
Abstract
Impairments in learning and memory occur in as many as 50% of patients following traumatic brain injury (TBI). Similar impairments occur in rodent models of TBI, and the development of new memory testing procedures provides an opportunity to examine how TBI affects memory processing in specific neural memory systems. Specifically, metric, topological, and temporal ordering tasks are object-based tests for memory of spatial orientation and temporal sequencing working memory developed for use in rodents. Previous studies demonstrated that specific lesions of the dentate gyrus/CA3 of the hippocampus and the parietal cortex resulted in deficits in the metric and topological spatial orientation tasks, respectively. Lesions of the CA1 impaired a rat's ability to recall the temporal order of odors. The purpose of the following study was to determine whether moderate lateral fluid percussion TBI would generate deficits in these working memory tasks, and whether observed deficits were associated with cell loss in the CA2/3 and/or CA1 of the hippocampus. Two weeks following a moderate lateral fluid percussion TBI, adult rats demonstrated significant deficits in both the metric and temporal ordering tasks (p<0.05) but not in the topological task. Stereological analysis identified a significant reduction in neurons in the CA2/3 (p<0.05) but not the CA1 of the hippocampus. These data demonstrate the utility of three object-based tasks to expand our understanding of how different neural memory systems are affected by TBI.
Introduction
The Morris Water Maze (MWM), a test of spatial memory in which animals utilize spatial cues to find a submerged platform in a water tank, has been the most extensively examined model of spatial maze learning and memory tested in rodent models of TBI in both adult 17 –23 and pediatric animals. 24 –27 Other spatial maze tasks such as Barnes maze 28 –31 and the working memory novel object recognition (NOR) 32 –36 have also been adapted to determine the effects of TBI on cognitive function.
Recently, several object-related working memory tasks have been developed that 1) take advantage of a rodent's natural exploratory behavior, 2) are run in a low-stress environment, 3) have specific hippocampal and cortical anatomical correlates, 4) allow data to be collected in a single day for each task, and 5) can be translated for testing of a clinical population. 37,38 Each task requires an animal to explore either objects or odors on an open table or in an opaque box with four visible extra-maze spatial cues. Specifically, the metric task assesses an animal's ability to recall the relationship of two objects in space based on angles and distances, 39 –41 whereas the topological task assesses an animal's ability to recall relationships among four objects in space based on spatial cues, but without changes of angles and distances separating the objects. 39,40 Finally, the temporal ordering task assesses an animal's ability to discriminate the temporal order in which different odors were presented over time. 42,43 Based on the results of brain lesion studies in rats, processing of spatial and temporal information in each of the tasks appears to be associated with different brain regions. Specifically, bilateral dorsal dentate gyrus and CA3 hippocampal lesions impaired metric memory whereas bilateral parietal cortex lesions impaired topological memory. 37,38 Finally, bilateral lesions of the CA1, especially the more caudal and ventral aspects, significantly impair performance in the temporal ordering task of odors. 42,44
A consistent pattern of neuronal damage following moderate (∼2.2 atm) lateral fluid percussion (LFP) brain injury in the rat has been described using a variety of different staining methods. 21,23,45 –47 Overall, by 7 days post-injury there is significant neuronal loss bilaterally in all subregions of the hippocampus, especially in the CA2/3 and dentate gyrus. Neuronal loss is also observed unilaterally in the ipsilateral parietal cortex after LFP injury. 21 The development of novel cognitive tasks that can be used along with traditional spatial learning tasks are key in better characterizing the relationship between cell death/dysfunction and long-term changes in cognitive performance following a LFP. A better understanding of the nature of these relationships has the potential to improve the design of future pharmacological or behavioral therapeutic strategies.
Methods
Subjects
A total of 65 Sprague–Dawley subjects (Harlan) weighing 300–330 g were used in this study. Animals were housed in individual cages in a temperature (22°C) and humidity- controlled (50% relative) animal facility with a 12 h light/dark cycle. Animals had free access to food and water for the duration of the experiments. Animals remained in the animal facility for at least 7 days prior to surgery. The Institutional Animal Care and Use Committee at the University of California at Davis approved all animal procedures in these experiments.
Surgical procedure
Rats were anesthetized with 4% isoflurane in a 2:1 nitrous oxide/oxygen mixture, intubated, and mechanically ventilated with a rodent volume ventilator (Harvard Apparatus model 683, Holliston, MA). A surgical level of anesthesia was maintained with 2% isoflurane. After rats were mounted in a stereotaxic frame, a scalp incision was made along the midline, and a 4.8 mm diameter craniectomy was performed on the right parietal bone (centered at −4.5 mm bregma and right lateral 3.0 mm). Two skull screws (2.1 mm diameter, 6.0 mm length) were placed into burr holes, 1 mm rostral to bregma and 1 mm caudal to lambda to secure the injury cap. A modified Luer-lock connector 2.6 mm in inner diameter was secured to the craniotomy with cyanoacrylate adhesive and dental acrylic (Yates and Bird, Chicago, IL). Rectal temperature was continuously monitored and maintained (36–37.5°C) throughout the surgical preparation using a feedback temperature controller pad (CWE model TC-1000, Ardmore, PA). Temporalis muscle temperature was measured by insertion of a needle temperature probe (Physitemp unit TH-5, probe MT-29/2, Clifton, NJ) between the skull and temporalis muscle.
LFP TBI
Experimental TBI was produced using a fluid percussion device (VCU Biomedical Engineering, Richmond, VA) 48 with the lateral orientation for impact. 49 The LFP device consists of a cylindrical Plexiglas reservoir filled with isotonic saline. One end of the reservoir has a Plexiglas piston mounted on O-rings and the opposite end has a transducer housing with a 2.6 mm inside diameter male leur lock opening. An extracranial transducer (model SPTmV0100PG5W02; Sensym ICT) connected to a digital storage oscilloscope (model TDS 1002; Tektronix Inc., Beaverton, OR) is used to determine the atmospheric pressure (ATM) generated during each injury. Moderate TBI (2.14±0.003 ATM) was produced by rapidly injecting a small volume of saline into the closed cranial cavity (over the right ipsilateral hemisphere).
Immediately following LFP, rats were returned to ventilation with a 2:1 nitrous oxide/oxygen mixture in the absence of isoflurane, and returned to the heating pad to maintain normothermic body temperature. The plastic injury tube and skull screws were removed and the scalp incision was closed with 4-0 braided silk sutures. When spontaneous breathing was observed, animals were extubated, and righting reflex was assessed at regular intervals (∼20 sec) to test the rat's ability to spontaneously recover to a prone position. The duration of suppression of the righting reflex was used as an additional indicator of injury severity. Animals were then returned to their cages and placed on a heating pad until they were ambulatory. They were then returned to the vivarium.
Behavioral analysis
Separate animals were used for each of the three behavioral tasks to allow for testing exactly on post-injury day 14. For each of the tasks, behavior was video-recorded by a camera mounted above the apparatus (Logitech Webcam Pro 9000, Logitech® International S.A., Romanel-sur-Morges Switzerland) and stored using computerized software (Dell Vostro 400, Dell Inc., Round Rock, TX). Behaviors were visually scored off-line in a blinded fashion by a single, highly trained investigator, using a stopwatch that measured tenths of a second (Thermo-Fisher Scientific, Waltham, MA). For each behavior animals were habituated to the environment prior to the task. Data for each task was collected on post-injury day 14.
Metric task
The metric spatial information task, described by Goodrich-Hunsaker and colleagues, 40 investigated rats' ability to recognize differences in the distance between two distinct objects based on angles and spatial cues. A large white acrylic circular platform (2 m in diameter and 0.6 cm thick) was placed on top of a table 1.09 m above the floor. A white curtain surrounded the platform to reduce the number of extra-maze cues. Four distinct visual cues were hung equally spaced around the curtain. Animals were habituated to this environment for 10 min on post-injury day 13, the day prior to behavioral testing. Behavior in the metric task was assessed on post-injury day 14.
The metric spatial task consisted of two continuous periods, habituation (15 min) and test (5 min). Two novel objects were positioned in the center of the platform 68 cm from each other (Fig. 1A). During the habituation period, the rat was placed on the edge of the platform equidistant from the two objects, and allowed to freely explore the environment including the two objects for 15 min. After the habituation phase animals were returned to their home cage for 5 min while the platform and objects were cleaned with 70% ethanol and the distance between the two objects was reduced to 34 cm (Fig. 1A). The animals were then returned to the platform for a 5 min test period. In both the habituation and the test, animals were scored for the time (in seconds) spent actively exploring objects, with active exploration defined as object sniffing, physical contact, and rearing.
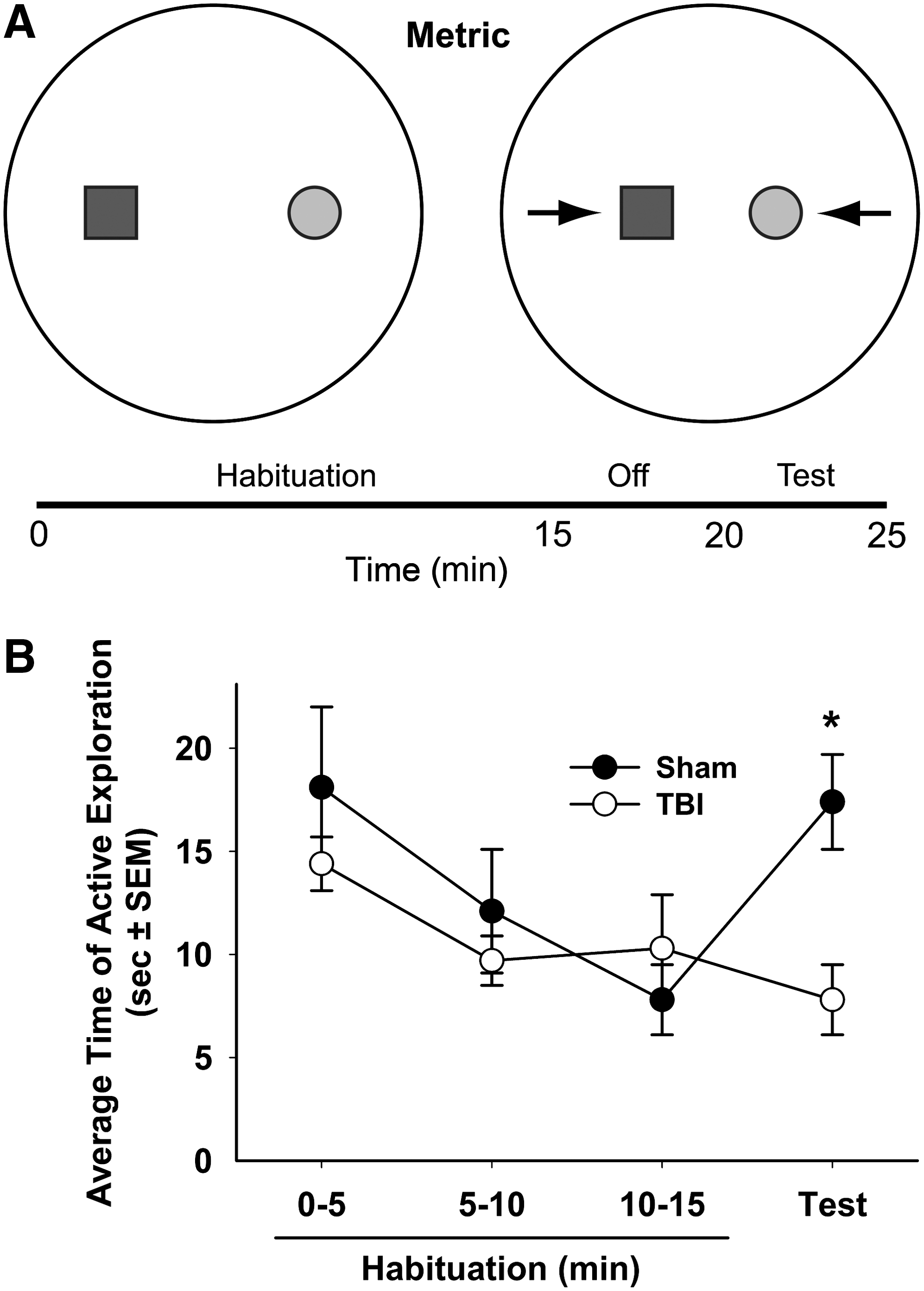
The metric task is a working memory task in which, after a 15 min habituation period, an animal is tested for the ability to discriminate a change in distance between two objects
Topological task
The topological spatial information task, modified from the task described by Goodrich-Hunsaker and colleagues, 40 examined rats' ability to recognize differences in the placement of objects relative to each other using only spatial cues. The platform, room and experimental timeline used for this task were the same as those used for the metric task. Similar to the metric task, animals were habituated to this environment for 10 min on the day prior to analysis of behavior. Performance in the topological task was assessed 14 days post-injury. Four novel objects were positioned around the center of the platform 68 cm from each other (Fig. 2A) and at the beginning of each trial rats were placed on the edge of the platform equidistant from two of the objects. During the 15 min habituation period, animals were allowed to freely explore the environment including the four objects. After this habituation phase, animals were returned to their home cage for 5 min. During these 5 min, the platform and objects were cleaned with 70% ethanol and the positions of the front two objects were transposed (Fig. 2A). The animals were then returned to the platform for a 5 min test. Animals were scored (in seconds) for the time spent actively investigating the front two objects in both the habituation and test.

The topological task is a working memory task in which, after a 15 min habituation period, an animal is tested for the ability to discriminate a change in the spatial relationship between two objects
Temporal ordering task
The temporal memory ordering task 42 investigated rats' ability to remember a sequence of odor presentations. The temporal ordering task was performed in a test chamber (white acrylic box with an open top [0.66 m2 surface area with 0.5 m high walls]). The test chamber was placed on top of the table that was utilized for the metric and topological tasks. Animals were habituated in the test chamber for 2 days prior to the test, spending 5 min each day in the box in the absence of any objects. Performance in the temporal ordering task was assessed 14 days post-injury. The temporal ordering task consisted of three 5 min acquisition periods and a 5 min test each separated by a 15 min interval in which the animals were placed in their home cages (Fig. 3A). Three odors were utilized to assess an animal's ability to recall a sequence of temporal presentations. The odors used in this task were based on those previously described. 42,50 Briefly, the common spices of mace, ginger, and cloves (McCormick & Company, Inc., Sparks, MD) were mixed with unscented sand and poured into specimen cups (40 mL clear polypropylene, Thermo-Fisher Scientific, Waltham, MA). During each of the three 5 min habituation periods, animals were presented with two cups, filled with the same odor, each 160 cm away from opposing corners of the test box (Fig. 3A). The order of the odor presented was always mace followed by ginger, and then cloves, with mace being defined as the “initial odor” and cloves the “recent odor.” After the 15 min delay period following the presentation of the cloves odor, animals were tested for recall of temporal ordering of odor presentation. Specifically, animals were given 5 min with two specimen cups. One contained the initial odor and the second the most recent odor. Animals were scored (in seconds) for the time spent actively investigating each odor during acquisition and the test, with active investigation defined as sniffing, and physical contact with the specimen cup. To determine whether animals preferred the initial odor during the test phase, time actively interacting with the initial odor was compared with total odor exploration time (excluding non-interaction time): (time with initial odor/time with initial+recent odor) *100%.
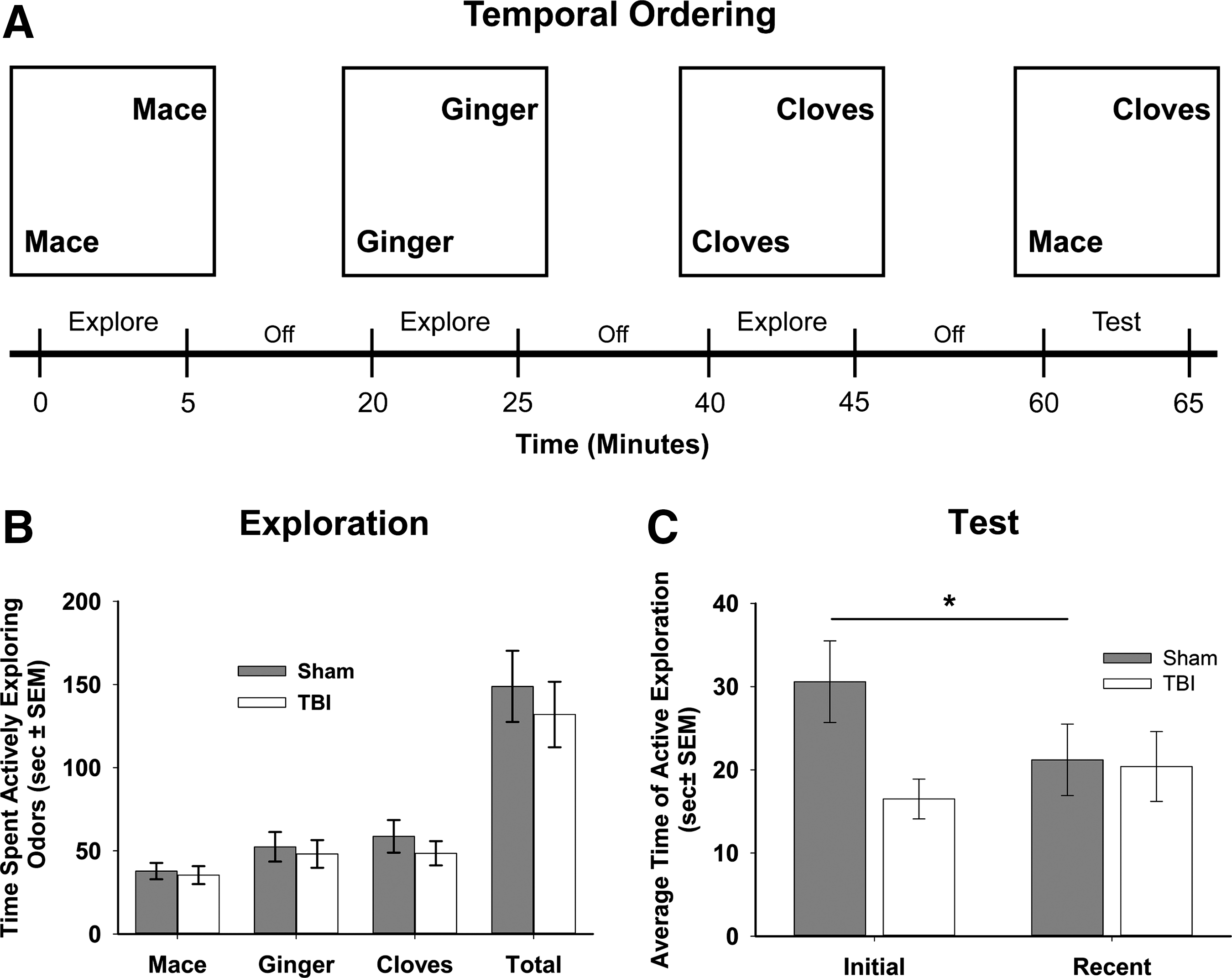
The temporal ordering task is a working memory task in which animals are tested for their ability to discriminate two odors that are presented over a 1 h period
Tissue collection and sectioning
Rats were euthanized on post-injury day 16 by deep sodium pentobarbital anesthesia (100 mg/kg, ip), transcardially perfused with 100 mL of 0.1 M sodium phosphate buffer (pH=7.4), followed by 350 mL of 4% paraformaldehyde (pH 7.4). Brains were removed and post-fixed for 24 h in 4% paraformaldehyde at 4°C. Brains were immersed in 10% sucrose solution for 24 h followed by immersion for 48 h in a 30% sucrose solution. After buffer exchange, brains were frozen on powdered dry ice, and 45 μm coronal sections were cut on a sliding microtome (American Optical, Model 860). Every serial section starting at −2.12 mm bregma and ending at −4.80 mm bregma was saved in 24-well cell culture plates, and every fifth section throughout the hippocampus was then mounted on gelatin-coated glass microscope slides.
Cresyl violet staining
Slide-mounted brain sections were dehydrated at room temperature by immersion in a succession of ethanol baths: 70% (2 min×1), 95% (2 min×2), and 100% (2 min×2), followed by xylene treatment for 16 min. Sections were then rehydrated in 100% (2 min×2), 95% (2 min×2), and 75% (2 min×1) ethanol baths and rinsed with distilled water (30 sec×2). Afterwards, sections were stained with cresyl violet acetate (0.1%) for 6 min, rinsed in distilled water (15 sec×2), differentiated in 95% ethanol with 0.15% acetic acid, and dehydrated in 95% (30 sec×2) and 100% (30 sec×2) ethanol and xylene (5 min×2). After dehydration, the sections were cover-slipped with Permount (Thermo-Fisher Scientific, Waltham, MA).
Stereological cell counts
Serial cresyl violet stained tissue sections from a random subset of brains were selected across five surgical dates for stereological analysis of the numbers of hippocampal CA2/3 and CA1 pyramidal neurons (sham n=12, TBI n=13) ipsilateral to the fluid percussion injury. Estimation of the total number of neurons in the ipsilateral hippocampus was made on a brightfield microscope (Nikon E600, Nikon, Tokyo) with a motorized stage (Bioprecision2, Ludl Electronic Products, Inc., Hawthorne, NY) using stereological software (Stereo Investigator™ 8.0, Microbrightfield, Inc., Williston, VT). For the CA2/3 counts, the region of interest was defined as the stratum pyramidale of the hippocampal CA2/3 with the borders of the region defined as the pyramidal layer entry into the dentate gyrus at the lateral tips of the dorsal and ventral blades of the dentate granule cells and the narrowing of the stratum pyramidale at the intersection of the CA1 to CA2. For the CA1 counts, the borders were defined as the intersection of the stratum pyramidale of the CA1 to CA2 through the most medial aspects of the CA1. The regions of interest were outlined using a 10×objective (Plan Apo, NA 0.45, Nikon). Neuronal cell counting was performed with a 100×oil immersion objective (Plan Apo, NA 0.95, Nikon). The criterion for selection and quantification of surviving neurons was a morphologically distinct neuronal cell body.
Statistical analysis
Data analysis was performed using SPSS software (Version 18, Chicago, IL), which adheres to a general linear model. Alpha level for type I error was set at 0.05 for rejecting null hypotheses. All data are expressed as mean±standard error of the mean (SEM). Data for body weights and righting times were analyzed using independent samples t tests (two tailed). Performance during the initial 15 min habituation period for the metric and topological tasks was analyzed using a repeated measures ANOVA, with object exploration over the three 5 min blocks of exploration used as the repeated within-groups variable. Object exploration during the last 5 min of the habituation period and during the 5 min test period was analyzed using paired-samples t tests (two tailed) to determine whether animals re-explored following alterations in the spatial environment. Time spent actively exploring the scents in the temporal ordering task was analyzed using an independent samples t test (two tailed). The scent preference among the three odors was analyzed using a one-way ANOVA with a Bonferroni post-hoc analysis. The stereological estimate of the number of neurons between groups was analyzed with an independent samples t test (one tail). Finally, a two tailed Pearson's correlation was used to determine if there was a linear relationship between CA2/3 and/or CA1 neuron number with performance in the metric and temporal tasks.
Results
There was a significant difference in righting time between sham (2.5±0.1 min) and TBI (13.5±1.1 min) animals (p<0.001). At the time of behavioral analysis, there was a significant difference in weight between sham and injured animals (352.6±5.4 and 325.2±5.2 g, respectively; p<0.001). However, between post-injury days 7 and 14, sham animals gained weight at an average of 3.12±0.45 g/day whereas TBI animals gained weight at an average of 4.55±0.58 g/day (p=0.06) indicating that injured animals were ambulatory and consuming food in a similar manner to sham-injured animals. Each animal was randomly selected for one of each of the three behaviors tested.
Metric task
Analysis of exploration time across the habituation period did not detect an overall difference in exploration between sham and injured animals (p=0.6; Fig. 1B), indicating that animals explored and habituated to the objects in a similar manner regardless of injury. Over the 15 min habituation, there was a significant decrease in time the animals spent actively exploring the objects (F (2,36)=5.80, p<0.01). Furthermore, the interaction between time and group was not significant (p=0.30) indicating that sham and TBI rats habituated in a similar fashion. After a 5 min period spent in their home cages, the animals were returned to the environment with the same two objects, but with the distance between objects reduced to 34 cm. During the test, sham animals had a significantly increased exploration of the objects (9.6 sec or 122% increase) in the new configuration (t=−2.97, df=8, p<0.05) as compared with the final 5 min period of the habituation. In contrast, TBI animals had no significant change in exploration (2.2 sec or 23% decrease) as compared with the final 5 min period of habituation (t=0.99, df=10, p=0.35; Fig. 1B).
Topological task
There was no difference in the exploration of the front two objects between the two groups during the initial 15 min habituation period (p=0.78; Fig. 2B). Over the 15 min habituation, there was a significant decrease in time the animals spent actively exploring the objects (F (2,42)=19.56, p<0.001). Furthermore, the interaction between time and group was not significant (p=0.36) indicating that sham and TBI rats habituated in a similar fashion. After the position of the front two objects was transposed, both sham (t=−2.99, df=10, p<0.05) and injured (t=−2.87, df=11, p<0.05) animals had a significant increase in exploration of the objects in the novel configuration (Fig. 2B).
Temporal ordering task
One TBI animal was removed from the analysis for failure to explore the odors during the acquisition period. There was no difference in exploration time (Fig. 3B) of each odor between sham and TBI rats for mace (p=0.74), ginger (p=0.73), or cloves (p=0.42). There was also no difference in total exploration time across all three scents between sham and TBI injured animals (Fig. 3B; p=0.57). Additionally, there was no preference for any particular odor (F (2,66)=2.86, p=0.064), although there was a trend toward a preference of the final odor, cloves, as compared with the initial odor, mace (p=0.08). In the test phase, both sham and TBI animals spent a similar amount of time exploring the most recent odor (21.2±4.3 and 20.4±4.2 sec respectively; p=0.9; Fig. 3C). There was a significant increase in the amount of time sham animals spent re-exploring the initial odor, 30.6±4.9 sec, as compared with the most recent odor (a 9.4 sec or 44% increase; t=2.50, df=21, p<0.05; Figure 3C). In contrast, TBI rats showed no preference, as they spent a similar amount of time exploring the initial odor, 16.5±2.4 sec, as they did exploring the most recent odor (a 3.9 sec or 19% decrease; p=0.135). Additionally, there was a significant difference in the percent of total exploration time during the test spent with the initial odor, mace, between sham and TBI animals (60.6±4.3 and 47.1±3.5% respectively; t=2.41, df=21, p<0.05). Overall, these data suggest that TBI animals have a deficit in working memory related to the temporal ordering of three odors.
Hippocampal cell counts
Of the 25 brains randomly selected for stereological analysis, three were not included because of histological artifacts. The final sample size for sham was n=9 (n=3 each from the metric, topological, and temporal ordering groups) and n=13 for TBI (n=5, n=3 and n=5 from the metric, topological, and temporal ordering groups). Estimates of the number of pyramidal neurons in the stratum pyramidale of the hippocampal CA2/3 were made using unbiased, randomized, stereological techniques. There was a significant reduction in the estimated number of neurons by weighted section thickness in the CA2/3 of TBI (38,547.3±2060.8) as compared with sham (44,466.5±1731.5) injured brains (t=0.861, df=20, p<0.05; Fig. 4A). There was no significant difference in the number of CA1 neurons between sham (60,212.0±2741.7) and TBI (56,498.5±3036.8) animals (p=0.16; Fig. 4B). Coefficients of error ranged between 0.05 and 0.11 (mean values: sham CA3=0.079±0.002, TBI CA3=0.085±0.004, sham CA1=0.057±0.002 and TBI CA1=0.064±0.004). There was not a linear relationship between CA2/3 or CA1 neuronal number and performance in the metric (r[6]=0.34, p=0.41 and r[6]=0.54, p=0.17, respectively) or temporal ordering (r[6]=0.02, p=0.96 and r[6]=0.16, p=0.71, respectively) tasks.
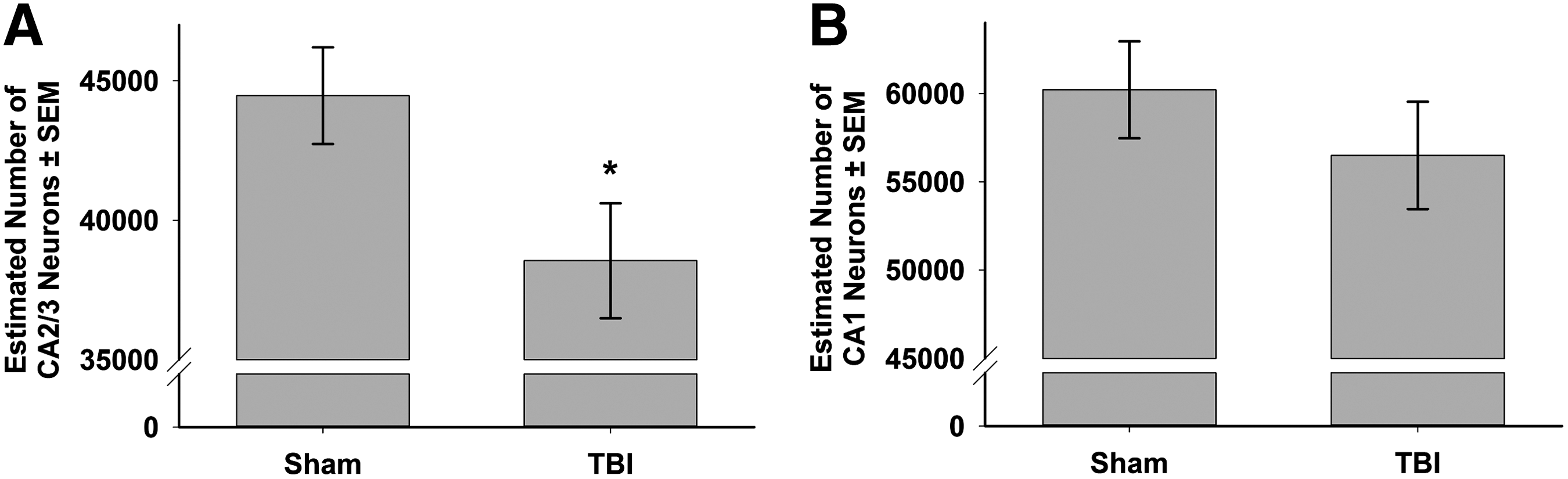
Stereological estimation of the number of pyramidal neurons in the CA2/3
Discussion
Adult animals received a sham or moderate LFP and were tested for their performance in three working memory-based cognitive tasks: the metric, topological, and temporal ordering tasks. In the metric task, animals were tested on the ability to discriminate the distance between two objects in space. In the topological task, the animals were tested on their ability to discriminate when two objects had their locations transposed in space. Finally, in the temporal ordering task, animals were tested on their ability to discriminate the temporal order of the presentation of three odors. In each case, sham animals were able to discriminate changes in object location and temporal order as previously described. 37,40,42 In contrast, following moderate LFP, injured animals were unable to discriminate a change in distance between two objects in the metric task, as they continued to show habituation to the two objects after they had moved closer together. However, injured animals were able to distinguish when two objects were transposed in space in the topological task and performed identically to shams. Finally, injured animals were unable to distinguish between the temporal ordering of odors.
The metric, topological, and temporal ordering working memory tasks were designed as low stress cognitive tasks that utilized a rodent's natural exploratory behaviors. Furthermore, unlike many of the spatial maze tasks, these tasks require minimal training (habituation to the environment for 1-2 days), and data can be collected in a single day. Additionally, these tasks can be useful in functionally evaluating the roles of cell death and neuronal connectivity following TBI, as these tasks have specific neuroanatomical correlates in rodents. 37,38 Specifically, in the metric task, control animals re-explore two objects after the distance between them is reduced. Lesions of the dorsal hippocampus, and specifically those that damage the dentate gyrus, hilus, and CA3 of the hippocampus significantly disrupt object re-exploration during the test phase on the metric task. 38,40 Lesions of the parietal cortex did not significantly alter performance in the metric task. 38 In a separate task that requires use of metric cues (e.g., distance and angles between objects), animals with dorsal hippocampal lesions failed to distinguish between an object that was placed in a “correct” location and one that was placed in an “incorrect” location 50 cm away. 51,52 These findings led the authors to conclude that the dentate gyrus and CA3 of the hippocampus are critical to the formation of object location memory based on relationships of distance and angle. 53 In the present study, LFP resulted in a significant decrease in the number of CA2/3 neurons in the dorsal hippocampus ipsilateral to injury. Consistent with previous lesion studies of the CA3 in the dorsal hippocampus, LFP- injured animals with a loss of CA2/3 neurons failed to re-explore two objects when there was a change in the distance between the objects.
In the topological task, animals are exposed to four objects over 15 min. After a 5 min interval, two of the objects are transposed in space. Unlike the metric task, which relies on changes in the distance between two objects and therefore a change in the visual angle between them, 39,54 topological information is based on the spatial relationships among the four objects and the four external cues. 39,54,55 Whereas lesions of the CA2/3 dorsal hippocampus led to deficits in working memory in the metric task, these lesions did not disrupt performance in the topological task, 40 as animals habituated and then re-explored the transposed objects in a similar fashion to uninjured control animals. Unlike the metric test, lesions of the parietal cortex disrupt performance in the topological task such that animals with bilateral lesions of the parietal cortex fail to re-explore the transposed objects. 37,40 Previous studies have demonstrated that LFP injury generates significant cell death in the parietal cortex, 21,56,57 and, therefore, we hypothesized that we would see a significant impairment in performance in the topological task. However, in the current study, the patterns of habituation and re-exploration between sham and TBI animals were indistinguishable. This suggests that the severity of LFP investigated in these studies does not cause a lesion sufficient to impair performance in the topological task. It is possible that surviving neurons and circuitry in the ipsilateral cortex were sufficient to support completion of the task or that neurons and circuitry from the contralateral parietal cortex compensated for the injured ipsilateral cortex, allowing LFP-injured rats to process topological associations.
Previous studies have shown that naive rats presented with three different odors prefer (i.e., show increased exploration of) the first presented odor, as compared with the most recently presented odor when given a choice, demonstrating the ability to process the temporal order of odor presentation. In contrast, rats with excitotoxic lesions in CA1 prefer the last odor presented, suggesting a role for hippocampal CA1 in processing of temporal order. 42,58 In the present study, rats with LFP injury showed no preference for the initial versus the most recently presented odor. This is unlikely to be because of impairment in odor discrimination because both groups show clear and similar preferences for specific odors during the acquisition phase (i.e., clove>ginger>mace). The difference between the findings may be the result of the more diffuse pattern of neuronal injury produced by LFP, which includes damage to the hilus, CA2/3, and parietal cortex in addition to CA1. 21 –23,45,47 Following LFP injury, there is also evidence of limited cell death in the thalamus, CA1, and entorhinal cortex 21 as well as changes in functional connectivity within the hippocampus including depressed long-term potentiation 59 –62 and theta oscillatory rhythms. 29 Therefore, changes in neuronal number and function throughout the hippocampus may contribute to the impaired processing of temporal order.
In the CA2/3, there was a significant reduction in the number of neurons, whereas in the CA1, there was not (p=0.2). The loss of CA2/3 neurons was associated with a diminished performance in both the metric and temporal ordering tasks. Within the constraints that the histology was analyzed on a randomly chosen subset of animals, we performed a Pearson's correlation between cell counts in the CA2/3 and the CA1 with metric and temporal ordering performance, respectively, but no significant relationship was observed. One explanation is that TBI causes not only cell death but also cell dysfunction following injury, and, therefore, analysis of cell death alone may not be predictive. However, we feel that the lack of the correlation lies more in the nature of the two tasks. In the metric task, animals that remember the original position of the objects re-explore during the test phase, whereas if they forget, they continue to habituate. In neither the lesion studies 40 nor in the current study, did injury alter habituation; therefore, cell loss in the CA2/3 does not affect habituation. TBI animals fail to re-explore and continue habituating in contrast to the shams that re-explore. As TBI animals continued to habituate during the test phase, it is not surprising that no relationship was detected between cell number and performance. As we have already described, CA2/3 cell loss does not have any effect on how animals habituate. In the temporal ordering task animals were allowed to explore both the initial and most recent odor. Whereas sham animals preferentially re-explored the initial odor, injured animals showed no preference. If injured animals do not remember the temporal order of the odors, then there is an equal probability that they will spend equivalent time with the initial odor and the most recent odor, in other words, exploration will be random. Therefore, it is not surprising that random performance in the temporal ordering task does not correlate with CA1 cell number.
One concern is that deficits in the metric and temporal tasks could be related to a general working memory deficit, rather than being specific for each task. In the metric and topological tasks, both injured and sham animals habituated to the objects, and, therefore, there was no deficit in working memory associated with habituation. Additionally, in the initial characterization of the topological task, 40 animals were habituated in three separate 5 min bins, each separated with a 5 min interval. Animals continued to habituate to the objects until the orientation was modified. These data suggest that simply removing and replacing an animal in an environment is insufficient to induce re-exploration. In the temporal ordering paradigm, animals were exposed to new odors every 15 min. Animals explored the second and third odors in a similar fashion to the initial odor, and, therefore, there was not a working memory deficit for detecting a novel odor. Finally, injured animals performed identically to shams in the topological task. If TBI induced a general deficit in working memory, one would predict that injured animals would also fail to recognize the transposed objects in the topological task.
One of the goals when designing cognitive tasks for rodents is to generate paradigms that are representative of learning and memory that could be assessed in humans. Kessels and colleagues distinguished several different object-based paradigms that can be assessed in patients, including route learning (i.e., using visual cues to find a target), object-location memory (i.e., where objects exist in an environment), and spatial working memory. 63,64 There is a large body of data exploring the effect of TBI on route learning, using primarily the MWM, 17 –23 as well as the Barnes Maze. 29,30 Additionally, there are different types of spatial working tasks that have been utilized in models of TBI including the T-maze 65 –69 and the radial arm maze. 18,21,70 There are also studies that have evaluated object recognition using the novel object task. 32,33,36 Object location memory and spatial working memory assessed in the metric and topological tasks, as well as temporal sequencing, have not been well addressed in experimental models of TBI.
In a meta-analysis of multiple clinical experiments, Kessels and colleagues concluded that patients with hippocampal lesions had deficits in maze learning, working memory, object location memory, and positional memory as compared with controls. 63 Importantly, they found that lateralization of the lesion (i.e., whether the lesion was in the left or right hippocampus) was significantly related to whether the patient had deficits in maze learning, object-location memory, or positional memory. Working memory was not lateralized. A group of 25 amygdalohippocampectomy patients were evaluated for performance in object location memory. Using a touch screen computer, patients were asked to remember the orientation of objects with or without having to recall changes in distance and angle. Patients with amygalohippocampal lesions were able to perform the topological but not the metric task. 64 A population of patients with right or left parietal cortex strokes was tested for their ability to discriminate changes in metric and topological space using a matching paradigm on a computer. Stroke patients made significantly more errors in both the metric and topological tasks regardless of the location of the lesion, as compared with controls. 71 In a group of hypoxic patients with hippocampal damage, memory for a procedural sequence was assessed. Using a computer, patients were instructed to recall the correct sequence for order of entry into 16 different arms on a maze. Patients with hippocampal damage performed significantly worse on the task than did controls. 72 Finally, a group of patients with temporal lobe epilepsy were asked to recall autobiographical stories. Whereas those patients could remember the details of their stories, they could not consistently remember the temporal sequence of events that happened. Problems with temporal sequencing were present regardless of whether the events were clustered in a very short period of time or over a long period of time. 73 These clinical studies demonstrate how tests of metric and topological spatial working memory, as well as temporal sequencing, can be translated to test cognitive function in patients with neurological disorders.
Conclusion
In conclusion, these data demonstrate the utility of exploring new assessments of cognitive function both in rodents and in the clinical setting following TBI. The tasks adapted in this article are low stress in nature, and take advantage of the rodent's natural behaviors. They are tasks that can each be completed in a single day, and they have specific anatomical representations in the rat. Our data indicate that damage to the CA3, perhaps related to a significant loss of CA2/3 neurons, leads to failure to recall the relationship of two objects in space using information related to distance and angles in the metric task. Despite cell death in the parietal cortex, injured rats performed identically to sham animals in a task using topological information related to the connectivity and containment of four objects in space and spatial cues. Finally, there was a disruption in injured animals' ability to recall the temporal sequence of three odors presented to them. Whereas our present data failed to detect an overt loss of cells in the CA1, these data suggest that the connectivity related to the CA1 has been disrupted following injury. Better understanding of the different types of cognitive impairments following injury, as well as information related to the anatomical loci of these behaviors, may be critical in our understanding of deficits in learning and memory in TBI patients. Furthermore, by having a range of tasks to assess the specific nature of an individual's chronic cognitive disability, we may be able to design more effective patient-specific therapeutic approaches.
Footnotes
Acknowledgments
This work was supported by the NSF Center for Biophotonics Science and Technology, a designated NSF Science and Technology Center managed by the University of California at Davis under the cooperative agreement number PHY0120999. We thank Eric West, Vanessa Atkinson, and M. Hiram Dominguez for their expert technical assistance on this project.
Author Disclosure Statement
No competing financial interests exist.