
Abstract
Microparticles are cell-derived, membrane-sheathed structures that are believed to shuttle proteins, mRNA, and miRNA to specific local or remote target cells. To date best described in blood, we now show that cerebrospinal fluid (CSF) contains similar structures that can deliver RNAs and proteins to target cells. These are, in particular, molecules associated with neuronal RNA granules and miRNAs known to regulate neuronal processes. Small RNA molecules constituted 50% of the shuttled ribonucleic acid. Using microarray analysis, we identified 81 mature miRNA molecules in CSF microparticles. Microparticles from brain injured patients were more abundant than in non-injured subjects and contained distinct genetic information suggesting that they play a role in the adaptive response to injury. Notably, miR-9 and miR-451 were differentially packed into CSF microparticles derived from patients versus non-injured subjects. We confirmed the transfer of genetic material from CSF microparticles to adult neuronal stem cells in vitro and a subsequent microRNA-specific repression of distinct genes. This first indication of a regulated transport of functional genetic material in human CSF may facilitate the diagnosis and analysis of cerebral modulation in an otherwise inaccessible organ.
Introduction
MP-mediated transcellular delivery is hypothesized to be of special importance in the blood by enabling the specific targeting of distal cells 8 and by preventing dilution of information in this expansive circulating system. Reports of MPs in cerebrospinal fluid (CSF), which is pumped around an analogous circulatory system comprising the ventricles, the subarachnoid space, and spinal cord, are in contrast limited. 10,13 –16 The rationale for MP-based communication is nonetheless equally strong.
The morphological and proteomic analysis performed by Harrington and coworkers 17 that demonstrated the presence in CSF of discrete spheres associated with biochemically distinct components, such as prostaglandin H synthase, acetylcholine, or adenosine diphosphate-ribosylation factor protein, presently provides the best evidence that CSF contains MPs. In parallel intercellular RNA transfer by exosomal microvesicles was considered to be a possible mode of signaling within the nervous system. 18,19 This assumption was, however, focused mainly on intersynaptic communications.
This study is based on the hypothesis that CSF harbors a communication network comparable to the system described in blood. It was our aim to establish the presence of MPs in the CSF (CSF-MPs) derived from non-injured subjects (NIS) and traumatic brain-injured patients (TBIP) and to analyze their potential as vehicles that shuttle distinct biological and genetic information.
Methods
Ethics statement
The ethics committee of the Medical University of Graz specifically approved this study on February 10, 2009. The study and the potential to participate were explained to patients before they were discharged from the clinic. At 4 to 6 months later, letters were sent to the patients with the same information and the option to call for more information. Only patients who consented to the study by returning a signed form by mail at this time point were included. Six of the trauma patients called back for more information before sending a signed form. In two cases, guardians consented on behalf of participants.
Clinical parameters of TBIPs and healthy controls included in the study
CSF samples were collected from patients with severe TBI traumatic brain injuries (Glasgow Coma Scale [GCS]≤8) when ventricular drainage was implemented as a measure of intensive care treatment. Because of ethical considerations, samples were not taken at standardized time points but rather when ventricular drainage was indicated from acute increased intracranial pressure. A higher risk of ventricular drain infections resulting from additional study dependent interventions was thereby avoided.
There were 26 samples collected from 11 patients over 2 years. The number of samples per patient and the time points of sample collection relative to the time point of primary injury varied considerably (time of sampling corresponding to the time of injury is indicated in the respective figures). Further, differences in the location and type of brain injury as well as additional injuries to different body parts and organ systems render the group of brain-injured patients a rather heterogeneous study population (Table 1). Hence, statistical analysis of results is a mere attempt to draw some general conclusions, but must be interpreted with caution. The analyzed parameters are additionally presented for each subject, thereby allowing for a more personalized interpretation of data.
The GCS initially diagnosed is provided.
CSF samples from adults who received a lumbar puncture for diagnostic purposes were collected for control studies (n=26). Only subjects without a confirmed subarachnoid hemorrhage or infection (confirmed central nervous system [CNS] diseases [CCD]) were included as non-injured controls (NIS, n=17; Table 1; see online supplementary Fig. S1,S2 at
Preparation of CSF
CSF was obtained from patients with TBI by ventricular drainage and from NIS by lumbar puncture. Samples were taken directly from the external ventricular drain tube. Sample volumes varied between 3–7 mL (TBIP) and 0.2–1 mL (NIS). Samples were stored no longer than 30 min at 4°C before further processing. CSF samples were centrifuged at 400 g for 5 min and filtered through a 0.45 μm syringe filter; 1 μL of Protease Inhibitor Cocktail (Sigma)/mL of CSF was added. The CSF was then stored at −80°C until further analysis. MPs were isolated by thawing CSF on ice, followed by ultracentrifugation (Optima L-90k; Beckman Coulter) for 40 min at 170,000 g at 4°C. Supernatant was carefully discarded, and the resulting MP pellets were processed according to the intended experiment.
Flow cytometric detection of MPs
There were 100 μL MP suspensions transferred into Trucount™Tubes (BD Bioscience) containing a known number of fluorescent beads (diameter: 1 μm) to enumerate MPs. Flow cytometry were performed using the LSR II BD Bioscience and the BD FACSDiva™ 6.0 software. Forward and side scatter were set in logarithmic scale. MPs were differentiated from signal noise by threshold settings of 200 forward scatter and 200 side scatter. The acquisition was terminated after 2000 bead counts. The number of MPs were calculated by the following formula described by Shet and associates 20 : MP [counts/mL]=[(total beads per tube/beads counted)×events counted]×dilution factor.
microRNA (miRNA) preparation, miRNA array, and bioinformatic analysis
For miRNA isolation, at least 50 mL of CSF pooled from 10 patients with TBI was centrifuged as described above. The MP pellet was prepared according to the protocol of the Qiagen miRNeasy Kit. Rat brain RNA and miRNA were isolated for use as internal controls. RNA and miRNA quality and concentrations were determined using a Bioanalyzer 2100 (Agilent).
MP samples and two miRNA rat brain samples were analyzed using the Affymetrix GCS300 (Affymetrix) with FlashTag Biotin HSR (Gensiphere).
The miRWalk (
Overrepresented neuron-related biological processes with a p value less than 0.001 were used for visualization. These processes were searched for in the initial GO network and rearranged without altering their connections to the other processes.
PCR-mRNA
For the polymerase chain reaction (PCR), the RNA concentration in the CSF was determined using the Ribogreen assay kit (Invitrogen) in accordance with the manufacturer's protocol; RNA content was measured using a POLARstar optima fluorometer. To detect mRNAs for activity-regulated cytoskeleton-associated protein (ARC), ß-ACTIN, microtubule-associated protein 2 (MAP2), and LIM domain kinase 1 mRNA (LIMK1), Dicer, fibroblast growth factor receptor 1 (FGFR1), and CD133 cDNA was synthesized with the First Strand cDNA Synthesis Kit (Fermentas) according to the manufacturer's protocol using 0.5 ng of total RNA. Real-time PCR (RT-PCR) was performed using the FastStart PCR Master Mix (Roche) and specific primers (Eurogentec) (ARC [412 bp]; ß-ACTIN [327 bp]; MAP2 [319 bp], LIMKI [264 bp]). PCR conditions were kept within the linear range determined for every product. Expression levels were normalized to the corresponding PCR product from NT2 cell cDNA. For primer sequences, see online supplementary Table S1 at
PCR miRNA
For miRNA analysis, NCode™ miRNA qRT-PCR Kits (Invitrogen) were used according to the manufacturer's protocol. cDNA was synthesized using 0.5 ng of total RNA. Universal Primer was provided in the reaction kit, specific miRNA primers were designed using the NCode™ miRNA Database (Invitrogen), and RT-PCR was performed using a Roche LightCycler® 480 according to the manufacturer's instructions. PCR for mRNA and miRNA was performed three times per cDNA sample; cDNA was prepared three times from each CSF sample.
PCR reaction products were separated on a 2–4% agarose gel, scanned, and band intensities quantified by densitometry using Photoshop CS5 software. For primer sequences see online supplementary Table S1 at
Electron microscopy
MP pellets were resuspended in 100 μL phosphate buffered solution (PBS) for electron microscopy. Then 10 μL of the sample were transferred to pioloform-and-carbon-coated grids, blot dried, and rinsed with water. Negative staining was performed by applying 2% uranyl acetate for 1 min. Samples were then air-dried and viewed with a Zeiss EM 901 transmission electron microscope.
Western blot
CSF pellets derived from CSF samples pooled from five patients were homogenized in 100 μL radioimmunoprecipitation assay buffer containing 5% sodium dodecyl sulfate (SDS), phenylmethanesulfonylfluoride (PMSF), iodoacetamide, and aprotinine (each 1 mM). Protein content was determined using a BCA Protein Assay Kit (Novagen®). Ten μg of protein was loaded on a 10% SDS-PAGE gel followed by transfer to nitrocellulose (Schleicher & Schuell; BA85) in a Tank Blotter (Biorad). Blots were blocked with 1% non-fat milk (Sigma) in Tris-buffered saline (TBS) for 3 h. Filters with specific antibodies were incubated overnight in blocking solution (rb α eIF2C 1:100 [argonaute2 (AGO2); Santa Cruz]; gt α Staufen (STAU2) 1:50 [Santa Cruz]; mou α GFI-1 1:500 [Sigma]), followed by 2×TBST and 3×TBS washing steps and incubation with biotinylated antibodies for 1 h. For visualization, blots were washed again and developed by the ABC-horseradish peroxidase (Vectashield) method using diaminobenzidine as chromogen.
Cytochemistry
Ten mL of CSF was incubated with 2 mg EZ-Link® Sulfo-NHS-Biotin (Thermo Scientific) for 30 min at room temperature (RT), followed by ultracentrifugation as described above. Pellets were resuspended in 400 μL PBS and streptavidin Cy5-conjugated (1:200; Invitrogen), and RiboGreen (1:100) was added and incubated for 30 min at RT. The fraction was then washed with 10 mL PBS and again ultracentrifuged as described above. Ten μL of the MP suspension was transferred to an object slide and air-dried at 37°C. The MPs were finally washed (3×PBS) and mounted with Aquatex (Merck) and examined under the microscope (amplification 40×; images were also digitally enlarged).
For antibody staining, MPs were treated with 1% bovine serum albumin (BSA)-PBS blocking solution for 1h. Antibody gt α hnRNP A2/B1 1:100 (Santa Cruz) was then incubated overnight followed by 3×PBS washing and incubation with biotinylated antibodies for 1 h. For visualization, slides were again washed and developed using the ABC-horseradish peroxidase method using diaminobenzidine as chromogen. Slides were mounted with Eukitt® (Kindler).
Incubation of NT-2 cells with MPs
NTERA2 clone D1 (NT2.cl.D1) embryonal carcinoma stem cells were grown and maintained in Dulbecco's Modified Eagle's Medium, supplemented with 10% fetal calf serum and 2 mM
For PCR analysis, cells were incubated with MPs containing 5 ng or 10 ng RNA. The RNA amount was measured using the RiboGreen assay. After 1 h and 3 h incubation time, cells were washed twice with 10 mL PBS harvested and RNA was isolated using the RNeasy micro Kit. RNA amount was measured using Nanodrop,® and cDNA libraries were prepared as previously described, followed by PCR for Dicer1, CD133, FGFR1, miR-451, and U6 as housekeeping gene as already described above.
Silencing of miR-451
NTERA2 cells were cultured as described above to a confluence about 30–50%. Cells were transfected using X-tremeGENE siRNA transfection reagent (Roche) combined with a final concentration of 50 nM of either hsa-miR-451 inhibitor (miRCURY LNA microRNA inhibitor) or scrambled (Negative Control A) (Exiqon) following the manufacturer's protocol and preincubated for 2 h. MPs were prepared as described above, and RNA content was measured using the RiboGreen assay. MPs with a content of 10 ng RNA were then supplied to the cells for 1 h. Cells were washed, harvested, and analyzed as described above.
Statistical analyses
Comparison of CSF-MPs (n/100 μL) and RNA content (ng/μL) in TBIPs and NIS as well as comparison of content of ARC, LIMKI, MAP2, and β-ACTIN in TBIPs and NIS were performed with the non-parametrical Mann-Whitney U test for independent samples. Statistical significance was set at the level of α=0.05.
Frequency of occurrence of miRNA species in CSF samples of TBIPs and NIS was evaluated statistically with the logistic regression analysis. NIS were defined as 0, TBI patients as 1; miRNA not present was defined as 0, miRNA present as 1. The Nagelkerke's R2 and the omnibus-test for statistical significance were performed in SPSS 18 (PASW). Statistical significance was set again at the level of α=0.05.
Comparison of CSF-MPs (n/100 μL) and RNA content (ng/μL) in NIS and CCD patients (see online supplementary Fig. S1,S2 at
Results
Quantification of MPs in CSF from patients with TBI and healthy subjects
CSF was collected from TBIP by ventricular drainage when indicated as a measure of intensive care treatment. Control CSF from NIS was obtained after lumbar puncture implemented for diagnostic purposes. CSF samples of subjects without subarachnoid hemorrhage or CSF infections were included as non-injured controls (NIS, Table 1). Patients diagnosed for subarachnoid hemorrhage or CNS infection (CCD) were not included in the comparative analysis of CSF-MPs. CCD patient diagnosis as well as the number of MPs and RNA content of respective samples are presented in Figure S1 (see online supplementary Fig. S1 at
CSF was collected and stored by a standardized protocol (see Methods). Putative MP-containing fractions were derived from CSF samples by serial centrifugation and filtration (<400 nm). High magnification transmission electron microscopy showed the samples to contain a heterogeneous population of intact particles ranging in size from 50 to 400 nm (Fig. 1A). An increasing number of reports suggest MPs to be membrane sheathed shuttles for a variety of RNA molecules and proteins.
11
Staining of the CSF-MPs with RiboGreen (RNAs) and membrane-specific Sulfo-NHS-biotin confirmed the presence of membrane covered particles that carry RNA molecules (Fig. 1B–D). Furthermore, mRNA in CSF was protected from RNaseI digestion underlining the finding that RNA is shuttled by membranous particles (see online supplementary Fig. S3 at
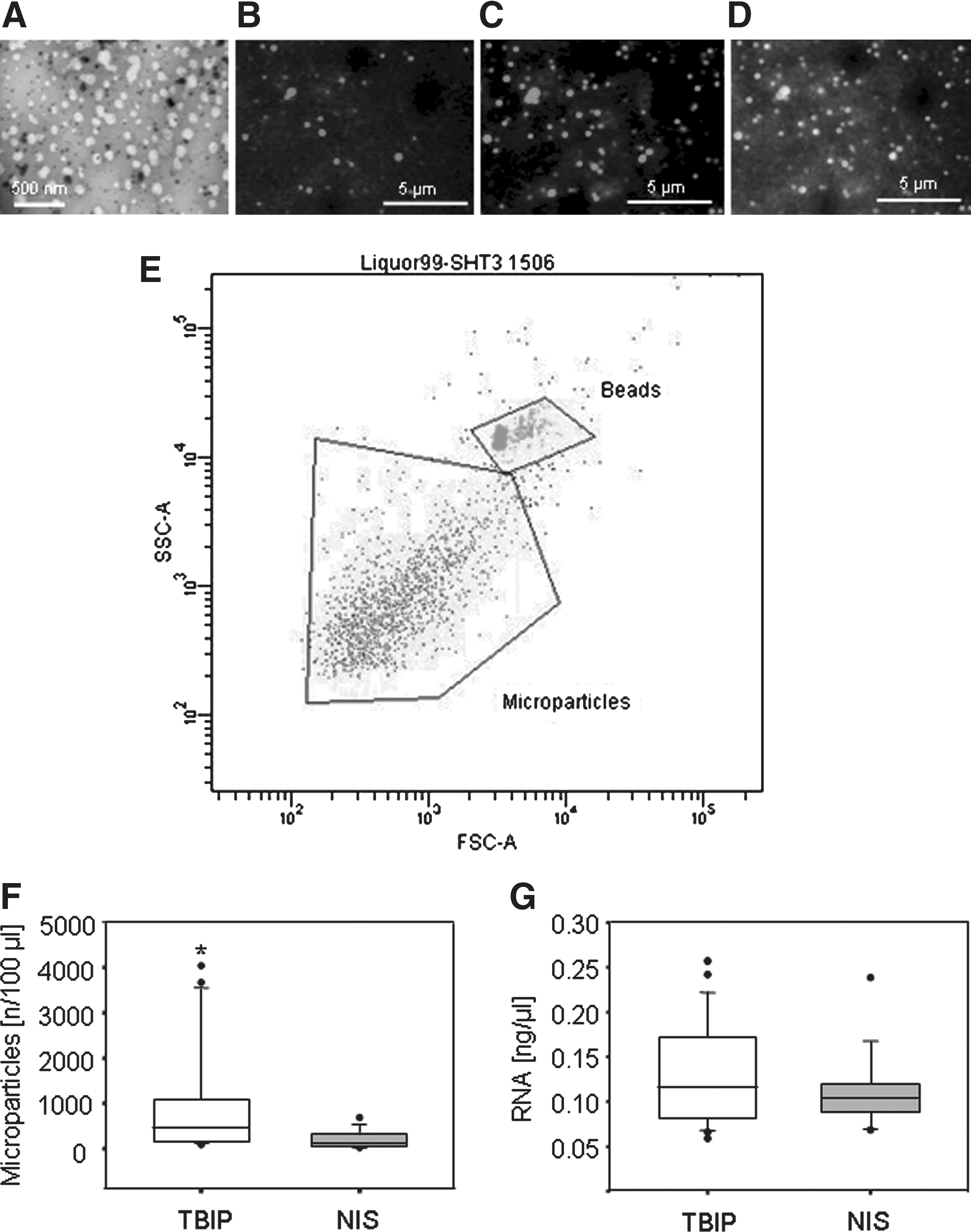
Detection and quantification of microparticles derived from cerebrospinal fluid (CSF-MPs). (
The particles were quantified by flow cytometry and shown to be significantly more abundant in TBIPs than in NIS (Fig. 1E,F). They were also more abundant in CCDs, suggesting an association between CNS damage or disease and elevated numbers of CSF-MPs (see online supplementary Fig. S1 at
CSF-MPs are enriched in mature mRNAs and proteins that are associated with neuronal RNA-granules
There is evidence that MPs carry ribonucleoproteins, which are associated with the transport of RNA to distal cellular sites, such as dendrites, to enable localized translation.
23
In accordance with this observation, we detected typical neuronal granule proteins such as MAP2, ARC, and LIMK1 mRNA, as well as ß-ACTIN in CSF-MPs from both patients and controls (Fig. 2). A semi-quantitative comparison of the relative levels of distinct mRNAs in single samples that would allow a correlation of RNA levels with disease progression was disregarded, because validated normalization candidates were not available for CSF-MPs derived mRNA. An attempt to evaluate the average level of these mRNAs in CSF-MPs derived from patient or NIS samples was implemented by using external signals derived from NT-2 cell cDNA amplification for normalization (see online supplementary Fig. S4A–D at
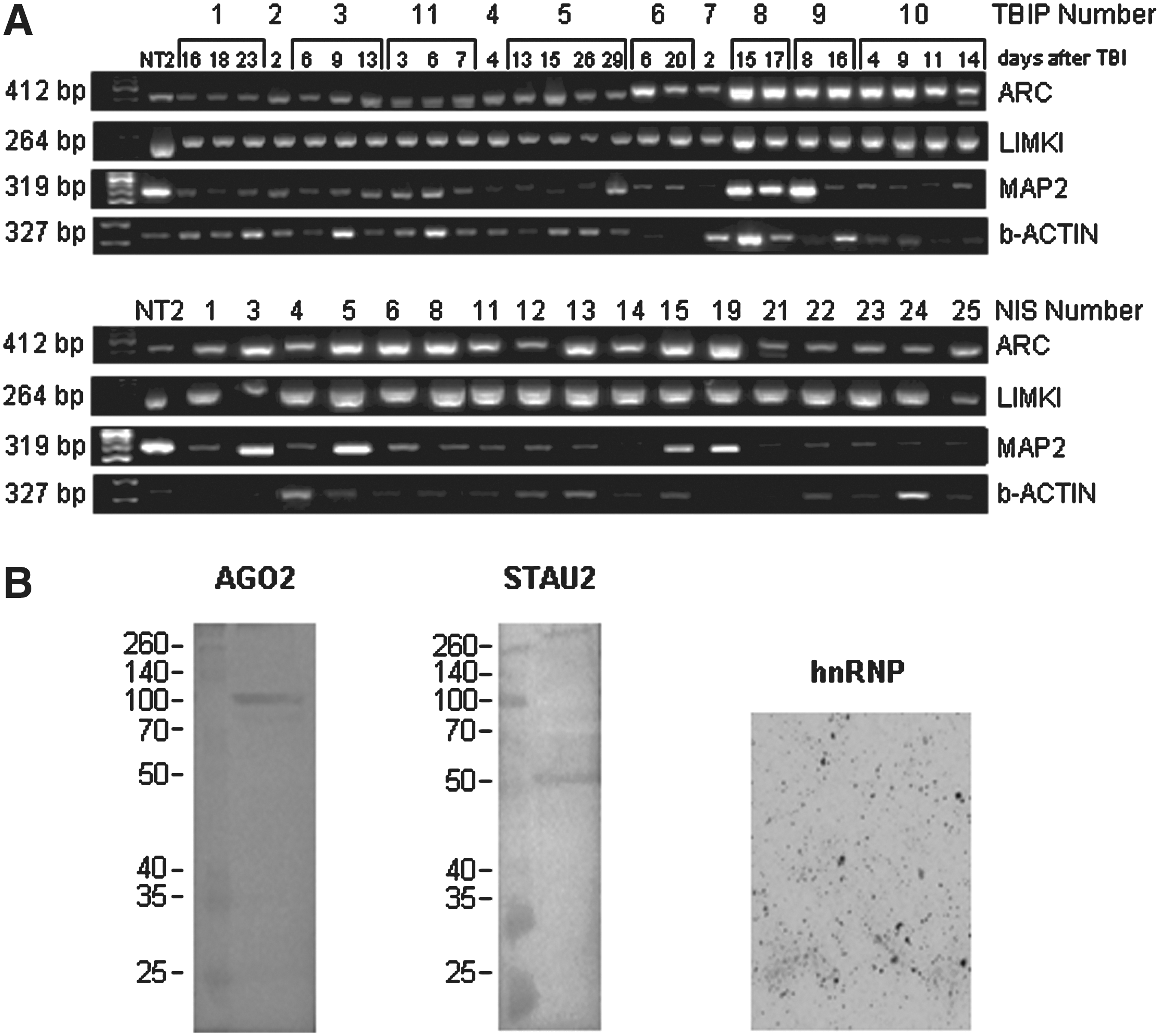
Detection of mRNAs shuttled by cereb rospinal fluid microparticles (CSF-MPs). (
We also verified the presence of RNA granule-associated proteins in CSF-MPs derived from TBIP. AGO2 and STAU2 were detected by Western blotting of CSF-MPs proteins isolated from pooled CSF (Fig. 2B). This pool was also used to visualize hnRNP in CSF-MPs by immunocytochemistry.
The very small volumes of CSF available precluded a direct comparison of the protein levels in CSF-MPs from NIS by Western blotting.
Profiling of CSF-MP microRNAs
A bioanalyzer profile of CSF-MP RNA revealed the presence of a broad range of RNA sizes including a prominent peak of small RNA species (51% of total RNA) (Fig. 3A) consistent with an enrichment in miRNAs. Based on this result, we hybridized RNA from pooled CSF-MPs from 10 brain-injured patients with Affymetrix GeneChip miRNA arrays. Eighty-one distinct miRNAs were identified (see online supplementary Table S2 at
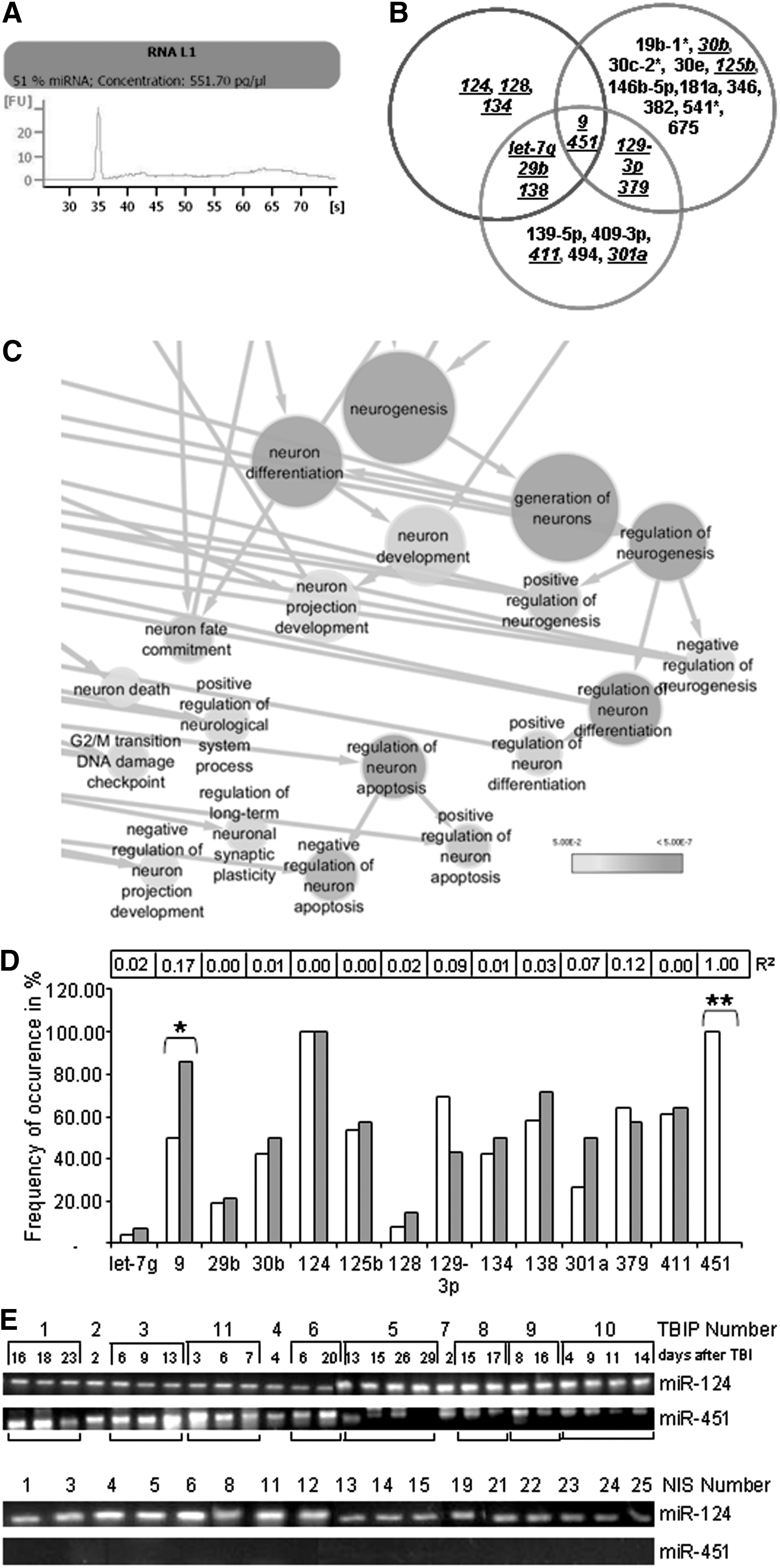
Profiling of microRNAs derived from cerebrospinal fluid microparticles (CSF-MPs). (
A total of 1659 target genes were linked to the identified miRNAs using miRWalk database (see online supplementary Table S3 at
CSF derived MPs from TBIP and healthy subjects contain a distinct pattern of microRNAs
The presence of 14 distinct miRNA species in CSF-MPs derived from individual TBIPs and NIS was verified by qRT-PCR (Fig. 3B, underlined). A miRNA was deemed to be present in the CSF-MPs of a given patient or NIS when the intensity of the respective miRNA RT-PCR signal was reproducibly 20% higher than the background signal. The frequency of occurrence of distinct miRNAs was determined by logistic regression analysis (Fig. 3D-E). Using this approach, we verified the presence of miR-124 in all samples (R2: 0.00). miR-451 in contrast was only detected in CSF-MPs from TBIPs (R2: 1, p=0.00) and could therefore play a regulatory role associated with TBI. The presence of miR-9 on the other hand was shown to be more prevalent in CSF-MPs derived from NIS (R2: 0.172, p≤0.025). A statistically significant or predictive frequency of occurrence was not observed for any of the other miRNA species analyzed.
Functional transfer of CSF-MP components to undifferentiated NTERAs
The ability of CSF-MPs to deliver their constituents to target cells was investigated by incubating membrane-specific biotinylated, RiboGreen-stained MPs with undifferentiated NTERAs (NT-2), a neuronally committed human teratocarcinoma cell line. 26 Undifferentiated NTERAs express nestin and vimentin, intermediate filament (IF) proteins expressed in neuroepithelial precursor cells, as well as MAP1b, expressed in human neuroepithelium. 27 Uptake of RiboGreen stained RNA was shown by fluorescence microscopy (Fig. 4A–E). The non-superposition of green fluorescent RNA and red fluorescent membranes might indicate that RNA is released from MPs during uptake (Fig. 4E).
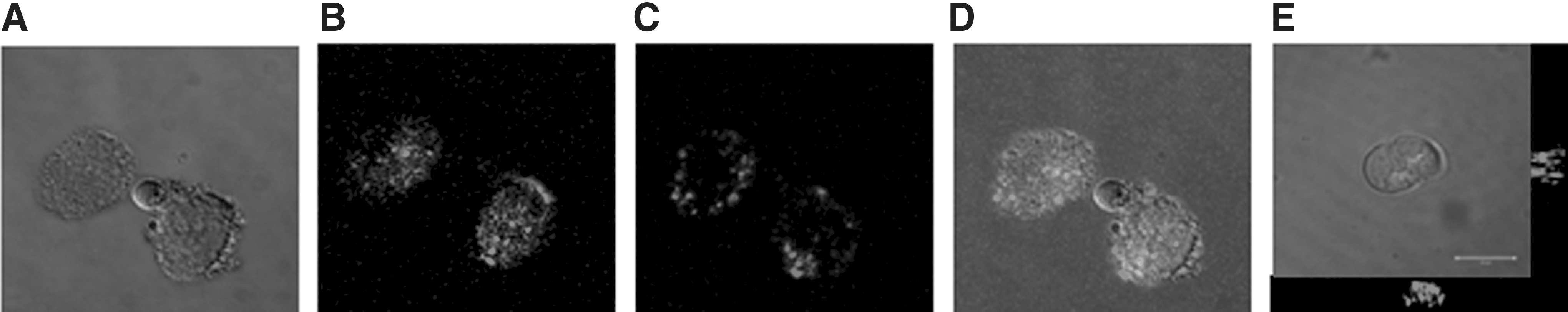
Transfer of CSF-MP content to undifferentiated NTERAs. (
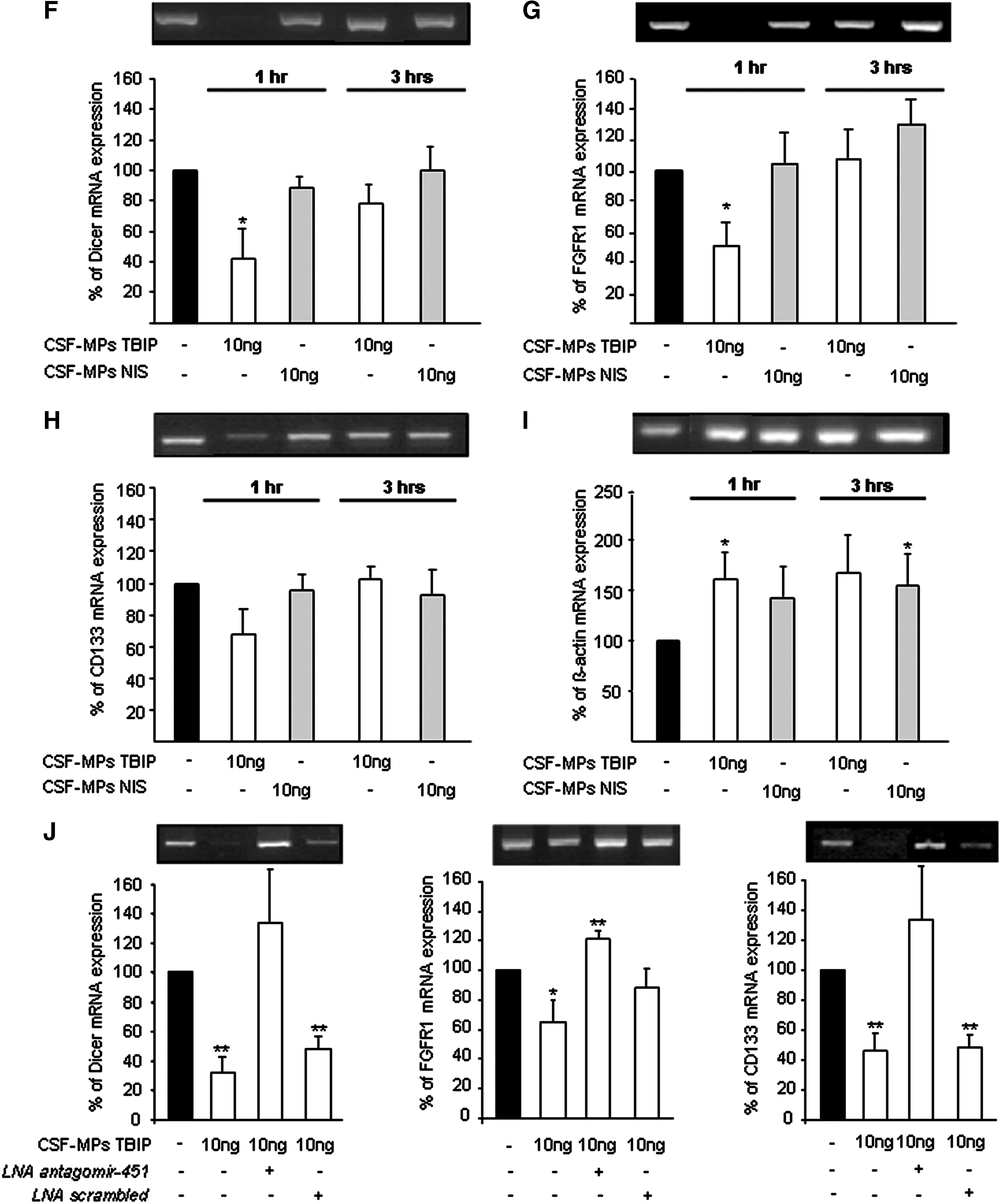
On addition of CSF-MPs derived from TBIPs to NTERAs, a rapid (1 h) concentration and time-dependent miR-451 mediated decrease of Dicer, FGFR1, and CD133 mRNA expression was observed (Fig. 4F–H). These genes are listed as putative miR-451 target genes in the miRWalk database. The decrease in putative target gene expression was accompanied by an increase in miR-451 in these cells (see online supplementary Fig. S1B at
miR-451 was inhibited by a LNA miR-451 inhibitor but not by LNA scrambled when added to the cells before incubation of NTERAs with CSF-MPs derived from TBIPs (Fig. 4J). These results demonstrate the down-regulation of Dicer, FGFR1, and CD133 mRNA to be specifically mediated by miR-451.
Discussion
In this study, we show for the first time that human CSF contains membrane-sheathed MPs that carry genetic information and proteins. We demonstrate that the genetic information comprises mRNAs associated with RNA granules and miRNAs implicated in the regulation of neuronal processes. CSF-MPs are more abundant in subjects with brain injury and shuttle a distinct set of miRNAs, including miR-9 and miR-451, both of which have previously been shown to be regulated in cerebral tissue after experimental traumatic brain injury. 24,25 We confirm a transfer of genetic material from CSF-MPs to cultured NTERAs and a subsequent decrease in putative miR-451 target gene expression, suggesting CSF to harbor a transcellular delivery system that contributes to the signaling between cells.
Experimental design
Analyzing CSF samples derived from TBIP is to some extent restricted by a limiting study design. A primary impediment is the absence of true controls; i.e., CSF of healthy subjects. Observed differences between CSF samples derived from TBIP and NIS should therefore be interpreted with care. Further, samples are taken from different locations (ventricle/subarachnoid space lumbar region). To exclude a potential rostro-caudal gradient of CSF-MPs and the associated content, a study analyzing MPs and RNA content in lumbar versus ventricular CSF samples in the same patient population is necessary. 28 –33
MPs in CSF
We confirmed the presence of CSF- MPs in human CSF samples by electron and fluorescent microscopy. Consistent with descriptions in body fluids and in cell cultures, CSF-MPs were highly heterogeneous with sizes ranging from 50–400 nm, which suggests that they are generated by a combination of membrane budding and exocytosis (MPs in the lower size range). 13,14,17,34,35
Whereas the range of CSF-MP sizes was comparable in patients and NIS, the abundance of CSF-MPs was clearly elevated in the former group (see above), consistent with reports in other non-CNS disease states, 7,8 albeit the numbers detected were considerably below those reported in a previous CNS study. The discrepancy, however, most likely reflects the use of different isolation protocols; i.e., sample filtration through a 0.45 micron mesh after high speed centrifugation might result in significantly reduced numbers of particles. 17,36
RNA carried by MPs derived from CSF
The amount of RNA per MP sample varied considerably between samples. The composition and function of MPs are dependent on their cellular origin, the agonist responsible for MP formation, and the microenvironment of the parental cell. 37,38 The heterogeneity of the MP population in CSF samples might be the reason why a correlation between RNA levels and the health status of the donor was not detected.
The variable total RNA content of CSF-MPs comprises ß-ACTIN, MAP2, LIMK1, and ARC mRNA as well as Staufen-2 and Argonaute-2 protein, molecules that have been associated with the regulated intercellular transport of RNA. 39,40 Our findings thereby confirm the results of a previous study that showed circulating MPs and MPs derived from cultured stem cells to carry RNA granule associated ribonucleoproteins. 23,41 Collino and associates 23 have suggested a role of ribonucleoproteins in RNA transport and stability in MPs derived from cultured mesenchymal stem cells. A similar role of ribonucleoproteins and respective mRNAs has still to be established in CSF-MPs.
microRNAs shuttled by MPs
microRNAs constitute a high percentage of ribonucleic acids shuttled by CSF-MPs. We determined a predictive value for the presence of miR-451 in CSF-MPs of brain injured patients. miR-9 was detected with a higher frequency of occurrence in CSF-MP samples derived from NIS than from TBIPs. Intriguingly, miR-9 and miR-451 were the only two miRNA species independently identified in rat brains after TBI. 24,25 Also, other disease-dependent released MPs differ significantly in their miRNA content profile, as shown by Diehl and colleagues. 42
miR-9 is specifically expressed in the mammalian nervous system and has been implicated in the regulation of a variety of neuronal processes such as neuron development 43 or axis formation. 44 The prevalence of miR-9 in CSF-MPs from NIS might indicate a role in the homeostasis of neuronal regulation.
miR-451 was detected in patient CSF-MPs only. A potential role of miR-451 as a damage associated regulator of gene expression was also verified by in vitro studies. In undifferentiated adult neuronal stem cells, expression of putative miR-451 target genes Dicer, FGFR1, and CD133 was silenced within 30 min of scratch induced cell damage (see online supplementary Fig. S5 at
Taking into consideration the role of miR-451 in erythropoiesis, however, it should be noted that the prevalence of miR-451 in CSF-MPs of brain injured patients might indicate the presence of vascular MPs in the CSF. As previously discussed, transfer of MPs from the nervous to the cardiovascular system and in this case vice versa might constitute an additional novel transborder communication channel. 18
The emerging role of the identified miRNAs in cerebral function was further affirmed by GO analysis. The result suggests 34 of 37 neuron related cellular processes to be overrepresented in target genes regulated by the distinct set of miRNAs (see online supplementary Table S4 at
Transfer of miRNA from MPs to cultured cells
We confirmed the ability of CSF-MPs to transfer their “RNA cargo” to target cells in vitro. Attachment of plasma derived MPs, and the transfer of their miRNA to human umbilical vein endothelial cells in vitro has been previously demonstrated. 42 Our observations are also consistent with results showing miRNA transfer from stem cell MPs to fibroblasts, 34 from mesenchymal stem cells to tubular epithelial cells or tumor cell lines, 23,45 and from embryonic stem cell microvesicles to mouse embryonic fibroblasts. 46
The uptake of CSF-MP content by cultured NTERAs was paralleled by the regulation of gene expression. The decrease of putative miR-451 target genes in NTERAs incubated with CSF-MPs from brain injured patients demonstrates a functional transfer of messages and a role of this miRNA in damage induced cerebral regulation. A MP mediated reprogramming of target cells has been shown for cell culture derived MPs. 5,47 –50
Conclusion
The results of the present study provide strong evidence for the shuttling and cell-to-cell transfer of brain injury associated miRNA and mRNA by CSF-MPs. The transport of genetic information in CSF and subsequent reprograming of target cells signifies a communication network that promotes signal transduction between adjacent and distal cells in the ventricular system. The uncovering of the extracellular transport of signals in the CSF will facilitate a more detailed analysis of the regulation of cerebral function and might thereby support the identification of novel diagnostic markers or even therapeutic strategies for damage associated or neurodegenerative cerebral modulations.
Footnotes
Acknowledgments
We thank Chris Wrighton for editing the article and helpful comments. This work was supported by a grant from the Styria government.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.