
Abstract
The majority of clinical spinal cord injuries (SCIs) are contusive and occur at the cervical level of the spinal cord. Most scientists and clinicians agree that the preclinical evaluation of novel candidate treatments should include testing in a cervical SCI contusion model. Because mice are increasingly used because of the availability of genetically engineered lines, we characterized a novel cervical hemicontusion injury in mice using the Infinite Horizon Spinal Cord Impactor (Precisions Systems & Instrumentation, Lexington, KY). In the current study, C57BL/6 mice received a hemicontusion injury of 75 kilodynes with or without dwell time in an attempt to elicit a sustained moderate-to-severe motor deficit. Hemicontusion injuries without dwell time resulted in sustained deficits of the affected forepaw, as revealed by a 3-fold decrease in usage during rearing, a ∼50% reduction in grooming scores, and retrieval of significantly fewer pellets on the Montoya staircase test. Only minor transient deficits were observed in grasping force. CatWalk analysis revealed reduced paw-print size and swing speed of the affected forelimb. Added dwell time of 15 or 30 sec significantly worsened behavioral outcome, and mice demonstrated minimal ability of grasping, paw usage, and overground locomotion. Besides worsening of behavioral deficits, added dwell time also reduced residual white and gray matter at the epicenter and rostral-caudal to the injury, including on the contralateral side of the spinal cord. Taken together, we developed and characterized a new hemicontusion SCI model in mice that produces sufficient and sustained impairments in gross and skilled forelimb function and produced primarily unilateral functional deficits.
Introduction
W
During the past decade, there has been intensive research into molecular and cellular factors determining neurological recovery after SCI in the hope of finding an effective therapeutic strategy. 3 –6 Genetically engineered mouse models that permit (conditional) expression or inactivation of genes have served as valuable in vivo tools to test molecular pathomechanisms and provide proof of principle for promising therapeutics. 7 –10 Though many SCI studies in mice have been performed at the thoracic level, few have involved cervical lesions of the spinal cord. 11 –15 It is noteworthy that findings in thoracic injury models may not be directly applicable to injuries at the cervical level. Thoracic SCIs produce behavioral deficits by damaging long axons of ascending and descending tracts in the white matter. Little functional abnormalities observed can be attributed to loss of gray matter, such as interneurons and/or motor neurons at the level of injury.
A number of models of traumatic or atraumatic cervical SCI have been previously reported on, in which the spinal cord is injured by contusion, dislocation, distraction, compression, crush, sharp transection, excitatory neurotoxins, or irradiation injuries. 16 –28 Though partial and complete transection models are useful to study axonal regeneration, 29,30 these models reflect the small percentage of humans that suffer a sharp, penetrating SCI. Besides, the mechanism of recovery can also vary as a result of the type of injury sustained. 31,32 Additionally, after contusion injuries, a rim of spared axons persists adjacent to the injury site that is affected by secondary degeneration, which may result in ongoing neural damage. For this reason, the contusion model is more suitable for the investigation of neuroprotective therapies, unlike the transection model, where there are no spared fibers. 32
Among the current injury models available, blunt contusion and compression injuries best represent the pathophysiologic process observed in human SCI. 30,33,34 According to a recent survey of the SCI research community, 72% of the 324 respondents agree that contusion injury is currently the most clinically relevant injury model of SCI. 34 A number of mouse SCI models at the cervical level have been developed, including a model of cervical spondylotic myelopathy, sharp axotomy models of the right lateral dorsal funiculus, and complete lateral spinal hemisections. 16,20,25,35,36 However, to date, no attempts have been made to characterize a cervical hemicontusion injury in mice. It has been shown in rats that unilateral cervical injuries produce forelimb deficits ipsilateral to the side of injury while sparing the function of the forelimb contralateral to injury. 28,33,37 This would significantly reduce the need for chronic intensive care after SCI, as observed with a bilateral cervical contusion injury in mice. 38
The Infinite Horizon Spinal Cord (IH) Impactor (Precisions Systems & Instrumentation, Lexington, KY) is a commercially available SCI contusion device. Initially, the IH Impactor was developed for use in a thoracic SCI model and has rapidly become widely utilized in the SCI research community. Recently, Lee and colleagues 28 described the systematic development of a cervical unilateral contusion model using the IH Impactor in rats. In the current study, we extended this model in C57/Bl6 mice with behavioral and histopathological outcome assessments. Besides evaluating trauma resulting from contusive injury, we also examined the influence of continued compression on injury by increasing the programmable dwell time from 0 to 15 and 30 sec.
The injury parameters established in these experiments generated a primarily unilateral injury model with sustained functional deficits over 12 weeks. This injury model will provide an additional preclinical tool that can be used to test molecular hypotheses in transgenic (Tg) mice and evaluate promising therapeutic agents before moving to clinical trials.
Methods
All animal procedures were performed in accord with the guidelines of the Canadian Council for Animal Care and approved by the University of British Columbia Animal Care Committee (Vancouver, British Columbia, Canada). All behavioral and histological analyses were performed by a person blinded to the treatment.
Animals and general housing conditions
Adult male C57BL/6 mice were purchased from Charles River Laboratories (Wilmington, MA) and housed in the central animal facility, with room temperature (RT) controlled at 21°C and an artificial 12-h light/dark cycle (lights off, 9:00
Preoperative care
Mice were initially anesthetized with a mixture of ketamine hydrochloride (72 mg/kg) and xylazine hydrochloride (9 mg/kg). To achieve a surgical plane of anesthesia, animals were additionally anesthetized by inhalation with isoflurane (2%; O2 1L/min). To alleviate pain, buprenorphine was injected (subcutaneously [s.c.]; 0.03 mg/kg; Temgesic; Schering-Plough Corporation, Kenilworth, NJ) and the eyes were lubricated with an eye gel (Liposic® Ophtalmic Liquid Gel) to prevent desiccation. In addition, animals were s.c. injected with 1 mL of lactated Ringer's (Hospira Inc., Montreal, Quebec, Canada) to replace blood lost during surgery. Skin in the neck region was shaved and disinfected with betadine and 70% ethanol.
Left C5 hemicontusion
C5 hemicontusion injury was performed as previously published for rats. 28 Animals were placed in a stereotaxic frame (David Kopf Instruments, Tujunga, CA), and a 1-cm dorsal midline skin incision over the upper cervical area was made. Connective and muscle tissue were dissected to expose the posterior vertebral elements from C3 to C6. A laminectomy was performed on the left side at C5 to expose the spinal cord. To stabilize the spinal column, the C3–C6 spine was held with a custom-built clamp (material, polyether-ether-ketone [PEEK]; McMaster-Carr, Elmhurst, IL) to firmly grasp the vertebrae (Fig. 1A–C). The clamp was then positioned in a frame that was tilted at a 22.5-degree angle to the horizontal plane. 28 With the animal positioned in the clamping system, the impactor tip (diameter, 0.75 mm with rounded edges; material, carbon steel) was aimed at the gray matter of the dorsal spinal cord between the C5 and C6 dorsal nerve roots (Fig. 1D). The impactor was then triggered to deliver a set force of 75 kilodynes (kdyn) at 100 mm/s, followed by 0, 15, or 30 sec of continued compression (i.e., dwell time; n=8, n=6, and n=7, respectively). The IH Impactor device performed the dwell times as it was programmed to produce, and only accurate dwell times of 15 or 30 sec were measured. A zero dwell time implies that the impactor was programmed to not add any time in addition to the turnaround time of several milliseconds, which is a constraint as a resulf of the inertia of the tip.

Spinal cord clamping system to prevent movement of the vertebral column on impact. (
After perfusion and exposure of the spinal cord, injury level was anatomically verified by counting the dorsal roots during dissection. All the impacts were confirmed to be between the C5 and C6 nerve roots.
Postoperative care
Muscle was sutured with coated Vicryl 5-0 (Ethicon, Markhem, Ontario, Canada). Mice were kept in a temperature-controlled incubator set at 32.0°C until animals regained their ability to move around freely, after which they were placed back in a recovery cage. To alleviate postoperative pain and prevent dehydration, buprenorphine (0.03 mg/kg) and 1 mL of lactated Ringer's (Hospira Inc.) was given s.c. twice-daily for an additional 2 days. After surgery, an easily accessible supply of moisture and nutrients, such as Transgel and Ensure
Grooming
After tap water was applied to the animal's head and back with soft gauze, the animal was placed in a plexiglas cylinder (15 cm in diameter and 15 cm in height). 33 Grooming activity was recorded with a video camera for a total time of 15 min. Frame-by-frame video playback was used to score each forelimb independently by the maximal area contacted during grooming 33 : 0=the animal was unable to contact any part of the face or head; 1=the animal's forepaw touched the underside of the chin and/or mouth area; 2=the animal's forepaw contacted the area between the nose and eyes, but not the eyes; 3=the animal's forepaw contacted the eyes and the area up to, but not including, the front of the ears; 4=the animal's forepaw contacted the front, but not the back, of the ears; and 5=the animal's forepaw contacted the area of the head behind the ears.
Cylinder rearing
Forelimb usage during spontaneous vertical exploration was done as described previously. 26,39,40 Briefly, animals were placed in a clear Plexiglas® cylinder (15 cm in diameter and 15 cm in height) for 15 min per session. During the session, animals were videotaped, and a frame-by-frame analysis of forelimb usage during 20 independent rears was done after.
Usage of orepaws for support during rearing was scored as either “ipsilateral, “contralateral,” or “both.” Forelimb usage of the affected paw was represented as the “percentage of ipsilateral forelimb usage,” calculated by adding the number of times the animal placed either the ipsilateral only or with both paws simultaneously and dividing this by the total number of paw placements.
Grip-strength test
Forelimb grip strength was measured using an automated grip-strength meter (Linton Instrumentation, Diss, Norfolk, UK), as described previously. 38,41 During the pretraining period, mice were trained to grip a mesh bar attached to the grip-strength meter apparatus. On each trial, the animal was held by the base of the tail and gently pulled in a backward direction. Over 7 days, mice were allowed to grip with both forepaws, after which a 1-week baseline grip-strength assessment was performed. For each animal, maximum peak grip strength (g) at which the mesh was released was measured during 10 trials. After injury, grip-strength data were conducted weekly for a total of 5 weeks (maximum peak force during 10 trials).
CatWalk
CatWalk automated gait analysis was used to quantify locomotion. 42 CatWalk measurements were calculated from five runs during uninterrupted locomotion at a consistent speed. Different gait parameters related to affected front and hind paw pairs were analyzed, including print area, print width, print length, swing speed, base of support, and stance duration. Print area is the surface area (pixel) of the complete paw print. Print width is the width (vertical direction) of the complete paw print during stance. Print length is the length (horizontal direction) of the complete paw print. Swing speed is the speed (pixels/sec) of the paw during the swing phase. Base of support (BOS) is the average width between either the front or hind paws. Stance is the duration of contact with the glass plate in a step cycle.
Montoya staircase
Forelimb reaching performance was assessed using the Montoya staircase test, scaled down for mice based on the design previously published for rats. 43 –45 During this test, animals freely enter a narrow translucent Plexiglas corridor and are allowed to reach down for food pellets placed within eight wells of a descending “stair case” (Dustless Precision Pellets® chocolate; 20 mg; 45% fat, 34% carbohydrates, and 11% protein [%w/w]; Bio-Serv, Frenchtown, NJ).
Before injury, mice were first acclimatized to the apparatus by placing food pellets on the staircase steps in their home cage on 3 consecutive days preceding the start of the training session. Subsequently, animals were trained individually on the staircase task for seven sessions of 15 min (one session per day; two pellets per well). To motivate animals to grasp for food, mice were food-deprived for 20 h, having access to standard lab chow for only 4 h per day (12:00
During baseline and postinjury assessment, the numbers of pellets remaining on each well during 30 min were counted. Pellets were color-coded according to the level of the well on the staircase to keep track of which pellets were obtained during scoring. Success rates were measured as number of pellets eaten or maximum distance reached (i.e., the lowest step from which the mouse managed to retrieve a pellet). Because animals could use their tongue to consume pellets from the top two steps, reaching performance was only assessed for the bottom six steps (numbered 3–8; maximum, 12 pellets).
Spinal cord tissue processing
After intracardiac perfusion with 0.1 M of phosphate-buffered saline (PBS; 15 mL) and ice-cold 4% paraformaldehyde in PBS (30 mL), the spinal cord was post-fixed overnight and cryoprotected in graded concentrations of sucrose (12, 18, and 24%) in phosphate buffer. Twenty-micrometer coronal serial sections were cut on a cryostat (Microm International GmbH, Heidelberg, Germany) and mounted onto Superfrost/Plus slides (Fisher Scientific, Fair Lawn, NJ).
Immunofluorescence and lesion area quantification
Blocking of nonspecific proteins was accomplished by incubating the slides in 10% normal donkey serum (Jackson ImmunoResearch Laboratories, West Grove, PA) in 0.1% PBS/Triton X-100 for 30 min. Sections were incubated overnight at RT in mouse anti-GFAP (glial fibrillary acidic protein) antibody (Ab; 1:500; Sigma-Aldrich, St. Louis, MO) and rabbit anti-fibronectin (1:200; Dako, Glostrup, Denmark). After incubation with DyLight-conjugated secondary Abs (1:200; Jackson ImmunoResearch Laboratories, West Grove, PA), slides were stained with Hoechst (1:5000) and cover-slipped (Fluoromount-G; Southern Biotechnology Associates, Birmingham, AL).
Each slice was digitally photographed using a Zeiss AxioPlan2 imaging microscope (Zeiss, Inc., Thornwood, NY) equipped with a QImaging camera (QImaging, Burnaby, British Columbia, Canada). Lesion area (mm2) was manually traced using imaging analysis software (SigmaScan Pro5; Systat Software Inc., San Jose, CA) and was defined as a central GFAP-negative area of the spinal cord.
Eriochrome cyanine staining
To analyze spared white and gray matter, coronal sections through the cervical injury site were stained with Eriochrome cyanine (EC) to visualize myelinated white matter. Frozen spinal cord sections were rehydrated through graded ethanol solutions (twice in toluene, twice in 100%, once in 95%, and then once in 70 and 50% ethanol). After rehydration in dH2O, slides were stained in EC Solution (0.16% EC, 0.4% sulphuric acid, and 0.4% ferric chloride) at RT for 10 min and gently rinsed in dH2O. Slides were differentiated in 0.5% ferric ammonium sulphate at RT for 2 min, rinsed in dH2O, dehydrated in graded ethanol solutions (once in 50, 70, and 95% ethanol and two changes in 100% ethanol), cleared in xylene two times, and cover-slipped using Entallan mounting medium (EM Science, Gibbstown, NJ).
Each slice was digitally photographed (Zeiss AxioPlan2 imaging microscope; Zeiss) and manually traced using imaging analysis software (SigmaScan Pro5; Systat Software). The amount of total white-matter sparing was based on positive staining for myelin visualized with EC staining. Spared gray matter was defined as tissue containing normal gray matter cytoarchitecture based on EC staining.
Statistical analysis
All data are presented as the mean±the standard error of the mean (SEM). Because our data are not normally distributed and the sample sizes are rather small (<10), Kruskal-Wallis' nonparametric test was used, followed by Dunn's post-hoc testing. Spared white and gray matter for the ipsilateral, as well as for the contralateral, side were summed as a measure of total spared tissue. Pearson's R-value was used to evaluate correlations between total spared tissue and behavioral scores. Statistical significance was set at p≤0.05.
Results
Injury parameters
No significant differences were detected between groups for the actual force, displacement, or velocity measured. The average force in the 0-, 15-, and 30-sec dwell groups was, respectively, 79±1.0, 79.8±1.2, and 80.1±1.2 kdyn. An average displacement of, respectively, 830.6±24.5, 791.3±39.5, and 844.1±35.1 μm was measured for the 0-, 15-, and 30-sec dwell groups. The average velocity measured with the different dwell time settings was 122.9±0.4 (0-sec), 122.7±0.3 (15-sec), and 123.9±0.5 mm/sec (30-sec).
General health
After SCI, all injured animals demonstrated a clubbed ipsilateral forepaw the first few days after injury days. Daily observation of mice in their home cage revealed some transient hindlimb deficits and slight postural instability. This hindlimb impairment was typically observed early after SCI and was absent at 2 weeks postinjury. Before and after injury, mice were weighed weekly as a gross measure of general health (Fig. 2). After an initial drop of ∼16% during the first 3 days, the body weight of the 0-sec dwell group gradually increased and returned to preoperative baseline values by 14 days after surgery. Increasing dwell time from 0 to 15 or 30 sec affected the time course of body-weight recovery considerably (Fig. 2). Mice of both groups displayed an initial ∼24% decline in body weight after injury. By day 5, animals progressively gained weight and recovered their preoperative weight by 12 weeks. However, at the end of the experiment, body weights of animals subjected to prolonged dwell times were still significantly below that of the 0-sec dwell group (0-sec dwell, 27.6±0.4; 15-sec dwell, 25.7±0.4; 30-sec dwell, 25.5±0.4).
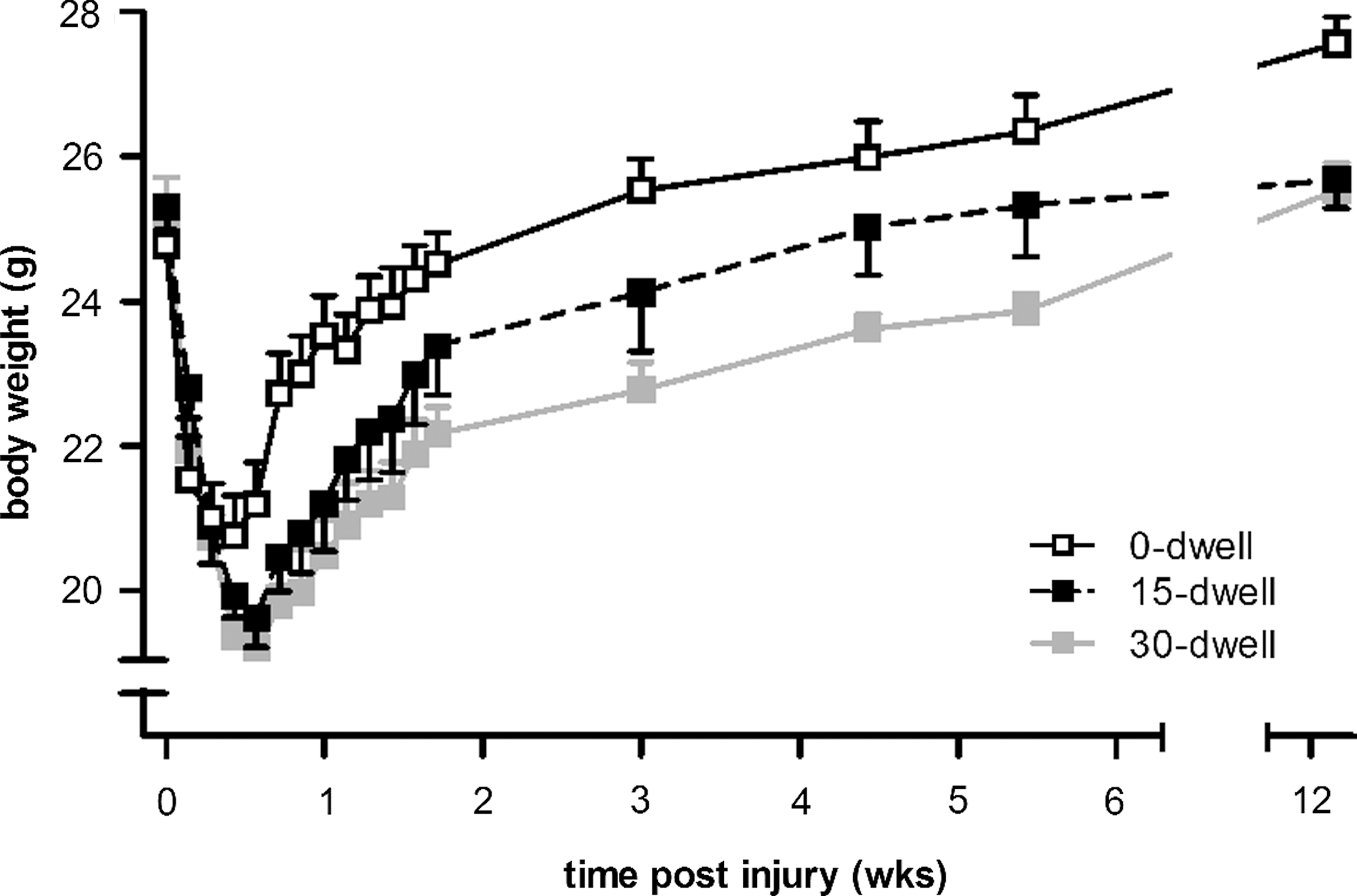
Body-weight change of mice receiving a C5 contusion injury without or with dwell time. During the first 3 days, body weight dropped significantly more if the contusion injury was followed by 15 or 30 sec of dwell time, compared to animals receiving no dwell time (0-dwell). At 12 weeks postinjury, the average body weight of both dwell time groups was still ∼7% below that of weight levels of the 0-sec dwell time group. No differences were observed between the 15- or 30-sec dwell groups. Error bars indicate SEM. *p≤0.05. SEM, standard error of the mean.
Cylinder rearing
The cylinder rearing task is effective in examining forelimb use for postural support and vertical exploration (Fig. 3). Before injury, animals typically use both limbs equally during wall placements, that is, in ∼20% of placements, they use only the right or only the left paw independently (Fig. 3A), and in ∼60%, both paws are placed simultaneously on the wall (Fig. 3B). All groups showed a near-complete loss in independent usage of the affected (ipsilateral) paw during rearing in the weeks after injury (Fig. 3A) and used the ipsilateral paw mostly in combination with the contralateral paw (Fig. 3B). Compared to preinjury levels, the 0-sec dwell group displayed a ∼40–60% reduction in ipsilateral forelimb usage during rearing events (Fig. 3B). The addition of dwell time worsened the outcome considerably (p<0.01), and at 2 weeks, very little usage of the ipsilateral paw was observed (Fig. 3B). Over time, a slight improvement was noted in usage of the ipsilateral paw, and at 12-weeks postinjury, the differences between the three injury groups were no longer significant (Fig. 3B).

Forelimb usage during rearing in C5 hemicontused mice without or with dwell time. (
Grooming test
Compared to preinjury levels, all rats showed lower grooming scores with the ipsilateral paw after contusion injury (Fig. 4A). At 2 weeks postinjury, animals in the 0-sec dwell group were able to raise their affected paw up to eye level (score 3) without any further improvement over time. Grooming deficits were more pronounced when the contusion injury was followed by a dwell time of 15 or 30 sec (p<0.003; Fig. 4A). After injury, animals were, at most, capable of touching the underside of the chin (score 1), which was maintained until the end of the study independent of length of dwell time (Fig. 4A).
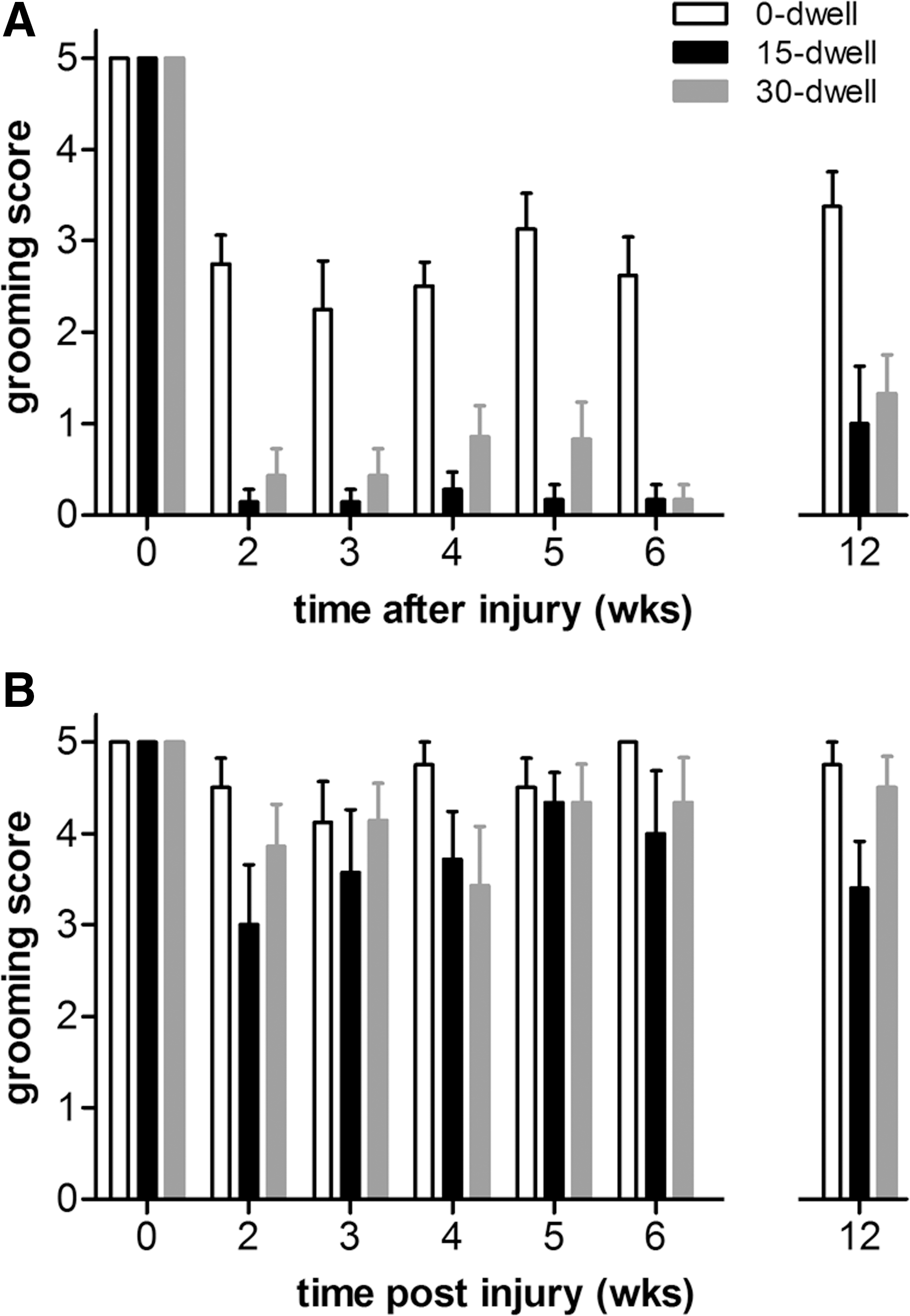
Face grooming scores in mice after C5 hemicontusion without or with dwell time. (
For the contralateral paw, maximum grooming scores were observed for both pre- and postinjury time points. Some animals showed slight postural instability during the first 2 weeks post-SCI, which may have contributed to their grooming deficit of the contralateral paw early after injury. No significant differences were observed between groups for the contralateral paw (Fig. 4B).
Grip-strength test
After injury, mice were assessed for grip-strength performance (Fig. 5), commonly used to evaluate forelimb muscle strength as an indicator of neuromuscular function. 46,47

Forepaw grip strength from SCI mice after C5 hemicontusion injury without or with dwell time. Two weeks postinjury, the maximal gripping force (g) exerted by both forepaws declined in all three groups. Over time, however, animals in the 0-sec dwell time group showed recovery to preinjury baseline levels. Animals in 15- and 30-sec dwell groups showed no recovery of their dual grip strength, and at week 12 postinjury, dual forepaw grip strength was ∼50% of preinjury levels. Dashed lines represent the grip strength from mice that pulled the bar with their contralateral forepaw only. Error bars indicate SEM. *p≤0.05. SCI, spinal cord injury; SEM, standard error of the mean.
Postinjury, all mice in the 0-sec dwell group gripped with both forepaws and recovered dual forepaw grip strength comparable to pre-SCI baseline levels (0-dwell preinjury, 96.0±4.2 g; week 12 postinjury, 101.1±8.6 g; Fig. 5). Contusion, followed by 15 and 30 sec of static compression, resulted in a significant deficit in the strength test, compared to injury without additional compression (p<0.001; Fig 5). Initially, these animals were completely unable to use their affected forepaw to grip, which resulted in a level that was ∼50% of preinjury levels (15-sec dwell, 48.0±4.8; 30-sec dwell, 51.0±7.8; Fig. 5). Although, over time, these animals gradually regained the ability to use the impaired paw to grasp, grip strength remained severely affected over a period of 12 weeks independent of compression time (15-sec dwell, 55.4±5.7; 30-sec dwell, 63.8±3.6).
Montoya staircase test
In the Montoya staircase test, animals reach from a central platform with their forelimbs to retrieve food pellets from eight descending steps with shallow wells. 43 –45 The pellets in lower wells are more difficult to grasp than those in wells higher on the staircase. Analysis of video recording revealed that mice were able to use their tongue to retrieve pellets from the top two steps. Therefore, reaching was only scored for the bottom six steps (numbered 3–8; maximum, 12 pellets).
Mice were trained for 6 days on the Montoya staircase test to ensure baseline scores for all animals that reached a plateau (usually achieved within four sessions). Uninjured mice retrieved approximately five pellets before injury (0-sec dwell, 5.25±0.75; 15-sec dwell, 5.00±0.61; Fig. 6A). Typically, pellets placed below well number 6 were beyond reach (Fig. 6B). No overall difference in performance was observed between groups.
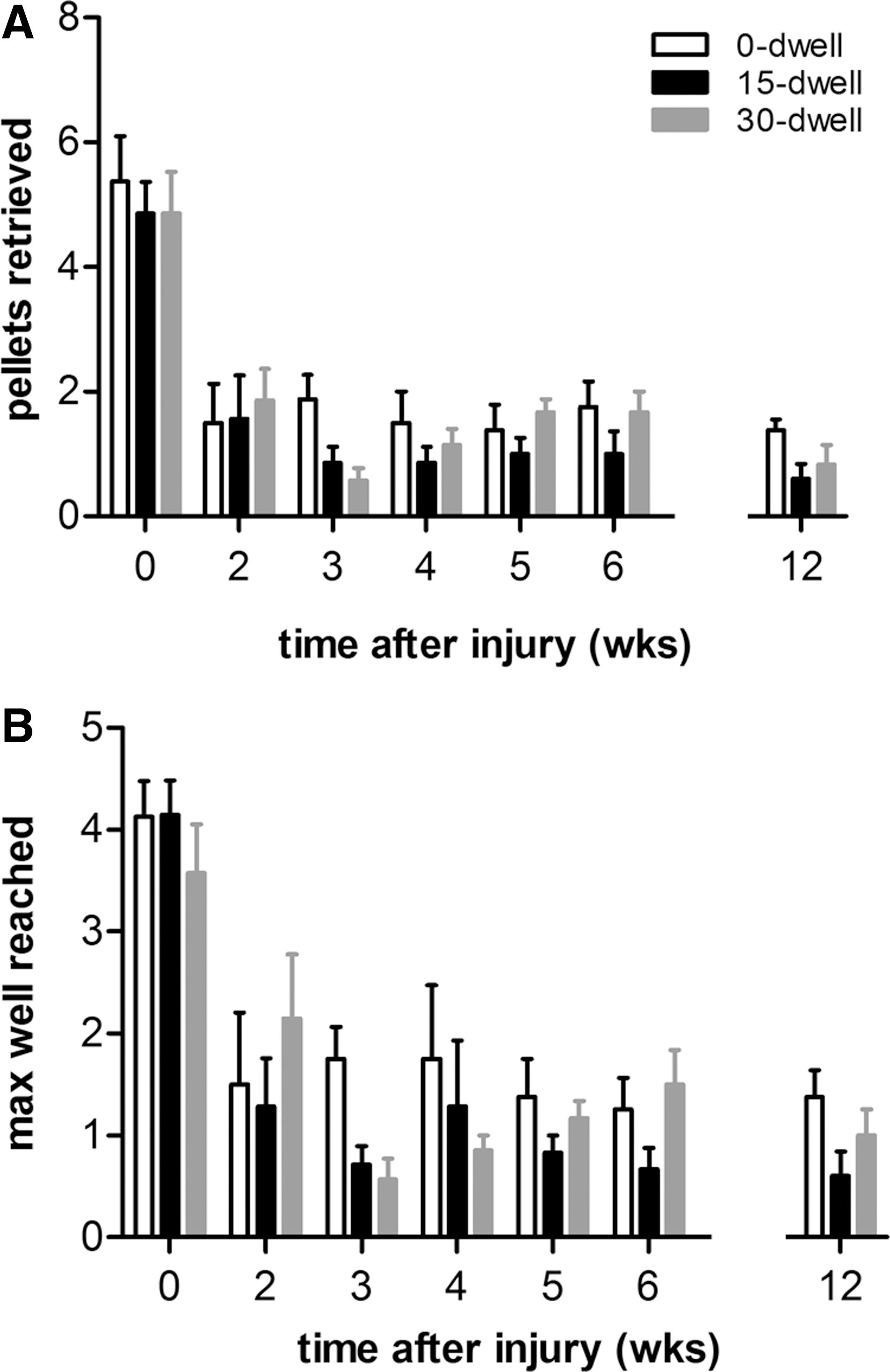
Effect of C5 hemicontusion without or with dwell time on Montoya staircase skilled reaching performance. (
After C5 hemicontusion, mice showed severe impairments not only in retrieval success with the ipsilateral paw, but also in maximum distance reached. At 2 weeks after SCI with or without additional dwell time, mice demonstrated reaching scores ∼60% of baseline values, which remained stable until 12 weeks postinjury (0-sec dwell, 1.38±0.18; 15-sec dwell, 0.6±0.24; 30-sec dwell, 0.83±0.31; Fig. 6A). Animals could only reliably retrieve pellets from well number 3 and sometimes well number 4 (Fig. 6B). No differences in time course and extent of reaching deficit were observed between groups.
Besides an ipsilateral deficit in paw reaching, the number of pellets consumed on the contralateral side was also significantly reduced, when compared to preinjury levels. At 12 weeks postinjury, the number of pellets eaten successfully collected with the contralateral paw for the 0-, 15-, and 30-sec dwell groups was, respectively, 2.00±0.65, 1.17±0.41, and 0.91±0.42. No differences were observed between groups.
CatWalk analysis
After hemicontusion injury, an overall decrease was observed in area, width, and length of the complete paw print during stance in all animals (Fig. 7A,B). In the 0-sec dwell group, print area decreased from 27.8±0.53 to 13.8±2.03 mm2 at 2 weeks after contusion injury, with almost no recovery during the 12-week observation period (Fig. 7A). Analysis further revealed that the reduction in print area observed after contusion resulted from a decrease in both print width and length (Fig. 7B,C). Further, during the swing phase of the affected front paw, a ∼40% drop in average speed of the moving limb could be detected (Fig. 7D).
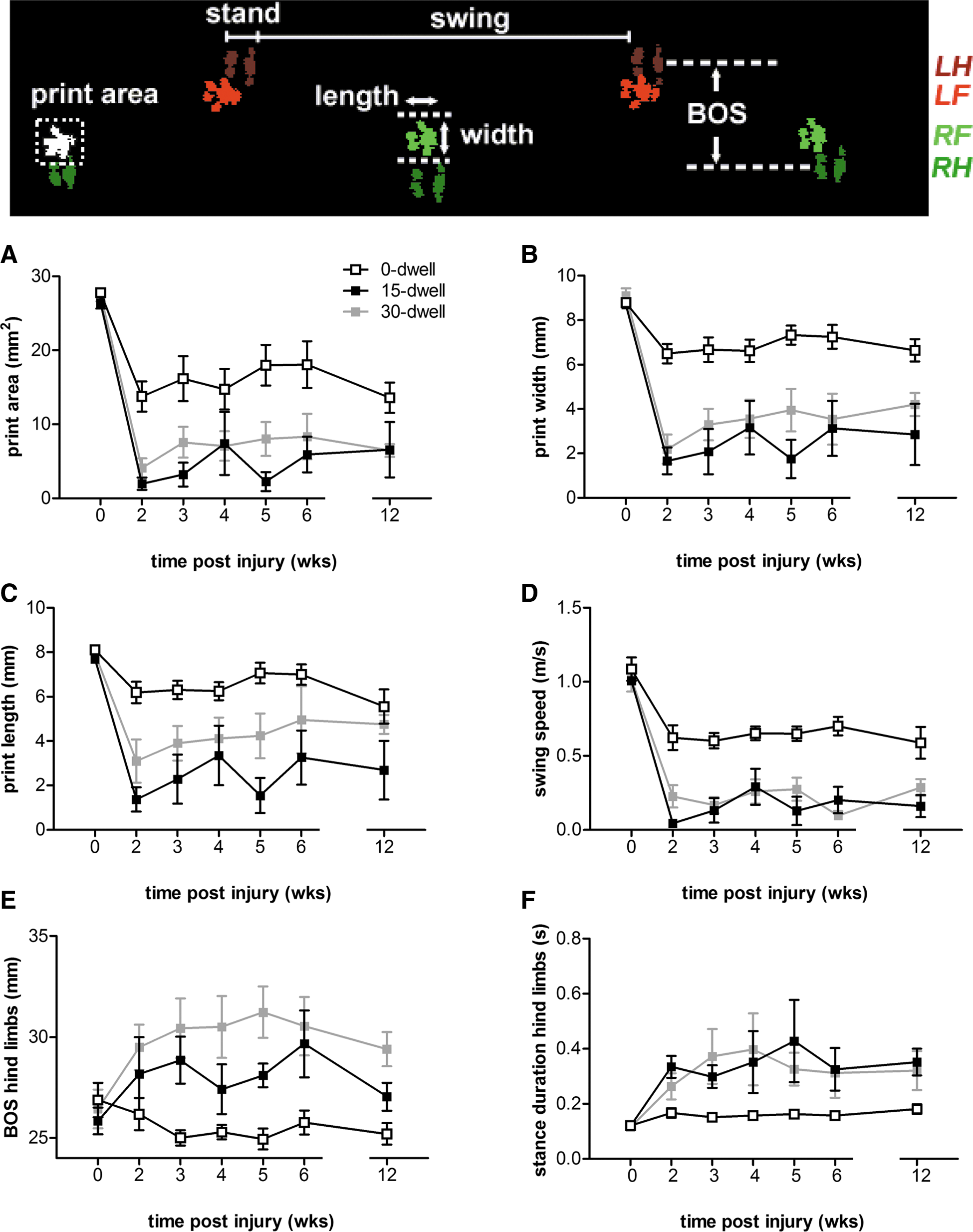
CatWalk analysis of paw parameters during overground locomotion. Upper panel: foot prints (LF, left front paw; RF, right front paw; LH, left hind paw; RH, right hind paw) digitized by CatWalk software measuring print area, print length, print width, and BOS during the stand phase of the step cycle. (
In both the 15- and 30-sec dwell groups, print area was reduced to less than 10% of its preinjury value (Fig. 7A; 12 weeks postinjury; 15-sec dwell, 6.57±3.73; 30-sec dwell, 6.45±0.86). Both the width and length of the affected front paw was generally smaller than that of mice in the 0-sec dwell group (Fig. 7B,C). However, differences in print length disappeared at the later time points investigated. In addition, CatWalk analysis showed a marked decrease in swing speed of the affected paw, which was significantly lower, when compared to the 0-sec dwell group.
Analysis revealed that both BOS and stance duration of the hind limbs were significantly affected by dwell time (Fig. 7E,F). Both groups (15- and 30-sec dwell) showed a compensatory increase in both parameters of the hind limbs, when compared to SCI animals without dwell time (Fig. 7D). A larger BOS and/or longer stance duration of the hind limbs can compensate for an unstable gait caused by the affected forepaw. Notably, CatWalk footprints revealed that in 60–71% of the animals in both dwell time groups, the affected forepaw was not always detected during the step cycles. In the 0-sec dwell time group, this absence of the ipsilateral forepaw print was only observed in 1 of 9 animals (11%). Although BOS may also depend on the size of the animal, 48 no correlation between body weight and hindlimb BOS was present.
Lesion area
After SCI, mice do not develop cystic lesions typically observed in rat SCI; however, the lesion does get filled with dense fibrous connective tissue (as illustrated in Fig. 8). 49,50 This fibrous structure contains an extracellular matrix rich in fibronectin-positive material, surrounded by GFAP-positive cells (Fig. 8). GFAP staining was used to outline the lesion edge in the injured spinal cords of our mice. At the epicenter, the average lesion area of the 0-, 15-, and 30-sec dwell groups encompassed, respectively, 46.9±5.3, 44.9±8.3, and 48.4±2.9% of the hemicord ipsilateral to the injury (Fig. 9A). The lesion involved both gray and white matter and extended for ∼400 μm rostral and caudal to the lesion epicenter (Fig. 9A). Although the primary damage was observed ipsilateral to the injury, a small percentage of GFAP-negative area was observed on the contralateral side (Fig. 9B). There was no difference in GFAP-negative lesion area between the groups (Fig. 9A–C).

GFAP and fibronectin immunostaining of mice spinal cords after C5 contusion injury without or with dwell time. Representative pictures of spinal cord cross-sections at week 13 postinjury showing the outline of the lesion epicenter stained with fibronectin (red) and GFAP (green). Note the increased lesion area characterized by the dense fibronectin expression and GFAP-negative area when the contusion injury is followed by 15- or 30-sec dwell time. The lesion area is defined as a central GFAP-negative region of the spinal cord. GFAP, glial fibrillary acidic protein. Color image is available online at

Quantification of the GFAP-negative lesion area after spinal cord contusion injury with or without dwell time. (
Gray and white matter sparing
To evaluate changes in spared gray and white matter area, spinal cord sections were stained for Eriochrome/neutral red to visualize myelin. Tissue damage in white and gray matter after a 75-kdyn contusion injury was predominately confined to the ipsilateral side (Figs. 10 and 11). At the epicenter, damaged tissue included parts of the dorsal and ventral funiculus medially and parts of the lateral funiculus (Fig. 10). The dorsal and ventral horns of the gray matter were entirely damaged (Figs. 10 and 1A). Areas of spared tissue included a thin spared rim in the ventrolateral funiculus (Figs. 10 and 1B). Spared white matter at the injury epicenter (i.e., ipsilateral) for the 0-, 15-, and 30-sec dwell groups was, respectively, 0.35±0.058 (range, 0.24–0.59), 0.32±0.070 (range, 0.19–0.50) and 0.26±0.024 mm2 (range, 0.16–0.37 mm2) (Fig. 11B).

Eriochrome/neutral red-stained spinal cord sections of C5 hemicontused mice without or with dwell time. Photomicrographs of representative spinal cord cross-sections stained with myelin-specific EC at the epicenter and 200-μm increments rostral-caudal to the epicenter. At week 13 postinjury, EC-stained sections demonstrated large lesions on the ipsilateral side around the epicenter region, with a varying amount of remaining gray matter and a peripheral rim of spared white matter. Note the reduced residual white and gray matter contralateral to the injury side in both the 15- and 30-sec dwell time groups. In the contusion-only group (0-sec dwell), damage was mainly contained to the ipsilateral side. EC, Eriochrome cyanine. Color image is available online at

Tissue sparing quantification of spinal cord tissue after C5 contusion injury without or with dwell time. Measurement of the caudal-rostral extent of ipsilateral (
Although the extent of spinal cord damage did not extend more than approximately 600 μm along the rostrocaudal axis of the epicenter (Fig. 10), gray and white matter area was significantly reduced in all SCI groups, compared to the corresponding segmental level of uninjured control spinal cords (Fig. 11A,B). These results suggest that contusion injury significantly reduces ipsilateral spinal cord size. This phenomenon was not observed for the contralateral site (Fig. 11C,D).
Although no differences were obtained in extent of tissue damage to the ipsilateral side (Fig. 11A,B), it was observed that dwell time of both 15 and 30 sec caused extensive white and gray matter damage at the hemicord contralateral to injury (Fig. 10C,D). Spinal cords of mice that received an extended 30-sec dwell time resulted not only in greater contralateral tissue damage at the epicenter, but also at 220, 440, and 660 μm distance rostrally from the epicenter of the injury (Fig. 11C,D).
Correlation between motor function recovery and pathophysiology
The amount of spared tissue at the epicenter was plotted against week 12 postinjury behavioral results, as assessed with the cylinder rearing test, grooming test, and grip-strength test (Fig. 12). To determine whether there was a linear relationship between behavioral tasks and histological assessment, Pearson's correlation coefficient was calculated (R-value; Fig. 12). Linear regression indicated a significant positive correlation between the amount of spared tissue at the epicenter and their performance in the cylinder rearing test, grooming test, and grip-strength test. There was no correlation between tissue sparing and staircase performance.
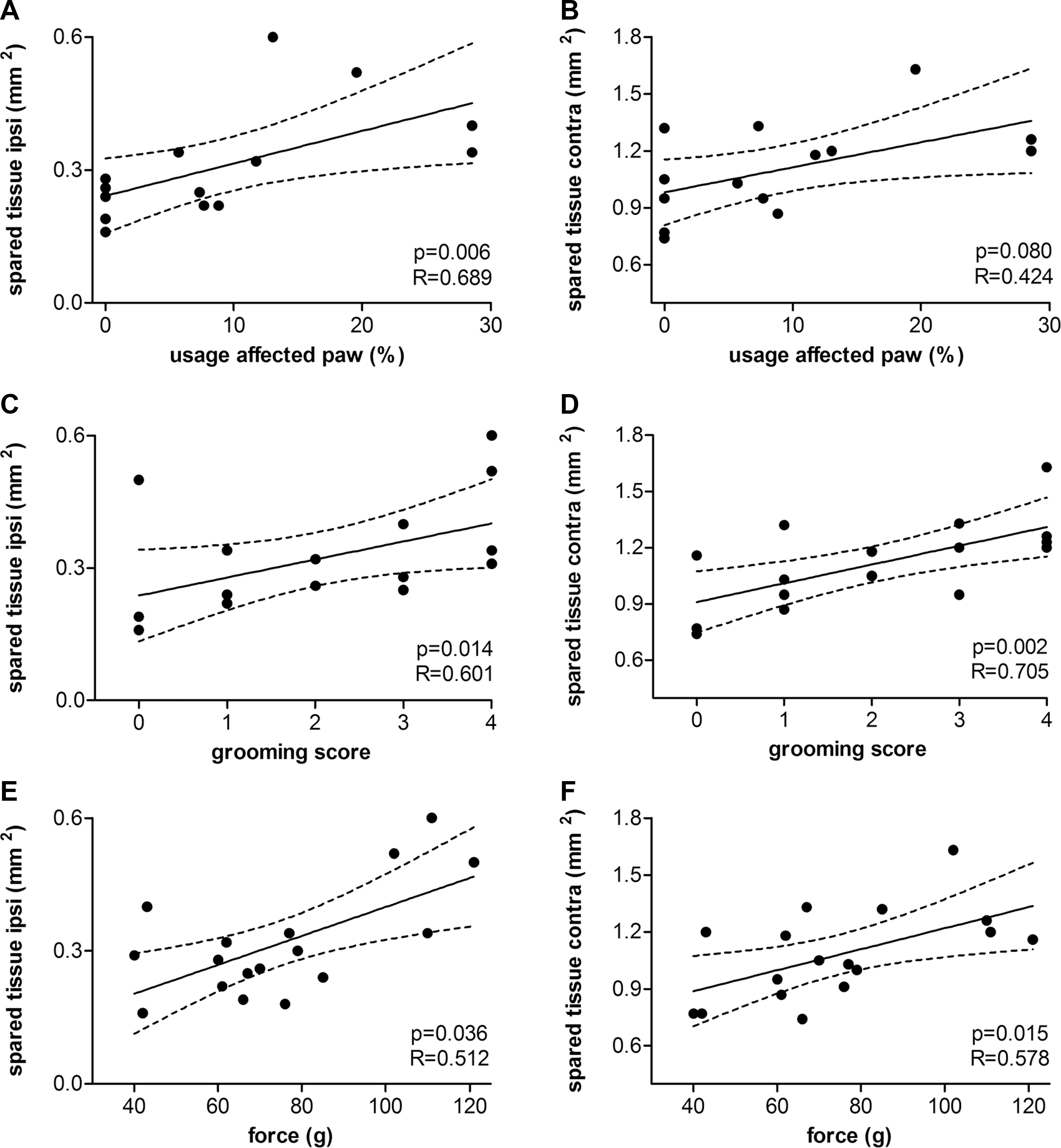
Pearson's correlation and linear regression analysis between behavioral and histological outcomes. Linear regression was observed showing a significant positive correlation between (
Discussion
In this article, we describe the development of a mouse cervical hemicontusion model using the Infinite Horizon Spinal Cord Impactor at a set force of 75-kdyn. With this setting, we were able to produce sustained behavioral deficits in the ipsilateral forelimb, with parenchymal damage largely contained to the ipsilateral side. There are numerous upper extremity tasks proven effective for assessing various aspects of motor impairment and recovery after SCI. We selected a battery of tests to examine fine and gross motor components of the overall forelimb functions, including the grooming test, cylinder rearing, Montoya staircase, grip-strength test, and CatWalk performance. Animals had clear deficits in forelimb usage during wall placements, grooming, as well as overground locomotion. Additionally, we observed that transient spinal cord compression (i.e., dwell time of 15 and 30 sec) after initial injury worsened ipsilateral forelimb function and resulted in parenchymal damage past the midline to the contralateral side. Given that there were obvious functional differences between injuries with or without dwell time shows the importance of this lesion extension to the opposite side. Although dwell time itself was associated with worsened outcomes, duration was not (at least not between 15 and 30 sec).
Development of this model occurred after a series of pilot studies. With respect to the target area of injury, we intended to damage both the corticospinal (CST) and rubrospinal (RST) tracts because these both play an important role in forelimb function in rodents. 51 In a pilot study, we observed that a 75-kdyn hemicontusion injury with the animal in neutral rotation (i.e., vertical impact) did not produce significant functional forelimb deficits, as assessed with the cylinder rearing and grooming tests. Further, the lateral nature of the injury appeared to have spared the dorsal CST, which may have contributed to the minimal functional impairments in these animals (data not shown). Strategies of increasing impact forces to 100 kdyn, however, resulted in low survival rates of ∼25% as a result of respiratory complications. We hypothesized that by angling the trajectory of the impactor, we would be more likely to damage both the CST and RST and therefore induce a more substantial and sustained functional impairment, as we previously reported in rats. 28 Continued spinal compression of 15 or 30 sec was applied in an attempt to produce various degrees of severity, because static compression early after onset has been shown to exacerbate damage and hindlimb function in a rat model of thoracic SCI. 52 –54
In our hands, we experienced some variability in holding the spinal column firmly with the clamps provided with the IH device and therefore preferred to design a custom-built clamping system scaled down to mice that firmly grasps the transverse processes of the cervical spine, as previously described. 27,28 The inability to hold animals firmly with the clamps provided with the IH device and to prevent subtle movement at the time of injury should, theoretically, not influence the injury mechanics significantly, because the impactor is designed to impart the same force each time. However, a previous pilot study indicated that even with a force-controlled impactor, subtle movements during injury caused bybreathing or clamp failure resulted in aberrant degrees of parenchymal damage and unpredictable functional deficits (J.H.T. Lee, personal communication).
Interestingly, mice injured without dwell time exhibited recovery of grip strength, whereas mice with added dwell time exhibited more-permanent deficits. The rodent grip-strength test was introduced more then 30 years ago to screen for neurobehavioral toxicity. 46,47 Currently, it has also been used as an accepted measure of forelimb muscular strength after SCI. 38,55 It has been recognized, however, that other nonmotor changes, such as loss of or failure to gain body weight, can further contribute to forelimb weakness without evidence of central nervous system lesions. 56,57 Though this could be a possible explanation for the decline in grip-strength force in both dwell time groups, this is highly unlikely because the degree of recovery of body weights in these mice did not coincide with recovery of gripping ability.
Spinal cord compression after injury did not result in a more severe reaching deficit in the Montoya staircase test, as indicated by the equivalent number of pellets retrieved in spinal-cord–contused animals without and with dwell time. Although the magnitude of the reaching deficit was similar between groups, unpublished preliminary data from our lab suggests that the underlying nature of the impairments might be different. It was observed that during cereal manipulation with the affected forelimb, the elbow of the majority of animals receiving additional dwell time were either fully extended or only partially flexed (complex movements associated with cereal manipulation is recently described by Irvine and colleagues). 58 Spinal-cord–contused animals without added dwell time, however, demonstrated only subtle deficits in elbow and shoulder motion.
The loss of these functions after contusion injury are likely related to a combination of both axonal disruption of sensomotor pathways and to the direct abolition of motor neurons, second-order sensory neurons, and interneurons at C5, which directly innervate proximal and distal forelimb muscles, such as the deltoid (shoulder), biceps (upper forelimb), extensor carpi radialis longus (dorsal surface of the upper forelimb), extensor carpi radialis brevis (dorsal surface of the lower forelimb), and flexor carpi radialis (ventral surface of lower forelimb). 59,60 Because there was a clear relationship between the degree of total spared tissue at the injury site and function of the impaired forelimb, we suggest that the diminished recovery of grooming, rearing, and gripping performance after dwell time could be the results of differences in the extent of the lesion. High correlations exist with most of the behaviors, but not all tasks. We found no correlation between the amount of gray- and white-matter sparing and forelimb reaching abilities using the Montoya staircase paradigm. One possibility is that the extent of parenchymal damage as a result of contusion injury may be beyond the threshold for development of compensatory mechanisms, independent of dwell time.
Besides worsening of behavioral deficits, compression time reduced residual white and gray matter at the epicenter and rostral-caudal to the injury. This suggests a distinct vulnerability of white and gray matter to compression injury to the spinal cord. Changes in local strains and stresses within the microenvironment during transient compression may have contributed to a greater degree of both neuronal and microvascular disruption. Others have shown that the physical properties of spinal gray and white matter are highly distinctive, with enhanced capillary density 61 and reduced tolerance to compression before blood–spinal cord barrier disruption in gray, compared to white, matter. 62 Intraparenchymal hemorrhage and hemorrhagic necrosis is a direct consequence of trauma and has a typical pattern of distribution into the contused spinal cord, spreading from the central region; this may involve several segments of the spinal cord. Hemorrhage has been shown to positively correlate with injury severity and has also been found to be predictive of functional outcomes. 63 –67 Further, recent work by Simard and colleagues 68 suggests that, because of the anisotropy of spinal cord tissues, the same force can generate large differences in severity of neurological injury, with severity correlating with amount of primary hemorrhage. These investigators showed that differences in apparent injury severity were the result of the specific location of the impact, which determined whether the primary hemorrhage would be uni- or bilateral. 68
In addition, post-traumatic ischemia and the presence of inflammatory cells leads to the increased production of free radicals, resulting in oxidation of lipids, proteins, and nuclei acids, degradation of cytoskeletal proteins, and mitochondrial dysfunction, thereby exacerbating tissue damage. 69 –73 Thus, compression subsequent to impact may be multi-factorial, triggering specific molecular and cellular events that evolve into secondary injury mechanisms, which exacerbated the injury.
The less-severe forelimb impairments without dwell time might, in part, depend on spared residual neurons and axons within the area of partial damage. There is evidence that the RST in the lateral column and the CST, which are important for distal flexor function, can compensate for each other, to a certain degree, after injury. 74,75 Further, spared fibers of the ventral and dorsal portion of the CST ipsilateral to and/or sprouting of corticospinal midline-crossing projections can contribute to improved behavioral recovery after SCI. 76 –79 The relationship between CST and RST damage in our model and the functional impairments is only postulated. Retro- and anterograde tracing studies would be needed to quantify RST and CST sparing (if any) and correlate these with forelimb function to strengthen this association.
In summary, considering that, clinically, the most common SCI occurs at a cervical level resulting from a contusive impact, we decided to develop a mouse cervical unilateral injury model. Although there are various cervical contusion injury models available for rats, hitherto, there has been no unilateral cervical contusion injury model characterized for mice. In this study, we describe the development of a clinically relevant cervical unilateral contusion injury using the IH Impactor. A force of 75 kdyn without additional dwell time was generally well tolerated by the animals and produced consistent, sustained functional deficits over 12 weeks with characteristic parenchymal damage to the spinal cord. This injury model will provide an additional preclinical tool that can be used to perform mechanistic proof-of-principle’ experiments in Tg mice. This will enable significant steps in the understanding of pathophysiology after SCI as well as the molecular mechanisms underlying promising therapeutic agents before moving to clinical trials.
Footnotes
Acknowledgments
This research was supported by The Craig Neilsen Foundation (to W.T.), The Christopher and Dana Reeve Foundation (to F.S.), and the Canadian Institutes for Health Research (to W.T). W.T. holds the Edie Ehlers Chair in Spinal Cord Research funded by the Rick Hansen Foundation. The authors thank Miss Roya Elahi and Grace Kim for their help on data analysis. The authors also thank Miss Lauren Whitehead for her invaluable help with organizing and analyzing the data. Further, the authors thank Tamara Godbey and Juan Felipe Molina in the Central Animal Facility for their help and advice with animal care.
Author Disclosure Statement
No competing financial interests exist.