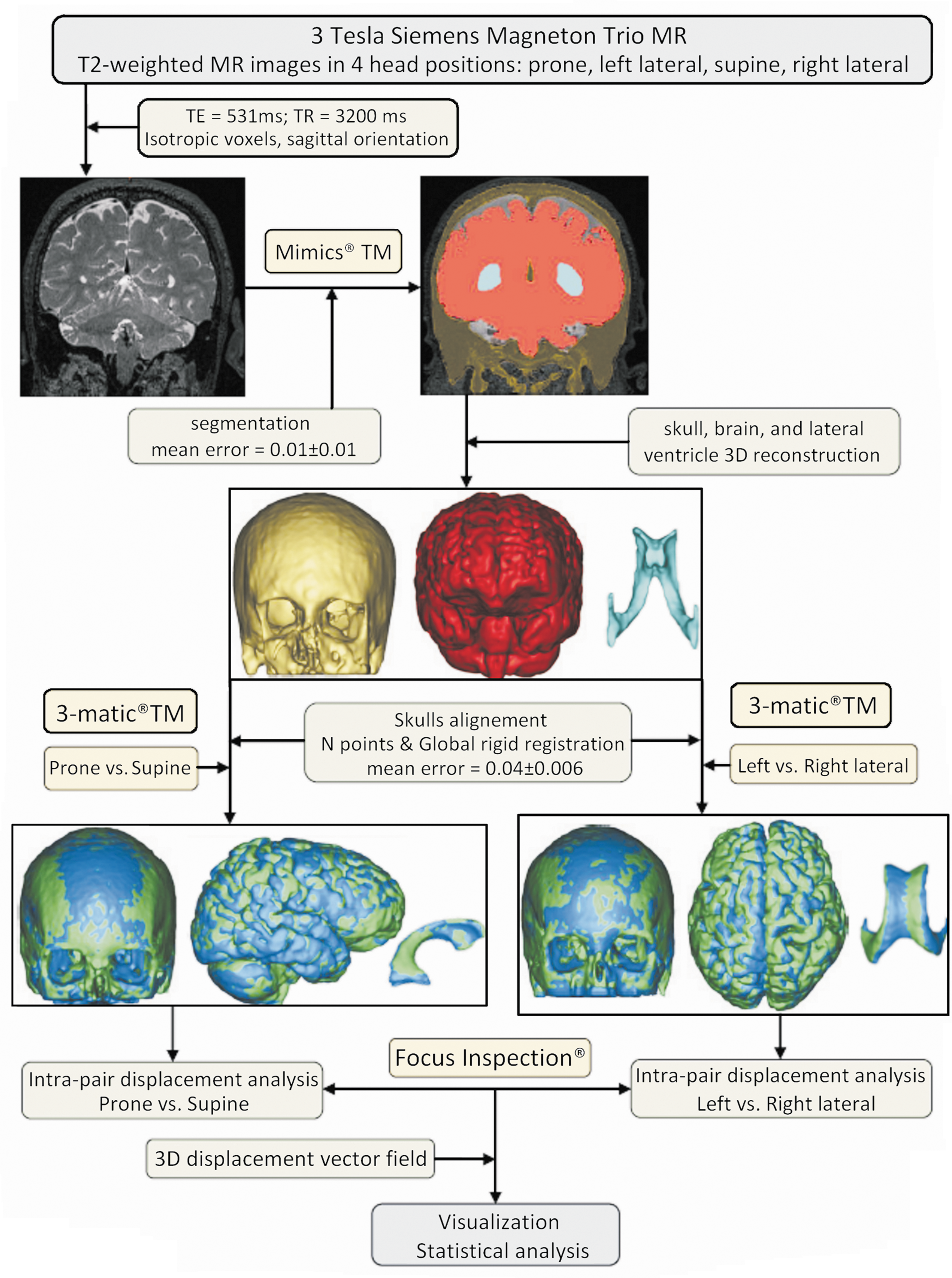
Schematic overview of the methodology used to assess the brain and ventricle positional shift, describing the intermediary steps. First, T2-weighted magnetic resonance (MR) scans of human volunteers were acquired using a 3-T MR scanner, in four head positions. During the segmentation phase, the gray values of the brain, skull, and ventricles are accurately separated by applying thresholding and various morphology operations (close, dilate, and erode), in the native space orientation. The resulted voxel masks were used to reconstruct the 3D volumes of the brain, skull, and ventricles, for each individual head position. Then the cortical and ventricular volumes were pair-wise repositioned (prone with supine and left lateral with right lateral) in a reference orientation space by rigidly registering the skulls. The cortical and ventricular 3D deviations were calculated after performing pair-wise comparisons of the brain and ventricle volumes in the same orientation space, and visualized by means of color plots. Color image is available online at
Results
Scans with insufficient image quality or with errors in the obliqueness information from the DICOM header were excluded from image and data processing. In total, successful comparisons were performed and analyzed for 26 subjects in sagittal head positional change, and for 24 subjects in coronal head positional change. A mean relative error to the reference cerebral volume of 0.01±0.01 SD (min=0.0001 and max=0.032) was calculated for the segmentation phase.
Inter-position brain 3D deviation analysis
Based on the color plots, regional descriptions of the brain movement for different lobes, individual gyri, and sulci were extracted individually for each volunteer and separately per sex and age.
For a head movement in the sagittal plane, the cortical deviations were distributed between 1.92 and 5.71 mm for the positive values, between −1.30 and −7.86 mm for the negative values, and with mean values ranging from −1.08 to −0.47, for a 95% CI. In 16 of 26 analyzed sagittal positional head changes, a high density of high positive amplitudes was observed at the level of the inferolateral and supraorbital aspects of the frontal lobes, frontal poles, inferolateral aspects of the temporal lobes, and the temporal poles. In addition, high negative amplitudes with parieto-occipital localization were described (Fig. 3).
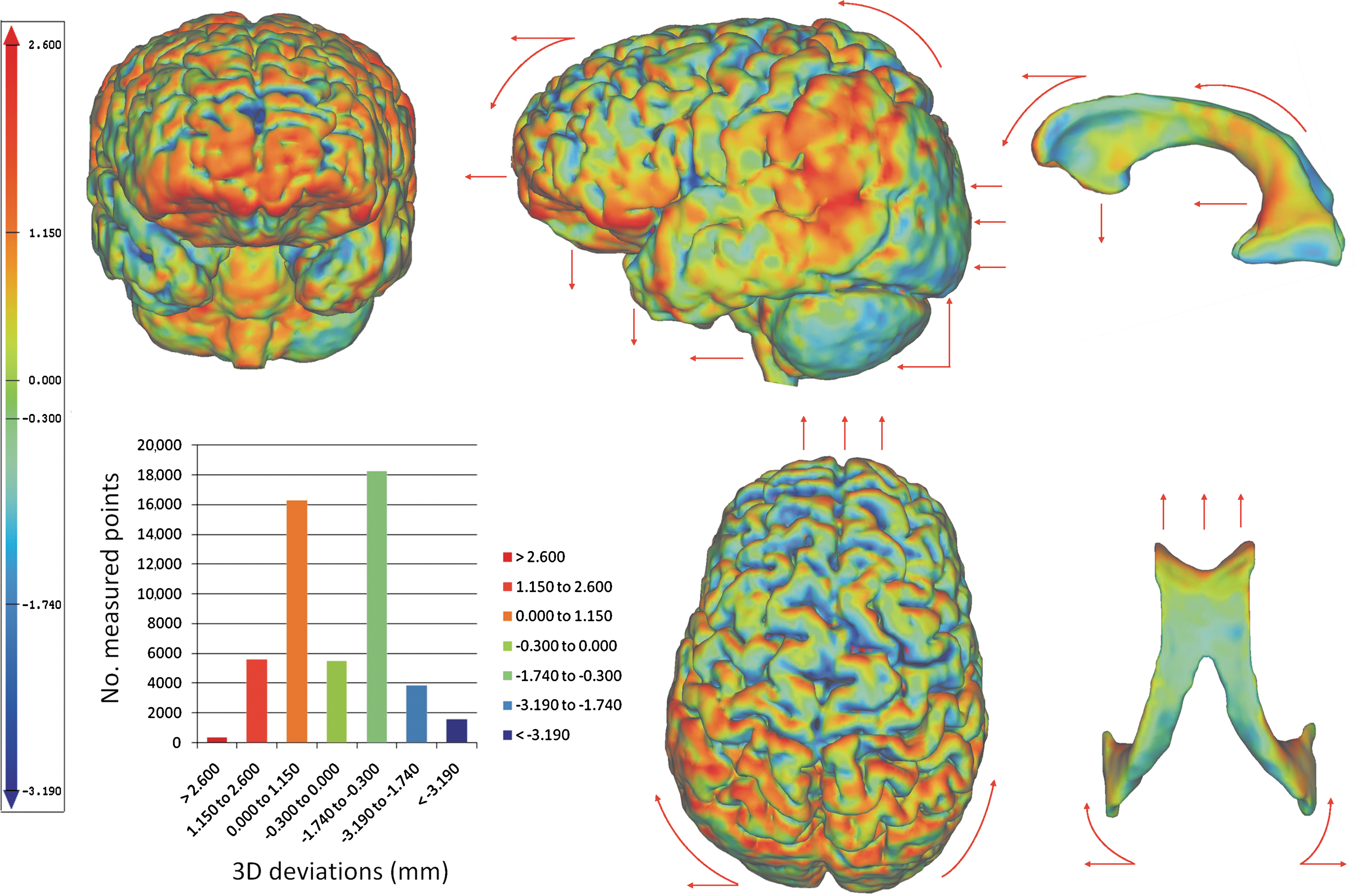
Typical distribution of the cortical (left figures) and ventricular (right figures) deviations resulted from a head movement in the sagittal plane, plotted by means of a color code. The red arrows indicate the direction of the whole-brain shift resulting from a prone-supine head positional change. In the color code, the red describes maximum positive amplitudes resulting from an outward brain deviation, and the blue corresponds to maximum negative amplitudes resulting from an inward brain deviation, compared with the reference position (supine). The cortical plots reveal large variations of the displacement amplitudes, with high amplitudes identified at the level of the inferior and lateral aspects of the frontal and temporal cerebral lobes, at the level of the deep borders of several cerebral frontal sulci (lateral sulcus, central sulcus, and pre- and post-central sulcus), and with a symmetrical distribution on the cerebral hemispheres. The head movement in the sagittal plane leads to a whole-brain translation in the same plane, and a brain rotation around the axial axis. The orbital surfaces of the frontal lobes shift down in the anterior cranial fossa; at the parieto-occipital level the brain rotates in the sagittal plane, and the inferior surfaces of the occipital lobes and cerebellar hemispheres move upward. The lateral ventricles follow the whole-brain shift, and due to brain internal deformation, the inferior and posterior horns of the ventricles are collapsing. The histogram on the left shows an example of the typical distribution of the 3D cortical deviations (mm) per number of measured points. Color image is available online at
For a head movement in the coronal plane, the cortical deviations were distributed between 1.45 and 8.35 mm for the positive values, between −1.68 and −11.46 mm for the negative values, and with mean values ranging from −2.08 to 0.51, for a 95% CI. In 19 of 24 analyzed cases of coronal head positional changes, higher amplitudes were found at the level of the inferolateral and supraorbital aspects of the frontal lobes, inferior temporal gyri, and temporal poles (Fig. 4).
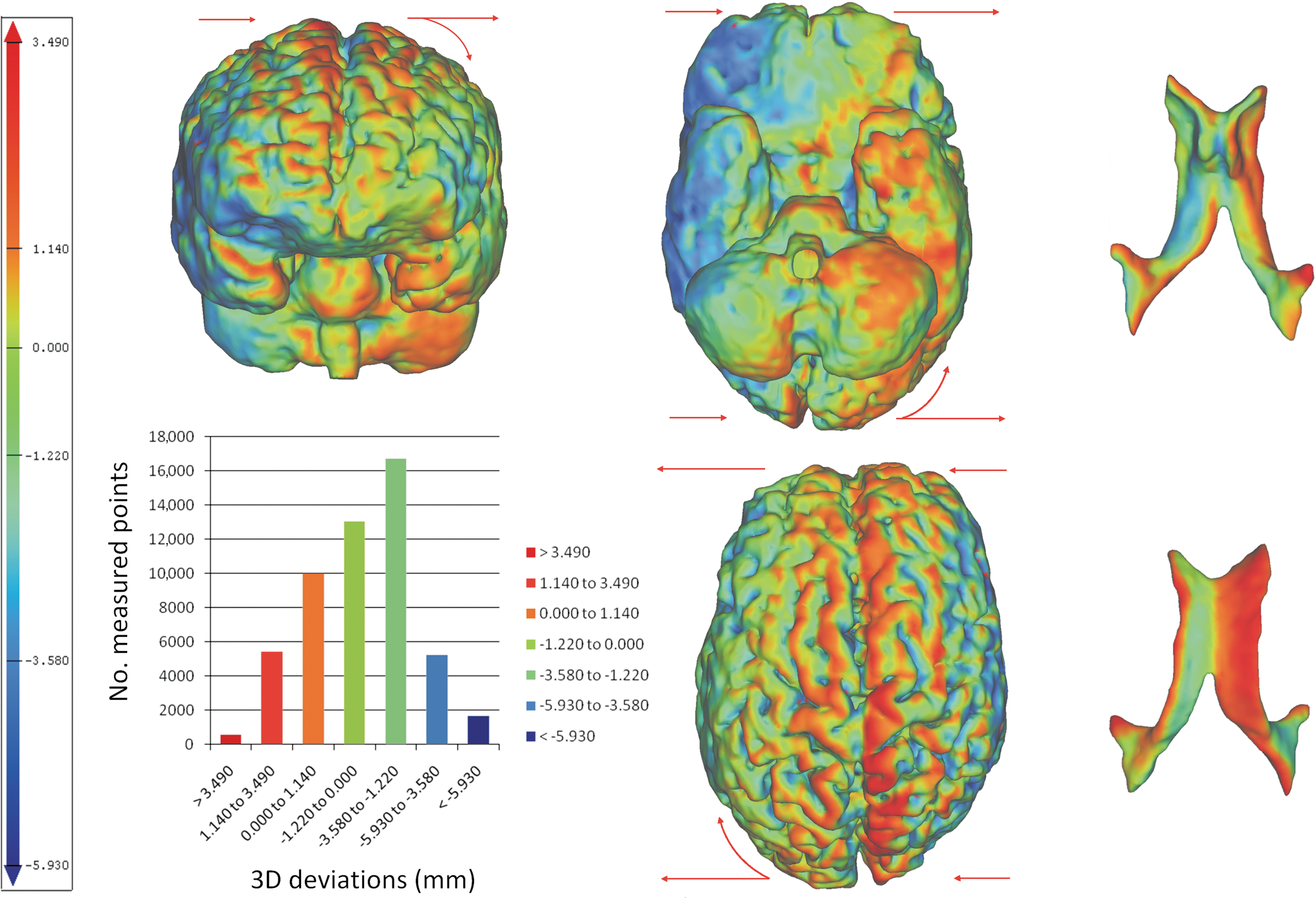
Typical distribution of the cortical (left figures) and ventricular (right figures) deviations resulting from a head movement in the coronal plane, plotted by means of a color code. The red arrows indicate the direction of the whole-brain shift resulting from a lateral head positional change. In the color code, the red describes maximum positive amplitudes resulting from an outward brain deviation, and the blue corresponds to maximum negative amplitudes resulting from an inward brain deviation, when compared with the reference position (right lateral). The cortical plots reveal large variations of the displacement amplitudes, with high amplitudes identified at the level of the inferior and lateral aspects of the frontal and temporal cerebral lobes, at the deep borders of the interhemispheric fissure, and with asymmetrical distribution, in the direction of gravity. A head positional change from left to right lateral leads to a whole-brain translation in the coronal plane, followed by slight rotations in the axial and coronal plane, behavior more pronounced at the level of the frontal and temporal cerebral lobes. The lateral ventricles follow the whole-brain shift, and due to internal brain deformation, the inferior and posterior horns of the ventricles are collapsing in the direction of the head movement. The histogram on the left shows an example of the typical distribution of the 3D cortical deviations (mm) per number of measured points. Color image is available online at
Isolated elevated amplitudes were revealed at the deep borders of several cerebral frontal sulci (lateral sulcus, central sulcus, pre and post central sulcus) for the head movement in sagittal plane (Fig. 3), and at the deep borders of the interhemispheric fissure, for the head movement in coronal plane (Fig. 4).
In Table 1 the cortical shift values for sagittal as well as for coronal head positional changes are presented as mean±SD, for a 95% confidence interval. The data reveal slightly larger amplitudes for the coronal head movement than for the head movement in the sagittal plane.
The data are presented as mean±SD for 95% CI for each volunteer and for age group (<20 years, 25–40 years, and > 60 years). The table includes the personal data of the volunteers and the measured cortical volume. The accuracy of the presented data is supported by the large number of measurement points describing the geometry of the cerebral hemispheres. The calculated deviations vary intra-individually between the two head positional changes considered for the study, and inter-individual for the same type of head positional change. The data presented in the table reveal no noteworthy age effect on the calculated brain shift amplitudes. Negative values depict inward movement and positive values depict outward movement.
CI, confidence interval; SD, standard deviation; MRI, magnetic resonance imaging; n=30, 17 males and 13 females.
For head movement in the sagittal plane, the maximum positive amplitudes of the cortical shift decreased with age (Spearman's rho=−0.47, p=0.02), but the age influence on the maximum negative amplitudes was not significant (Spearman's rho=0.29, p=0.15). Although a significant influence of age on the maximum positive amplitudes of the brain shift was found, the statistical analysis revealed no significant effect of age on the mean 3D cortical deviations (Spearman's rho=0.05, p=0.82; Fig. 5A). In the coronal head positional change, a moderate, though statistically significant, effect of age on the mean 3D cortical deviation was established (Spearman's rho=0.43, p=0.04; Fig. 5B).
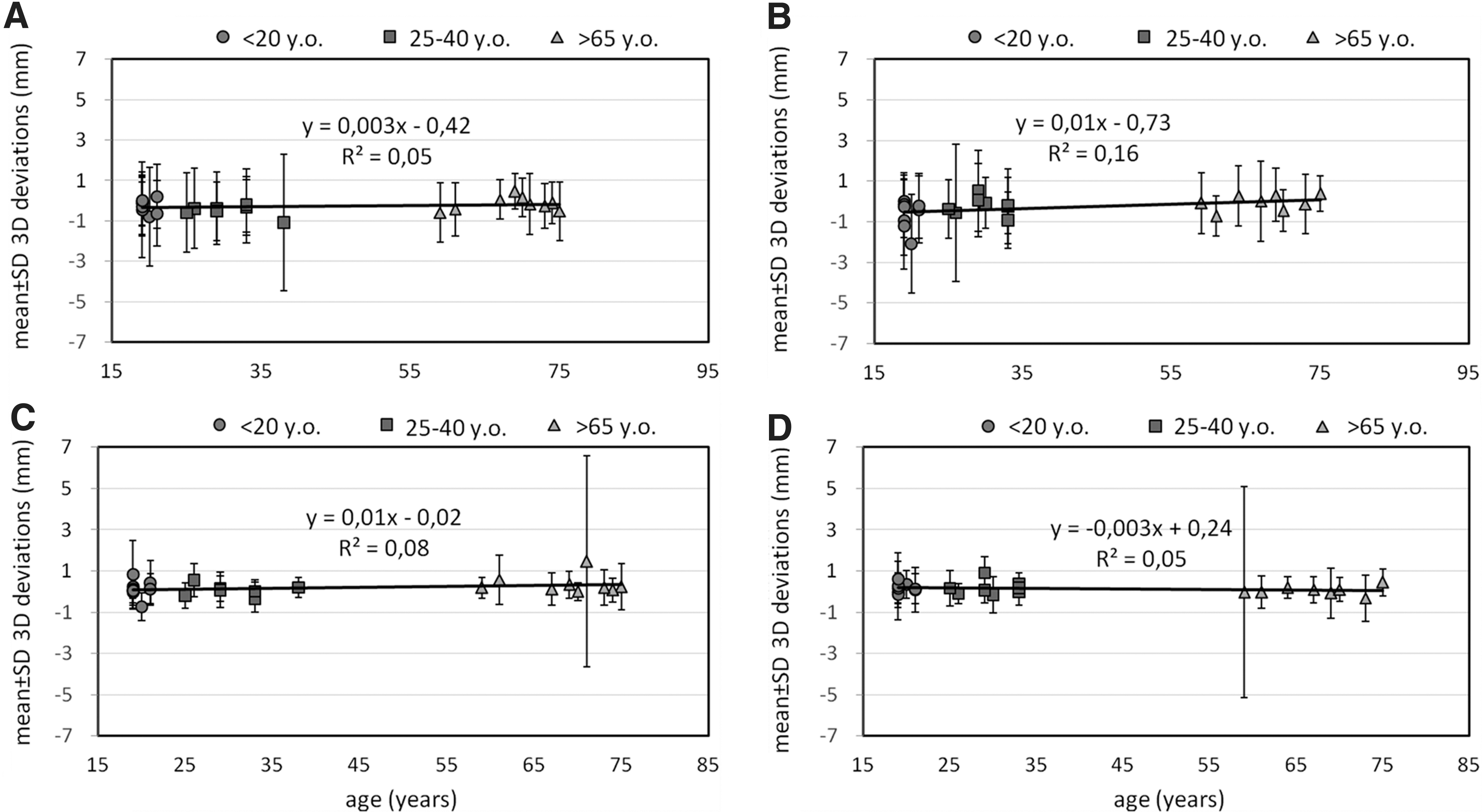
Graphical distribution of the 3D cortical and ventricular deviations function of age, separately for sagittal (total n=26; n=9 for those age <20 years, n=8 for those age 25–40 years, and n=9 for those age >65 years), and for coronal head movement (n=24; n=8 for those age <20 years, n=8 for those age 25–40 years, and n=8 for those age >65 years), and represented as mean±standard deviation. (
No significant influence of sex was found for the 3D cortical shift for a positional head change in the sagittal plane (Spearman's rho=0.22, p=0.29), or the head movement in a coronal plane (Spearman's rho=0.10, p=0.63).
Inter-position ventricle 3D deviation analysis
For a head movement in the sagittal plane, the ventricular deviations were distributed between 0.61 and 4.11 mm for the positive values, and between 0.01 and 0.83 mm for the negative values, with average amplitudes ranging from −0.72 to 0.83, for a 95% CI. For a head movement in the coronal plane, the ventricular deviations were distributed between 0.95 and 6.71 mm for the positive values, and between −0.97 and −4.90 mm for the negative values, with average amplitudes ranging from −0.32 to 0.90, for a 95% CI. The ventricular shift values in conjunction with the corresponding cortical deviations are summarized in Table 2. The color maps in Figures 3 and 4 show that the shift of the lateral ventricles followed the whole-brain motion. For a head movement in the sagittal plane, higher amplitudes of ventricular deviations were obtained at the level of the anterior horns of the ventricles, and at the inferior-posterior surface of the ventricular bodies. Moreover, under gravity, the temporal horns of the ventricles collapsed, suggesting brain deformation.
The data are presented as mean±SD for 95% CI for each volunteer and for age group (<20 years, 25–40 years, and > 60 years). The table includes the personal data of the volunteers and the measured cortical volume. The accuracy of the presented data is supported by the large number of measurement points describing the geometry of the cerebral hemispheres. The calculated deviations vary intra-individually between the two head positional changes considered for the study, and inter-individual for the same type of head positional change. The data presented in the table reveal no noteworthy age effect on the calculated brain shift amplitudes. Negative values depict inward movement and positive values depict outward movement.
CI, confidence interval; SD, standard deviation; MRI, magnetic resonance imaging; n=30, 17 males and 13 females.
Statistical analysis revealed no significant effect of age (two-tailed Spearman's rho=0.16, p=0.42 for sagittal head movement, Fig. 5C; Spearman's rho=−0.18, p=0.41 for coronal head movement, Fig. 5D), or of sex (Spearman's rho=0.00, p=1.00 for sagittal head movement; Spearman's rho=−0.30, p=0.16 for coronal head movement), on the 3D deviations observed at the ventricular level, for any of the inter-positional comparisons performed.
Brain and ventricle volumetric analysis
The brain volume calculation yielded a mean value of 1342.54±156.92 mL, with individual values ranging from a minimum of 1160.02 mL to a maximum of 1752.04 mL. In comparison with the group of adolescents, the mean brain volume was 4.57 % smaller for the group of young adults, and was 7.06% smaller for the group of elderly adults (Table 1). This indicates a significant reduction in brain volume with increasing age.
In contrast, the ventricular volume was significantly larger in the groups of young adults and the elderly, suggesting that at this level, age had an inverse effect. The ventricular volume varied between volunteers, ranging between a minimum of 14.04 mL and a maximum of 53.27 mL, yielding a mean value of 28.63±9.88 mL SD. In comparison with the group of adolescents, the mean ventricular volume was 4.9% larger for the group of young adults, and 29.17% larger for the group of elderly adults (Table 2).
The statistical analysis showed that the brain volume decreased with age (Spearman's rho=−0.36, p=0.06), but the relationship was inverted for the ventricular volume (Spearman's rho=0.39, p=0.04). However, no significant correlation between brain volume and mean brain shift for either type of head positional change was found (Spearman's rho=−0.11, p=0.60 for sagittal head movement, and Spearman's rho=−0.34, p=0.11 for coronal head movement). In addition, sex exerted no significant influence on the brain volume (Spearman's rho=−0.19, p=0.34), or on ventricular volume (Spearman's rho=0.09, p=0.97).
Skull geometry and influence on brain shift
The influence of skull geometry on positional brain shift was studied for two skull dimensions: skull anteroposterior diameter (APD) and maximum cranial breadth (MCB). In the group of male volunteers, the APD ranged from 176.3 mm to 205.34 mm, with a mean value of 188.85±8.26 mm; the MCB varied from 127.14 mm to 151.32 mm, with a mean value of 138.19±6.47 mm. For female volunteers, the APD values were distributed between 168.15 mm and 190.84 mm, with a mean value of 179.03±6.10 mm; the MCB varied from 128.26 mm to 142.08 mm, with a mean value of 135.54±4.21 mm.
The simple regression analysis showed no significant relationship between the cortical shift and MCB (Spearman's rho=−0.19, p=0.37 for sagittal head movement, and Spearman's rho=−0.29, p=0.16 for coronal head movement), or between cortical shift and APD for either sex (Spearman's rho=−0.10, p=0.62 for a head positional change in the sagittal plane, and Spearman's rho=−0.34, p=0.10 for a head positional change in the coronal plane; Fig. 6). However, a correlation was established between sex and APD (Spearman's rho=0.52, p=0.005); larger APDs were found in the group of male volunteers.
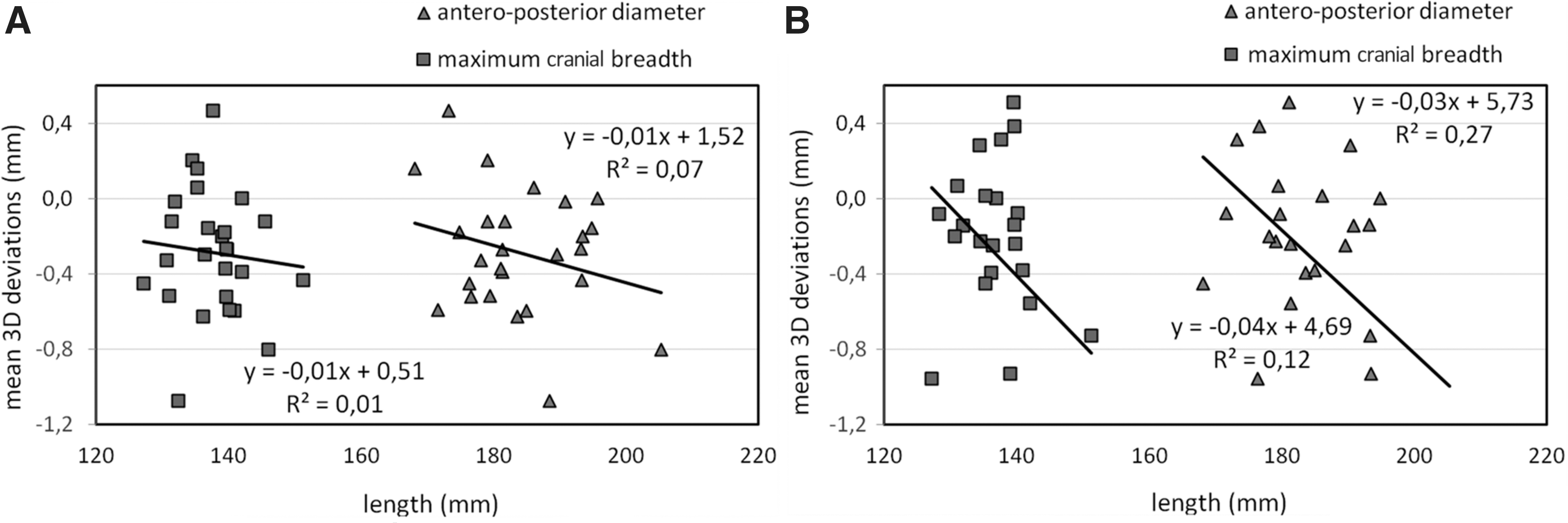
Graphical distribution of the 3D cortical deviations in relation to skull geometry, represented by two anthropometric dimensions: anteroposterior diameter (APD) and maximum cranial breadth (MCB). The graphs show no significant relationship between skull geometry and the calculated brain shift amplitudes, with less sensitivity for brain movement in a sagittal direction (
Multiple linear regression analysis
For head movement in the coronal plane, age and MCB were identified as independent predictors for the mean brain shift, explaining 35% of the variance seen in the brain motion amplitudes. The multiple regression model revealed that age was a significant predictor for the brain shift (coefficient=0.01, p=0.01), but MCB contributed more to the displacement variations (coefficient=−0.05, p=0.02) (Table 3).
Significant at the 0.01 level.
The resulting regression model shows that only age and MCB are statistically significant predictors for the brain shift resulting from a lateral head movement. The model explains 59% of the variation in the brain shift, indicating that for each extra year in age the brain shift will increase 0.01 mm, but will decrease 0.05 mm for each millimeter added to the skull MCB. In the model, age was the most significant predictor (p=0.01), but made the smallest contribution to the resulting cortical shift. In contrast, MCB, although less significant (p=0.02), had the largest contribution to the resulting brain shift.
For head movement in the sagittal plane, the multiple regression analysis failed to identify any independent relationship between brain shift and age, sex, MCB, or APD.
Discussion
Here we describe an MRI study that was undertaken to investigate the type and range of relative brain-skull movement in quasistatic circumstances in vivo. In addition, the influence of age, sex, and skull geometry (MCB and APD), on the calculated 3D cortical and ventricular deviations was evaluated. From the 30 human volunteers included in the study, the 3D brain shift was successfully processed and analyzed in 26 cases of head sagittal positional changes, and in 24 cases of coronal head positional changes.
Inter-position brain and ventricle 3D deviation analysis
While gravity may be a major driving force in brain movement, our results show that brain shift is utterly complex, with wide regional variations in the measured 3D cortical deviations. These local variations were mainly determined by changes between the moment of the gravitational forces of each measured point as the distances between the measured cortical point and the brain's axis of rotation were changing. Moreover, the brain shift maximum amplitude and distribution of the brain shift varied among volunteers and between head positions. For many subjects, regions where the direction of the displacement field was not aligned with the direction of gravity were observed, indicating that under gravitational load, the brain is also deforming. Therefore, we believe that the direction of brain shift is likely to be determined by a complex interaction between gravity, geometry of the contact surfaces, fluid pressure, and material properties.
Based on the 3D color maps we can suggest two theoretical scenarios that describe the overall brain-skull relative motion under gravitational loading. A positional change of the head in the sagittal plane will most probably lead to a brain translation in the same plane and a brain rotation around the axial axis. The orbital surfaces of the frontal lobes will shift down in the anterior cranial fossa; at the parieto-occipital level, the brain will rotate in the sagittal plane and the inferior surfaces of the occipital lobes and cerebellar hemispheres will move upward. A change of head position from left lateral to right lateral will most likely lead to a translation of the brain in the coronal plane, followed by slight rotations in the axial and coronal planes.
After a sagittal head movement, the brain shift was equally distributed along the interhemispheric fissure, mainly due to the skull's coronal symmetry. On the other hand, the brain motion resulting from a coronal head movement was no longer ruled by symmetry, leading to regional differences in brain shift due to local inhomogeneous strains. Additionally, for all four head positions, the distribution of the cortical deviations showed regions with elevated amplitudes in the areas of the inferolateral aspects of the frontal lobes and temporal poles. Lower amplitudes were observed at the level of the cerebellum and brainstem, in agreement with previous observations made by Ji and Margulies (2007). Slightly larger cortical deviations were calculated for coronal head movement in the group of young adults, suggesting higher mobility for the brain when the gravitational forces are acting in the coronal plane. However, as age increases, the differences between the 3D cortical deviations resulting from a head movement in the coronal plane or from a sagittal head movement are less important. Based on our results, it is reasonable to conclude that an important role in determining the brain shift is played by the tentorium and falx, which act as boundaries restricting the motion of the brain, and by the brainstem, which acts as an anchorage point around which the brain is rotating. The lateral ventricles followed the whole-brain shift and the pronounced movement observed for the frontal lobes; the maximum displacement was reached at the anterior horns of the lateral ventricles in both comparisons. However, the average ventricular displacement amplitudes appeared to be slightly smaller than the cortical amplitudes, suggesting the presence of a deformation component, which contributes to the whole-brain movement.
The results obtained in our study are in good agreement with previously published data by Hardy and associates (1997 and 2001) (∼3–5 mm brain displacement with local peak displacements of ±5 mm), and by Al-Bsharat and colleagues (2002) (∼3 mm brain-skull relative displacement). In addition, previous work by our group described a major craniocaudal motion component at the base of the frontal lobes and peak displacements of 3.2 mm at the vertex after sagittal head impacts (Depreitere et al., 2004b).
The present study shows that in the quasistatic situation, the brain shift seen was not solely a whole-brain rigid-body motion inside the skull. Important variations occurred between individual sulci and gyri due to internal inhomogeneous strains under gravitational loading. This is in contradiction to the conclusions of Zou and colleagues (2007), who found after performing low-velocity impacts on cadaver heads, that the brain displacement was described mainly in terms of rigid-body motion (4–5 mm and ±5 degrees), while the deformation was considered to account for most of the additional movement at higher speed.
It is necessary to mention that the present work only assesses brain shift under gravitational forces, which does not reflect real-life traumatic situations. There are strict limitations on work done with healthy human volunteers. It is likely that under high-impact loading conditions, the head's soft tissues exhibit strong nonlinear behavior. In addition, the skull deformation and resulting pressure waves might influence where and how the brain will deform. In contrast with the in vivo research, the use of cadaver models allows high-impact conditions to be tested. However, post-mortem changes in brain material behavior and in brain boundary conditions will undoubtedly influence the results obtained through cadaver research, representing a major limitation of this approach. Moreover, the efforts to re-establish in vivo–like boundary conditions by means of vascular and cerebrospinal re-pressurization techniques have been shown to produce a more rigid response of the head (Nusholtz et al., 1984). More recently, attempts to assess brain shift and deformation in low and mild impact conditions in vivo using tagged MR techniques (Bayly et al., 2005; Feng et al., 2010) found a general 2–3 mm brain-skull relative displacement and regions with maximum strains of up to 5%. While the reported data are in good agreement with the observations made in the current study, their approach only allows for a two-dimensional assessment of a behavior occurring in a three-dimensional space. Therefore we believe that by coupling both in vivo approaches, more realistic behavior of the brain during an impact can be obtained.
Influence of age, sex, and skull geometry on brain 3D deviations
It was previously suggested that age and sex influence cortical volume (Blatter et al., 1995), and the brain aging process (Scahill et al., 2003), and might further influence brain displacement (Ji et al., 2004). The influence of age on the cortical and ventricular volumes described in our study is in agreement with previously published reports. Yet the statistical analysis revealed no significant influence of sex on the cortical and ventricular volume. For a head movement in the sagittal plane, the multiple regression analysis revealed no independent associations between mean brain shift and age, sex, APD, or MCB. The insignificant relationship between age and brain displacement amplitudes contradicts our initial hypothesis that the brain will shift more as age increases due to a decrease in brain volume with age. One reason for this behavior may be a reduction of brain tissue elasticity and deformability with age. The present study strongly suggests that the deformation component plays an important role in whole-brain movement due to gravitational loadings. This observation is further supported in our study by the incongruence between ventricular and cortical displacements, which was less pronounced in the elderly age group. Moreover, if the aging process determines changes in the brain's local tissue density (Good et al., 2001), the brain will sink less into the cerebrospinal fluid (CSF) in order to follow basic principles of mechanics. Age did exert a significant negative effect on the maximum positive displacement in sagittal head movement (p=0.02), suggesting regional specificity in the relationship between age and brain motion. This observation is supported by the brain's anisotropic structure and morphology, as well as by the inhomogeneous nature of the brain's aging process (Gur et al., 2002).
When gravity acts in the coronal plane, the multiple regression model identified age and MCB as predictors for the brain shift, with age being the most significant variable (p=0.01), but with MCB having the largest contribution to the final output (p=0.02). This behavior could be explained by the relationship between the position of the center of mass and the vertical axis of the brainstem, which will contribute to the amplitude and to the distribution of the frontotemporal shift.
Finally, the large number of measured points covered any intra-individual biological bias, ensuring the validity of our data. However, the study sample size might be too small to cover inter-individual bias, both age- and sex-induced.
Contribution to cerebral contusion research
Over the years, many authors have considered the brain-skull relative motion as playing an important role in the pathogenesis of brain contusion. Yet the precise mechanisms are not completely understood. Based on the color maps provided by this study, regions with high displacement amplitudes can be identified at the level of the orbitofrontal and the inferolateral temporal cortex. These regions were previously identified as being predilection sites for contusions in a study of 192 contusions by Depreitere and colleagues (2004b). That study also showed a higher frequency of symmetrical orbitofrontal contusions in sagittal impacts, and a higher frequency of asymmetrical temporal contusions in lateral head impacts. Considering compressive stress as the causative mechanism of contusions at the tissue level, the data from the present study support the hypothesis that the bouncing of the frontal and temporal cortex surfaces on the skull base of the anterior and middle cranial fossa, secondary to inertial loading, and the higher amplitudes of cortical brain shift seen in these regions, may explain why these regions are predilection sites for contusions (Depreitere et al., 2004b).
Contribution to finite-element modeling techniques
Most contemporary head FEMs are validated against the cadaver data published by Hardy and associates (1997 and 2001), which refer only to the skull's center of gravity, and provides no information about local brain boundaries. Yet accurate FE simulations require a high-fidelity reproduction of the human head (subject-specific anatomy, material properties, and boundary conditions; Ho and Kleiven, 2009), as indicated by the high inter- and intra-individual variations of the brain displacement fields reported in this study. In addition, our study suggests that by assuming a fixed brain axis of symmetry, or by considering a non-sliding contact condition for the brain-CSF interface, the accuracy of the FEM will be affected. The data presented in this study will help to improve the accuracy of existing FEMs, and to develop more biofidelic head models, which can be used for accident reconstruction or to predict the risk of a specific brain injury.
Contribution to neuronavigation procedures
The gravitationally-induced brain shift amplitudes reported in the current study are sufficient to potentially cause clinical errors during surgical procedures requiring sub-millimeter accuracy, if head position differs between scanning and surgery. A shift of millimeters in the mid-brain can influence the results of stereotactic procedures (Miyagi et al., 2007), since the pre-operative planning is based on the assumption that anatomical structures do not shift between the scans acquired pre-operatively and the surgery. By using in vivo MRI on human volunteers, direct quantitative measurements of the brain shift and deformation can be obtained, and this can further increase the accuracy of computer-assisted neurosurgery.
Limitations of the current research
While the manual segmentation approach used in the current study allows for full user control, it is user-dependent, tedious, and time-consuming. In contrast, semi-automated approaches are generally faster and can provide subvoxel accuracy (Gelaude et al., 2008). Yet automated methods are also prone to errors by missing localized anatomical features, and by not functioning well in regions with poor contrast. We believe that by offering full control to the user, the manual approach can give more accurate results. This method is also supported in the literature by Bowers and associates (2008).
By being sensitive to the initial transformations, the rigid registration technique might be considered to be another limitation of the current research. However, we used an easy-to-implement global registration algorithm, which assumes a rapid nearest-neighbor search scheme, thus increasing the efficiency and accuracy of the registration computation.
Conclusions
This study presents a feasible approach to quantitatively and qualitatively assess the brain-skull relative displacement in vivo, using MRI techniques. The present approach allows for the determination of cortical displacement fields, and for the observation of 3D brain and ventricle deviations. In addition, the comparison of the 3D cortical deviations with the amplitudes obtained at the level of the lateral ventricles allows for a qualitative assessment of the internal brain deformation under gravitational loading. We were able to describe patterns for the brain-skull relative motion, and to identify brain regions with high mobility, suggesting intra- and inter-individual variability. Yet we did not find clear evidence of sex or age influence on the brain and ventricle motion in quasistatic conditions. Thus a broader study might be necessary to draw any final conclusions about these issues. The results contribute to our understanding of frontotemporal contusion mechanopathogenesis, and can be utilized for the improvement of navigational accuracy for intracranial surgery. The obtained data are directly applicable in the validation and testing of finite element head models used to investigate the mechanisms behind head trauma, allowing more realistic models to be developed.
Footnotes
Acknowledgments
This research was performed within the framework of the FWO Vlaanderen (Funds for Scientific Research from Flanders, Belgium), Project Levenslijn G.0001.08: “Improving head protection for child and adult pedal cyclists: approach based on lesion-specific mechanogenesis.” We would like to acknowledge Prof. Stefan Sunaert and Mr. Stefan Ghysels from the Radiology Section, University Hospital Gasthuisberg, for technical assistance offered during the MRI sessions.
Author Disclosure Statement
No competing financial interests exist.