
Abstract
The inhibitory environment of the injured spinal cord is an obstacle to functional recovery and axonal regeneration in adult mammals. We had previously shown that injection of adeno-associated virus (AAV) encoding the L1 cell adhesion molecule (AAV-L1) at the time of acute thoracic compression injury of adult mice promotes locomotor recovery, which is associated with ameliorated astrogliosis and improved axonal regeneration in the lumbar spinal cord. In the present study, we investigated whether delayed injection of AAV-L1, chondroitinase ABC (Chase), or the combination of the two agents into the mouse spinal cord 3 weeks after injury would also lead to improved recovery. The Basso Mouse Scale showed enhanced locomotor recovery 12 weeks after application of the agents in all treatment groups compared to the control group that was injected with AAV encoding green fluorescent protein (AAV-GFP). Investigation of hindlimb function using single-frame motion analysis revealed, however, that L1 overexpression, but not injection of Chase, improved voluntary movements without body weight support, whereas injection of Chase, but not L1 overexpression, enhanced body weight support during stepping. Mice with the combined application of AAV-L1 and Chase showed improvement in both parameters. Enhanced motor recovery after combined application correlated with increased densities of cholinergic and GABAergic terminals at motoneuronal cell bodies, and of lamina-specific glutamatergic sensory afferents 15 weeks after injury, indicating enhanced synaptic rearrangements in the lumbar spinal cord below the lesion site. These findings suggest that L1 overexpression combined with Chase application may contribute to the treatment of sub-chronic spinal cord injury.
Introduction
S
An alternate way to improve the outcome of SCI is to overexpress beneficial molecules, such as the neural cell adhesion molecule L1 that promotes neuritogenesis and axonal targeting, neuronal migration and survival, as well as myelination and synaptic plasticity (Buhusi et al., 2008; Demyanenko et al., 2011; Guan and Maness, 2010; Irintchev and Schachner, 2011; Maness and Schachner, 2007). Abundant evidence has accumulated showing that L1 expression levels and axon regeneration in the CNS are correlated. Retinal ganglion cells, nigrostriatal and cholinergic neostriatal neurons express relatively high levels of L1, and are able to regenerate axons along a peripheral nerve graft without upregulation of L1 expression (Becker et al., 2001; Jung et al., 1997; Woolhead et al., 1998). Neurons in the thalamic reticular nucleus express low levels of L1 and CHL1, but are capable of upregulating these proteins, and also successfully regrow into the graft (Chaisuksunt et al., 2000a; Zhang et al., 1995). Cerebellar deep nucleus neurons and precerebellar brainstem neurons also show a substantial capacity for axon regeneration, accompanied by increased expression of L1 (Chaisuksunt et al., 2000b). Zhang and associates (2005) demonstrated that Purkinje cells, which do not express or upregulate the expression of L1, its close homologue CHL1, or growth-associated protein-43 (GAP-43), and do not regenerate into grafts (Chaisuksunt et al., 2000b), show remarkable axon regrowth when both L1 and GAP-43 are transgenically expressed in these cells. In accord with the latter finding are observations in zebrafish showing that the expression of L1.1, a homolog of mammalian L1, is increased after SCI in successfully regenerating descending axons, but not in ascending projections that fail to regenerate (Becker et al., 2005), and that morpholino-mediated reduction of L1.1 expression in descending neurons leads to deficient axon regrowth and poor functional recovery (Becker et al., 2004).
In accord with the regeneration-promoting properties of L1 are the results of several studies demonstrating that exogenously-supplied L1 is beneficial for recovery after acute SCI. The experimental paradigms include delivery of L1 recombinant protein to the injured rat spinal cord (Roonprapunt et al., 2003), and injections into the spinal cord of injured mice of L1-overexpressing embryonic stem cells (Chen et al., 2005), neural aggregates (Cui et al., 2011), Schwann cells (Lavdas et al., 2010), and radial glia (Xu et al., 2011). Most importantly, overexpression of L1 in the injured mouse spinal cord via adeno-associated virus (AAV) resulted in improved locomotor recovery, regrowth/sprouting of severed axons, and reduced reactive astrogliosis (Chen et al., 2007). In the present study, we tried to take advantage of the regeneration-conducive effects of AAV-L1 overexpression, combined with the disinhibition effects of Chase, to achieve functional improvement after application at a late time point (3 weeks) after SCI in adult C57BL/6 inbred mice.
We report that AAV-L1, Chase, and most prominently their combination, promote motor recovery when injected 3 weeks after compression of the spinal cord using an electromechanical device, an incomplete lesion model allowing one to achieve reproducible graded levels of motor disability in the mouse (Apostolova et al., 2006). We further explored some mechanisms of this beneficial effect of L1 and Chase by analyzing synaptic rearrangements of cholinergic and GABAergic terminals at motoneuronal cell bodies, and glutamatergic terminals in several areas of the lumbar spinal cord (i.e., caudal to the lesion site), including laminae VII and IX and Clarke's column. Our results show that L1 is effective not only immediately, but also 3 weeks after SCI. Additionally, we confirm and expand the work of others by showing that the application of Chase alone or in conjunction with another recovery-enhancing measure promotes motor recovery and synaptic plasticity at a delayed time point after SCI. Importantly, our results indicate that L1 and Chase affect different aspects of locomotion and synaptic rearrangements, thus leading to additive effects on motor recovery.
Methods
Animal surgery
Female 3-month-old C57BL/6 mice were anesthetized by intraperitoneal injection of a ketamine-xylazine mixture (100 mg/kg ketamine, Ketanest; Parke-Davis/Pfizer, Karlsruhe, Germany; 10 mg/kg xylazine, Rompun; Bayer, Leverkusen, Germany), and surgery was performed as described by Lee and colleagues (Lee et al., 2009). In brief, laminectomy was performed at the T9 vertebra (i.e., the T10–T11 spinal cord segment), and the spinal cord was compressed using fine watchmaker's forceps driven by an electromagnetic device (Curtis et al., 1993). The force and duration of compression were regulated by an electrical current flow controller. Compression was performed at the highest force for 1 sec (100% compression according to the operational definition of Curtis and associates; Curtis et al., 1993). This type of lesion in the mouse, although leading to severe motor disability, is incomplete as shown by retrograde labeling of cells projecting beyond the lesion site to the distal (lumbar) spinal cord, including neurons in the cervical spinal cord, reticular formation, raphe nuclei, red nucleus, and motor cortex (Apostolova et al., 2006).
At 3 weeks after injury, the mice were anesthetized with the same dose of the ketamine-xylazine mixture as that used for SCI, and the compressed lesion site of the spinal cord was disclosed by removing the connective tissue at the laminectomized spinal segment. Injection was performed at the caudal end of the lesion site, as identified as a dark brown area, using a 1-μL Hamilton syringe in the same manner as in our previous AAV-L1 study (Chen et al., 2007). Two groups of mice were injected with the AAV-5 vectors constructed with the murine cytomegalovirus early (mCVM) promoter to express L1 (AAV-L1) or green flourescent protein (AAV-GFP; 3×107 transducing units in 1 μL), and the other two groups were injected with chondroitinase ABC (10 U/mL; Seikagaku Kogyo Ltd., Tokyo, Japan), alone or in combination with an additional AAV-L1 injection. Schematic illustration of the treatment regimens is shown in Figure 1. Previous work has shown that in both acutely-injured (Chen et al., 2007), and non-injured mouse spinal cords (unpublished observations), a single injection of AAV-GFP at a low thoracic level leads to early (before 1 week post-infection) and long-lasting (persisting at high levels for at least 5 weeks) transgene expression. The transgene is expressed in cells and cell processes, including axons and axon terminals, in the cervical and lumbar spinal cord for distances of at least 10 mm from the injection site. Most frequently transfected are neurons (48%), including motoneurons and interneurons in the ventral horn, followed by astrocytes (29%) and oligodendrocytes (3%).
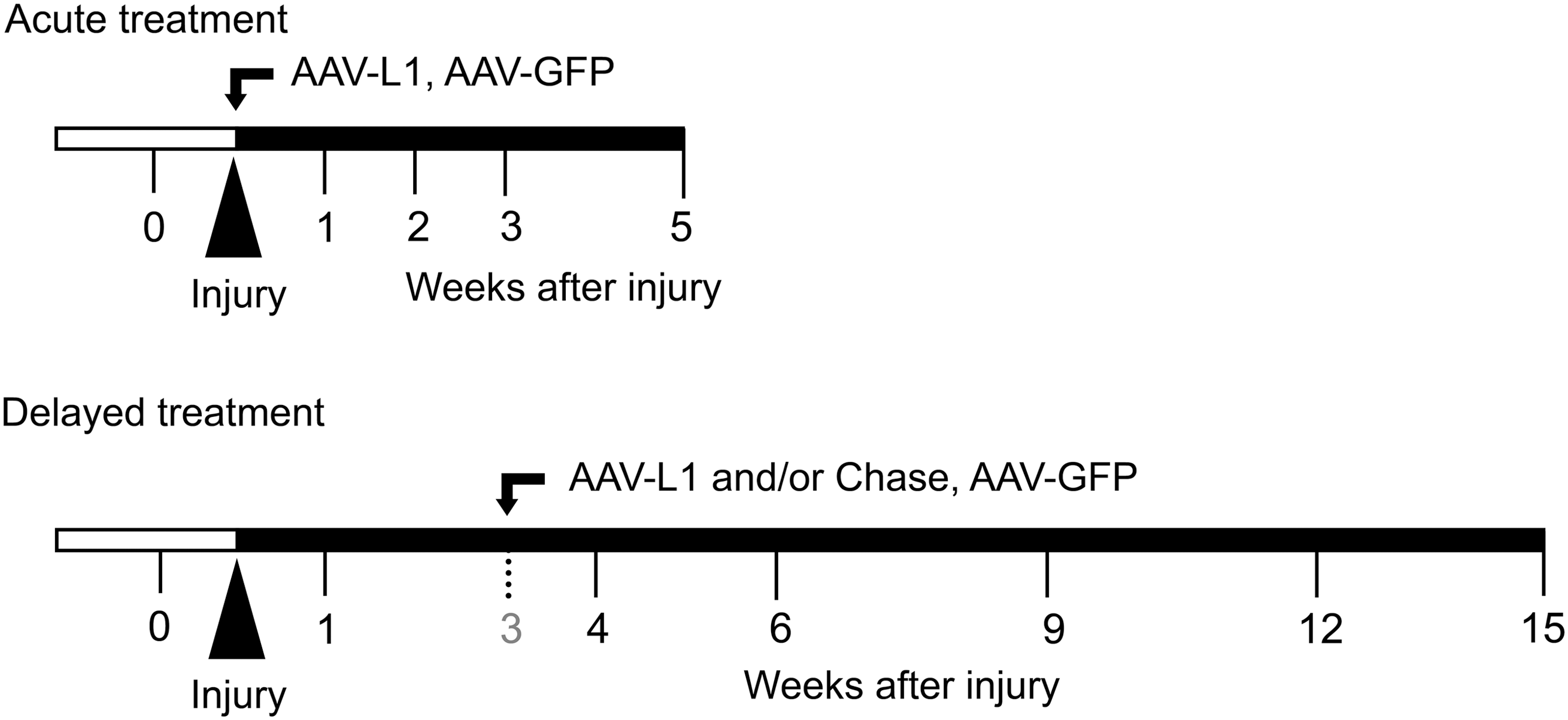
Schematic representation of the experimental design. In a previous study (Chen et al., 2007), an L1-expressing adeno-associated virus (AAV-L1) construct was injected into the lesion site immediately after compression spinal cord injury (SCI) of adult female C57BL/6 mice (“Acute treatment”), and locomotor functions were assessed and compared with mice treated with an AAV-GFP (green fluorescent protein) vector (as negative control) over a 5-week recovery period. In the present study, young adult female C57BL/6 mice were treated with a single intraspinal injection of AAV-L1, chondroitinase ABC (Chase), or a combination of these agents (AAV-L1 plus Chase) 3 weeks after injury (“Delayed treatment”). Motor function assessment was performed prior to SCI (0), and at 1, 4, 6, 9, and 12 weeks after injury, and compared with mice that had received control treatment (AAV-GFP). After the 15-week observation period, spinal cords of the mice from all four treatment groups (AAV-L1, Chase, AAV-L1+Chase, and AAV-GFP) were sampled and subjected to immunohistological analyses.
Analyses of motor function
The animals were subjected to evaluation of motor function before SCI (day 0), and at 1, 4, 6, 9, 12, and 15 weeks after injury (Fig. 1). Individuals performing the functional assessments, as well as morphological analyses, were unaware of the identity of the treatment groups to which the animals belonged. Open-field locomotion was assessed by a mouse rating scale, the Basso Mouse Scale (BMS; Basso et al., 2006). In addition, motor performance was examined using video-based single-frame motion analysis, an established approach for the evaluation of motor recovery after spinal cord lesions in mice (Apostolova et al., 2006; Chen et al., 2007,2010; Cui et al., 2011; Devanathan et al., 2010; Jakovcevski et al., 2007; Lee et al., 2009; Mehanna et al., 2010; Semler et al., 2011; Xu et al., 2011). Motor behavior of individual animals was recorded while the mice were performing two tasks: the beam-walking test and the pencil-grabbing test. The videos were recorded using a high-speed camera (A602fc; Basler AG, Ahrensburg, Germany), and SIMI Motion 7 software (SIMI Reality Motion Systems, Unterschleissheim, Germany) at 100 frames per second. Two parameters, the foot-stepping angle and the rump-height index, were measured using video frames of beam walking. Selected frames in which the animals were seen in defined phases of locomotion (Apostolova et al., 2006) were used for measurements performed with Image Tool 2.0 software (University of Texas Health Science Center, San Antonio, TX; free software available at
The second parameter, the rump-height index, was estimated from the recordings used for measurements of the foot-stepping angle. The parameter is defined as height of the rump (i.e., the vertical distance from the dorsal aspect of the animal's tail base to the beam, normalized to the thickness of the beam measured along the same vertical line). For each animal and trial, at least three frames in which the rump height was maximal were used for measurements. The rump-height index is a numerical estimate of the ability to support body weight. This ability requires coordination in different joints of both hind extremities, and is influenced by various factors such as stepping art (plantar versus dorsal) and muscle strength.
A third parameter, the limb extension:flexion ratio, was evaluated from video recordings of voluntary movements of the mice performed during the pencil-grabbing test. An intact mouse, when held by its tail and allowed to grasp a pencil with its forepaws, tries to catch the object with its hindpaws and performs cycling flexion–extension movements with the hindlimbs. For the spinal cord injury paradigm, left- and right-side view videos were recorded for each animal. The extension and flexion length of the extremity (distance from the most distal midpoint of the paw to a fixed, discernible point on the animal's body, such as the tail base), were measured for at least three extension–flexion cycles per animal and time point. Mean values for the two extremities from one animal were averaged. The extension:flexion ratio is a numerical estimate of the animal's ability to initiate and perform voluntary, non-weight-bearing movements. Such movements require connectivity of the spinal cord to supraspinal motor control centers but in the form evaluated here, no coordination or precision.
Tissue processing and immunohistochemistry
After the last evaluation of motor function, the mice were anesthetized with a 16% solution of sodium pentobarbital (Narcoren, 5 μL/g body weight; Merial, Hallbergmoos, Germany), and transcardially perfused with fixative consisting of 4% formaldehyde and 0.1% CaCl2 in 0.1 M cacodylate buffer (pH 7.3), for 15 min at room temperature. After perfusion, the spinal cords were left in situ for 2 h at room temperature, after which they were dissected out and post-fixed overnight (18–22 h) at 4°C in the solution used for perfusion. The lumbar enlargements of the spinal cords were then excised and immersed in 15% sucrose solution in 0.1 M cacodylate buffer (pH 7.3) for 24 h at 4°C, embedded in embedding medium, and frozen for 2 min by immersion in 2-methyl-butane (isopentane) precooled to −80°C. Serial transverse sections (25-μm thick) were cut on a cryostat (CM3050; Leica, Nussloch, Germany). The sections were collected on SuperFrost Plus glass slides (Roth, Karlsruhe, Germany) in a standard sequence so that six sections 250 μm apart were present on each slide.
Immunohistochemistry was performed according to a published protocol (Irintchev et al., 2005). The following commercially available antibodies were used at optimal dilutions: goat anti-choline acetyltransferase (ChAT, 1:100; Chemicon, Hofheim, Germany), mouse anti-vesicular GABA transporter (VGAT, 1:1000; Synaptic Systems, Göttingen, Germany), mouse anti-vesicular glutamate transporter 1 (VGLUT1, 1:500; Synaptic Systems), guinea pig anti-5-hydroxytryptamine transporter (5-HT, 1:1000; Millipore, Schwalbach, Germany), rabbit anti-glial fibrillary acidic protein (GFAP, 1:500; Dako, Hamburg, Germany), and rabbit anti-NG2 proteoglycan (1:100; Chemicon). Water-bath antigen unmasking was performed in 0.01 M sodium citrate solution (pH 9.0) for 30 min at 80°C (Jiao et al., 1999). Non-specific binding was blocked using 5% normal serum from the species in which the secondary antibody was produced, diluted in phosphate-buffered saline (PBS, pH 7.3), and supplemented with 0.2% Triton X-100 and 0.02% sodium azide for 1 h at room temperature. Incubation with the primary antibody diluted in PBS containing 0.5% lambda-carrageenan (Sigma-Aldrich, St. Louis, MO), and 0.02% sodium azide, was performed for 3 days at 4°C. After washing in PBS (three times for 15 min each at room temperature), the appropriate Cy3- and Cy2-conjugated secondary antibody, diluted 1:200 in PBS/carrageenan solution, was applied for 2 h at room temperature. Following a subsequent wash in PBS, cell nuclei were stained for 10 min at room temperature with bis-benzimide solution (Hoechst dye 33258, 5 μg/mL in PBS; Sigma-Aldrich). Finally, the sections were washed again, mounted in anti-fading medium (Fluoromount G; Southern Biotechnology Associates via Biozol, Eching, Germany), and stored in the dark at 4°C. For double-labeling with ChAT and VGAT antibodies, the protocol was similar except that the primary antibodies were mixed together for section incubation. Photographic documentation was made with an LSM 510 confocal microscope (Zeiss, Oberkochen, Germany).
Quantification of motoneuron soma size and perisomatic terminals
Estimations of soma areas and perisomatic terminals were performed as described previously (Apostolova et al., 2006; Jakovcevski et al., 2007; Lee et al., 2009). Transverse spinal cord sections stained for ChAT and VGAT, which contained motoneurons caudal to the lesion scar, were selected for the analyses. Stacks of 1-μm-thick images were obtained with an LSM 510 confocal microscope using a 63× oil-immersion objective and digital resolution of 1024×1024 pixels. Four sections 250 μm apart were analyzed per animal and staining procedure to assure that motoneurons located both close and remote to the lesion scar were sampled. One image per motoneuron at the level of the largest cell body cross-sectional area was used to measure soma area, perimeter, and number of perisomatic terminals. Motoneurons were identified by ChAT staining, and by their large, pale nuclei (bis-benzimide staining), large soma size, and position in the ventral horn of the spinal cord. The sample size ranged between 20 and 30 motoneurons per animal. Areas and perimeters were measured using ImageJ software (National Institutes of Health [NIH], Bethesda, MD). Linear density was calculated as the number of perisomatic terminals per unit length. Mean values of individual animals were used to calculate group mean values.
Quantification of excitatory afferent input and expression of GFAP and NG2
To investigate densities of VGLUT1+ synaptic terminals, transverse lumbar spinal cord sections caudal to the lesion were used. Digital images were obtained on an LSM 510 confocal microscope using a 63× oil-immersion objective and a digital resolution of 1024×1024 pixels. The confocal images were converted to grey scale to adjust threshold consistently based on the histogram shape of the optimal color intensity using ImageJ software. Control sections stained immunohistochemically without secondary antibody were used as a black reference of grey values. VGLUT1+ synaptic terminals were detected using particle analysis. Criteria for identification of synaptic terminals were: objects with size > 2 μm2 and circularity 0–1. The number and perimeter of synaptic terminals were sampled in the Clarke's column, lamina VII, and lamina IX of each section. Density was calculated as the number of VGLUT1+ synaptic terminals per unit area. The values for left and right side were averaged for each section. Six sections 250 μm apart were analyzed per animal, and the mean values from individual animals were used to calculate group mean values. Expression of GFAP and NG2 was quantified similarly to VGLUT1, but instead of particle counts, we estimated the area of the immunopositive structures and normalized them to the total image area. In addition to GFAP and NG2, we pursued analysis of serotonergic (5-HT-positive) axons in the distal spinal cord, which are projections that we had previously shown to be increased by acute AAV-L1 application in parasagittal spinal cord sections (Chen et al., 2007). Unfortunately this attempt failed, since the staining pattern of the transverse spinal cord sections was variable enough to preclude meaningful quantifications.
Statistical analysis
All numerical data are presented as group mean values with standard error of the mean (SEM). Parametric tests were used for comparisons as indicated in the text and figure legends. Analyses were performed using the SYSTAT 9 software package (SPSS, Chicago, IL). The threshold value for acceptance of differences was 5%.
Results
Delayed AAV-L1 and/or Chase injections improve motor recovery after SCI
To investigate the effects of delayed overexpression of L1 and/or application of Chase after SCI, we performed a longitudinal evaluation of motor functional recovery (Fig. 1). In contrast to injection of AAV-L1 or, as a negative control AAV-GFP, immediately after injury as in previous experiments (Chen et al., 2007), adult female mice were injected with AAV-L1, Chase, AAV-L1 and Chase in combination (Comb), or AAV-GFP, 3 weeks after injury. The BMS, a scoring system for open-field locomotion of mice, indicated severe disability of walking in all groups 1 week after compression SCI (Fig. 2A). Injection of AAV-GFP at 3 weeks after SCI in the control group did not affect spontaneous recovery seen without injection of the virus (Chen et al., 2010; Lee et al., 2009), and the score increased in a linear fashion between 1 and 15 weeks (Fig. 2A). However, it must be noted that functional improvements achieved at 9–12 weeks, as estimated by the BMS and the parameters described below, was statistically insignificant compared with 6 weeks after injury (p>0.05 by ANOVA with Tukey's post-hoc test). The groups of mice injected with AAV-L1, Chase, and Comb, showed significantly better recovery of BMS scores at 12 and 15 weeks after injury compared to controls. Significant differences among the treatment groups (AAV-L1, Chase, and Comb) were not found up to 12 weeks after injection. The conclusion from these observations is that all three treatments improved functional recovery to a similar degree compared with control mice.
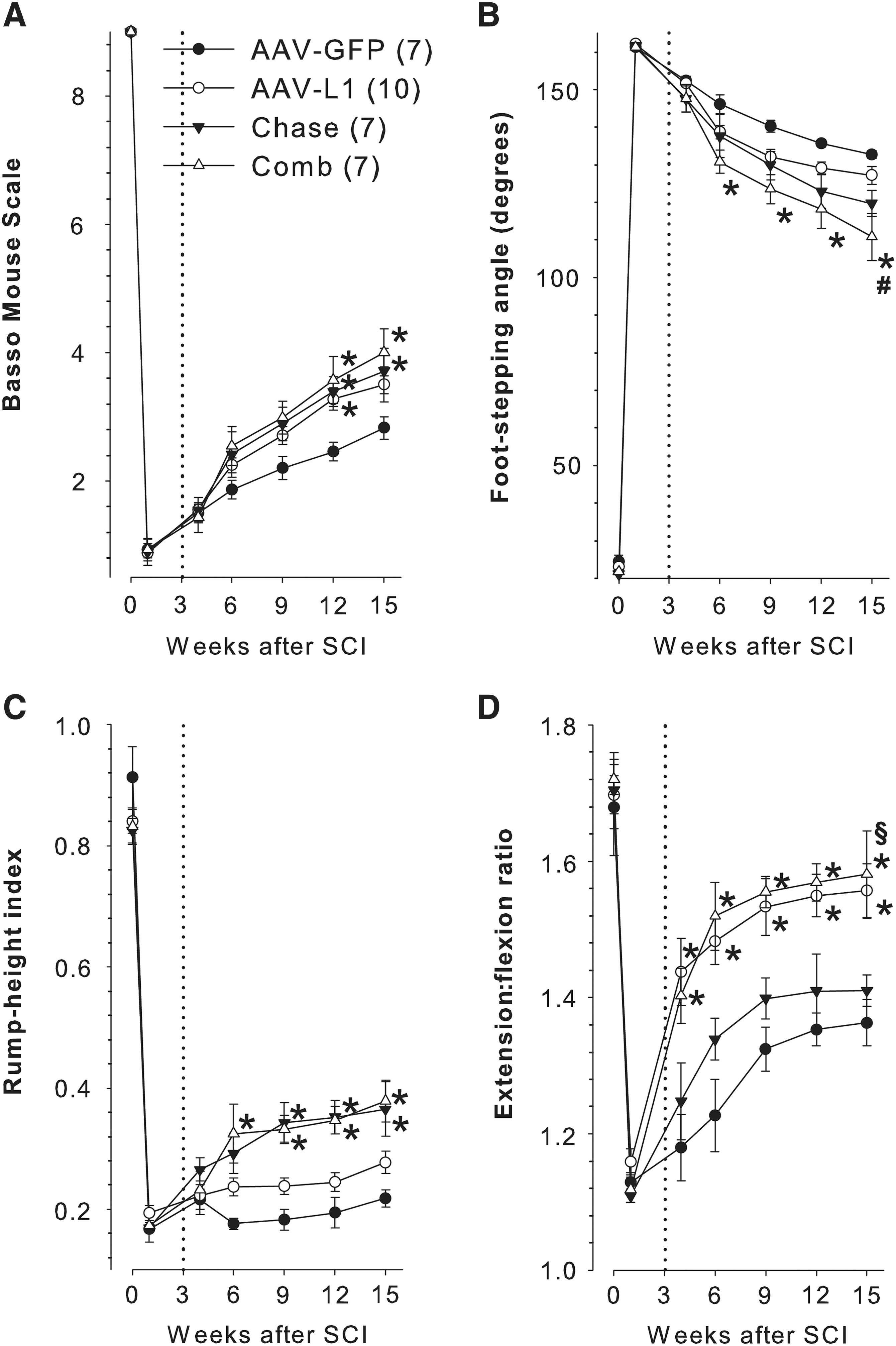
Improved functional recovery after compression spinal cord injury (SCI) and delayed injections of adeno-associated virus encoding L1 (AAV-L1) and/or chondroitinase ABC (Chase). Panels
We also performed a more elaborate motion analysis based on measurements using single video frames to examine recovery of distinct motor abilities as estimated by three different parameters (Apostolova et al., 2006). One of the parameters used was the foot-stepping angle, an objective measure of stepping ability (plantar or dorsal stepping) during walking, which typically correlates well with locomotor score (BMS and BBB) values (Apostolova et al., 2006; Chen et al., 2007,2010; Cui et al., 2011; Devanathan et al., 2010; Jakovcevski et al., 2007; Lee et al., 2009; Mehanna et al., 2010; Semler et al., 2011; Xu et al., 2011). The pattern of recovery of the foot-stepping angle in control AAV-GFP-treated mice (Fig. 2B) was similar to that of the BMS score evaluated in the same mice (Fig. 2A). In contrast to BMS, only Comb, but not AAV-L1- or Chase-treated mice, showed better recovery 6–15 weeks after injury compared with control mice (Fig. 2B). These findings indicate that significant improvement of the stepping abilities after SCI in mice is achieved only after the combined AAV-L1 and Chase treatment.
The second parameter analyzed was the rump-height index, a measure of hindlimb ability to support body weight during walking. This ability was severely impaired in all groups at 1 week after SCI, and barely improved during the following 14 weeks in mice treated with AAV-GFP or AAV-L1 (Fig. 2C). Compared with these two groups, the rump-height index was significantly higher 9–15 weeks after SCI in mice treated with Chase or the combined AAV-L1 and Chase treatment (Fig. 2C). Thus, injection of Chase leads to improvement of body weight support, a more complex function compared with stepping analysis by the foot-stepping angle (Apostolova et al., 2006; Semler et al., 2011), while L1 overexpression showed no effect on this aspect of locomotion.
We also analyzed the ability of the mice to perform limb movements without body weight support as estimated by the extension:flexion ratio. This parameter measures the change in limb length during reaching pursuits in the air (pencil-grabbing test), which is a more complex behavior compared with aspects of walking like stepping (foot-stepping angle) and body weight support (rump-height index; Apostolova et al., 2006), and is not estimated by the BMS. In all animal groups, limb movements were severely restricted 1 week after SCI compared with their pre-operative abilities (Fig. 2D). In control AAV-GFP mice, the extension:flexion ratio increased in a linear fashion up to the ninth week, reaching plateau values of about 40% of baseline (pre-operative) values at 12–15 weeks after SCI (Fig. 2D). Treatment with Chase did not significantly improve recovery of this parameter. In contrast, recovery of this parameter was accelerated and reached significantly higher values in AVV-L1- and Comb-treated mice 6–15 weeks after SCI (Fig. 2D). These results indicate that delayed application of AAV-L1, but not Chase, after SCI leads to improvement of the ability to perform limb movements without body weight support.
The conclusions from the functional analyses are that AAV-L1 and Chase positively affect recovery of different aspects of limb motion, and thus have an additive effect on the overall motor performance of SCI mice. At the same time, pain behaviors like over-grooming and licking or biting of the hind paws characteristic of spinal cord-injured BALB/C, but not C57BL/6 mice, the strain used here (Kerr and David, 2007), were not observed in either control or L1- and/or Chase-treated animals. These observations suggest that our treatments did not lead to exaggerated abnormal changes in sensation like allodynia or hyperalgesia (Hoschouer et al., 2010).
Rearrangement of synaptic terminals at the somata of motoneurons
To understand some of the cellular mechanisms contributing to the improved locomotor outcome of mice treated with AAV-L1 and/or Chase 3 weeks after SCI, we examined the synaptic terminals at motoneuron cell bodies caudal to the lesion site in the lumbar spinal cord 15 weeks after SCI in the mice used in the motor tests. We first analyzed numbers of choline acetyltransferase-expressing (ChAT+) cholinergic terminals (Fig. 3A), known to form C-type synapses on motoneurons (Davidoff and Irintchev, 1986; Hellstrom et al., 2003), based on our observations that attenuated loss of C-terminals after SCI correlates with better motor recovery of individual animals (Apostolova et al., 2006; Jakovcevski et al., 2007; Lee et al., 2009). The linear density (number per unit length) of perisomatic ChAT+ terminals 15 weeks after SCI was higher by 81% and 101% in groups treated with AAV-L1, alone or in combination with Chase, respectively, than in the AAV-GFP-injected control group (Fig. 3B).

Enhanced rearrangements of perisomatic terminals after adeno-associated virus encoding L1 (AAV-L1) treatment. Shown are motoneuronal soma size and densities of synaptic terminals at motoneuron cell bodies 15 weeks after spinal cord injury. Representative confocal images show ChAT+ and VGAT+ synaptic terminals around motoneuron cell bodies in the ventral horn identified by ChAT immunostaining (
Additionally, we estimated the numbers of vesicular GABA transporter-immunopositive (VGAT+) terminals, which reflect numbers of inhibitory (GABAergic and glycinergic) synapses at the cell bodies of motoneurons identified by ChAT labeling (Fig. 3A). In groups treated with AAV-L1 and the combination of AAV-L1 with Chase, the density of perisomatic inhibitory puncta was higher by 41% and 52%, respectively, compared to the control group (Fig. 3B). In the course of spontaneous recovery from SCI in C57BL/6J mice, the number of VGAT+ inhibitory terminals was no different from that of non-injured mice 6 weeks after SCI (Apostolova et al., 2006; Jakovcevski et al., 2007), but was significantly increased at 12 weeks after injury compared to non-injured animals (Lee et al., 2009). It is likely that this late increase in the density of VGAT+ terminals at motoneuron cell bodies contributes to the functional recovery observed late after SCI.
Another parameter to consider in functional recovery is the size of motoneuron cell bodies, as reflected by the cell body areas. There was no difference between the treatments (Fig. 3B). Thus, enhanced density of ChAT+ and VGAT+ synaptic terminals is due to an increased number of synapses, rather than a reduction of cell body area. These observations indicate that AAV-L1, but not Chase treatment, significantly influences rearrangements of modulatory (cholinergic), and inhibitory (GABAergic), inputs to motoneuronal cell bodies.
Excitatory afferent inputs to the spinal cord after injury
Preservation and/or sprouting of primary afferent inputs to the spinal cord is critical for functional recovery of hindlimb locomotion after SCI (Lavrov et al., 2008). We have demonstrated that area-specific alteration of excitatory afferent inputs in the lumbar spinal cord was related to the excitability of motoneuron pool and motor recovery after injury (Chen et al., 2010; Mehanna et al., 2010). Thus in the mice used in all previous assays we examined vesicular glutamate transporter 1-immunopositive (VGLUT1+) synaptic terminals known to be derived from medium- to large-sized neurons in the dorsal root ganglia, which conduct mechanoceptive and proprioceptive information to the spinal cord (Brumovsky et al., 2007). Three specific areas where the VGLUT1+ terminals appear prominently were selected for analysis (Chen et al., 2010): lamina VII adjacent to the central canal, where the last-order interneurons that innervate motoneurons in the spinal cord are located (Fig. 4A); the ventral horn region, lamina IX, where proprioceptive and mechanoreceptors are connected directly with motoneurons (Fig. 4B); and the Clarke's column, which receives proprioceptive information (Fig. 4C).

Combined adeno-associated virus encoding L1 (AAV-L1) and chondroitinase ABC (Chase) treatment enhances glutamatergic input to motoneurons in the lumbar spinal cord caudal to the lesion site. Confocal images of vesicular glutamate transporter 1-positive (VGLUT1+) terminals were obtained at the locations indicated on the diagram of a transverse spinal cord section shown at lower left. They were from the medial part of lamina VII (
The analysis revealed that the density (number of puncta per unit area) of VGLUT1+ terminals in the Clarke's column was higher by 29% and 26% for animal groups injected with Chase alone and AAV-L1 combined with Chase, respectively, compared with the AAV-GFP group (Fig. 4D). Interestingly, the Chase-treated groups showed an increased rump-height index, a measure of body weight support during walking, accompanied by increased excitatory afferent terminals in the Clarke's column, where proprioceptive sensory input is processed (Fig. 2C and 4D). On the other hand, only combined injections of AAV-L1 with Chase resulted in increased density of VGLUT1+ afferent inputs to the laminae VII and IX compared with the AAV-GFP controls. Taken together, these data suggest that altered plasticity of area-specific afferent inputs after SCI is associated with task-dependent motor recovery. Combined injections of AAV-L1 and Chase led to an overall improvement in both motor recovery and synaptic rearrangements.
Astrogliosis and NG2 proteoglycan expression in the injured spinal cord
Previously we have shown that immediate post-traumatic application of AAV-L1 leads, at 5 weeks after SCI, to reduced expression of GFAP, a marker protein for reactive astrocytes, and NG2 proteoglycan, a major axon growth inhibitory chondroitin sulfate proteoglycan (CSPG), compared to AAV-GFP treatment (Chen et al., 2007). We therefore analyzed whether delayed application of AAV-L1 and/or Chase have similar effects at 15 weeks after SCI. We found that Chase and the combined Chase/AAV-L1 application, but not AAV-L1 alone, led to reduced GFAP expression compared with AAV-GFP application (Fig. 5A). This lack of an L1 effect on astrogliosis was not associated with failure of L1 overexpression in astrocytes (Supplementary Fig. 1; see online supplementary material at
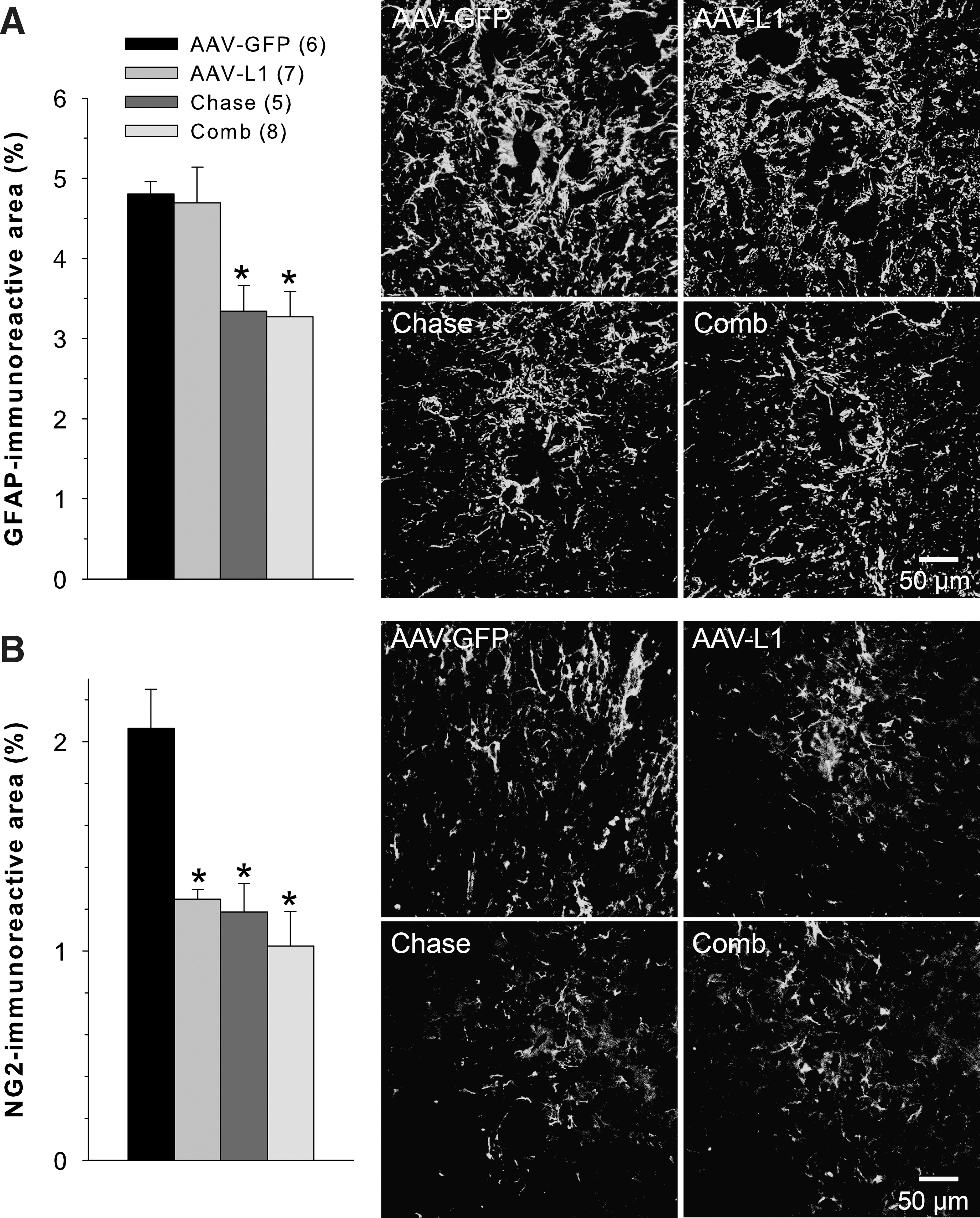
Effects of delayed application of chondroitinase ABC (Chase) and adeno-associated virus encoding L1 (AAV-L1) on astrogliosis and chondroitin sulfate proteoglycan (CSPG) expression 15 weeks after spinal cord injury (SCI). Astrogliosis was assessed in transverse sections distal to the lesion site using GFAP as a marker of reactive astrocytes (representative images from each of the four treatment groups are shown on the right hand side in
Discussion
In this study we show that delayed treatment of compression SCI in adult mice by a single parenchymal injection of L1 and Chase leads to improved functional outcome. This improvement was greater if the two agents were combined, and it was treatment-specific in the sense that L1 and Chase predominantly affected different motor abilities. At the histological level, analyses of chemically-defined synaptic terminals indicated that the two treatments augmented synaptic rearrangements that are likely to remodel synaptic activity and/or plasticity in the spinal cord in an area-specific, and when applied in combination, also additive manner.
Functional benefits of the L1 and Chase treatments
Based on the most commonly used measure of motor functions after SCI in mice, the locomotor BMS, the conclusion from the present experiments is that the application of L1, Chase, or their combination leads to similar degrees of functional improvement compared with the control AAV-GFP treatment. Motion analysis based on single-frame analysis from video-recordings showed, however, distinct functional effects of the different treatments. Compared with control mice, plantar stepping was improved only by the combined treatment, body weight support was augmented by the combined treatment and Chase, and voluntary target-reaching movements were enhanced by the combined treatment and AAV-L1. It is noteworthy that compared with the BMS locomotor rating, several individual parameters of the single-frame motion analysis provide a more differential and versatile assessment of distinct motor functions. This conclusion is not unexpected since rating scales give, on the one hand, a single score for a variety of walking abilities like, for example, stepping, coordination, and body weight support (Semler et al., 2011). On the other hand, the parameters of single-frame motion analysis are specifically designed to measure distinct and fine attributes of walking (locomotion), such as stepping estimated by the foot-stepping angle, and body weight support measured by the rump-height index, or functions that are not evaluated by BMS locomotor scales, such as target-reaching movements (motion) without body weight support estimated by the extension:flexion ratio (Apostolova et al., 2006; Semler et al., 2011). We propose that the dissection of overall locomotor behaviors into simpler motion attributes may aid the search for anatomical substrates underlying functional changes (see below), a search that will be more difficult if complex score measures are used (Fouad and Pearson, 2004).
Two functional effects observed in this study require special attention. First, we found a significant improvement of locomotor abilities by delayed application of Chase. This finding is in accord with the documented efficacy of Chase treatment, when initiated in the acute phase of SCI, leading to augmented axonal sprouting/regrowth and functional recovery (Barritt et al., 2006; Bradbury et al., 2002; Bradbury and Carter, 2011; Caggiano et al., 2005; Garcia-Alias et al., 2008; Jefferson et al., 2011; Tester and Howland, 2008). However, in the chronically injured spinal cord of rats, Chase effectively promotes co-treatments, but appears to be ineffective when applied alone (Karimi-Abdolrezaee et al., 2010; Tom et al., 2009a; Wang et al., 2011). The reasons for the efficacy of Chase in our model could be attributed to, for example, the type of injury (contusion, and complete or incomplete transection of the spinal cord), mode of Chase application (intraparenchymal versus intrathecal in other studies), or species (mouse versus rat). Regardless of the reasons for the different efficacy of Chase treatments, including findings that the enzyme may promote sprouting without influencing functional recovery (Tom et al., 2009b), our study provides strong support of previous work showing that Chase is beneficial, especially in combination with other efficient agents, for the sub-chronically injured spinal cord (Karimi-Abdolrezaee et al., 2010; Tom et al., 2009a; Wang et al., 2011). We also consider of special interest our observation of the beneficial effect of AAV-L1 applied alone or in combination with Chase on the voluntary movements analyzed by the extension:flexion ratio. This effect was larger than the effect of L1 on locomotor parameters (foot-stepping angle and rump-height index). In contrast, the results of acute SCI treatment with AAV-L1 showed enhanced recovery of gait parameters, and no effect on the extension:flexion ratio 5 weeks after injury (Chen et al., 2007). These results indicate differential effects of exogenously applied L1 depending on the time point of its application, although the relationship between time of treatment and magnitude of effect remains to be precisely determined. Promotion of voluntary movements in the sub-chronically injured spinal cord is particularly encouraging from a clinical point of view. Ground locomotion, such as walking, is to a large degree controlled by spinal mechanisms and can be efficiently trained in humans and experimental animals with SCI (Barbeau and Rossignol, 1987; Barriere et al., 2008; Rossignol et al., 2004; Wernig et al., 1995). Initiation and performance of voluntary movements require higher levels of supraspinal control, and are generally more impaired than stepping in any given SCI patient and are more resistant to training (Maegele et al., 2002).
Spinal cord plasticity and recovery of function
The differential functional effects of L1 and Chase suggest that the two treatments affect different anatomical targets. This notion is largely supported by our analyses of synaptic terminals in different areas of the spinal cord. First, we observed that L1, and to a similar degree the combined treatment with Chase, lead to an increase of ChAT+ and VGAT+ terminals at motoneuronal cell bodies 15 weeks after SCI, while Chase alone did not yield this effect. These findings indicate that L1 overexpression promotes plasticity of perisomatic modulatory (cholinergic) and inhibitory (GABAergic and glycinergic) synapses. This effect is explainable by the finding that AAV-L1 injection into the injured mouse spinal cord leads to L1 overexpression in both motoneurons and glial cells in the lumbar enlargement (Chen et al., 2007). Enrichment of the local environment in L1 is a well-known stimulus for compensatory axonal sprouting in the injured CNS (Irintchev and Schachner, 2011). Considering previous observations suggesting that functional performance after acute SCI in mice is related to the numbers of cholinergic and inhibitory perisomatic terminals (Devanathan et al., 2010; Jakovcevski et al., 2007), it is likely that the observed effects elicited by L1 in the sub-chronically injured spinal cord are functionally beneficial. It is surprising that Chase does not affect rearrangements of the perisomatic terminals, since these terminals are embedded in the perineuronal nets of motoneurons, which are extracellular matrix formations thought to restrict plasticity in adult mammals, and believed to be the main target of Chase (Bradbury et al., 2002; Carulli et al., 2010; Fawcett, 2009; Massey et al., 2006). Furthermore, we found that application of neither L1 nor Chase alone affect VGLUT1+ (excitatory glutamatergic) terminals at motoneuronal somata and pre-motor interneurons (lamina IX and VII, respectively). However, the combined application of L1 with Chase leads to a prominent increase of these glutamatergic synapses compared with the other treatments, indicating an additive effect. Finally, we found that these excitatory terminals are more frequent in the Clarke's column after Chase or combinatorial treatments than in single treatment with L1- and control-treated mice. The results of the VGLUT1+ terminal quantifications show that spinal excitatory afferents targeting different areas of the spinal cord are differentially affected by the treatments. Altogether, our findings support the idea that both treatments promote spinal cord plasticity, and thus motor recovery after SCI. The novel finding of dissimilar effects of these two treatments on synaptic rearrangements, and thus most likely also on synaptic activity/plasticity in several spinal cord compartments, may explain the observed differential and additive functional effects of L1 and Chase. However, the precise contribution to functional recovery of each specific aspect of plasticity observed here is unknown. It is a challenging task for future researchers to understand which alterations in spinal and supraspinal circuitries in different SCI models favor or impair restoration of the specific motor abilities after SCI. The importance of this notion becomes apparent when considering that enhanced plasticity in the adult mammalian spinal cord may have few apparent beneficial functional consequences (Tom et al., 2009b), or may even be detrimental by, for example, eliciting negative effects on motor skills that are not specifically trained (Garcia-Alias et al., 2009). In accord with these considerations, we can speculate that the enhanced glutamatergic input to the Clarke's column after Chase or combinatorial treatment observed in this study is a sign of plasticity with little or no contribution to functional recovery. The neurons of the Clarke's column send ascending projections through the ipsilateral spinocerebellar tract, and the functionality of this output pathway is certainly largely, if not entirely, compromised after the incomplete spinal cord compression injury.
In our previous study (Chen et al., 2007), we observed that L1 inhibits astrocyte proliferation and migration in vitro, and L1 overexpression in vivo leads to attenuated expression of GFAP and NG2 at 5 weeks after SCI compared with AAV-GFP treatment. Here we found no effect of delayed AAV-L1 application on GFAP expression by astrocytes at 15 weeks after SCI. A possible explanation for these differences is that delayed L1 overexpression after the third week after injury, in contrast to acute application, cannot influence astrocyte proliferation, which occurs mostly during the first week after injury (Lytle and Wrathall, 2007; White et al., 2010). In contrast to L1, Chase treatment was associated with a significantly lower level of GFAP expression, whereas treatment with both agents, AAV-L1, or Chase, led to reduction of NG2-positive glial cells. Overall, attenuated astrogliosis and/or NG2 expression at 15 weeks after SCI suggest that the delayed treatments lead to beneficial changes in the injured spinal cord environment, which may underlie the observed enhanced synaptic rearrangements, a notion supported by observations that enhanced L1 expression in the CNS promotes axonal sprouting (Aubert et al., 1998; Jucker et al., 1996; Styren et al., 1995).
In conclusion, the results of the present study show that delayed L1 overexpression is a potentially promising therapy for sub-chronic SCI. Especially attractive features of this treatment are its potential to improve voluntary movement capacities, its differential effects compared with another beneficial agent, Chase, and thus its additive mode of action on functional recovery and synaptic rearrangements/remodeling for synaptic plasticity in combination therapies.
Footnotes
Acknowledgments
The authors are grateful to the New Jersey Commission for Spinal Cord Research, Deutsche Forschungsgemeinschaft, and the Li Kashing Foundation for support (to M.S.). Bin Wu is supported by the National Natural Science Foundation of China (81000520). We are also grateful to Emanuela Szpotowicz for excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.