
Abstract
In the present study, we compared the roles of gracile neurons in mechanically-induced neuropathic pain caused by spinal injury and L5 spinal nerve ligation in rats. Behavioral and electrophysiological methods were used to measure mechanical allodynia in the hindpaws, and excitability of the gracile neurons in the medulla, respectively. In the spinal hemisection and spinal contusion models, mechanical allodynia developed in both hindpaws and lasted over a month. Three weeks following the hemisection, gracile neurons identified as wide-dynamic-range (WDR) and low-threshold (LT) neurons, showed increased neuronal activity to non-noxious mechanical stimuli compared to control groups, whereas the spinal contusion groups did not show evoked activity (*p<0.05). A lesion of the gracile nucleus partially reversed the existing mechanical allodynia in both hindpaws compared to prior to the injury in the hemisection group, whereas the spinal contusion groups did not show significant changes (*p<0.05). In the spinal nerve ligation model, mechanical allodynia developed at the ipsilateral (injured) side of the hindpaw. In addition, WDR neuronal activity at the ipsilateral gracile neurons showed a significant increase with non-noxious mechanical stimuli, whereas the LT neurons did not show significant changes (*p<0.05). Similarly to the hemisection model, a lesion of the gracile nucleus attenuated the mechanical allodynia in spinal nerve ligation models. The present data suggest that gracile neurons contribute to the maintenance of non-noxious mechanically-induced neuropathic pain in both hemisection- and ligation-induced neuropathic pain in rats.
Introduction
In the spinal cord, two ascending pathways mediate sensory input from peripheral receptive fields to the higher nervous system, including the brainstem and cortex. Spinothalamic tract pathways (STTs) mediate nociceptive inputs to the ventral posterolateral (VPL) nucleus of the thalamus, whereas dorsal column-medial lemniscus pathways (DC-MLs) mediate non-noxious input to the gracile nucleus of the medulla. 5 In the spinal dorsal horn regions, two distinct ascending pathways discriminate the properties of ascending somatosensory input to the higher nervous system. However, both pathways converge at the contralateral side of the thalamic VPL regions; STTs decussate at the spinal cord, whereas DC-MLs decussate at the gracile nucleus of the medulla.
Over the course of the past decade, the literature has suggested that neuropathic pain caused by SNL and hemisection produced similar intracellular and intercellular spinal mechanisms that are primarily mediated via the STTs. Activation of astrocytes and microglia, 6,7 upregulation of glutamate receptors, 8,9 increased production of reactive oxygen species (ROS) and proinflammatory cytokines, 3,10 and decreased endogenous GABAergic inhibitory tone, 4,11 are predominant pathophysiological changes in the spinal dorsal horn that result in neuropathic pain following injury. These changes contribute to the persistent neuronal hyperexcitability observed in response to both non-noxious and noxious mechanical stimuli in the dorsal horn neurons. Consequently, enhanced neuronal activity causes chronic neuropathic pain with both SNL and hemisection injuries. In addition, recent literature suggests that supraspinal mechanisms also contribute significantly to the incidence of neuropathic pain following SNL and hemisection. We and others have reported that hyperexcitability of the thalamic VPL neurons contributes to mechanical allodynia in the hindpaw, which includes similar mechanisms of neuropathic pain in the spinal dorsal horn. 12,13
In contrast to STTs, DC-MLs mediate non-noxious inputs to the supraspinal site of the gracile nucleus, and is considered an important ascending pathway for mechanical allodynia. However, the contribution of gracile neurons to the neuropathic pain seen with peripheral and spinal cord injuries remains unclear. In the present study we compared the roles of gracile neurons in the maintenance of mechanical allodynia, and the changes in neuronal activity to mechanical stimuli with peripheral and central neuropathic pain in a rat model.
Methods
A total of 89 male Sprague-Dawley rats (200–250 g) were used. We randomly divided the animals into three groups: hemisection, contusion, and ligation. For the central neuropathic pain model, the spinal hemisection (HEMI) injury was performed by spinal transverse hemisection at T13 as previously described. 2 After anesthesia using masked-isoflurane inhalation (2–3%), the surgical field was shaved and a longitudinal incision was made exposing several segments. After a laminectomy (T11–T12 vertebrae), the left side of the T13 spinal cord was unilaterally hemisected just cranial to the L1 dorsal root entry zone with a micro-dissecting knife. An insulin syringe (28-gauge needle) was inserted dorsoventrally at the midline of the cord, and the syringe was pulled laterally to ensure completeness of the hemisection without damage to the major dorsal vessel or vascular branches. Only one rat with deficits of the right hindpaw was excluded from the study. To compare the differential effect of ascending dorsal column pathways, bilateral spinal cord injury was performed by contusion injury at the T10 level using an Infinite Horizon Impactor. After deep anesthesia by intraperitoneal (IP) injection of sodium pentobarbital (60 mg/kg), laminectomy of the T8/T9 vertebral segment was exposed to the T10 spinal level and contused by a 150-kdyn force with 1 sec dwell time. After contusion, enrofloxacin (2.5 mg/kg) was intradermally injected for 5 consecutive days. The peripheral neuropathic pain model was produced by unilateral left L5 spinal nerve ligation (SNL). 10 Under isoflurane anesthesia, the left L5 spinal nerve was aseptically isolated and tightly ligated with 7-0 silk thread. For control groups (sham surgery), rats with the corresponding body weight were subjected to surgery without the spinal cord or L5 spinal nerve injuries. The bilateral lesions of the gracile nucleus were made by mechanical crush using fine forceps. To confirm the gracile lesion size, immunohistochemistry was performed. After perfusion with cold saline and 4% paraformaldehyde solution, the gracile region (bregma −13.68 to −14.60 mm) was removed according to the rat brain atlas (Paxinos and Watson, 1997), 14 and stored overnight in the cold room for post-fixation. After neuroprotection with 30% sucrose solution, the gracile region was mounted with mounting compound before sectioning. Using horizontal serial sections (60 μm), we selected each fifth section by a free-floating method using 0.1 M phosphate buffer (pH 7.4). The sections were evaluated using Radiance 2000 imaging software. Following the gracile lesion, 2 rats exhibiting abnormal behaviors (vocalization and unstable posture) were excluded from the analysis.
Mechanical allodynia was characterized by abrupt withdrawal response of the hindpaws with supraspinal responses, such as head turning, biting, licking, and flinching, to von Frey filament stimuli applied to the center of the glabrous surface of the hindpaw. To test the withdrawal response, the rats were individually housed in clear acrylic glass boxes (8×8×24 cm) above a metal mesh (0.5×0.5 cm), and acclimated for 15 min to avoid the stress of environmental change. Six applications of calibrated von Frey filaments, beginning with the 4.31-log unit filament [3.61 log units (0.45 g), 3.84 log units (0.74 g), 4.08 log units (1.26 g), 4.31 log units (2.04 g), 4.56 log units (3.31 g), 4.74 log units (5.50 g), 4.93 log units (8.32 g), and 5.18 log units (14.45 g)] were applied with 10-sec intervals between stimuli. The paw withdrawal response was quantified by the 50% withdrawal mechanical threshold using the formula: log (50% threshold)=Xf +κδ, where Xf=value of the final von Frey filament (log units), κ=correction factor (from a calibration table), and δ=mean differences of log units between stimuli. 15,16 The single neuronal activity of gracile neurons was measured by in vivo extracellular recording. After anesthesia (sodium pentobarbital, 60 mg/kg IP), a laminectomy exposed the medullary region including the gracile nucleus. A tracheal cannula was used for artificial ventilation, and a jugular vein cannula was used for supplemental anesthesia (sodium pentobarbital, 5 mg/kg/h, IV). The end-tidal CO2 concentration (3.5–4.5% by carbon dioxide analyzer), and body temperature (37°C maintained by homeothermic blanket control unit) were monitored. After mounting the rat on the stereotaxic frame, the head was fixed using a rat head adaptor with ear bars, and the animal was turned to expose the gracile region. Mineral oil was applied to prevent dryness of the gracile region and electrical insults from the external environment. Single-unit recordings of gracile neurons (GN; up to 500 μm from the surface) were made using carbon filament-filled glass electrodes (0.4–0.8 MΩ; Kation Scientific, Minneapolis, MN) during the application of mechanical stimuli to the receptive portions of the hindpaw. The neuronal activity was amplified, and then sent to an oscilloscope and a spike counter (Cambridge Electronic Design, Cambridge, U.K.), allowing real-time recording of peri-stimulus time histograms (PSTH), and waveforms of the spikes, using spike2 software (Cambridge Electronic Design). After isolation of a single unit, the activity was recorded of background activity (without stimulation, 20 sec); mechanical stimuli (10-sec duration at 20-sec intervals) induced by brushing the receptive area; pressure stimuli (using a clamp to produce pressure); and pinch stimuli (using a small Serrefine clamp to produces a painful sensation). The phenotypes of gracile neurons were classified according to previous studies: 17 wide-dynamic-range (WDR), low-threshold (LT), and high-threshold (HT) neurons. Neurons that showed the best response to brush stimuli (brush), or noxious stimuli (pressure and pinch), were classified as LT neurons; neurons that showed the best response to both pressure and pinch stimuli were classified as HT neurons; and neurons that showed graded response patterns to increasing mechanical intensity were classified as WDR neurons. Note that we did not record any HT neurons in the peripheral and spinal injury groups. We identified four single cells (two cells from each side of the gracile region) in each rat. Statistical analysis of the behavioral and electrophysiological data were performed using repeated-measures two-way analysis of variance (ANOVA), with the Newman-Keuls and Student's t-tests, respectively. Significance was set at p<0.05. All data are displayed as the mean±standard error of the mean (SEM).
Results
Compared to sham controls (n=2, no lesion), the sizes of the lesion in the gracile region groups (n=4) were 0.424±0.32 mm (left) and 0.449±0.32 mm (right) from the midline, which is determined by the obex and the central canal. 14 In addition, the lesion depth was 0.384±0.27 mm (left) and 0.431±0. 27 mm (right) from the surface (Fig. 1A). These gracile lesion sizes were confirmed by the rat brain atlas. 14
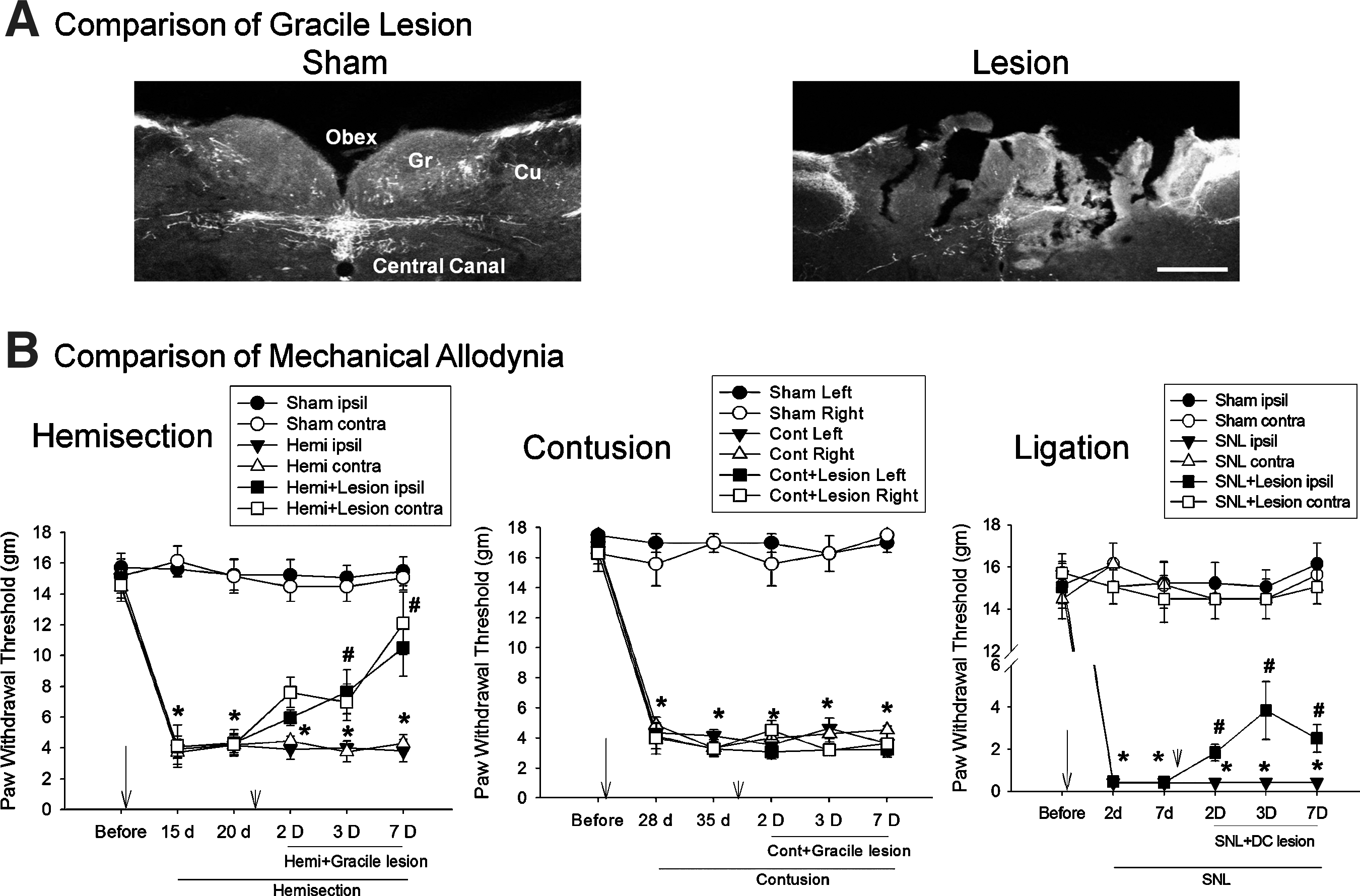
Attenuation of mechanical allodynia by lesioning of the gracile regions. (
To determine whether gracile neurons are involved in the maintenance of central or peripheral neuropathic pain, we tested the effects of gracile lesions on hemisection-, contusion-, and SNL-induced neuropathic pain behaviors. Three weeks after hemisection (HEMI, n=6), the mean withdrawal thresholds of the hindpaws were 4.3±0.9 g (ipsilateral) and 4.2±0.7 g (contralateral), and were significantly decreased compared to before injury (15.1±0.8 g and 14.5±0.9 g), or sham-injury (n=6; 15.1±0.8 g and 14.8±0.8 g) control groups (*p<0.05; Fig. 1B), indicating the maintenance of bilateral mechanical allodynia as a result of spinal hemisection. Five weeks after the contusion (Cont, n=5), the mean withdrawal thresholds of the hindpaws were 4.1±0.4 g (left) and 3.7±0.5 g (right), and were significantly decreased compared to pre-injury (17±0.6 g and 17±0.6 g; *p<0.05; Fig. 1B), indicating the maintenance of bilateral mechanical allodynia as a result of spinal contusion. Seven days after lesioning of the gracile region (n=5), however, the mean withdrawal thresholds of the hindpaws were 3.2±0.5 g (left) and 3.6±0.4 g (right), and did not show a significant increase compared to pre-lesion. Seven days after the SNL (n=7), the mean withdrawal threshold of the ipsilateral hindpaw was 0.4±0.1 g, which was significantly decreased compared to pre-injury (18±0.5 g), and sham controls (n=8; 18±0.5 g; *p<0.05; Fig. 1C). The paw withdrawal threshold of the contralateral side did not show any significant change. Two days after lesioning of the gracile regions (n=10), however, the mean withdrawal threshold of the ipsilateral hindpaw was significantly increased (2±0.5 g), compared to pre-lesion (*p<0.05; Fig. 1B).
To examine whether the electrophysiological responses of gracile neurons were changed in peripheral or central neuropathic pain conditions, we tested the excitability of gracile neurons (Table 1). The classification of WDR (Fig. 2A) and LT (Fig. 2B) neurons was determined by their response properties, which were determined by the pattern of the waveforms. 4,17 Three weeks after HEMI, the mean activities of WDR neurons (7 cells) to brush stimuli were 30.8±3.5 spikes/sec (ipsilateral) and 29.5±2.5 spikes/sec (contralateral), and showed significant increases compared to sham controls (6 cells; *p<0.05; Fig. 2C). The responses to pressure and pinch stimuli did not show any significant changes. After the HEMI, the mean responses of LT neurons (17 cells) to brush stimuli were 19.5±1.4 spikes/sec (ipsilateral) and 21.9±1.5 spikes/sec (contralateral), and showed significant increases compared to sham controls (10 cells; *p<0.05; Fig. 2C). However, we did not record evoked activity in the spinal contusion groups (Table 1). Three weeks after SNL, the mean activity of WDR neurons (7 cells) to brush stimuli was 39±4.3 spikes/sec (ipsilateral), and was significantly increased compared to sham controls (6 cells; *p<0.05; Fig. 2C), whereas pressure and pinch stimuli did not show any significant changes. The LT neurons (SNL=13 cells and sham=14 cells) also did not show any significant changes. In a comparison of spontaneous activity, the ipsilateral side of WDR neurons (7.7±1.4 spikes/sec) in the SNL group showed a significant increase compared to sham controls (3.8±1 spikes/sec, *p<0.05), whereas the HEMI groups did not show significant changes.
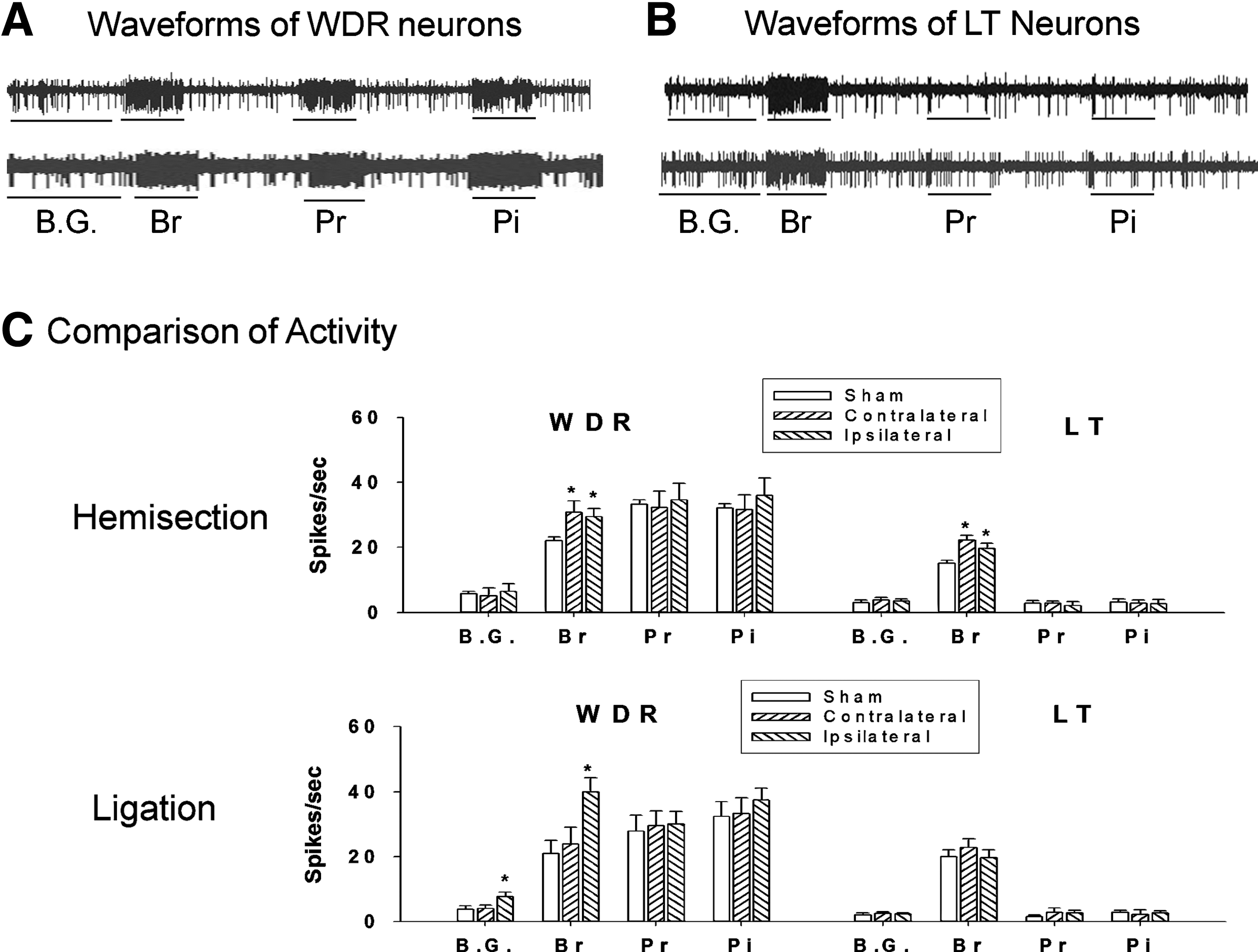
Hyperexcitability of gracile neurons after hemisection and spinal nerve ligation. The phenotypical neuronal activity of wide-dynamic-range (WDR;
The intact (sham) and unilateral injury groups (L5 spinal nerve ligation and T13 hemisection) showed responses to mechanical stimuli in both wide-dynamic-range (WDR) and low-threshold (LT) neurons in the gracile region, whereas the T10 contusion injuries did not show neuronal activity. However, no high-threshold (HT) neurons were detected in the gracile region in the unilateral and bilateral injury groups. In vivo extracellular recordings of four single units were made per rat (two units from each side of the gracile region).
Discussion
Herein we are the first to report that L5 spinal nerve ligation and T13 spinal hemisection result in enhanced activity of gracile neurons and mechanical allodynia in the hindpaw. In addition, the lesion of the gracile region attenuates mechanical allodynia in both injury groups, suggesting that gracile neurons may contribute to mechanical allodynia with non-noxious mechanical stimuli applied to the hindpaw following both types of injuries.
The peripheral neuropathic pain model with L5 spinal nerve ligation showed mechanical allodynia and increased gracile neuronal activity with non-noxious mechanical stimuli in the ipsilateral (injured) hindpaw, whereas the central neuropathic pain model with the T13 spinal hemisection showed mechanical allodynia and increased gracile neuronal activity in both the ipsilateral and contralateral hindpaws. In addition, lesions of the gracile nucleus attenuated mechanical allodynia in both the hemisection and spinal nerve ligation models. The lesion of gracile neurons in the hemisection groups produced large increases in paw withdrawal thresholds, whereas the ligation groups showed only slight increases. Although the mechanism behind the differential effect of gracile lesions on the attenuation of mechanical allodynia in the two types of injuries is unknown, a possible reason is that gracile neurons receive non-noxious tactile input mediated by DC-MLs that are relayed by myelinated primary afferent fibers from the hindpaws. The SNL does not interrupt the ascending DC-MLs in the spinal dorsal horn, whereas hemisection interrupts the ascending DC-MLs at the ipsilateral side of the spinal cord. Therefore, hemisection-induced synaptic reorganization and discharge activity probably induce greater activity of gracile neurons than ligation-induced changes. We and others have documented that SNL elicits mechanical allodynia in the ipsilateral hindpaw and neuronal hyperexcitability on the ipsilateral side of the spinal dorsal horn, whereas spinal hemisection produces bilateral mechanical allodynia in both hindpaws, and bilateral hyperexcitability of the spinal dorsal horns. 1,4 In addition, we and others have reported that hemisection results in bilateral hyperexcitability of the thalamic VPL neurons, whereas SNL produces hyperexcitability in the contralateral side of the thalamic VPL neurons. 12,13 The present study, combined with previous results, suggests that spinal hemisection produces bilateral neuropathic pain, whereas spinal nerve ligation produces unilateral neuropathic pain that runs through pathways including the spinal dorsal horn neurons, gracile neurons, and thalamic VPL neurons.
Previous studies suggest that activation of the dorsal column pathways contributes to various pain symptoms, including neuropathic, 18,19 visceral, 20 and bone pain. 21 For example, Gosselin and associates 22 reported that upregulation of GABA transporter-1 (GAT-1) with astrocytic activation results in a decrease in GABAergic inhibitory tone in gracile neurons. As a consequence, the low levels of GABAergic inhibitory tone contribute to neuropathic pain. Therefore, the neuronal-glial synaptic circuits may contribute to the hyperexcitable condition of the gracile neurons, resulting in neuropathic pain following both peripheral nerve injury and spinal cord injury. In addition, the activation (phosphorylation) of mitogen-activated protein kinase p38 (p38MAPK) in gracile neurons is associated with mechanical allodynia in the hindpaw. 23 It is well documented that glial activation, upregulation of GAT-1, and activation of the MAPK family (p-p38MAPK and extracellular signal regulated kinase), are predominant intracellular events that contribute to mechanical allodynia and neuronal hyperexcitability following peripheral nerve injury and spinal cord injury. 6,7 Therefore, pharmacological blockade of MAPK family activation or molecular approaches to prevent the upregulation of GAT-1 may be useful therapeutic tools for the attenuation of mechanical allodynia and neuronal hyperexcitability. The activation of ionotropic glutamate receptors in the gracile regions is also an important factor in the mechanical allodynia and neuronal hyperexcitability seen following peripheral and spinal cord injuries. 12,24 It has been noted that neurons overexpressing protein kinase Cγ (PKCγ) produce long-term hyperexcitation in nociceptive transmission. Approximately, 90% of gracile neurons show co-localization between PKCγ and the ionotropic glutamate receptor AMPA2/3 (GluR2/3). In addition, PKCγ-positive neurons are co-expressed with c-fos in the gracile nucleus, and are associated with mechanical allodynia after peripheral nerve injury. 25
However, there is controversy regarding the role of gracile neurons and their contribution to neuropathic pain following spinal hemisection. Previously, Kim and colleagues 26 reported that dorsal column pathways contribute to the induction, but not the maintenance, of mechanical allodynia following spinal hemisection in rats. The Kim group bilaterally transected the spinal cord at T8 to interrupt the dorsal column pathways. In the present study, we used a direct lesion at the gracile regions, and showed that gracile neurons contribute to the maintenance of neuronal hyperexcitability and mechanical allodynia in hindpaws with non-noxious mechanical stimuli after hemisection injury. It is interesting that bilateral spinal contusion injuries, such as spinal contusions, did not result in the appearance of hyperexcitable neurons at the gracile regions. However, we and others consistently reported that bilateral spinal cord injuries, such as spinal contusion injury, produce mechanical allodynia in both hindpaws, and hyperexcitability of spinal dorsal horn neurons and thalamic VPL neurons, suggesting that STTs, and not DC-MLs, are critical contributors to neuropathic pain following spinal contusion injury. 27,28 Taken together, the present data suggest that the intact (contralateral) side of the spinal dorsal horn mediates bilaterally-enhanced nociceptive transmissions to the gracile regions, which result in mechanical allodynia in the hindpaws and hyperexcitability of the gracile neurons following spinal hemisection. 29 However, the neuroanatomical and neurochemical changes in both the supraspinal regions and gracile neurons following spinal hemisection require further study.
In conclusion, the present data suggest that unilateral neural injuries, such as spinal hemisection and spinal nerve ligation, produce hyperexcitability of the gracile neurons and contribute to the maintenance of neuropathic pain after non-noxious stimuli.
Footnotes
Acknowledgment
This work was supported by the National Research Foundation of Korea, funded by the Korean government (MEST no. 2011-0030124)
Author Disclosure Statement
No competing financial interests exist.