
Abstract
A peripheral indicator of the presence and magnitude of brain injury has been a sought-after tool by clinicians. We measured neuron-specific enolase (NSE), myelin basic protein (MBP), and S100B, prior to and after scaled cortical impact in immature pigs, to determine if these purported markers increase after injury, correlate with the resulting lesion volume, and if these relationships vary with maturation. Scaled cortical impact resulted in increased lesion volume with increasing age. Concentrations of NSE, but not S100B or MBP, increased after injury in all age groups. The high variability of S100B concentrations prior to injury may have precluded detection of an increase due to injury. Total serum markers were estimated, accounting for the allometric growth of blood volume, and resulted in a positive correlation of both NSE and S100B with lesion volume. Even with allometric scaling of blood volume and a uniform mechanism of injury, NSE had only a fair to poor predictive value. In a clinical setting, where the types of injuries are varied, more investigation is required to yield a panel of serum markers that can reliably predict the extent of injury. Allometric scaling may improve estimation of serum marker release in pediatric populations.
Introduction
I
NSE (EC 4.2.1.11) is a glycolytic enzyme existing as isoenzyme homodimers (γγ and αα), and heterodimers (αγ), with the γγ and αγ dimers being highly abundant in the brain and at low levels in all tissues. 6 Measurement of NSE in the peripheral serum is only expected to occur after both release of intracellular NSE due to structural damage of the cell membrane, and disruption of the blood–brain barrier, although severe damage to internal organs can also result in detectable serum concentrations of NSE. 6,7 Though the half-life (t1/2) of NSE is hypothesized to be between 24 and 48 h, to our knowledge, no actual kinetic studies have been performed. 2,4,7
S100B is a 10-kD protein and is a member of a large class of EF hand calcium binding proteins produced and secreted by a variety of glial cells in the central and peripheral nervous systems (CNS and PNS), and is also present in many tissues, including bone, myocardium, and adipose. Within the CNS, astrocytes are the main source of S100B. S100B usually exists as homodimers or heterodimers with S100A1, and measurement of S100B alone indicates all states of S100B. 8 Secreted S100B binds to the receptor for advanced glycation end products, inducing autocrine actions and paracrine actions on neurons. 9 At physiological concentrations (nanomolar), S100B induces neurite outgrowth, prevents apoptosis, increases the rate of neurogenesis, and enhances cognition after TBI. 9 –11 Conversely, high concentrations of S100B (micromolar) secreted during pathological conditions (e.g., Alzheimer's disease), stimulate proinflammatory cytokines and apoptosis. 9,11 Sustained elevations of S100B in the serum are likely due to disruption of the blood–brain barrier, 12 but can occur with or without brain injury. In blood, S100B has a t1/2 of 25 min, 13 and is detectable in healthy neonates (1.8 μg/L) and children (0.3 μg/L), with concentrations declining in adulthood (0.1 μg/L). 14
Classic MBP is a heavily post-translationally modified 18.5-kD protein needed for the tight wrapping of the cytoplasmic membrane of oligodendrocytes around axons, and transduction of intracellular signaling cascades. 15 During development, myelination begins mid-gestation and continues through adolescence, with myelin containing a variety of splice isoforms of MBP transcribed from genes of oligodendrocyte lineage, resulting in proteins that range widely in size (14 to 21.4 kD) and function. 15,16 In healthy individuals, MBP would not be expected to be detected peripherally, and indicates disruption of the blood–brain barrier.
Although serum concentrations of NSE, S100B, and MBP are purported to positively correlate with clinical severity and/or pathoanatomic magnitude of injury, prediction is mainly limited only to very adverse outcomes such as death or severe disability in those with a low Glasgow Coma Scale score (GCS). 5,8,17 –19 The success of NSE, S100B, and MBP in predicting outcome in children with mild or moderate TBI (GCS score ≥9) has been varied. 2,4,5 Furthermore, the magnitude of mechanical forces and types of TBI (focal versus diffuse; limited versus extensive damage) in much of the clinical research of these serum markers are unknown. As an additional consideration, measurements of serum markers expressed as concentrations may not optimally estimate biomarker release, because estimation of the amount released from the injured brain is the desired information, rather than estimation of the hormone's action on tissues. This is particularly pertinent in growing subjects in whom there is allometric growth of blood volume, and the volume of blood relative to body weight decreases with age in both humans and piglets.
The objectives of this experiment were to determine whether (1) concentrations of NSE, S100B, or MBP in the peripheral blood 7 days following scaled cortical impact correlate with histologic lesion volume on day 7 after injury; (2) concentrations of serum markers following injury are age-dependent; and (3) accounting for allometric changes in blood volume increases the predictive value of serum markers in indicating the presence and extent of TBI.
Methods
Animal care and use
Sixty-seven male and female Yorkshire swine (Earle Parsons & Sons, Inc., Hadley, MA) were housed in a temperature-controlled facility (21–22°C) with a 12-h light/dark cycle with lights on at 0700 h. Piglets less than 28 days old were given access to a heated farrowing pad and milk replacer (Grade A® Ultra 24™; Milk Products LLC, Chilton, WI) ad libitum. One- and 4-month-old pigs were given pig and sow pellets (Blue Seal, Londonderry, NH) twice daily. Twelve hours prior to surgery, 1- and 4-month-old pigs were removed from feed, and 5- to 7-day-old piglets were removed from milk replacer and given a liquid electrolyte solution (BlueLite; TechMix, Inc., Stewart, MN). All protocols and procedures were in accordance with the guidelines of the American Veterinary Association and the National Institutes of Health (NIH), and were approved by the Animal Care and Use Committee at Dartmouth College, and aimed to limit the number of animals used and animal distress.
Scaled cortical impact
Swine aged 5 to 7 days, 1 month, or 4 months, in which the stages of brain development and anatomy parallel human infants, toddlers, and pre-adolescents, 20 underwent scaled cortical impact under isoflurane anesthesia as described previously. 20,21 Briefly, the swine were sedated with midazolam and anesthesia was induced with 5% isoflurane and maintained with 1–2% isoflurane and 21% oxygen (room air). Atropine and buprenorphine were administered, and the pigs were intubated and mechanically ventilated. Heart rate, blood pressure, end-tidal CO2, oxygen saturation, and body temperature were monitored and maintained within a narrow range appropriate to subject age. 21 A craniectomy was performed at the junction of the right coronal and sagittal sutures, and the dura was removed to expose the cortical surface. Scaled cortical impact was achieved via a spring-loaded device secured to the skull, and an indentor tip scaled to the size of the brain for each age of animal, such that 1% of the volume of the brain was displaced over 4 msec. This resulted in clinically-silent lesions located on the rostral gyrus, which is the somatosensory cortex corresponding to the snout. The mechanical input (strain) was thus identical among ages, since strain is the parameter best correlated with resultant histologic injury. 20
Blood collection and ELISAs
Venous blood was collected after anesthetic induction prior to craniectomy and at 15 min, 1 day, 4 day, and 7 days after scaled cortical impact. For the 1-, 4-, and 7-day post-injury blood collections, swine aged 5–7 days and 1 month were briefly anesthetized with 1–2% isoflurane, and swine aged 4 months were sedated with ketamine and xylazine (6.3–8.3 mg/kg; 1.3–1.7 mg/kg IM), followed by 1–2% isoflurane. Blood was collected in EDTA-coated Vacutainers® (Becton, Dickinson and Company, Franklin Lakes, NJ1), and spun at 3000g within 15 min of collection. Plasma was divided into aliquots and stored at −80°C until analysis.
In subjects with a complete set of samples (5–7 days, n=18; 1 month, n=11; 4 months, n=18; samples collected at least pre-injury, and on days 1 and 4), concentrations of S100B, NSE, and MBP, were determined by commercially available sandwich enzyme-linked immunosorbent assays (ELISAs) per the manufacturer's instructions in non-hemolyzed samples (Point of Care Division of Nanogen, Toronto, Ontario, Canada). The intra-assay coefficients of variation and inter-assay coefficients of variation, respectively, were as follows: NSE 5.7% and 17.1%, S100B 4.4 % and 11.8 %, and MBP 11.8% and 8.2%. The NSE ELISA used murine monoclonal antibodies raised against the γ subunit of human NSE, allowing the assay to detect both common isoforms, and had a detection limit of 0.6 ng/mL. NSE is a product of the ENO2 gene, and is highly conserved among species, consistent with other glycolytic enzymes with a sequence similarity of 97% between rats and humans. 22 Sus scrofa ENO2 (GenBank accession number XM_001925729.3; National Center for Biotechnology Information [NCBI]) has 93% sequence similarity with ENO2 in Homo sapiens (accession number BT007383.1), as determined by the Basic Local Alignment Search Tool (BLAST®, NCBI). Assays employing antibodies generated against human NSE are successfully used to determine serum concentrations of NSE in rodents and piglets. 23 –25 Recently, in a model of cardiac arrest in piglets, NSE was measured using antibodies generated against porcine NSE, resulting in concentrations of NSE similar to the values reported here. 26 The S100B ELISA used murine monoclonal and polyclonal rabbit antibodies raised against the whole human S100 ββ protein, and had a detection limit of 0.01 ng/mL. The porcine S100B gene has not been sequenced, but the amino acid sequence of the Ca2+-binding portion of the S100B protein (cd05027) is 96% similar between Homo sapiens (1MQ1_A) and Bos taurus (1CF_A), indicating that this domain is highly conserved among mammalian species. The MBP ELISA used polyclonal goat antibodies and murine monoclonal antibodies against the human 18.5-kD classic MBP protein, and had a detection limit of 0.2 ng/mL. A portion of the gene encoding MBP has 85% sequence similarity between Homo sapiens (accession number NM_002385) and Sus scrofa (accession number XM_003360586). Using this ELISA, MBP was increased in serum from piglets after rapid non-impact rotational injury (data not shown), demonstrating that detection of porcine MBP is not limited using this ELISA.
Lesion analysis via histology
On day 7 after scaled cortical impact, the swine were deeply anesthetized with 5% isoflurane, transcardially perfused with 0.9% saline followed by phosphate-buffered formalin (10%), and the brains were collected. Processing and analysis of the tissue was performed as previously described.
21
Briefly, 5-mm coronal slabs were paraffin-embedded, and 10-μM sections were stained with hematoxylin and eosin. For each subject, sections corresponding to the 0, 25, 50, 75, and 100th percentile of the area underlying the indentor tip were analyzed by a single neuropathologist who delineated the lesion.
21
For each subject the lesion size was averaged for the five sections, and taken as a percentage of the mean of the five sections of the corresponding uninjured contralateral hemisphere, allowing comparison of lesion size across ages. To estimate lesion volume for correlations of a serum marker to lesion volume, Cavalieri's estimator of morphometric volume (Vc) by the following equation was used:
Statistical analysis
To assess whether lesion size increased with age, and if serum marker concentrations varied prior to injury, increased over time after injury, or varied at each time point after injury when compared among ages or genders, several tests were used; p values < 0.05 were considered significant. First, the effects of age and gender, and the interaction of lesion size, estimated lesion volume, and the concentrations of NSE, S100B, and MBP at each time point were determined by a two-way analysis of variance (ANOVA) using SAS software (version 9.1.4; SAS Institute Inc., Cary, NC). There was no main effect of gender on concentrations of NSE, S100B, or MBP, lesion size, or estimated lesion volume, and therefore the genders were combined. The effects of age and time and their interaction with serum marker concentrations were tested using the repeated measures function procedure of SAS, followed by post-hoc t-tests when the interaction was significant comparing age groups at specific time points. Finally, the same was done after estimating total NSE or S100B, taking into account the allometric growth of blood volume.
We also assessed whether there was a correlation between estimated lesion volume and the peak serum marker concentration for each individual within each age group, as well as among age groups, using Pearson correlation coefficients. Finally, we assessed whether correlations between peak NSE or S100B and estimated lesion volume emerged when using allometric scaling for blood volume.
Using previously published data on milliliters of blood per kilogram of body weight measured for pigs of various weights, 27 a segmented quadratic equation for 5- to 7-day-old and 1-month-old piglets (y=106+(−1.29*kg)+0.0012*kg2; r2=0.992), and linear regression equation for 4-month-old piglets (y=85.68+(−0.32*kg); r2=0.863) was fitted using SigmaPlot® 11.2 (Systat Software, Inc., San Jose, CA), to estimate the total blood volume per subject based on known body weights (Table 1). The estimated blood volume per individual was then multiplied by the nanograms per milliliter of serum marker to determine the total estimated serum marker. Concentrations of serum markers that do not adjust for differences in blood volume among ages are expressed as concentrations (ng/mL), while serum markers for which the allometric growth of blood is accounted for are expressed as total estimated serum marker (ng).
From Bush et al. 27
To determine if NSE (both concentrations and total estimated) could indicate the presence of cortical impact, the pre-injury values were used as true negatives, and cut-off values, specificity, and sensitivity for NSE were calculated using Microsoft Excel, and receiver-operator curves were generated by plotting sensitivity versus 1 – specificity for four cut-off values to determine area under the curve using SigmaPlot software. Qualitatively, the predictive value of the serum marker for indicating cortical injury was scored using the area under the curves by the following system: 0.90–1=excellent, 0.80–0.89=good, 0.70–0.79=fair, 0.60–0.69=poor, and 0.50–0.59=fail.
Results
Lesion pathology
In our model of cortical impact in which the mechanical strain created by the impact is equivalent among ages, lesion size scaled to the size of the brain increased progressively with age, presumably reflecting altered susceptibility and/or repair mechanisms as a function of age, as previously reported. 21 In the present study, both estimated lesion volume (Fig. 1A), and lesion size scaled to the contralateral hemisphere, increased with increasing age (Fig. 1B).
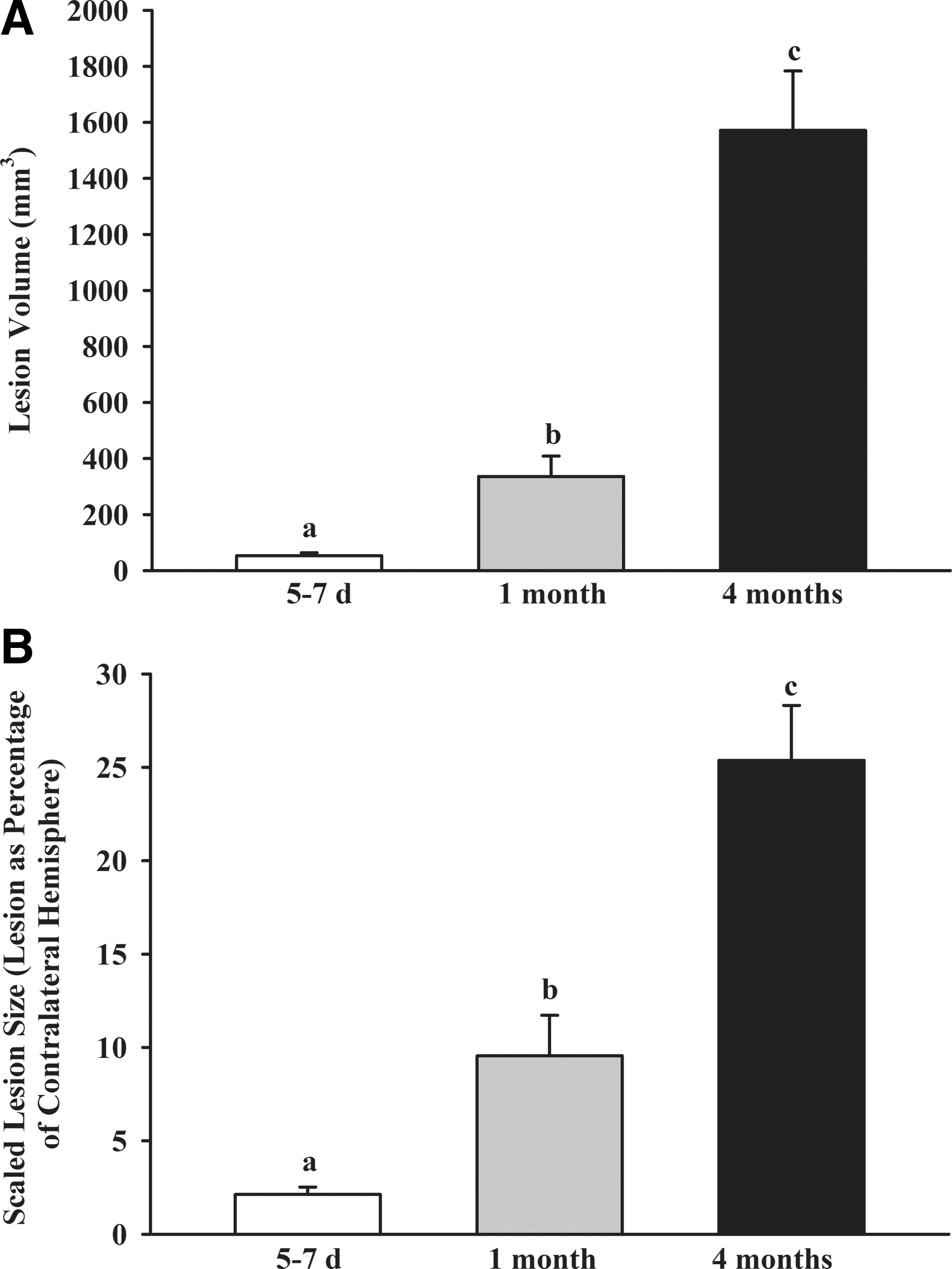
Lesion volume (
Neuron-specific enolase
There was a significant effect of time (F4,148=26.77, p<0.0001), and the time×age interaction (F8,148=3.76, p<0.0001), on the concentrations of NSE before and after injury, with concentrations of NSE that were greater on day 1 after injury compared to pre-injury in all age groups (Fig. 2). Prior to injury, concentrations of NSE did not vary among ages, but despite the large difference in lesion size at day 7 among age groups, there was no difference in the concentrations of NSE among age groups at day 1 (p>0.05, Fig. 2). Individually, the increase of NSE in response to injury was consistent, as only 3 of 45 of individuals failed to exhibit a numerical increase. Though concentrations of NSE increased after injury, they were not correlated with lesion volume at day 7 among all age groups or within age groups.

Neuron-specific enolase (NSE) prior to, and 15 min, 1, 4, and 7 days after scaled cortical impact in piglets aged 5–7 days, 1 month, and 4 months. (
When total NSE was estimated accounting for allometric growth of blood volume (Table 2), estimated total NSE increased in all age groups from pre-injury to day 1. Total estimated NSE was no different among age groups prior to injury, but at day 1 it was lowest in 5- to 7-day-old piglets, intermediate in 1 –month-old piglets, and greatest in piglets aged 4 months, the age group with the largest lesion volume (Figs. 1 and 2). Total estimated NSE tended to be positively correlated with estimated lesion volume when compared among all age groups (r=0.27, p=0.07), and also when compared among individuals within the 5- to 7-day age group (r=0.68, p=0.02), but not within the 1- or 4-month age groups.
The cut-off values of NSE chosen were values below the level considered negative (no injury), and the level above that considered positive (presence of injury), to determine specificity and sensitivity. Taking into account the allometric growth of blood volume by estimating total NSE did not increase the predictive value of NSE, though using age-specific cut-off values did improve the area under the curve.
Receiver operator curves were generated for concentrations of NSE (ng/mL) and total estimated NSE, and taking into account the allometric growth of blood volume (total estimated NSE). These changes did little to alter specificity, sensitivity, or the area under the curve (Table 2). The predictive value of NSE to indicate the presence of cortical impact was slightly improved by age-specific cut-off values, but only had fair predictive value at best.
S100B
Concentrations of S100B did not increase after injury in any age group (Fig. 3), likely due to the high variation in S100B concentrations among subjects prior to injury, particularly in the 4-month age group, in which 4 of 16 individuals had higher S100B concentrations prior to injury than at any time point after injury. Concentrations of S100B after injury did not correlate with lesion volume on day 7 after injury. When total S100B was estimated, it did not increase after injury (Fig. 3), but it was positively correlated with estimated lesion volume among all age groups (r=0.7, p<0.001), but not within any age group.

S100B prior to, and 15 min, 1, 4, and 7 days after scaled cortical impact in piglets aged 5–7 days, 1 month, and 4 months. Neither concentrations of S100B (
Myelin basic protein
In this model of scaled cortical impact in immature subjects, concentrations of MBP did not differ over time in any age group (5–7 days: 0.16±0.05 versus 0.29±0.09 ng/mL; 1 month: 0.04±0.05 versus 0.02±0.07 ng/mL; 4 months: 0.17±0.11 versus 0.16±0.05 ng/mL).
Discussion
In this model of scaled cortical impact in immature subjects, we sought to measure proteins previously evaluated as serum markers in children to characterize our model and to determine if purported markers of TBI correlate with lesion volume at day 7 after injury. To our knowledge, this is the first report measuring candidate serum markers prior to and after injury, comparing the serum marker to the volume of the resulting lesion in any species. When evaluated on a concentration basis, only NSE (ng/mL) was elevated after injury, peaking on day 1. Neither concentrations of S100B nor MBP were elevated in response to cortical impact. However, when the allometric growth of blood volume was taken into account, correlations emerged between lesion volume and both total estimated NSE and total estimated S100B.
Developmental changes in organ size and fluid compartment size are well characterized in both swine and humans. 27,28 However, to our knowledge, there has been little discussion of the influence of these factors on serum marker estimation after TBI, though allometric estimation of the pharmacokinetics of anesthetics in the blood compartment in infants and children has been performed (y=1.2×body weight1.1). 29 As a first attempt, we used a single-compartment pharmacokinetic model, for which the concentration of serum marker is directly proportional to the administered “dose,” and inversely proportional to its volume of distribution. The distribution volume of marker in this single-compartment model is assumed to be the blood volume, which changes in relation to body weight developmentally in piglets, as it does in infants and toddlers. 27,28 We also assumed an instantaneous release of serum marker from the brain, complete mixing within the blood volume, and equivalent clearance of the serum markers among ages, though the actual process of release, distribution, and clearance is likely more complex than we have assumed.
In the current study, using a cut-off value of 1.2 ng/mL for NSE, we found that cortical impact and concentrations peaked at 24 h in all age groups in a pattern similar to that seen in children. In children, NSE levels over 11.36 ng/mL indicate TBI, and they peak 8–40 h after injury, with an earlier peak after accidental TBI (higher mean GCS scores), and a later peak in cases of inflicted TBI (lower mean GCS scores). 2 In adult rodents, NSE peaked at 6 h, which may be earlier than both our current study and in humans, though we were limited by sampling frequency in the current study. 30 Although the pattern of release of NSE was similar in our piglets and what has been reported in children, peak concentrations in the piglets in the current study were lower (2.5–5 ng/mL) than what has been reported in both children and adult rodents (24.3 and 31.5 ng/mL). 2,30 Total estimated NSE was higher in those age groups with larger lesions and correlated with lesion volume, which is similar to work in adult rodents, where greater mechanical force resulted in greater concentrations of NSE after injury (though lesion pathology was not evaluated). 30 In children classified as having mild TBI, concentrations of NSE had little ability to predict outcome, 4 which may be due a reduced release of NSE in clinically mild injuries; however, allometric scaling may increase the predictive value of NSE in growing subjects.
We did not find an increase in the concentration of S100B after injury, which is consistent with previous work in humans demonstrating that S100B has a limited ability to indicate injury or predict outcome in cases of mild to moderate TBI (as determined by GCS score or Injury Severity Score). 4,5 In children with a mean GCS score of 10.8±4.5, S100B was elevated compared to non-injured children (0.026 versus 0.016 ng/mL), and yielded a specificity of 77% when using 0.017 ng/mL as a cut-off value. 2 The specificity of S100B (correctly-identified negatives) is generally low, ranging from 52% 1 to 72%, 2 and S100B was elevated in 90% of ill children without brain injury. 3 In the present study, a large portion of subjects had high S100B concentrations prior to injury. In adult rats, concentrations of S100B after injury did not correspond to the severity of controlled cortical impact. 31 The inability of S100B to indicate brain injury and predict outcome after injury may be also be due in part to the neurotrophic actions of S100B, which remains elevated in brain tissue after peripheral serum S100B concentrations have returned to normal. 11
Because this model of TBI requires anesthesia, it is possible that markers of brain injury may increase as a result of the method used to induce the injury rather than reflecting the injury due to cortical impact. General anesthesia achieved with multiple agents has been shown to induce neuronal apoptosis in immature rodents. 32 Prospective trials examining the potential for anesthetic-induced neurotoxicity in children are underway, but a single anesthetic exposure using a single anesthetic (isoflurane) in immature large animals has resulted in mixed outcomes, and it is not known if anesthetic toxicity is a rodent-specific phenomenon. 33 –35 Serum markers are increasingly being employed to monitor cerebral ischemia during the perioperative period to improve post-surgical neurologic outcomes. 36 It is evident that the blood–brain barrier becomes sufficiently permeable during ischemia to allow escape of proteins purported to be serum markers, 36 but detection of peripheral serum markers during anesthetic exposure in the absence of cerebral ischemia requires further study. In the current study, the increase in NSE was sustained through day 1, perhaps longer than would be expected if exposed to anesthesia alone.
We did not observe an increase in concentrations of MBP in response to cortical impact, indicating that our injury model may have created an average injury load similar to other forms of TBI classified as mild or moderate in other studies. In children with a mean GCS score of 10.8±4.5, MBP concentrations were no different from those of uninjured children. 2 Even after severe TBI in adults, concentrations of MBP did not correlate with GCS scores, but were only different among patients that survived versus patients that died. 19
In this paradigm of scaled cortical impact, concentrations of NSE increased in all age groups regardless of the degree of resulting histopathology; however, when total NSE was estimated taking into account the allometric growth of blood, peak NSE and S100B after injury correlated with lesion volume at day 7. Concentrations of S100B and MBP did not change due to cortical impact, indicating that these markers may not be suitable to detect this type of injury. However, even though this model used a uniform injury mechanism, and NSE consistently increased after cortical injury, the receiver operator curves indicate that the predictive value of NSE alone as a serum marker of TBI is only poor to fair. The use of a single serum marker to predict outcome in cases of mild TBI should be discouraged, especially in a clinical setting, where injuries are varied. Future studies endeavoring to increase the predictive value of serum markers in children should use panels of markers and allometric scaling.
Footnotes
Acknowledgments
This study was supported by an R01 (HD45364-5) from the NIH's National Institute of Child Health and Human Development, the Pediatric Neurosurgery Research Fund from the Children's Hospital at Dartmouth, and the Nicolas T. Zervas Endowed Chair Fund of Massachusetts General Hospital Department of Neurosurgery.
We thank Ms. Rachel Curtis, Ms. Ying Lee, Ms. Leslie Adams, and Mr. Michael Simoni, for their assistance with animal handling and blood collection, and Dr. Susan Margulies for critical review of the manuscript.
Author Disclosure Statement
No competing financial interests exist.