
Abstract
We investigated the role of the axon guidance molecule EphA4 following traumatic brain injury (TBI) in mice. Neutralization of EphA4 improved motor function and axonal regeneration following experimental spinal cord injury (SCI). We hypothesized that genetic absence of EphA4 could improve functional and histological outcome following TBI. Using qRT-PCR in wild-type (WT) mice, we evaluated the EphA4 mRNA levels following controlled cortical impact (CCI) TBI or sham injury and found it to be downregulated in the hippocampus (p<0.05) but not the cortex ipsilateral to the injury at 24 h post-injury. Next, we evaluated the behavioral and histological outcome following CCI using WT mice and Emx1-Cre-driven conditional knockout (cKO) mice. In cKO mice, EphA4 was completely absent in the hippocampus and markedly reduced in the cortical regions from embryonic day 16, which was confirmed using Western blot analysis. EphA4 cKO mice had similar learning and memory abilities at 3 weeks post-TBI compared to WT controls, although brain-injured animals performed worse than sham-injured controls (p<0.05). EphA4 cKO mice performed similarly to WT mice in the rotarod and cylinder tests of motor function up to 29 days post-injury. TBI increased cortical and hippocampal astrocytosis (GFAP immunohistochemistry, p<0.05) and hippocampal sprouting (Timm stain, p<0.05) and induced a marked loss of hemispheric tissue (p<0.05). EphA4 cKO did not alter the histological outcome. Although our results may argue against a beneficial role for EphA4 in the recovery process following TBI, further studies including post-injury pharmacological neutralization of EphA4 are needed to define the role for EphA4 following TBI.
Introduction
T
Ephrins are axon guidance proteins that are either anchored to the cell membrane by a glycosylphaditylinositol linkage (ephrinA) or have a cytosolic domain (ephrinB) that binds to its respective tyrosine kinase receptor (EphA or EphB). 10 Ephrins and their Eph receptors have been implicated in many aspects of neural development, including the formation of a variety of neural structures such as the corticospinal tract and anterior commissure. 11 –15 Since Eph-ephrin signaling may regulate axon guidance through contact repulsion, modified by the extracellular matrix, 16 a role in CNS injury has been implicated. 17 –19 Of particular interest in this regard is the multifunctional receptor EphA4, interacting with both ephrinA and B ligands, which may influence neuronal migration, differentiation, and connectivity. 17,20 –23 EphrinB3 is also expressed in adult CNS myelin where it inhibits neurite outgrowth, possibly via an EphA4 interaction. 24 Increased EphA4 expression was observed following corticospinal tract lesion and spinal cord injury (SCI), 25 –28 and, importantly, infusion of a peptide antagonist of EphA4 resulted in increased corticospinal tract axonal sprouting and a markedly improved motor recovery relative to controls. 26,29 Using EphA4-/- mice after SCI, increased axonal regeneration, reduced astrogliosis, and improved functional recovery were also observed. 28 These reports suggest an important role for EphA4 after CNS injury, which however has not to date been evaluated in experimental TBI. To evaluate the role for EphA4 in TBI, we first evaluated TBI-induced changes in gene transcription of EphA4 in wild-type mice. Then, mice deficient in EphA4 were subjected to TBI and we evaluated behavioral (cognitive, motor) and histological (hemispheric tissue loss, astrogliosis, and mossy fiber sprouting) outcome for up to 4 weeks post-injury. Since conventional EphA4 knockout mice develop a marked hopping gait, we instead used Emx1-Cre-driven conditional knockout (cKO) mice, 30 –32 in which EphA4 is absent from embryonic day 16; in these cKO mice, a normal pattern of locomotion is observed.
Methods
Animals
Male adult C57Bl/6 mice (B&K Laboratory, Stockholm, Sweden) were used for the qRT-PCR experiments. Both male and female mice with a conditional knockout (cKO) of EphA4 (see below) and their wild-type (WT) littermate controls were used in the behavioral testing and subsequent histology. Only homozygous cKO mice were used in this study. All animals had an initial body weight of 16–30 g and were housed at 24°C in cages with three to five mice per cage with free access to food and water in a 12-h light/dark cycle. The animals were kept in the colony for a minimum of 1 week prior to any surgical procedures. All mice were handled for 5 min per cage twice daily for 2 days before taking part in behavioral experiments. All behavioral and histological experiments and their subsequent evaluation were performed by an experimenter blinded to the genotype and injury status of each animal. Surgery of all animals was performed by an investigator (F.C.) blinded to the genotype of each animal. The histological analyses were performed on all animals that underwent functional testing. All procedures described herein were approved by the Uppsala County Animal Ethics board and followed the rules and regulations of the Swedish Animal Welfare Agency.
Generation of conditional EphA4 knockout mice
Non-conditional EphA4 knockout mice have severe coordination deficits and adopt a hopping gait. 15,33 To avoid these pre-existing neurological motor deficits, a conditional knockout model was used. 30 Here, Cre-expression was driven by the Emx1-promotor 31 and used to conditionally excise the EphA4 gene. In this model, Cre-recombinase is expressed in the hippocampus and most neocortical regions but not the basal ganglia. 31 In addition, the mice were also bred to contain one allele of the TaumGFP reporter that in the presence of Cre initiates production of β-galactosidase with a nuclear localization tag and green fluorescent tag (eGFP) with a membrane localization tag. 32 The genetic modification was done on a mix of 129Sv/J and C57BL/6 backgrounds. No obvious phenotypical differences between the cKO mice and their littermate controls were observed and no gait disturbances were detected in baseline, pre-injury motor tests (see discussion below).
Western blot analysis for EphA4 expression
CNS tissue samples from three controls and three cKO adult animals were homogenized in 10 times vol/ wt RIPA buffer (150 mM NaCl, 50 mM Tris-Cl [pH 7.4], 1% Triton X-100, 0.5% sodium deoxycholate, and 0.1% SDS) supplemented with 1X protease inhibitor cocktail (Roche, Basel, Switzerland). Protein concentration was measured by Bradford method using BSA as standards. Thirty μg total protein were separated on a 4–15% gradient polyacrylamide gel (Bio-Rad, Hercules, CA) and blotted onto nitrocellulose membrane (Bio-Rad). EphA4 and actin proteins were visualized by immunoblotting with the following antibodies and concentrations: mouse monoclonal antibody to EphA4, 1:5000 (Becton Dickinson Biosciences, Franklin Lakes, NJ); mouse monoclonal antibody to actin, 1:10,000 (cat. AC-15, Novus Biologicals, Littleton, CO); HRP-conjugated goat anti-mouse IgG, 1:2500 (cat. 62-6520, Invitrogen, Life Technologies, Grand Island, NY). The intensity of the resulting protein bands were quantified using ImageJ. Protein levels are presented as (EphA4 intensity/actin intensity).
Controlled cortical impact brain injury
Brain injury was induced using the controlled cortical impact (CCI) technique as described previously. 34 Anesthesia was induced using 4% isoflurane in air and maintained with 1.2% isoflurane in a 3:7 mixture of nitrous oxide and oxygen. The scalp was anesthetized using bupivacaine (Marcain, AstraZeneca, Sweden). The mice were placed in a stereotaxic frame on a heating pad and maintained at 37.0±0.5°C monitored by a rectal thermometer (CMA150, CMA Microdialysis, Stockholm, Sweden). A 4×4 mm craniotomy, keeping the dura mater intact, centered at 2.5 mm posterior to bregma and 2.5 mm lateral to the midline over the right hemisphere was created. Brain trauma was induced using a pneumatic cortical impact device (Amscien, Richmond, VA) with a 2.5 mm flat-tip metal impounder striking the exposed dura at 3.0 m/sec set to penetrate 0.5 mm into the exposed cortex. In all animals, the bone flap was then firmly reattached to the skull using tissue adhesive (Dermabond, Ethicon, Somerville, NJ). The incision was closed using interrupted resorbable sutures, and the animals were allowed to recover from the surgery in a wake-up cage equipped with an overhead heating lamp. Sham-injured controls (both WT and cKO mice) were subjected to anesthesia and craniotomy but not to CCI brain injury. Animal weights were monitored pre- and post-injury on regular intervals (Table 1). Animals losing more than 10% body weight following the injury were excluded from the study.
There were no statistically significant differences among the groups.
WT, wild-type mice; cKO, conditional knockout mice.
Quantitative RT-PCR
We used quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR) to evaluate TBI-induced changes in the gene expression of EphA4. Five brain-injured and two to three sham-injured (Table 2) mice from each time point (1, 3, and 7 days post-CCI) were anesthetized and sacrificed using rapid decapitation. To obtain RNA samples, bilateral neocortex and hippocampi were dissected (neocortical tissue were obtained from the injured area including the penumbra zone, and the whole hippocampus) and preserved in RNAlater solution (Qiagen, Valencia, CA). Total RNA was isolated using RNeasy Mini kit (Qiagen). Sample quality was assessed using the A260/A280 ratio and RNA concentration (ng/μL) was normalized from the absorbance at 260 nm, determined by a NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE) for each sample. The following primer pair was used: EphA4 (GenBank accession no. NM_007936; upper and lower primer: 5′-TGGAATTTGCGA CGCTGTC-3′ and 5′-CTTCCAGAGGGCTTGCTATC-3′). One of the primers spanned an intron-exon boundary to avoid contamination from genomic DNA. The Bio-Rad iScript One-Step RT-PCR Kit with SYBR Green (reverse transcription at 50°C for 30 min) amplified fragments of about 100 base pairs. qRT-PCR was run for 36 cycles (94–95°C for 10–15 sec, 60°C for 30 sec), using an MyIQ thermal cycler (Bio-Rad). Afterward, melting curves were obtained by increasing the temperature from 55.0°C to 94.5°C in increments of 0.5°C and examined to ascertain specificity of the PCR products (a symmetrical single peak). RNA concentration was also evaluated using the expression of 28S rRNA (X00525, 5′-GGGAGAGGGTGTAAATCTCGC-3′ and 5′-CTGTTCACCT TGGAGACCTGC-3′) and corresponded with the A260 values. Threshold cycle values (Ct) for the injured brain RNA samples were subtracted from average Ct values of an uninjured animal. The differences (ΔΔCt) were transformed to linear fold increases. All samples were run in duplicates.
WT, wild-type mice; cKO, conditional knockout mice.
Rotarod
The rotarod technique has been described as a sensitive test to detect motor deficits. 35 The rotarod used (Panlab, Barcelona, Spain) was set to accelerate from 0 to 40 rpm during 300 sec. The rotarod test was performed twice before surgery for baseline evaluation and then at days 2, 7, 13, 19, and 29 post-injury and four trials were run on each day. Jars of water were placed under the rods to increase the motivation of the animal to remain on the rotarod due to its natural instinct to avoid water. To further enhance motivation of the test, the trials were stopped at 0, 25, 100, and 300 sec, and the mice were then allowed to return to their home cage via a bridge. Animals that fell before the termination of the trial were placed in a holding cage until the next run. Latency to fall was recorded and if the animal remained on the rod until the termination of the trial this time was used. The longest latency achieved in the four trials was used for further analysis. Data are presented as percent of the pre-injury, baseline value.
Cylinder test
A unilateral injury to the motor cortex results in an asymmetry in the forelimb used for support during rearing, which can be measured using the cylinder test. 36 The mice were placed in a transparent glass cylinder, 7 cm in diameter and 38 cm high, containing wood chips at the bottom of the cylinder to encourage rearing, and were filmed at 10 frames/sec for 5 min or until 10 rears were observed. The animals were evaluated twice prior to surgery for baseline function and then at days 2, 7, 13, 19, and 29 post-injury. Two mirrors were placed behind the cylinder to enable simultaneous viewing of the mice from three different angles. Each rear was analyzed frame by frame in the films to see which paw made contact with the cylinder wall. 37 Each frame where a paw was in contact with the cylinder wall was scored as either “ipsi” (used the right forepaw), “contra” (used the left forepaw), or “both” (used both forepaw) based on which paws were used for support against the cylinder wall. The result was calculated as (“contra”+“both”/2)/(“ipsi”+“contra”+“both”), which is adapted from Woodlee and colleagues 37 and modified by replacing “ipsi” with “contra” in the numerator. With this modification, reduced use of the contralateral paw observed in brain-injured animals results in decreased, rather than increased, values compared to controls.
Morris water maze
The Morris water maze test evaluates how fast the mice learn the location of a hidden platform using visual cues surrounding the pool. 38 The 1.4 m diameter pool was filled with 22°C water and the 10 cm in diameter platform submerged 1 cm below the waterline, positioned in the south-east quadrant with its center 35 cm from the edge of the pool. The visual cues were placed on roller curtains surrounding the pool, assuring consistent cues between trials and allowing the experimenter to be out of sight during the trials. An automated tracking system was used to record the position of the mice (HVS Image, Buckingham, United Kingdom). Four daily trials, starting by placing the mouse in the tank at one of four designated entry points (N, S, E, and W), were made from post-injury days 20–23 in the acquisition, learning phase. Each trial was terminated when the mouse located the platform, at which the animal was allowed to sit for 15 sec to acquire the visual cues. Mice that failed to locate the platform in 90 sec were guided to it by the experimenter. The memory probe trial (platform removed) was performed at day 26 to enable evaluation of the ability to recall the previously learned task. In the probe trial, the latency to cross the platform position and the percentage of time spent in the quadrant previously containing the platform were recorded. In order to control for the visual ability of the animals and their motivation to find the platform, a black cylinder-shaped object was placed on the platform to reveal its location during the last MWM trial (visible platform test). Following all MWM trials, including the probe trial, the animals were placed under a heating lamp to maintain normothermia.
Perfusion and sectioning
At the end of the behavioral evaluation, 29 days post-injury, all animals were over-anesthetized with sodium pentobarbital (200 mg/kg i.p.) and then transcardially perfused using 50 mM sodium sulphate (to enable Timm staining) followed by 4% formaldehyde (Histolab, Gothenburg, Sweden). The brains were then rapidly removed and post-fixed for 4 h in 4% formaldehyde, transferred to 30% (w/v) sucrose for 2 days, and then snap frozen in dry-ice chilled isopentane. Following post-injury processing, each brain was cut into 25 μm coronal sections on a cryostat (HM500, Microm, Walldorf, Germany).
Evaluation of tissue loss
To measure the TBI-induced tissue loss, nine coronal sections from bregma levels 0 to −4 mm, 500 μm apart, were stained with Mayer's hematoxylin and eosin (Histolab) and photographed using a stereomicroscope (Stemi 2000-C, Zeiss, Göttingen, Germany) equipped with a digital camera (Mcm5c, Zeiss). The tissue area, ventricles excluded, of the contralateral and ipsilateral hemispheres, as well as the neocortical area, was measured using the software SectionToVolume 39 by an evaluator blinded to the injury and treatment status of each animal. Using this program, brain tissue was automatically detected based on its color resulting in the same result as when manually tracing the outline of the section and ventricles. 39 The volume of each hemisphere and neocortex between two sections was calculated as d*(A1+A2)/2, where d is the distance between sections and A1 and A2 is the measured area in the two sections. Hemispheric and neocortical tissue loss was calculated by subtracting the ipsilateral volume from the contralateral volume.
GFAP immunohistochemistry
GFAP expression was evaluated at 29 days following CCI in four selected regions of interest (ROI; Fig. 6) in sections from bregma −1.5, −2.5, and −3.5 mm. Unspecific binding was blocked using 2% NGS (normal goat serum) in PBS with 0.1% Triton-X 100 (blocking buffer) for 30 min in room temperature. The sections were then incubated at 4°C overnight with a primary antibody against GFAP (Z0334, Dako, Glostrup, Denmark) at 1:500 in blocking buffer. The sections were then washed with PBS and the secondary antibody (anti-rabbit-Cy3, C2306, Sigma-Aldrich, St. Louis, MO) was applied in blocking buffer and incubated for 45 min. Finally the sections were washed in PBS and mounted using Vectashield with DAPI (4′, 6-diamidino-2-phenylindole, Vector Laboratories, Burlingame, CA). The slides were kept frozen at −20°C until analyzed using densitometry. A fluorescence microscope system (Zeiss Axiovision) was used to capture 10x images from the cortical ROIs (1×1 mm) and 20x images from the hippocampal ROIs (0.5×0.5 mm); the exposure time was identical for each image. The fraction of the ROI stained over a set threshold, kept constant throughout the analysis, was measured using the built-in software of the microscope. The mean value of the six cortical ROIs and the six hippocampal ROIs from the three different bregma levels was calculated. Data are presented as fold increase compared to the value of the sham-injured, WT group.
Timm staining
To evaluate sprouting of the zinc-containing mossy fibers in the hippocampus, Timm-stained sections were evaluated according to previously published protocols. 7,40 –42 Slides with sections were placed, without light exposure, in a developing solution containing 180 mL 50% (w/v) gum Arabic, 30 mL 2 M citrate buffer, 90 mL 0.5 M hydroquinone, and 1.5 mL 1 M AgNO3 until sufficiently stained (∼40 min). The Timm reaction was inhibited by a gentle rinse in tap water (in the dark) for 30 min, and glasses were then placed in 5% (w/v) Na2S2O3 for 12 min, washed in H2O 2×5 min, dehydrated, and coverslipped using Pertex (Histolab). We then evaluated the Timm stain in two hippocampal areas, both ipsi- and contralateral to the injury, in sections from bregma −1.5, −2.5, and −3.5 mm, according to the protocol originally published by Cavazos and colleagues. 42 In this protocol, sections are scored from 0–5, where “0” means no granules; “1” means sparse granules in the supragranular region and in the inner molecular layer; “2” means granules evenly distributed throughout the supragranular region and the inner molecular layer; “3” means an almost continuous band of granules in the supragranular region and inner molecular layer; “4” means a continuous band of granules in the supragranular region and in the inner molecular layer; and ”5” means a confluent and dense laminar band of granules that covers most of the inner molecular layer, in addition to the supragranular region.
Statistical analysis
Due to the rather low number of animals in each group, only non-parametric statistical tests were used. Data were analyzed using a Kruskal Wallis ANOVA that, if significant at p<0.05, was followed by Mann-Whitney U-test for pair-wise comparison between groups. In addition, since the animals were analyzed at several time points, all behavioral data were also analyzed using the Friedman's test for repeated measures. For clarity, data are presented as mean+SEM, and p<0.05 was considered significant. All statistical analyses were made using Statistica (V.9, StatSoft, Tulsa, OK).
Results
There were no deaths resulting from the surgical procedure or the brain injury. After both sham and brain injury, all mice decreased in body weight but rapidly recovered (Table 1), except for one mouse in the brain-injured cKO group that gradually lost weight and was excluded from the study 19 days post-injury. There were no significant differences in body weights pre- and post-surgery among the injury groups in the behavioral study (Table 1). In all, 79 mice were used in the study; the number of animals in each study group is presented in Table 2.
qRT-PCR for EphA4
The gene expression of EphA4 in cortex and hippocampus bilaterally was evaluated by qRT-PCR at 1, 3, and 7 days following sham or brain injury. In the neocortical region, there were no significant TBI-induced changes at any evaluated time point (Fig. 1A). In the hippocampal region, the gene expression of EphA4 was reduced in brain-injured animals compared to sham-injured controls, reaching statistical significance at 24 h post-injury but not at any other evaluated time points (p<0.05; Fig. 1B).
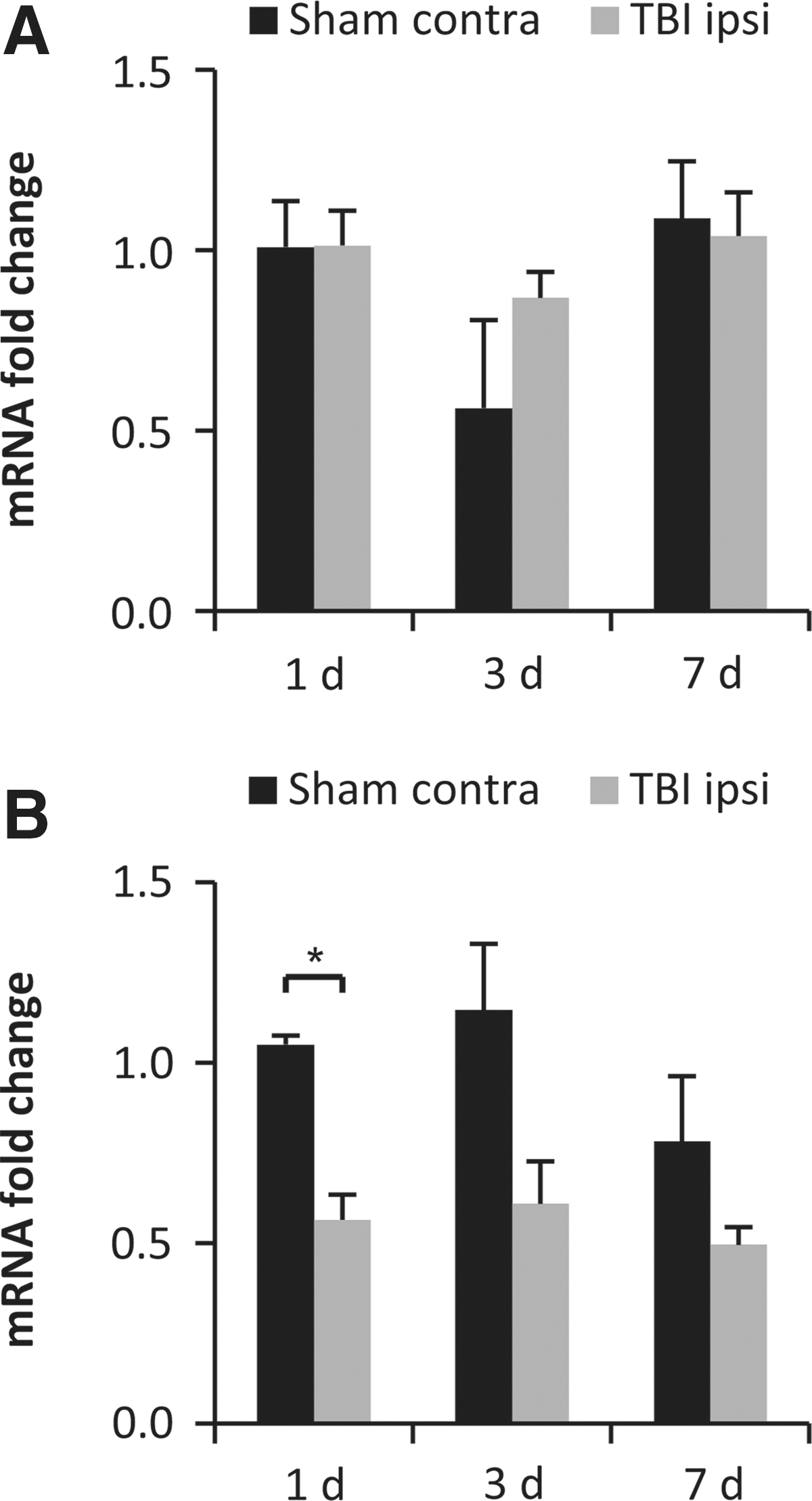
Gene expression of EphA4. Temporal alterations in neocortical (
EphA4 recombination efficiency in cKOs
The efficiency of recombination in EphA4 cKO mice was assessed by Western blot analysis (Fig. 2A). Only minimal amount of EphA4 was detected in hippocampal lysates and was reduced by 59% in cortical lysates. Low levels of EphA4 were detected in the midbrain and lumbar spinal cord. The midbrain EphA4 levels were not altered in cKO mice although they were increased in the lumbar spinal cord (Fig. 2B).
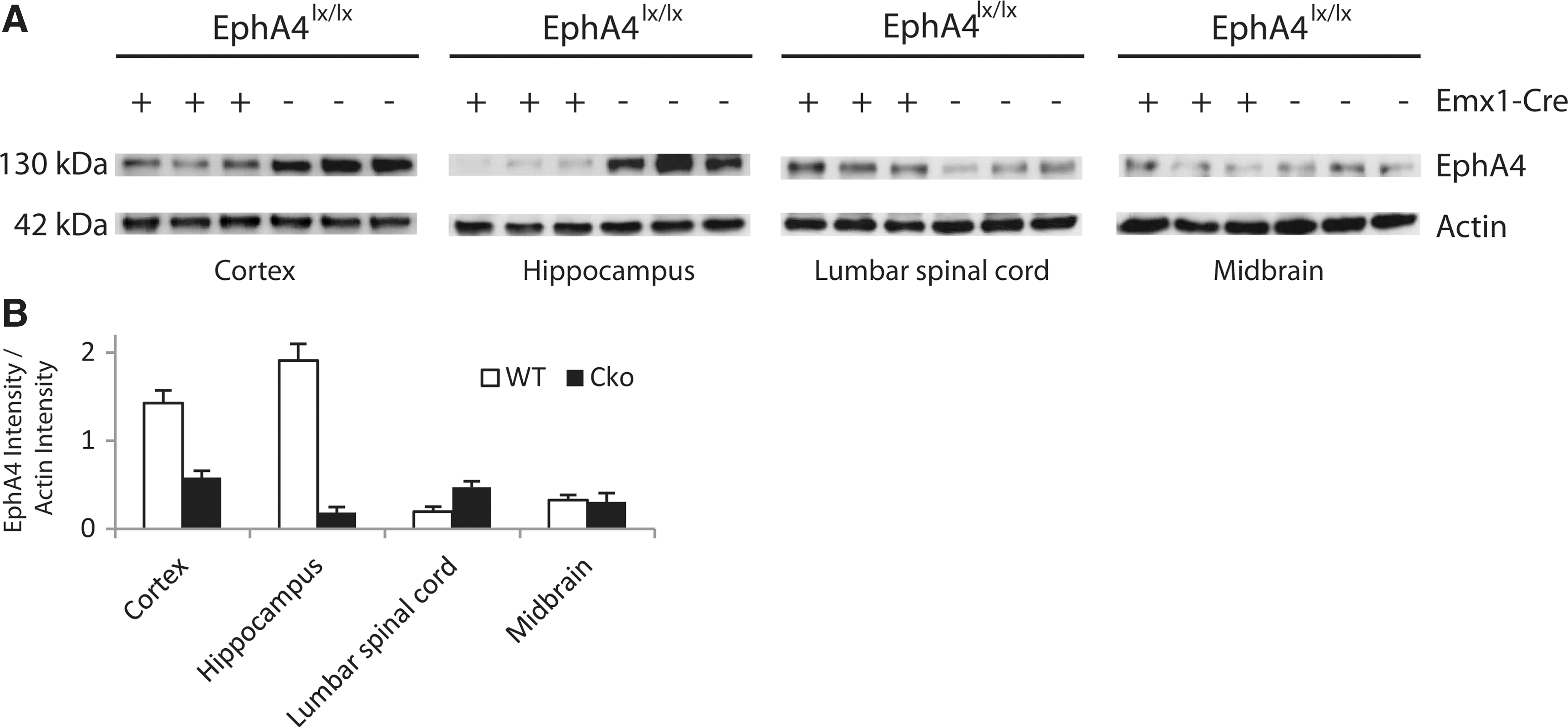
EphA4 protein expression in different CNS regions from EphA4 cKO mice and controls was assessed using Western blot analysis. The resulting protein bands are shown (
Rotarod
The rotarod latencies increased from the first to the second day of pre-training, demonstrating an initial learning aspect of this test (data not shown). There were no differences in pre-injury rotarod performance among the treatment groups (data not shown). A repeated measures analysis revealed a significant difference among the groups (χ2(6)=68.6, p<0.05). TBI resulted in a shorter latency to fall at day 2 post-injury in both WT and cKO mice compared to sham-injured mice (p<0.05; Fig. 3A). This TBI-induced motor deficit had recovered in both WT and cKO mice by day 13. Absence of the EphA4 gene did not result in a significantly altered rotarod performance in either brain-injured or sham-injured animals during the testing period.
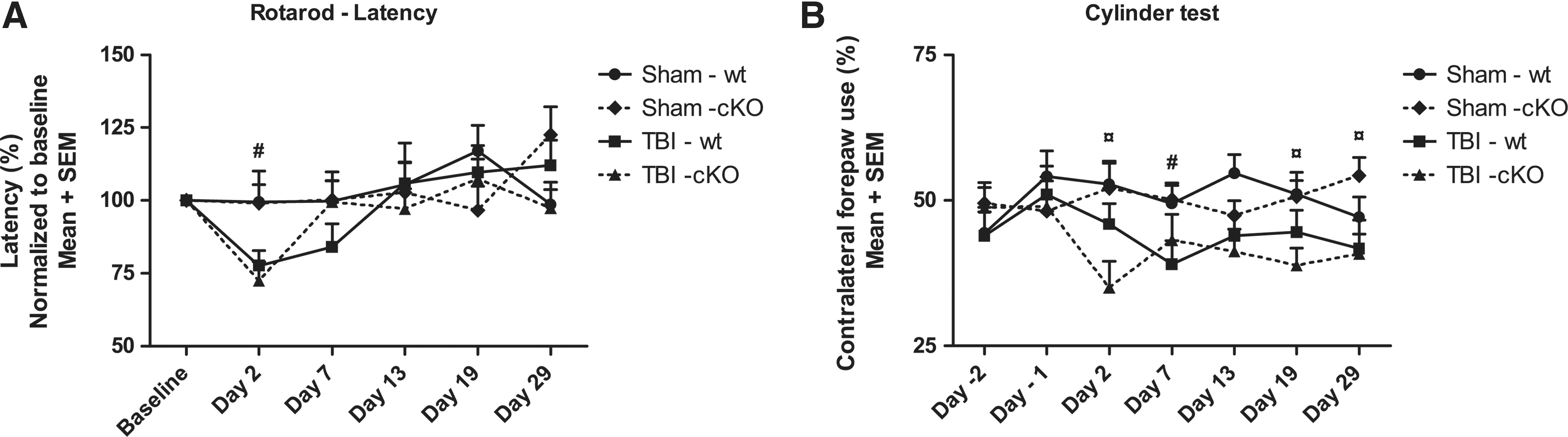
Motor function at baseline (pre-injury) and at days 2, 7, 13, 19, and 29 post-TBI. (
Cylinder test
Following TBI, both brain-injured groups had consistently lower use of contralateral forelimb use for the entire duration of the study and a repeated measures analysis revealed a significant difference among the groups (χ2(6)=71.9, p<0.05). At 29 days post-injury, brain-injured mice still used the ipsilateral forelimb (60%) more than the contralateral forelimb (40%) (Fig. 3B). Post-injury, sham-injured mice had higher values compared to brain-injured mice, reaching statistical significance at day 7 between the WT groups and at days 2, 19, and 29 between the cKO groups (p<0.05) although not at day 13 (Fig. 3B). In all, the conditional knockout of EphA4 did not significantly influence the performance in the cylinder test.
Morris water maze
At days 20–23 after CCI brain injury or sham injury, animals were evaluated for their ability to learn the position of a hidden platform in the MWM. A repeated measures analysis revealed a significant difference among the groups (χ2(3)=51.9, p<0.05). Sham-injured animals quickly learned the MWM visuospatial task defined by decreasing latencies to find the platform during the 4-day evaluation period (Fig. 4A).

Acquisition, learning (
Brain-injured, WT mice had a significantly reduced performance in the MWM on the second, third, and fourth day of testing compared to sham-injured controls. Genotype did not appear to affect MWM latency since there were only small, non-significant differences between WT and cKO animals (Fig. 4A). At 72 h following the last MWM learning trial, the platform was removed, the probe (memory) trial was performed, and the first 30 sec of the trial was analyzed for the latency to cross the platform position and the percentage of time spent in the correct quadrant of the pool. Brain-injured cKO mice had significantly longer latencies (p<0.05; Fig. 4B) to locate the platform area compared to their sham-injured controls. For the brain-injured WT mice, the difference compared to their WT sham-injured control did not reach statistical significance (p=0.09; Fig. 4B). Brain-injured mice spent less time in the correct quadrant compared to the sham-injured groups without reaching statistical significance (Fig. 4C). Brain injury did not alter the swim speed (data not shown) of the mice as previously reported. 43
Evaluation of tissue loss
The CCI brain injury predominately causes a focal loss of tissue in the ipsilateral cortical and hippocampal regions, although some bilateral axonal injury exists. 44 Here, the CCI brain injury caused a significant amount of tissue loss in both the cKO and WT mice compared to sham-injured controls (p<0.05; Fig 5A), to a similar extent in both brain-injured groups. Typically, the CCI brain injury results in a large necrotic cavity (Fig. 5B), although in 43% of the brain-injured mice of both genotypes, there was an obvious loss of cortical tissue without cavity formation (Fig. 5C). Thus, since several mice did not display a significant necrotic cavity, we evaluated loss of hemispheric tissue in addition to the neocortical tissue loss (Fig. 5A).

(
GFAP immunohistochemistry
Reactive astrocytosis is a consistent and robust response to TBI across animal species including humans, primates, and rodents.
45,46
To evaluate any involvement of EphA4 in the development of astrogliosis,
28
ipsilateral to the CCI injury we evaluated four regions of interest from three different bregma levels, for GFAP expression using immunohistochemistry and densitometry (Fig. 6A). In sham-injured animals, there were only a few scattered cells showing GFAP immunoreactivity. However, after TBI there was a markedly increased GFAP immunoreactivity showing an increased number of GFAP-positive cells that were larger and showed elongated and thickened processes (Fig. 6B,C). There were no differences in GFAP immunoreactivity between the sham-injured groups. When compared to sham-injured mice, brain-injured mice showed a markedly increased GFAP immunoreactivity in the majority of evaluated ROIs (Fig. 6B,C). Although not showing statistical significance, GFAP immunoreactivity in brain-injured cKO mice was higher than that of the brain-injured, WT mice (Fig. 6B,C). GFAP immunohistochemistry following sham or brain injury at 29 days post-injury. (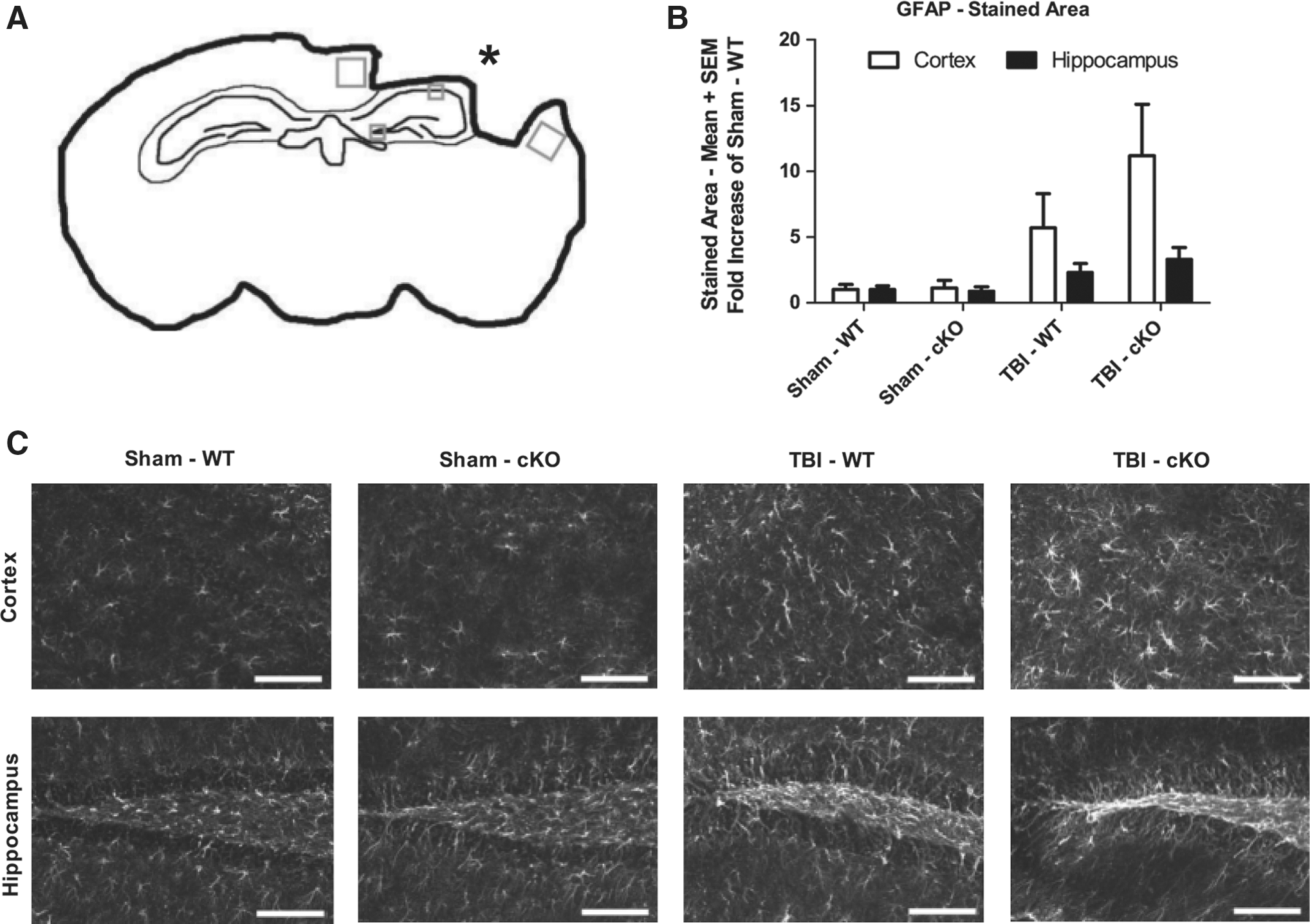
Timm staining
To study a measure of post-injury plasticity, we evaluated hippocampal mossy fiber sprouting, which was previously shown to increase after experimental TBI and seizures. 7,42 Using the Timm staining, the sprouting of mossy fibers was visualized. We evaluated two hippocampal areas at three different bregma levels (Fig. 7A–C). In only two of the evaluated areas did TBI per se increase Timm staining (p<0.05; Fig.7D), and in the remaining areas, the Timm staining score was similar in the brain-injured and sham-injured groups. Absence of EphA4 did not influence Timm staining score in either the sham- or brain-injured groups (Fig. 7D).
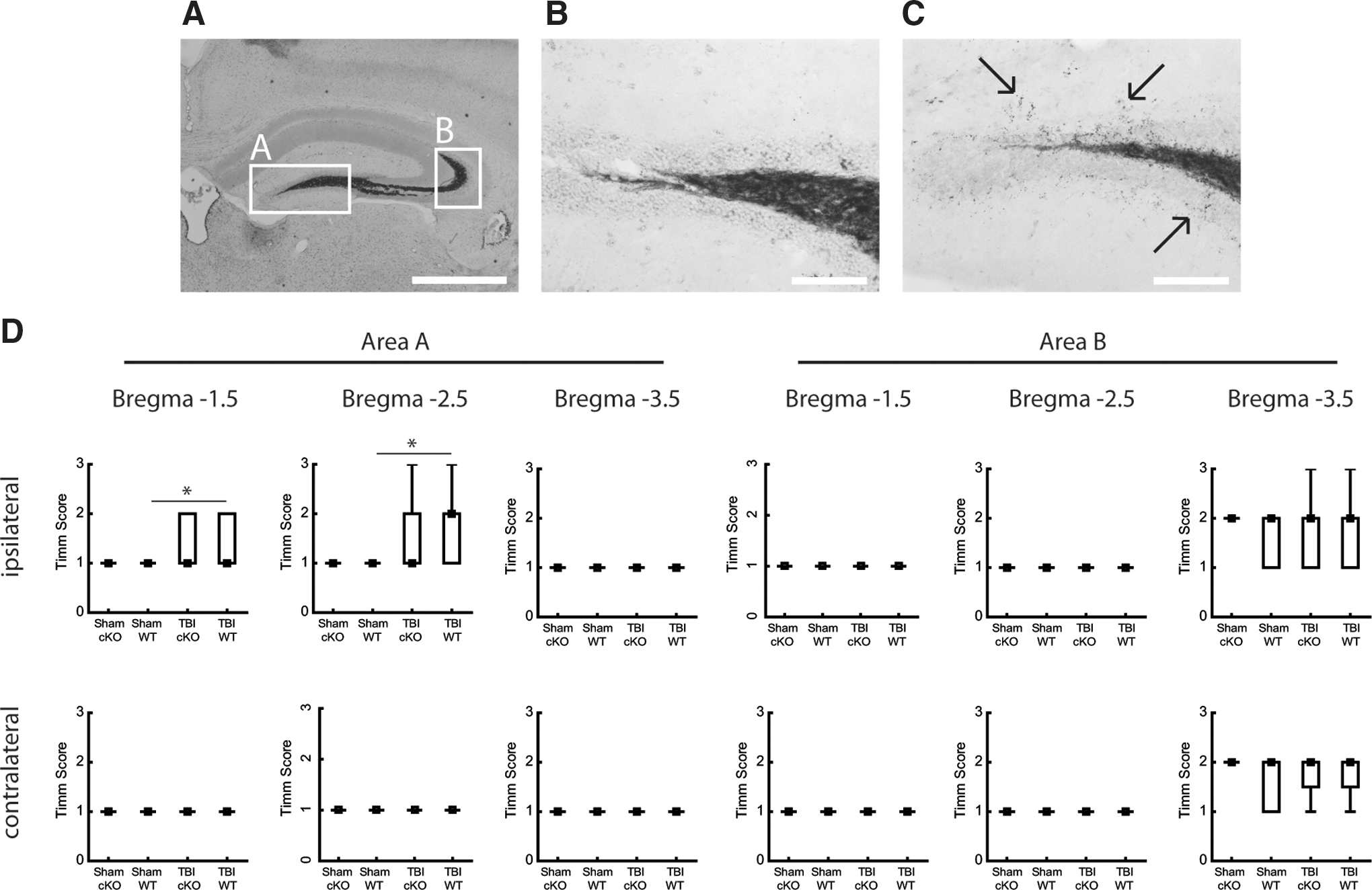
Timm stain evaluating mossy fiber sprouting at 29 days post-injury. (
Discussion
In the present report, we used the controlled cortical impact TBI model and evaluated the gene expression of EphA4. In WT mice, EphA4 mRNA was downregulated in the ipsilateral hippocampus at 24 h post-TBI although there were no TBI-induced changes in the ipsilateral cortex. We also evaluated the post-injury behavioral and histological outcome in conditional EphA4-knockout mice in which EphA4 is absent from certain forebrain regions from embryonic day 16. We used Western blot analysis and confirmed that only minimal levels of EphA4 were detected in the hippocampus and reduced to ∼40% of control values in the cortex. The increased EphA4 levels in the lumbar spinal cord of the cKO mice were unexpected, although the absolute EphA4 levels in both groups were low. The significance of this change is unclear, although may represent a compensatory upregulation of EphA4 caused by the reduction observed in other CNS regions. The reduction of EphA4 in the cortex and hippocampus was not associated with an improved behavioral outcome and the histological outcome was similar in brain-injured cKO and WT mice. In contrast to the results obtained with pharmacological neutralization or EphA4 knockout mice in models of spinal cord injury (SCI), 26,28 our results may argue against an important beneficial role for EphA4 inhibition in the recovery process following TBI.
EphA4 is broadly expressed and often coexists with ephrin ligands, suggesting a multifunctional role for EphA4. 47 –49 Following SCI, increased expression of Ephs occurs in adult rat and mouse. 28,50 –53 Additionally, EphA4 protein accumulates in transected, proximal axon stumps that may allow a direct inhibitory action on axons via ephrinB2-positive reactive astrocytes 27 and ephrinB3 on oligodendrocytes. 24 In post-mortem human TBI, the expression of EphA4 was increased and associated with reactive astrocytes. 54 Using qRT-PCR, we did not detect any significant TBI-induced changes in EphA4 mRNA expression in the cortical regions, similar to observations made in a mouse model of subdural hematoma. 50 Since this TBI model results in a marked cortical tissue loss, we cannot rule out that focal parts of the remaining cortical tissue had increased EphA4 expression. Our data suggest that the post-injury regulation of EphA4 differs markedly between SCI and TBI. However, we observed a significant downregulation of hippocampal EphA4 gene expression at 24 h post-injury, similar to that observed after deafferentation of the superior colliculus. 55 Yet the hippocampal downregulation was rather mild and transient and to evaluate the role for a prolonged and marked reduction of EphA4 signaling, we evaluated the behavioral and histological outcome after TBI in conditional EphA4 knockout mice.
After SCI, extensive axonal regeneration, reduction in post-injury astrogliosis, and a decreased RhoA activation were observed by 6 weeks post-injury that correlated with an enhanced functional recovery in EphA4-deficient mice. 28 Additionally, when an EphA4-blocking peptide was infused after SCI, markedly improved behavioral outcome was observed. 26 In contrast, no regeneration post-SCI was observed using antisense oligonucleotides to block EphA4 synthesis, 56 suggesting that inhibition of existing EphA4 is required to improve recovery and that merely inhibiting EphA4 synthesis is insufficient. We hypothesized that loss of the EphA4 gene from embryonic day 16 would lead to an improved behavioral outcome after CCI brain injury. A conditional KO mouse model was used, since conventional EphA4 KO mice display a hopping gait 33 difficult to evaluate in behavioral tests for cognition and motor function. Here, we used the rotarod and cylinder tests of neurological motor function and showed that sham-injured EphA4 cKO mice had similar pre- and post-injury motor function, arguing against a marked deficit induced by the absence of EphA4 per se. In addition, EphA4 cKO mice performed at a similar level in the motor tests compared to the wild-type, littermate brain-injured controls. Stimulation of EphA4 has previously been shown to exert both negative and positive consequences, depending on the level of stimulation and characteristics of participating cells. 57 –60 In vitro data suggest that ephrinB3 is a myelin-based inhibitor of neurite outgrowth suggesting a role for EphA4/ephrinB signaling in CNS injury. 24 In rodents, EphA4 is present in the hippocampus where it was suggested to play a role in adult synaptic plasticity and learning. 61 –64 The role for EphA4 in the cognitive function post-injury is unknown and we hypothesized, in view of the axonal growth-promoting actions observed following genetic and pharmacological neutralization of EphA4, that cognition should be enhanced in EphA4 cKO mice post-TBI. However, the brain-injured cKO mice performed similar to the brain-injured, WT mice in the cognitive tasks. Our data suggest that EphA4 does not influence the pathology leading to cognitive dysfunction after TBI. In fact, in an Alzheimer's disease (AD) mouse model overexpressing human amyloid-beta protein precursor, reduced expression of EphA4 receptors was associated with cognitive impairment. 65 These data, in addition to ours, suggest caution when neutralizing EphA4 in the early post-injury phase.
In the present report, EphA4 cKO mice had similar loss of hemispheric and neocortical tissue compared to brain-injured, wild-type controls. Although cell death after CCI TBI was increased in EphrinB3 KO mice and reduced in EphB3 KO mice, 66 our finding is similar to that observed following experimental SCI and indicates that EphA4 does not markedly influence cell survival after acute CNS injury. Instead, we focused on the astrocytic response to TBI. Astrocytes make up about 50% of all cells in the brain and supply substrates for neuronal energy metabolism. 67 An initial response to brain or spinal cord injury includes reactive astrocytosis, resulting in hypertrophy and proliferation of astrocytes and production of growth factors and cytokines. 68,69 Reactive astrocytosis isolates the lesion area by forming a barrier between healthy and injured tissue. 67 After experimental TBI, prolonged astrocytic hypertrophy and increased GFAP immunoreactivity have consistently been observed. 67,70 –74 Astrocytic hypertrophy/astrogliosis may be both beneficial and detrimental post-TBI since astrocytes may have both growth-inhibiting and growth-promoting properties, likely determined by injury model, time post-injury, and location of injury. 46,67,75 EphA4 was shown to be involved in the astrogliosis that contributes to scar formation after injury in adult rat and mouse 28,50 –53 and primates. 45 Surprisingly, absence of EphA4 did not significantly alter the astrocytic response after TBI in the present report, in contrast to the markedly reduced astrocytic scar formation observed following SCI in EphA4 KO mice. 28 The reasons for this difference is not clear although the injury model, the post-injury time point, and the genetic model are markedly different in the study by Goldshmit and colleagues 28 compared to those of ours. Astrocytes are also a very heterogeneous cell population, 73,76 and the response to EphA4 absence may differ between spinal cord astrocytes and cortical astrocytes. In support of our results, the astroglial scar was similar after SCI in mice with an EphA4 gene trap mutation compared to wild-type controls. 77 Although our data argue against a prerequisite for EphA4 signaling in mediating post-traumatic astrocytosis, it should be noted that GFAP immunostaining was evaluated only at one rather late post-injury time point and that the dynamics of the astrocytic response to TBI in this strain of EphA4 cKO mice still remains to be investigated.
Next we evaluated the effect of conditional gene deletion of EphA4 on post-traumatic hippocampal mossy fiber sprouting as a measure of brain plasticity. Ephs and their ephrin ligands have been implicated in the establishment of functional circuits, cell migration, and excitatory synapse regulation. 78 Since recovery after TBI must involve a degree of plasticity, modulation of sprouting may be an important aspect in pharmacological therapy after TBI. The Timm stain was used to evaluate mossy fiber sprouting, previously shown to be increased following TBI in the rat 79 and mouse. 80 Here, we evaluated the Timm stain in six different hippocampal regions and found it to be increased following TBI in two of the evaluated regions, with no differences between the brain-injured groups. It should be noted that the mossy fiber sprouting induced by TBI was slightly less pronounced than in a previous report from our group. 7 In that report, all animals had received intrathecal infusion via mini-osmotic pumps, although it is unlikely that this infusion influenced the Timm stain. Thus, the reason for the less marked Timm staining in the present report is unclear. It should be noted that increased sprouting per se may also be detrimental and increased (and aberrant) sprouting has been implicated in the development of post-traumatic epilepsy, 79,81 and the time course and magnitude of the sprouting need to be clarified. Although Eph/ephrinA interactions have been suggested to be involved in activity-dependent sprouting, 82 our present results argue that EphA4 does not influence post-injury sprouting of hippocampal mossy fibers. Evaluation of mossy fiber sprouting at various time points following TBI in cKO mice and evaluation of sprouting in other major fiber tracts are warranted.
Conclusion
We conclude that TBI transiently downregulates ipsilateral hippocampal gene expression of EphA4. However, additional reduction of EphA4 levels on the outcome after TBI using conditional knockout mice, in which EphA4 was virtually absent from the hippocampus and markedly reduced in the cortex from embryonic day 16, was not associated with improved behavioral or histological outcome after TBI. Our results, in contrast to results obtained in experimental spinal cord injury models, argue against a beneficial role for EphA4 after TBI. Differences in genetic background, a compensatory upregulation of other members of the Eph/ephrin family, or any residual EphA4 expression may, however, contribute to the differences in results between the acute CNS injury models. We suggest that post-injury pharmacological inhibition of EphA4 should be performed in rodent TBI models to further define the involvement of EphA4 in the pathophysiology of TBI.
Footnotes
Acknowledgment
The authors wish to thank Elaine Åstrand and Moritz Mayerle for excellent technical support. This study was supported by Upplandsstiftelsen, Uppsala University, Jeanssons Foundation, and the Swedish Brain Foundation. Dr. Kullander is a Royal Swedish Academy of Sciences Research Fellow and is supported by a grant from the Knut and Alice Wallenberg Foundation.
Author Disclosure Statement
The funding agencies did not have a role in the design and interpretation of the work presented here. No competing financial interests exist.