
Abstract
Although experimental traumatic brain injury (TBI) studies support estradiol as a neuroprotectant and potent stimulator of neuroplasticity, clinical studies suggest a negative association between endogenous estradiol profiles and mortality/poor outcomes. However, no studies have evaluated associations with cerebral spinal fluid (CSF) hormone profiles and aromatase gene (cytochrome P450 [CYP]19A1) variability on clinical TBI outcomes. We evaluated 110 adults with severe TBI. Average and daily estradiol, testosterone, and estradiol/testosterone ratios (E2:T) were measured using CSF and serum samples and compared to healthy controls. Eighteen tagging and four functional single-nucleotide polymorphisms (SNPs) for CYP19A1 were genotyped and compared to hormones, acute mortality, and Glasgow Outcome Scale (GOS) scores 6 months post-TBI. TBI subjects had lower CSF estradiol over time versus controls. CSF testosterone was initially high, but declined over time. E2/T ratios were initially low, compared to controls, but rose over time. Higher mean E2/T ratio in bivariate analysis was associated with lower mortality (p=0.019) and better GOS-6 scores (p=0.030). rs2470152 influenced CSF E2/T ratio and also serum and CSF testosterone (p≤0.05 all comparisons). Multiple-risk SNPs rs2470152, rs4646, and rs2470144 were associated with worse GOS-6 scores (p≤0.05, all comparisons), and those with>1 risk SNP variant had a higher risk for poor outcome, compared with those with ≤1 risk variant. TBI results in low CSF estradiol and dynamic CSF testosterone and E2/T ratio. In contrast to clinical serum hormone studies, higher CSF E2/T ratio was associated with better outcome. Further, genetic variation in CYP19A1 influences both hormone dynamics and outcome post-TBI.
Introduction
Estrogens are produced by conversion of androgens by aromatase, a cytochrome P450 (CYP) enzyme. 1 Aromatase is located on chromosome 15q21.1. It has a 30-kilobase (kb) coding region, with nine exons (II–X) and at least a 93-kb regulatory region consisting of variable tissue-specific promoters. 5 Promoter I.f is expressed in brain tissue. 6,6 Several brain regions make aromatase, 4 and the enzyme is located primarily in neuronal cell bodies and in presynaptic terminals. 2 Aromatase inhibition can reduce axon growth in developing neurons and reduce the number of hippocampal spine synapses. 8 However, the role of aromatase in central nervous system (CNS) hormone physiology and neuroprotection after traumatic brain injury (TBI) is not known.
After CNS damage, estradiol acts on oligodendrocytes, T cells, microglia, and astroglia to promote remyelination and proliferation of progenitor cells by growth factor expression to decrease acute edema and inflammation. 2 In a TBI model, daily 17β-estradiol in ovariectomized rats and males showed better cerebral blood flow and recovery. Other neuroprotective mechanisms for estradiol have been proposed, including increased prostacyclin synthase, increased prosurvival protein B-cell lymphoma 2 expression, reduced β-amyloid, and lowered glutamate-induced excitotoxicity. 9 Although hormone use in animal models has demonstrated the neuroprotective qualities of estradiol, systemic estradiol levels early after severe injury in humans are associated with mortality and poor outcomes. 10
Androgens can also influence neuronal growth and plasticity in the CNS. Removal of testes in rats and monkeys reduces the density of synaptic contacts on CA1 neurons, an effect reversed by administering testosterone. 11 Increased spine synapse density in hippocampal CA1 neurons occurred in ovariectomized female rats after treatment with testosterone or androgen dihydrotestosterone, the latter being a nonaromatized form of androgen. 11 Thus, testosterone may influence neuroplasticity through both increases in estradiol production and through aromatase-independent pathways. Testosterone pretreatment in models of TBI, however, can increase histological damage. 12
Our clinical research supports the dual and varied temporal effects of estradiol and testosterone on CNS neuroprotection and neuroplasticity, discussed in these studies cited above, by showing that high acute serum testosterone and estradiol levels are linked with worse outcomes, 10 whereas higher chronic endogenous serum testosterone and estrogen levels over the first year postinjury are associated with better outcomes in a small population of men with severe TBI (sTBI). 13 However, to date, no studies have established cerebrospinal fluid (CSF) estradiol and testosterone profiles after TBI, their relationships to serum hormone profiles, and if or how these profiles might relate to secondary TBI and outcome. Further, variability in the aromatase gene has not been evaluated for its role in mediating hormone profiles postinjury or with influencing outcome. We hypothesized that whereas elevated estradiol and testosterone in the periphery are associated with the acute stress response of sTBI, hormone production in CSF may represent CNS hormone production, with higher estradiol levels possibly relevant for TBI neuroprotection and favorable outcomes. Therefore, the aim of this exploratory study was to determine (1) CSF profiles for estradiol and testosterone (and their ratios) acutely after injury, (2) CSF/serum relationships between hormones, and (3) whether variation in the aromatase gene influences CNS estradiol and testosterone levels, E2/T ratios, and global outcome in an adult population with sTBI.
Methods
Study design and subjects
Subjects were enrolled in a study approved by our university's institutional review board. We evaluated 110 adults with sTBI admitted to a level 1 trauma center. Because of known racial differences in allelic distribution frequencies, subjects were included in this analysis if they were Caucasian. 14 Other inclusion criteria were age between 18 and 75, sTBI based on a Glasgow Coma Scale (GCS) ≤8, acute pathology confirmed by a cranial computed tomography (CT) scan, requirement of an extraventricular drainage catheter (EVD) for intracranial pressure (ICP) monitoring and management, a signed consent from next of kin for CSF collection and DNA sampling, and at least two CSF samples in the first 144 h post-TBI for analysis. Patients with cardiac or respiratory arrest before admission, documented prolonged hypoxia or hypotension before admission, evidence of brain death within the first 3 days after injury, an Abbreviated Injury Score of 5 in any region other than the brain, penetrating TBI, previous history of endocrine tumor, breast cancer requiring chemotherapy or tamoxifen treatment, prostate cancer requiring orchiectomy or luteinizing-hormone–suppressing agents, or untreated thyroid disease were excluded.
A small (N=13) healthy control group was used to compare CSF hormone levels. Control subjects did not have previous endocrine pathology and were not taking oral contraceptives, hormone replacement therapy, or hormone-modifying supplements. There were 6 healthy men and 7 healthy women enrolled (22–62 years of age), including 1 menopausal woman. For premenopausal women, 2 had samples collected during the follicular phase (days 5–10), and 4 had samples collected in the luteal phase (days 18–23). Potential control subjects who were pregnant, had hormone or contraception therapy, or history of an endocrine disorder were excluded. Control subjects had no history of TBI, neurological or bleeding disorder.
Critical care management
Enrolled TBI subjects received treatment consistent with The Guidelines for the Management of Severe Head Injury. 15 Monitoring included temperature, initial placement of an EVD, central venous catheter, arterial catheter, and pulse oximeter. ICP was kept at <20 mmHg, cerebral perfusion pressure was maintained at >60 mmHg, and mean arterial pressure (MAP) was maintained at >90 mmHg. If cerebral perfusion pressure remained low, pressors or inotropes to were used to maintain MAP levels. Sixteen of one hundred and ten subjects were enrolled in a randomized control clinical trial evaluating moderate hypothermia after sTBI, with 11 receiving a hypothermia protocol (temperature, 32.5–33.5°C for 48 h). The 5 randomized subjects not receiving hypothermia were treated to maintain a normothermic state as the standard of care and similar to the rest of the cohort. Twelve subjects were also involved in the Citicoline Brain Injury Treatment (COBRIT) study, a randomized, double-blind, placebo-controlled, multi-center trial studying 90 days of citicoline treatment on functional outcome after TBI. 16 Five subjects were randomly assigned to receive citicoline (1000 mg twice-daily), and 7 subjects were assigned to placebo for 90 days beginning 24 h after injury.
CSF sample collection and measurements
CSF samples (n=634) were collected passively by EVD, and samples were collected up to two times daily, for up to 6 days postinjury for those with TBI. CSF was obtained from healthy controls by a single lumbar puncture. Estradiol and testosterone were measured using high-sensitivity enzyme immunoassay kits (Salimetrics, LLC, State College, PA). Daily samples were averaged and assessed for each 24-h period postinjury. Weekly averages for each patient were calculated from daily averages for days 0–6 or the first 144 h post-TBI.
Serum sample collection and measurements
Blood samples (n=429) were collected at ∼7:00
DNA extraction
DNA for TBI subjects was extracted from whole-blood samples before transfusion or from CSF. Blood was collected into ethylenediaminetetraacetic acic vacutainer tubes, centrifuged, processed to retrieve the buffy coat, and DNA was extracted using a salting-out procedure. 18 If no blood sample for DNA extraction was available for a subject, DNA was extracted from CSF samples using a kit with a QIAamp DNA extraction protocol from Qiagen Corporation (catalog no. 51104; Qiagen, Hilden, Germany).
Genotyping
To assess the variability of the entire aromatase gene, CYP19A1, including the promoter regions located in intron 1, four functional single-nucleotide polymorphisms (SNPs) and 18 tagging SNPs (tSNPs) were analyzed: rs700519; rs28757184; rs2236722; rs56658716; rs2470144; rs2470152; rs4646; rs6493496; rs7168331; rs8041933; rs2470151; rs3751592; rs12901187; rs1062033; rs7172156; rs11632036; rs12591359; rs10459592; rs12592697; rs2899472; rs6493487; and rs2899470. tSNPs were chosen using HapMap database build 36, using a minor allele frequency of ≥20%, r 2 ≥0.80, and Caucasian ancestry. The functional SNP, rs700519 (Arg264Cys), and tSNP, rs6493496, were genotyped using 5′ exonuclease Assay-on-Demand TaqMan assays (AB17000 and SDS 2.0 software; Applied Biosystems, Foster City, CA). The i-PLEX Gold SNP Assay was used to genotype the other three functional SNPs, rs28757184 (Thr201Met), rs2236722 (Trp39Arg), and rs56658716 (Met364Thr), and 17 tSNPs.
Demographic and clinical variables
Independent variables included age, gender, initial GCS, body mass index (BMI), length of hospital stay (LOS), mechanism of injury, and injury type. The GCS was taken within 8 h of injury in a manner to limit the influence of alcohol, sedatives, or paralytics. Injury type was determined by clinical CT radiology reports.
Outcome measures
Outcome measures included acute care mortality and Glasgow Outcome Scale (GOS) scores at 6 months post-TBI. The GOS categorizes global functional outcomes into five levels: 5=good recovery; 4=moderate disability; 3=severe disability; 2=persistent vegetative state; and 1=death, 17 GOS categories were collapsed into 1 versus 2+3 versus 4+5 for analysis.
Statistical analysis
Statistical analyses were performed using SPSS (v20.0; SPSS, Inc., Chicago, IL) and SAS statistical software (version 9.2; SAS Institute Inc., Cary, NC). Summary statistics, including medians, means, and standard error of the mean (SEM), for continuous variables were computed. Frequencies and percentages were determined for categorical variables. The chi-square test, with Fisher's exact test, when appropriate, was used to determine associations between SNP genotypes, covariates as well as acute mortality and GOS-6 scores. The choice between parametric and nonparametric tests was based on the distribution of the variables being analyzed. Parametric tests were used when no violations of the normality assumption were observed. Otherwise, nonparametric bivariate tests were utilized. Because of skewed data distributions for mean hormone levels and ratios, associations between descriptive variables and mean CSF/serum hormone levels were analyzed using Mann-Whitney's test, Kruskal-Wallis' test, and Spearman's rank correlation, as appropriate.
Week 1 average CSF and serum estradiol and testosterone levels were calculated by first averaging available hormone measurements within each 24-h period post-TBI if multiple samples were taken on the same day. Means for the 7-day period were then calculated. CSF estradiol/testosterone ratios (E2:T) were calculated by dividing estradiol (pg/mL) by testosterone (pg/mL) for each individual sample at each day, after which mean ratios were calculated. Subjects had at least two ratios available for analysis. If subjects had a time point that included one hormone level but not the other, that time point was excluded from the ratio calculation. Nominal pairwise analysis was performed to describe daily hormones and ratios that comprised the mean values used for bi- and multi-variate analysis.
Eighteen tSNPs and four functional SNPs for the aromatase gene were analyzed. The functional SNPs, rs28757184 (Thr201Met), rs2236722 (Trp39Arg), and rs56658716 (Met364Thr), were monomorphic, with 100% having genotype C/C, T/T, and T/T, respectively, in our population. Thus, they were excluded from further analyses. We then performed analysis to determine that remaining SNPs were in Hardy-Weinberg's equilibrium as a test of data integrity and also to identify SNPs whose allegic frequencies were statistically independent of each other at the p≤0.05 level using the chi-square test with Fisher's exact test, as appropriate.
After the number of SNPs was reduced to three statistically independent SNPs, we then used chi-square tests, and Fisher's exact test, as appropriate, to examine bivariate associations between these SNPs and categorical outcomes. Statistically independent SNPs included for further analysis were rs2470152, rs4646, and rs1470144. Risk variants were identified as the variant at each of these three SNP locations as being more likely to be associated with poor outcomes. Because of a skewed distribution for hormones and their ratios, associations among CSF/serum hormone averaged over the first week, SNP genotypes, and outcomes were determined using Kruskal-Wallis' test. Allelic associations with hormone ratios and outcome were also performed.
Multi-variate analyses (linear and ordinal logistic regression) were performed to identify which variables were associated with hormone levels and with outcome. For linear regression, data were log transformed to meet the assumptions for normality required for this regression-modeling procedure. Independent variables, as well as covariates associated in bivariate analysis with hormones and/or outcome (at p<0.20), were initially included in the multi-variate models. Relevant interaction terms were generated to assess interrelationships between the primary genotypes, other covariates, and outcome. Final multi-variate models included covariates significant at the α=0.05 confidence level, except E2/T ratios, which was forced into its outcome model. The combined effects of hormones and genetic variant effects on outcome were also tested using multi-variate analysis. A cumulative total risk was generated using a simple counting approach with each of the three SNPs significant in single-SNP multi-variate analysis described above and while taking into account the effects of demographic and clinical variables and hormone levels. This variable was then dichotomized into presence of zero to one risk variants versus two to three risk variants; and its effect on outcome was assessed using multi-variate ordinal logistic regression.
Results
Population description: General
One hundred and ten TBI subjects fulfilling inclusion criteria had genotype analysis for the aromatase gene and outcome data collected at 6 months postinjury. Women comprised 18.2% of the population, mean patient age was 34.8±1.5 years, and median GCS score was 6. The mean BMI of the population was 27.2±0.6. Mean acute care LOS was 22.0±1.1 days. Eleven percent of the cohort participated in the COBRIT study and 15% in the hypothermia study. Primary mechanisms of injury were automobile accidents (38.2%), motorcycles (21.8%), and falls or jumps (17.3%). More men (71%) had subdural hematomas, compared to women (45%; p=0.036). For GOS-6 scores, 22.7% had a GOS of 1, 40.9% had a GOS of 2 or 3, and 36.4% had a GOS of 4 or 5. In our control population (n=13), 53.8% were women, and the average age was 38.7±14.7 years.
Bivariate analysis
Population description: genes
Genotype distributions for the primary SNPs assessed in this study are provided as follows. Genotype distributions for SNP rs2470152 showed 21.7% of subjects with C/C and 78.3% T-allele carriers. For SNP rs2470144, 34.3% had A/A and 65.7% were G-allele carriers. For SNP rs4646, 63.0% had C/C and 37.0% were A-allele carriers. Table 1 summarizes bivariate analysis of covariates by genotype. There were no significant differences in genotype frequencies with respect to sex, GCS, BMI, citicoline versus control, or hypothermia versus control based on genotypes.
rs240144 had missing genotype information for 5 subjects.
rs2470152 had missing genotype information for 4 subjects.
rs4646 had missing genotype information for 2 subjects.
p values based on chi-square analysis, Fisher's exact test, analysis of variance, and Kruskal-Wallis' test, as appropriate.
TBI, traumatic brain injury; GCS, Glasgow Coma Scale; BMI, body mass index; SEM, standard error of the mean; COBRIT, Citicoline Brain Injury Treatment study; GOS, Glasgow Outcome Scale; Veg, vegetative; Mod, moderate; CSF, cerebrospinal fluid.
Outcome: genes
Table 1 summarizes bivariate analysis comparing SNPs to outcome measures. SNPs were not associated with acute mortality. For rs2470144, A/A-genotype carriers had a significantly higher proportion of worse outcome (p=0.046), compared to G-allele carriers. rs2470152 C/C-genotype carriers tended to have fewer deceased subjects and more subjects with moderate/good GOS outcome, compared to the rest of the cohort (p=0.065), and those with the rs4646 C/C genotype also tended to have worse outcome, compared to A-allele carriers (p=0.103), but differences did not reach statistical significance.(Figure 1). There were no significant associations with outcomes in allelic analysis for each of these three SNPs (data not shown). Taken together, the three SNPs assessed here suggest three risk variants that are associated with (or trending toward) worse outcome and are assessed further in multi-variate analysis.

GOS 6-month outcome results by genotype in the TBI population. GOS categories included dead (GOS=1), vegetative-severe (GOS=2, 3), and moderate-good (GOS=4, 5). Categories were graphed in percentages of total genotype cohort. (
Outcome: covariates
Table 2 summarizes associations of covariates gender, GCS, age, and BMI with outcome at 6 months postinjury. Older age was associated with greater acute mortality (p=0.009) and worse GOS (p<0.001) scores. Gender and BMI were not associated with outcome. Hypothermia treatment was associated with better 6-month GOS outcome (p=0.003) in this cohort, but citicholine did not have an effect on outcome (p=0.946). These data helped guide formulation of multi-variate models presented below.
p values based on chi-square analysis, Fisher's exact test, analysis of variance, and Kruskal-Wallis' test, as appropriate.
GOS, Glasgow Outcome Scale; GCS, Glasgow Coma Scale; BMI, body mass index; SEM, standard error of the mean.
CSF hormone levels: other covariates and outcome
Figure 2 shows daily CSF estradiol, testosterone, and E2/T ratios, compared to values obtained in healthy controls. Daily CSF estradiol levels were similar between men and women, but lower than controls, on days 0–4 and day 6 during the time course evaluated (p<0.035, all comparisons). However, daily CSF testosterone levels were higher than controls for the whole cohort on day 0 and lower than controls on day 6 (p<0.019, all comparisons). CSF testosterone was significantly higher in men, compared to women, with TBI on day 1 (p<0.035). There were no sex differences with E2/T ratios across the time course. E2/T ratios were lower than controls for the whole cohort on days 0–3 (p<0.028, all comparisons), but increased over time.
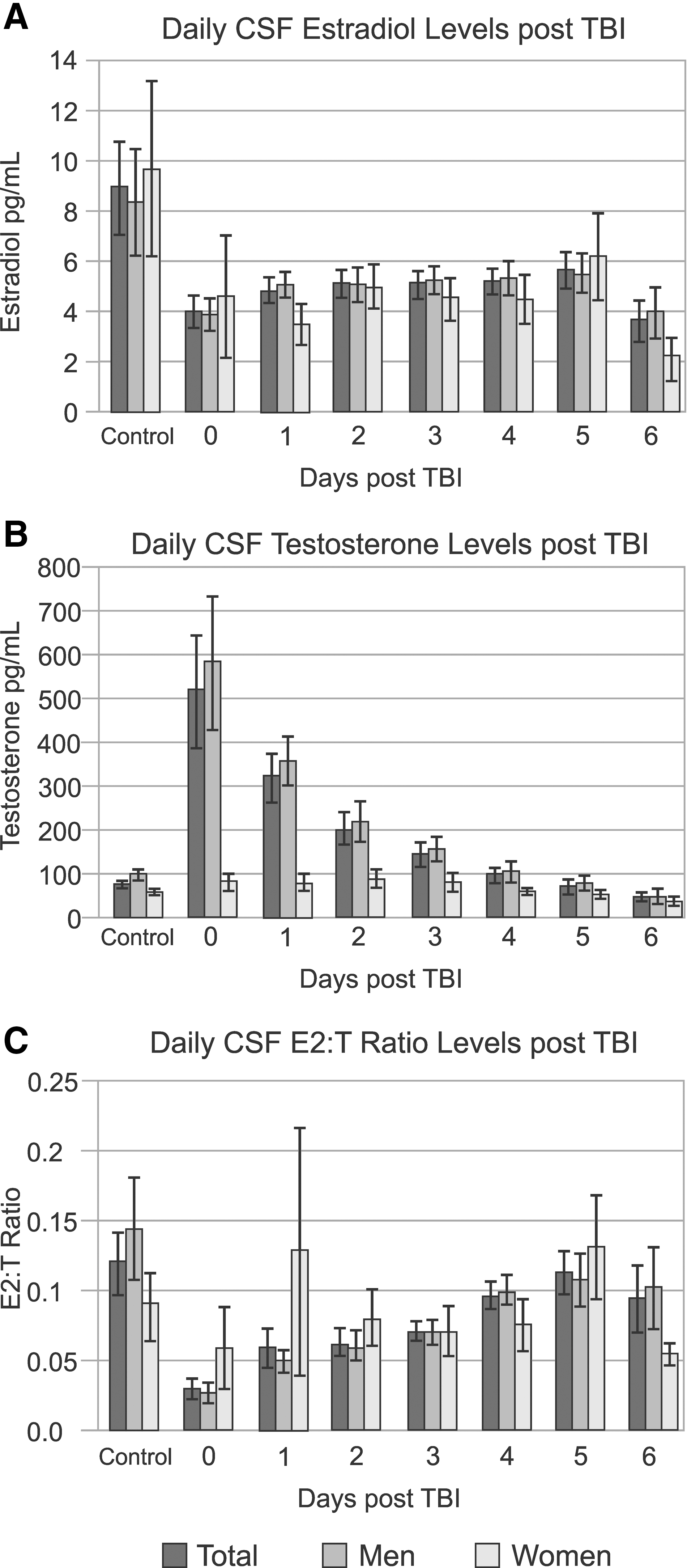
Daily CSF hormones levels in subjects post-TBI, compared to controls for (
In contrast to longitudinal profiles outlined above, mean CSF estradiol, testosterone, and E2/T ratios over the first week were calculated for each subject. TBI subjects, compared to controls, had significantly lower estradiol levels; a 54.6% decrease (4.90±0.35 vs. 8.98±1.85 pg/mL; p=0.001). Within the TBI population, there were no significant sex differences in CSF estradiol, though men had significantly higher CSF testosterone, compared to women (247.32±38.83 vs. 71.08±9.42 pg/mL; p=0.034). Within controls, there were no differences in CSF estradiol or E2/T ratios between men and women. However, male controls did have significantly higher CSF testosterone than women (99.06±10.62 vs. 62.14±6.07 pg/mL; p=0.009). These findings imply the possibility of sex-specific aromatase activity in the CNS under both physiological and pathological conditions, although other sex-specific activity and levels for metabolic enzymes within this pathway could be considered.
BMI, GCS, citicoline, and hypothermia treatment were not associated with CSF estradiol, testosterone, or E2,T ratio. Similar to reports regarding acute serum hormone levels, 10 older subjects had higher estradiol (p=0.033) and testosterone (p=0.051) levels. E2/T ratios were not affected by age. Average CSF testosterone and estradiol were not associated with outcome. However, higher CSF E2/T ratios were associated with lower acute mortality rates (p=0.019) and better GOS-6 scores (p=0.030).
CSF hormone levels: genes
In bivariate analysis, no SNPs were associated with average estradiol or testosterone levels (see Table 1). However, for rs2470152, C/C-genotype carriers had significantly higher E2/T ratios than T-allele carriers (0.07±0.01 vs. 0.10±0.02; p=0.026), and T-allele carriers had significantly lower E2/T ratios than controls (0.06±0.01 vs. 0.12±0.02; p=0.012; Fig. 3A). Allelic analysis substantiated the variant comparisons by showing that C carriers had significantly higher E2/T ratios than T carriers (0.09±0.01 vs. 0.06±0.01; p=0.044), and T carriers had significantly lower mean E2/T ratios than controls (0.04±0.01 vs. 0.12±0.02; p=0.004; Fig. 3D). For the rs2470144 variant and allelic comparisons, there were no significant differences with E2/T ratio between TBI genotypes (Fig. 3B). For rs4646, relatively increased E2/T ratios in the C/C genotype group did not reach statistical significance (0.06±0.01 vs. 0.09±0.01; p=0.059), when compared to the rest of the TBI cohort. However, A carriers had significantly lower E2/T ratios than controls (0.06±0.01 vs. 0.12±0.02; p=0.001; Fig. 3C). Allelic analysis among TBI groups showed that E2/T ratios for C carriers were significantly higher than E2/T ratios for A carriers (0.08±0.01 vs. 0.05±0.01; p=0.018), and A carriers had significantly lower ratios, compared to controls (0.05±0.01 vs. 0.12±0.02; p<0.001; Fig. 3F). Taken together, these results show that genetic associations with CSF hormone profiles among TBI groups, and among genetic variant groups for associations within the TBI population versus healthy controls, are distinctly different for each of the three statistically independent SNPs. These varied genetic associations with CSF hormone levels, in the context of TBI, are not surprising given the heterogeneity and regulatory complexity of the aromatase gene.
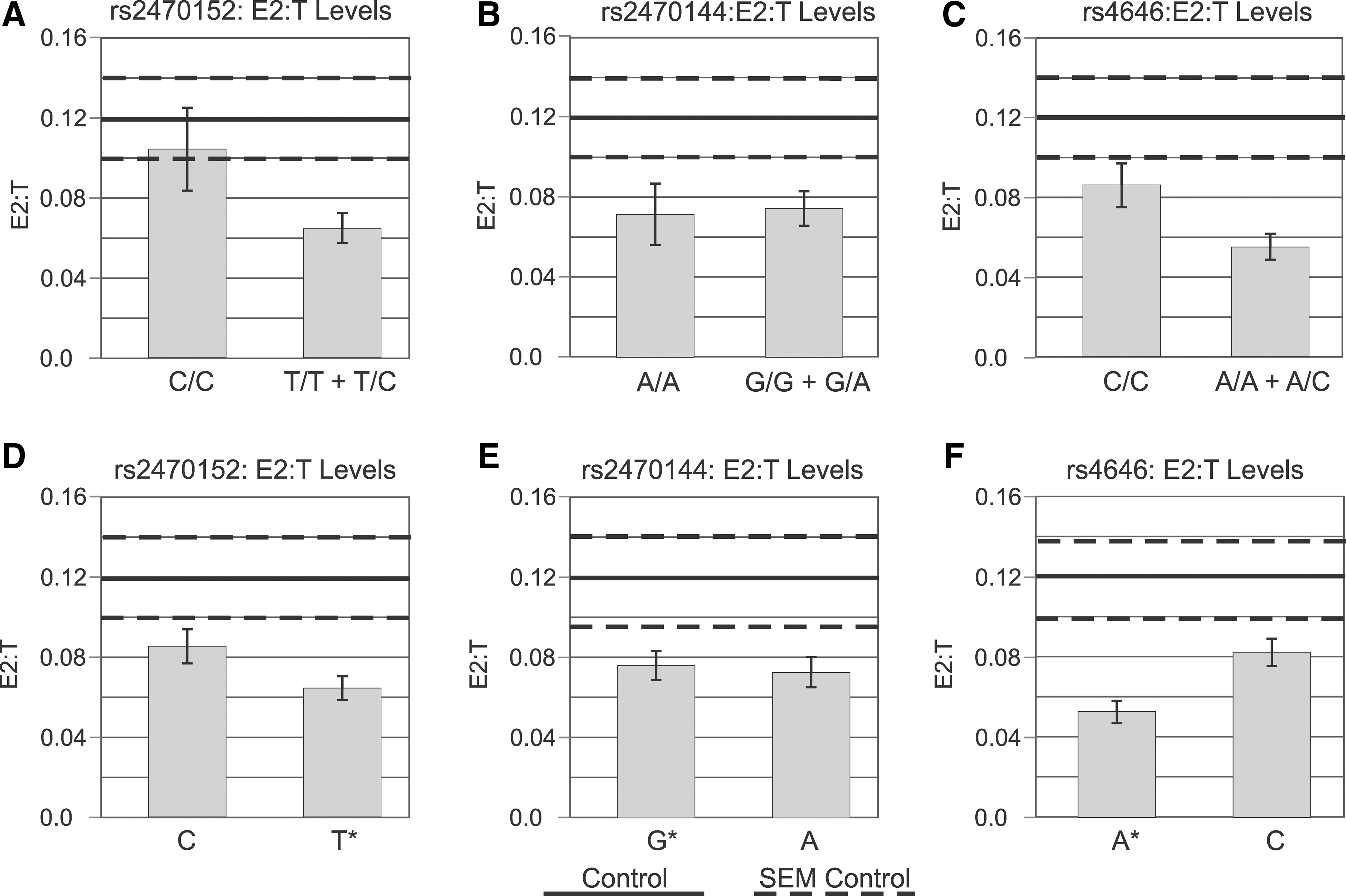
Week 1 average CSF E2/T ratios by genotype with control ratios. TBI subjects had lower ratios, compared to controls (p=0.052). Genotypes and alleles were compared within single-nucleotide polymorphisms using risk allele T for rs2470152 (
Serum hormone levels: genes and CSF hormone associations
Mean serum estradiol, testosterone, and E2/T ratios over the first week were calculated. In bivariate analysis, no SNPs were associated with average serum estradiol or E2/T ratios. For rs2470152, subjects with the C/C genotype had significantly lower testosterone levels than T-allele carriers (2.56±0.56 vs. 3.90±0.40 nmol/L; p=0.010). Neither rs2470144 nor rs4646 were associated with serum testosterone levels (Table 1). When correlating mean CSF hormone levels with mean serum hormone levels, we found no correlation between CSF and serum testosterone (R 2 =0.0202; n=245 samples; N=88 patients), but a moderate correlation between serum and CSF estradiol (R 2 =0.4127; n=240 samples; N=86 patients). These results suggest that serum estradiol may contribute, in some way, to CSF estradiol levels observed and/or that injury affects some elements of estradiol physiology similarly, irrespective of production location in the periphery versus CNS. Known age-related increases in both CSF and serum hormone levels post-TBI may be one contributor to this association.
Single SNP multivariate associations
CSF hormone levels
Multivariate linear regression analysis incorporated the covariates of gender, age, GCS, and BMI when evaluating SNP associations with average estradiol, testosterone, and E2/T ratio (models not shown). For rs2470152, average E2/T ratios were increased among those with C/C genotypes, compared to T carriers (p=0.019), and there were no significant covariate associations.
Outcome
Multi-variate ordinal logistic regression analysis examining single gene-variant associations with outcome were adjusted for age and GCS in final models. There were no associations between SNP genotypes and acute care mortality in single SNP models. However, subjects who were T carriers for rs2470152 had worse outcomes, based on GOS-6 scores post-TBI, compared to those with the C/C genotype (odds ratio [OR]=2.945; p=0.034). Subjects with the rs2470144 A/A genotype had significantly worse GOS-6 scores post-TBI, compared to G carriers (OR=2.333; p=0.040). Also, subjects with the rs4646 C/C genotype had worse GOS-6 scores, compared to A-allele carriers (OR=2.262; p=0.041). Age and GCS were associated with mortality and GOS-6 scores in all three SNP multivariate models (p<0.034 all comparisons). When examining E2/T ratios within each of the single SNP multivariate models, E2/T ratios were significantly associated with GOS-6 scores when paired with the rs4646 variant (Table 3). Other E2/T ratio associations with outcome, while adjusting for other gene variants, did not reach significance (models not shown). These results suggest that genetic variation within the aromatase gene influences global outcome, and that interrelationships between gene variants and hormone levels may have some shared prognostic variance with respect to GOS-6 scores.
OR, odds ratio; CI, confidence interval; GCS, Glasgow Coma Scale.
Multi-SNP multi-variate analysis
We tested the relationship between cumulative risk variants on outcome, using a simple counting approach with each of the three SNPs significant in single SNP multi-variate analysis above and while taking into account the effects of demographic and clinical variables and hormone levels (Table 3). Multi-variate ordinal regression analysis demonstrated that those carrying two to three of the risk variants alleles had worse GOS-6 scores than subjects carrying only zero to one risk variant (OR=4.758; p<0.001). Higher genetic risk variants were also associated with acute mortality (OR=3.510; p=0.048; full model not shown). These results indicate that the three SNPs are not only statistically independent, but also contribute independently to the cumulative risk for poor GOS scores at 6 months.
Discussion
This study demonstrates, for the first time, that TBI results in significantly decreased CSF estradiol levels, increased testosterone, and decreased in E2/T ratios over time. These findings in CSF contrast with low serum testosterone and relatively normal serum estradiol overall for TBI subjects, compared to controls. 10 In bivariate analysis with the TBI cohort, those who had higher CSF E2/T ratios had lower acute mortality rates and better GOS-6 scores. T-allele carriers for rs2470152 had significantly worse outcomes as well as lower CSF E2/T ratios, suggesting that the T allele may be a risk allele. It is possible that the effects of the T allele on outcomes may result, in part, from the effects of this allele on CSF E2/T ratios. G-allele carriers for rs2470144 had significantly better outcome, suggesting that the G allele is protective. Also, A-allele carriers for rs4646 had better GOS scores, albeit with lower CSF E2/T ratios, suggesting that it may be protective. These findings suggest that dynamic changes between CSF estradiol and testosterone, as well as genetic variation in the aromatase gene, influence global outcome after sTBI. The fact that risk for poor outcome increased with carriage of a higher number of “risk” variants in our multi-variant analysis, along with our data reduction approach a priori to reduce the primary SNPs evaluated to those that were statistically independent, suggests that SNP relationships with outcome presented each uniquely contribute to biosusceptibility for secondary injury.
The experimental literature suggests that estradiol has neuroprotective effects that can reduce the severity of brain damage. 19 –22 To date, it was unknown if or how CSF sex hormone levels change after TBI and what role genetic variation in the aromatase gene may play in both injury recovery and responses. Within controls, there were no sex differences in CSF estradiol; however, men had higher CSF testosterone levels, compared to women. These relationships between men and women were present in the TBI population, with higher CSF testosterone in men and no differences in estradiol. Together, these data support sex differences in aromatization in the CNS.
Mean CSF estradiol and testosterone were not associated with outcome; however, the relative balance of CSF hormones, as represented by E2/T ratios, was associated with both acute mortality and GOS-6 scores. This finding suggests that dynamic changes between the two hormones are more strongly associated with recovery, rather than individual levels. Overall, both men and women with sTBI had lower mean CSF estradiol, higher mean testosterone, and trends toward lower mean E2/T ratios, when compared to controls, a finding that may limit adequate neuroprotection in the setting of TBI. In fact, higher E2/T ratios, closer to the ratios observed in the control population, occurred in subjects with better outcomes.
Whether this relationship between higher ratios and outcome is reflective of simply (or more generally) less severe injury, or if normal E2/T ratios represent a relatively neuroprotected state, is hard to discern in this analysis. Changes in CSF testosterone tended to influence the E2/T ratio more than changes in CSF estradiol over time. Although estradiol is considered neuroprotective, some literature suggests that testosterone exacerbates CNS damage. 23 –25 Given our findings suggesting that higher ratios are protective, the dynamic balance between availability of testosterone as a substrate for estradiol, in contrast to the potential detrimental effects of testosterone on CNS function and survival, may be what the CSF E2/T ratios capture that is relevant to TBI outcome. For rs2470152, subjects with the C/C genotype had lower serum testosterone, compared to the remaining TBI cohort. As the primary enzyme catalyzing estradiol from testosterone, the relationship between aromatase genetics and CSF hormone associations with outcome may be influenced by both CNS and peripheral production.
Importantly, TBI is associated with an acute hypopituitary state, resulting largely from increased stress, and possibly elevated cortisol levels, leading to disruption of the hypothalamic/pituitary/gonadal axis. 10 Hypogonadism, for some, can be persistent. 13 The influence of peripheral hormone production on CSF hormone levels is complex. But, after TBI, the BBB is disrupted, temporarily allowing increased permeability 25 occurring over hours to days. Although BBB permeability is compromised initially after injury, and these hormones can cross an intact BBB, a significant contribution of peripheral hormones to CNS hormone levels is not strongly supported by our data.
Serum testosterone (over time) drops rapidly after TBI and follows a similar decreasing time course as with CSF levels observed in our analysis. 10 Although CSF testosterone also declines, it remains higher than controls for several days postinjury. Despite a similar pattern of decline, associations between mean CSF and serum testosterone levels showed no correlation. There could be multiple contributors to this finding. We hypothesize that there is differential activation of tissue-specific promoters for aromatase, with impaired CNS and overactive peripheral aromatase activity. The gonadotropins regulating gonadal synthesis of hormones in the periphery are greatly reduced very early after injury, 10 limiting the potential physiological effect of peripheral testosterone synthesis on CSF levels primarily to that synthesized in the adrenals. Although speculative, injury and genetic effects on sex-hormone–binding globulin (SHBG)-bound testosterone versus estradiol in the periphery may contribute to low correlation for CSF versus serum values. 27,27 Interestingly, some literature suggests that the biologically active free fraction of cortisol and cortisol-binding globulin concentrations are affected by brain injury, 29 and it may be informative to evaluate if and how trauma-induced changes occur with SHBG to affect sex hormone availability in the periphery for BBB passage after TBI.
As our previous literature shows, serum estradiol levels are elevated for many subjects with TBI. 10 Moderate correlations between CSF and serum estradiol may reflect less dramatic compartmental dynamics in hormone production over time, compared to testosterone. However, aromatase effects in the brain largely result in local increases in estradiol levels, 20 and changes in aromatase activity can influence estradiol levels within minutes. 1 As such, genetic variation in brain-tissue–specific promoter I.f 7 could have a considerable effect on local activity post-TBI. Based on this literature, persistently low estradiol in the CNS may suggest that the majority of CSF estradiol in our cohort was synthesized in the CNS. CNS aromatase function may improve because CSF E2/T ratios rise over time, yet CSF estradiol remains depressed throughout the time course. One alternative contribution to this finding simply may be decreased CNS availability of testosterone at later time points, because CSF testosterone concentrations are actually lower than control values. Despite this possibility, subjects with greater CSF E2/T ratios, influenced either by lower testosterone production or through genetic and/or temporal variation in aromatase function, have better outcome that reached statistical significance in bivariate analysis and also in multi-variate analysis, when modeled in conjunction with rs4646.
Clinical serum studies have associated high estradiol with worse outcome in severe trauma and critically ill populations. 30,30 In our study characterizing serum hormones acutely after TBI, elevated estradiol in particular was associated with mortality and poor outcomes 6 months postinjury. 10 These findings suggest that tissue-specific aromatase activity significantly influences the hormone profiles, and there are different mechanisms of hormone physiology at play in the CSF versus serum compartments. Also, though aromatase conversion of testosterone to estradiol in the CNS has some physiological role in neuroprotection, aromatase conversion of testosterone to estradiol (presumably) in adipose tissue largely represents an acute stress response (e.g., by tumor necrosis factor alpha, leading to increased peripheral aromatization). 3,3 Notably, increased serum estradiol and testosterone levels were associated with higher BMI in our previous studies. 10
Although the translated protein product is the same, each tissue expresses a different promoter region of aromatase: I.f for brain; I.3 and I.4 for adipose; and PII for ovary. 5 In the CNS, astrocyte cells manage both glutamate metabolism and neurosteroidogenesis, particularly, androgen production. However, neurons are the primary cell type that converts testosterone to estradiol. 33 Evidence that brain aromatase activity is acutely altered by glutamate agonists 33 provides further evidence that CSF estradiol post-TBI may be more controlled by local neurotransmitter levels and signaling pathways, rather than early BBB permeability from serum concentrations. Although excitatory signaling may be physiologically beneficial in regulating CNS aromatase activity and neurosteroidogenesis under normal conditions, perhaps acute excitotoxicity, and the cellular dysfunction that ensues, may limit effective aromatase activation when needed for CNS neuroprotection. Selective vulnerability of neurons to injury after TBI, relative to astrocytes, may be another hypothesis for decreased aromatase activity, specifically, leading to decreased CSF estradiol and increased testosterone, compared to controls. Ongoing damage from secondary injury may limit androgen production and may contribute to reduced CSF testosterone over time.
We used a tSNP approach, and statistically independent tSNPs, to characterize relatively unique blocks of DNA associated with hormones and/or outcome. Interestingly, whereas variability in each of our three independent SNPs evaluated was associated with better outcomes, they each had a different relationship with CSF E2/T ratios, with lower ratios associated with the rs2470152 (risk) T allele, no difference in ratios for rs24701144, and higher ratios for the rs4646 (protective) A allele. Yet, higher CSF E2/T ratios were associated with better outcome in bivariate analysis and some multivariate models, providing indirect evidence that DNA blocks represented by these tSNPs associated with hormones may differentially influence CNS versus peripheral hormone production.
rs2470152 is an SNP in intron 1 located in the I.4 promoter region. 27 Several studies report associations between rs2470152 and serum hormone levels, although the findings are mixed. 27,27 Importantly, the rs2470152 locus itself is a potential binding site for a transcription protein cyclic adenosine monophosphate response element (CRE). 36,36 In our study, T-risk-allele carriers had lower CSF E2/T ratios and worse GOS-6 scores. Importantly, the T allele was linked with higher testosterone in both serum and CSF. Because rs2470152 resides inside the adipose tissue promoter, 37 our findings showing testosterone differences in both serum versus CSF may be consistent with the potential functionality of this SNP, despite the limited correlation of testosterone across compartments. Because gene transcription is often affected by TBI, 38 injury-mediated changes in CRE activated transcription for the aromatase gene may be one mechanism by which this SNP is associated with testosterone in both compartments. rs2470144 is a tSNP located in intron 1 on the 5′ region of the gene. 39 When comparing A/A to G carriers, the latter group had better GOS-6 scores without differences in CSF or serum hormone levels, suggesting that the G allele may contribute to better outcomes, outside of a specific role on acute hormone levels.
rs4646 is a tSNP located on the 3′ untranslated region of the gene 40 and is in close proximity to the functional SNP, rs700519, a nonsyonymous coding SNP. The nucleotide exchange results in an amino acid change from arginine to cysteine. 41 Several breast cancer studies report on genetic influences at this locus with regard to hormone levels 41 and treatment effects. 40,40 A allele carriers had significantly lower CSF E2/T ratios, yet they tended to have better GOS-6 scores. The rs4646 A allele also tended to be linked to lower serum E2/T ratios. The rs4646 A allele association with lower CSF ratios and better outcomes contrasts with findings showing that elevated CSF E2/T ratios in general are linked to better outcomes. Interestingly, it is only in the rs4646 model that both CSF E2/T ratios and the risk allele for outcome were each independently associated with GOS scores (see Table 3). Taken together, this finding indirectly suggests that the primary effect of this SNP on outcome may be mediated through a reduction in peripheral E2/T ratios; however, there was only a nonstatistically significant trend for serum E2/T ratios to be influenced by variation in the rs4646 SNP.
In addition to genetic influences on hormones and outcome, other factors affected outcome and hormone levels. Age was an important predictor of acute mortality and GOS-6 and was associated with the rs2470152 T allele. However, even with adjusting for age, this SNP was still associated with CSF E2/T ratios and outcome. Older age was associated with higher CSF estradiol and testosterone levels, but not ratios. Serum estradiol is elevated in our previous report, and this finding may serve as indirect evidence that there is some degree of estradiol transport from the periphery to the CSF after TBI. Sex did not influence outcome. However, to date, there are mixed results on whether sex influences outcome after TBI. 44 –46 BMI also did not influence outcome or CSF hormone levels, despite the influence of BMI on serum estradiol and testosterone. 10 Subjects receiving hypothermia had better GOS-6 scores in bivariate analysis; however, findings in this small cohort do not generalize to the multi-variate models presented or to the findings from the previously reported multi-centered study on hypothermia 47 and should be interpreted with caution. Although not significantly affecting CSF hormone levels in our population, we cannot rule out more-subtle effects of these interventions on hormone profiles. Despite the possibility of increased heterogeneity in hormone profiles resulting from treatment variability, genetic associations with hormones and hormone associations with outcome were still identifiable in this population. In addition to the neuroprotective effects in experimental TBI, clinical studies suggest that pharmacological dosing of progesterone can improve neurological outcomes after TBI. 48 However, progesterone effects on other hormones and/or aromatase activity have not been conducted. Further, the role of estradiol treatment in the TBI population is still unknown.
Although this study is novel in its presentation of CSF hormone profiles and aromatase gene associations with outcome, there are some limitations. Frequencies of genetic variation for the aromatase gene differ among ethnic groups, and there is evidence that the association between aromatase genetic variations and serum hormone levels can differ based upon race. 14 Because our sample size for non-Caucasians would be uninformative, we limited our analyses to Caucasians. 27,27 Future work with larger populations may allow for stratification by race. Also, the small sample size decreases the ability to discriminate subtle genetic effects. Larger sample sizes will be required to validate the findings presented in this initial exploratory study. The averaged levels calculated for estradiol, testosterone, and E2/T ratios used in multivariate analysis do not address fully the dynamic changes that occur for specific subpopulations the first week, and longitudinal hormone analysis 10,10 may be informative. Because estradiol measurements for TBI subjects were below controls, the sensitivity in the lower range of the kit may be a limitation.
In summary, we report that TBI creates an environment where CSF estradiol levels are significantly lower than healthy controls, and higher E2/T ratios are associated with better outcome in some models. Further, genetic variation at rs2470152 influences CSF and serum hormone levels and global outcome. Specific analysis with individual SNPs, as well as cumulative genetic risk analysis, demonstrates that multiple elements of aromatase gene function may independently influence TBI physiology and outcome. Larger sample sizes for validation studies are needed, particularly to evaluate if and how some associations hold in an independent population. Future directions include targeted DNA haploblock exploration of aromatase gene variation and functionality in proximity to each of these SNPs reported here. Also, investigating CNS cortisol profiles over time, and how these profiles may influence CNS estradiol and testosterone, may be of interest. Importantly, further studies regarding the role of tissue-specific aromatase activity on TBI pathology is warranted. If and how sex hormone binding globulins and BBB permeability contributes to CNS and peripheral hormone profiles may also be informative. Future work assessing genetic relationships of free and bound hormones, as well as more-refined outcome measures among TBI survivors, is warranted.
Footnotes
Acknowledgments
The authors thank Sandra Deslouches for her technical support and the Brain Trauma Research Center for some elements of data collection. This work was supported by the Centers for Disease Control and Prevention (R49 CCR323155), the Department of Defense (W81XWH-071-0701), and the National Institutes of Health (5P01NS030318, R01NR008424, and R01 HD048162).
Author Disclosure Statement
No competing financial interests exist.