
Abstract
The majority of people who sustain a traumatic brain injury (TBI) have an injury that can be classified as mild (often referred to as concussion). Although head CT scans for most subjects who have sustained a mild TBI (mTBI) are negative, these persons may still suffer from neurocognitive and neurobehavioral deficits. In order to expedite pre-clinical research and develop therapies, there is a need for well-characterized animal models of mTBI that reflect the neurological, neurocognitive, and pathological changes seen in human patients. In the present study, we examined the motor, cognitive, and histopathological changes resulting from 1.0 and 1.5 atmosphere (atm) overpressure fluid percussion injury (FPI). Both 1.0 and 1.5 atm FPI injury caused transient suppression of acute neurological functions, but did not result in visible brain contusion. Animals injured with 1.0 atm FPI did not show significant motor, vestibulomotor, or learning and memory deficits. In contrast, 1.5 atm injury caused transient motor disturbances, and resulted in a significant impairment of spatial learning and short-term memory. In addition, 1.5 atm FPI caused a marked reduction in cerebral perfusion at the site of injury that lasted for several hours. Consistent with previous studies, 1.5 atm FPI did not cause visible neuronal loss in the hippocampus or in the neocortex. However, a robust inflammatory response (as indicated by enhanced GFAP and Iba1 immunoreactivity) in the corpus callosum and the thalamus was observed. Examination of fractional anisotropy color maps after diffusion tensor imaging (DTI) revealed a significant decrease of FA values in the cingulum, an area found to have increased silver impregnation, suggesting axonal injury. Increased silver impregnation was also observed in the corpus callosum, and internal and external capsules. These findings are consistent with the deficits and pathologies associated with mild TBI in humans, and support the use of mild FPI as a model to evaluate putative therapeutic options.
Introduction
T
Three rodent models: fluid percussion injury (FPI), controlled cortical impact injury, and weight drop, have been used to examine the behavioral and pathological changes associated with TBI. Although moderate-to-severe forms of injury have been extensively studied using these models, there have been relatively fewer investigations into the consequences of mild TBI. 8 –11 In humans, mild TBI is generally defined as an opening Glasgow Coma Score of 13–15, transient loss of memory for events immediately before or after the injury, loss or alterations of consciousness, and alterations in mental status at the time of injury. However, within this definition exists a range of injuries that can be classified as either uncomplicated or complicated, depending on neuroimaging abnormalities. Although most mTBI patients do not have gross structural changes that can be detected by head computerized tomography (CT), few animal studies have been carried out using magnitudes of TBI that do not result in brain contusion. 12 –16 For example, Shultz et al. 16 reported that 1.2 atm FPI did not cause overt brain damage but resulted in significant short-term memory impairments when tested 24 h after injury. Employing a 1.5 atm FPI injury, Wu et al. observed that animals pre-trained in the Morris water maze task had poor memory recall when tested over the first 3 days after injury. 17 As experimental treatments often need to be administered over a period of days after TBI, it is desirable that models give rise to cognitive deficits that persist beyond the acute stage of injury. With this in mind, we aimed to identify the magnitudes of mild FPI that can produce a significant cognitive deficit on day 5 post-injury that occurs in the absence of visible injury.
In this study, we examined the vestibulomotor, motor, and cognitive functions in rats injured using either 1.0 atm or 1.5 atm FPI. After the completion of behavioral testing, we performed ex vivo diffusion tensor imaging (DTI) to assess axonal integrity, and histopathological analysis to examine inflammation, neuronal loss, and morphological changes. Our results show that while a 1.0 atm injury did not cause a significant neurocognitive deficit, 1.5 atm injury caused a reproducible learning and short-term memory impairment that occurred in absence of visible contusion or neuronal loss, but that was associated with axonal damage and neuroinflammation. Our results indicate this level of injury would be suitable for pre-clinical drug screening studies to improve the outcome in this subset of mild TBI patients.
Methods
Materials
Male Sprague-Dawley rats (275–300 g) were purchased from Charles River Laboratories (Wilmington, MA). Antibodies to NeuN and GFAP (Millipore, Billerica, MA), IBA-1 (WAKO, Richmond, VA), amyloid precursor protein (APP, Invitrogen, Grand Island, NY), and myelin basic protein (Covance, Princeton, NJ) were obtained for use in these studies. A silver staining kit to identify degenerating neurons was purchased from FD Neurotechnologies (Columbia, MD).
Lateral fluid percussion injury
All experimental procedures were approved by the Institutional Animal Care and Use Committee and were conducted in accordance with the recommendations provided in the Guide for the Care and Use of Laboratory Animals. Protocols were designed to minimize pain and discomfort during the injury procedure and recovery. Fluid percussion injury was carried out as described previously. 18 –20 Briefly, rats were initially anesthetized using 5% isoflurane with a 1:1 N2O/O2 mixture and then maintained with a 2.5% isoflurane with 1:1 N2O/O2 mixture via a face mask. Animals were mounted on the stereotaxic frame, a midline 4.8 mm-diameter craniectomy was carefully made midway between bregma and lambda. A hub (modified from 20-gauge needle) was planted into the burr hole and affixed to the skull by contact adhesive and dental cement. Once the assembly was secured, the rat was removed from anesthesia and allowed to regain its tail pinch reflex. Immediately upon regaining this reflex, the rat was injured using a FPI device and a pressure of 1.0 or 1.5 atmosphere (atm) over base room pressure. After injury, the hub and surrounding dental cement were immediately removed, and the incision closed by wound clips. Sham-operated animals received all the aforementioned surgical procedures except hub implantation and the injury. Animals' body temperature was maintained at 37°C during the surgery, using a rectal thermometer coupled to a heating pad.
Assessment of acute neurological function
After injury, the duration of suppression of the paw withdrawal reflex and the self-righting reflex were monitored as acute measures of injury severity. Self-righting was monitored by placing the animal on its back and recording the time from injury required to right itself three consecutive times. For sham-operated animals, the duration of suppression of righting response was measured from the time of reinstatement of the paw pinch reflex (the point at which the FPI was delivered in injured animals).
Measurement of cerebral perfusion
Cerebral perfusion was measured using a PIM3 scanning laser Doppler device. Due to the prolonged anesthesia required for these measures, perfusion measures were carried out in animals separate than those used for behavior. Following the creation of the burr hole, but prior to hub implantation, the cerebral cortex was scanned through the burr hole to establish pre-injury perfusion values. After injury, the animal was placed back into the stereotaxic frame and maintenance of anesthesia reinstated. The hub was gently removed and cerebral perfusion monitored every 10 min for the first hour after injury. Additional scans were performed 24 h after injury to determine if perfusion had normalized by this time point. Sham-operated animals were treated identically with the exception of the injury. Five consecutive scans were generated for each time point, the perfusion values of which were average for each animal.
Behavioral assessments
All behavioral tests were conducted by an experimenter blind to the treatment groups.
Vestibulomotor and motor functions
Three different motor skill tasks (beam balance, rotarod, and foot-fault) were used to determine animals' vestibulomotor and motor performances as described previously. 21,22 Beginning on day 1 post-injury, animals were given three trials in the beam balance task during which the length of time spent balancing on the beam was recorded. For the foot fault task, the number of foot faults was evaluated by placing the animal on a wire grid (opening size of 2×2 cm) and counting the number of times a front paw missed and slipped below the plane of the grid out of a total of 50 steps. The rotarod procedure used was similar to that described by Hamm et al. 22 The animal was placed upon the stationary rotarod device for a period of 10 sec, after which rotation was initiated (beginning speed of 3 rpm that was incrementally increased by 3 rpm every 10 sec until a maximum of 30 rpm was reached). The time spent on the device (maximum time of 20 sec at 30 rpm) was recorded in each of the two daily trials. A trial ended, and time was recorded, when the animal either fell off the device or held onto the rotating rod for two consecutive rotations without attempting to walk.
Spatial learning and short-term memory
A common component of neuropsychological tests used to diagnose mTBI (e.g., military acute concussion evaluation, MACE; automated neuropsychological assessment metrics, ANAM) is a short-term memory test. In order to assess short-term memory, we used a modified version of the Morris water maze task in which rats are trained to find the location of a hidden platform within a single training session, followed by a probe trial administered 30 min later. 23,24 On day 5 post-injury, rats were given 10 consecutive training trials with an inter-trial interval (iti) of 4 min. Each trial was initiated by placing the rat into the water maze at one of four randomly chosen starting positions. The animal was allowed to search for the hidden platform for a period of 60 sec and the time to find the platform recorded. If the rat failed to find the hidden platform on any given trial, it was led there by the experimenter. Thirty min after the last training trial, animals were tested in a probe trial in which the platform was removed from the tank and the rat allowed to search for a period of 60 sec. Movement within the maze was monitored using a video camera linked to tracking software (Ethovision, Noldus Information Technology, Leesbury, VA, USA). Measures of memory including latency to first platform crossing, number of crossings, and quadrant preference were recorded.
Ex vivo diffusion tensor imaging (DTI)
At the conclusion of behavior testing, rats were given an overdose of sodium pentobarbital (100 mg/kg), transcardially perfused with ice-cold phosphate-buffered-saline (0.1 M PBS), followed by buffered 4% paraformaldehyde. Brains were carefully removed and stored in fixative at 4°C. Representative brains from each group were then submerged in Fomblin (Kurt J. Lesker Company, Livermore, CA) and images were acquired at the UTHealth MRI Core Facility using a Bruker Biospec 70/30 URS scanner, operating at field strength of 7 Tesla. A series of 3D EPI DTI scans (42 gradient directions) were obtained in axial orientation. These images were then imported into DTI Studio software to calculate maps of fractional anisotropy (FA), mean (MD), longitudinal (LD), and radial (RD) diffusivities. Regions of interest (ROIs) encompassing the genu of the corpus callosum, cingulum, internal and external capsules, fimbria, and cortex proximal to the injury site, were outlined in the ipsilateral hemisphere, and values compared between sham and FPI groups. Statistical comparisons were made using a two-way ANOVA, followed by a Bonferroni post-hoc analysis. Fiber tracking was carried out using MedINRIA 1.9 (
Immunohistochemistry
Brains not used for DTI scanning were transferred to a 30% buffered sucrose solution for cryopreservation. Brains were sectioned on a cryostat in the coronal plane at 40 μm through the rostro-caudal extent. Sections were incubated in primary antibody solutions (0.1-0.5 μg/mL antibody, 2.5% normal goat serum in PBS) overnight at room temperature. Following extensive washing, sections were incubated for 1 h in PBS containing species-specific secondary antibodies linked to AlexaFluor dyes (Alexa488 or Alexa568; Invitrogen). Sections were mounted onto glass slides and coverslipped with Fluoromount-G to retard fading. Slides were examined using an upright microscope with epifluorescence capabilities. Images were captured using a MagnaFire camera using settings that remained constant across groups.
Silver staining
Silver staining was carried out on free-floating sections using a kit from FD Neurotechnologies (Columbia, MD) essentially as described by the vendor. Of exception was the impregnation time was extended from 4 min to 6 min in order to maximize the signal-to-noise ratio.
Statistical analyses
Statistical comparisons were carried out using SigmaStat (Systat Software, San Jose, CA). Across group comparisons of data collected over time (e.g., behavioral training and blood flow measures) were evaluated using a repeated measures two-way ANOVA, followed by post-hoc analysis. Group main, or interactions of group and time, differences were used to compare the groups. Single measure data (e.g., probe trial data) was statistically compared using a Student's t-test for unpaired variables, whereas within group comparisons (e.g., quadrant preference) were tested using a repeated measures one-way ANOVA.
Results
1.0 atm FPI did not cause vestibulomotor, motor, or spatial learning, and memory deficits
The majority of human mTBI patients are head CT negative, indicating no overt physical damage to the brain, yet many will suffer from neurological, cognitive, and behavioral dysfunctions <

FPI of 1.0 atm did not cause learning or memory dysfunction when tested 5 days post-injury.
In order to determine the effect of 1.0 atm mFPI on vestibulomotor and motor functions, animals were tested on the balance beam, rotarod, and foot fault tasks, beginning on day 1 post-injury. No significant differences between the two groups was detected in either vestibulomotor (balance beam (day 1): sham, 56.30±2.01 sec; 1.0 atm FPI, 56.39±1.95 sec, Student's t-test: p=0.975), locomotor (rotarod (day 1): sham, 66.36±4.30 sec; 1.0 atm FPI, 73.33±7.63 sec, Student's t-test: p=0.401), or motor coordination (foot fault (days 1–3): ipsilateral two-way repeated measures ANOVA: F(2,66)=0.468, p=0.628; contralateral two-way repeated measures ANOVA: F(2,66)=0.311, p=0.734).
When learning was assessed using the abbreviated water maze task (on day 5 post-injury), a trend towards longer latencies to locate the hidden platform was observed in the 1.0 atm FPI animals compared to the sham controls (repeated measures two-way ANOVA: F(1,35)=3.51; p=0.069) (Fig. 1B). When short-term memory was assessed 30 min after the completion of training, there was no difference in latency to the previous location of the platform (Students t-test, p=0.42; Fig. 1C), nor in number of platform crossings (Student's t-test, p=0.56, Fig. 1D). Both sham (one-way repeated measures ANOVA: F(3,48)=7.85, p<0.001) and 1.0 atm FPI (one-way repeated measures ANOVA: F(3,57)=12.17, p<0.001) groups displayed significant quadrant preferences for the target quadrant (data not shown). As significant short-term memory impairments were not observed at 1 atm FPI, further characterization of this injury magnitude was not carried out.
1.5 atm FPI causes transient motor dysfunction
To determine if a higher injury magnitude could be used to mimic the cognitive deficits associated with uncomplicated mild TBI, groups of animals were injured using 1.5 atm FPI (n=11) or sham-operated (n=11) and the resulting pathobiologies compared. We observed a significantly longer suppression of the righting response in 1.5 atm injured rats as compared to sham animals (sham: 1.53±0.2 min; 1.5 atm mFPI: 7.35±0.3 min, Student's t-test: p<0.001). 1.5 atm animals also had significantly longer suppression of paw withdrawal reflexes (1.0 atm mFPI, 43.67±1.48 sec; 1.5 atm mFPI 90.73±7.28 sec, Student's t-test: p<0.001), further supporting increased injury. Similar to 1.0 atm FPI, 1.5 atm FPI did not cause overt damage to the brain (Fig. 2A). When tested for vestibulomotor and locomotor function, 1.5 atm FPI animals were not significantly impaired in their ability to perform the beam balance (sham: 56.28±2.08 sec; mFPI: 54.91±3.21 sec, Student's t-test: p=0.933; Fig. 2B) or the rotarod (sham: 66.36±4.30 sec; mFPI: 71.58±6.89 sec, Student's t-test: p=0.527; Fig. 2C) tasks. Transient deficits in motor coordination were observed, with 1.5 atm FPI animals making significantly more ipsilateral (two-way repeated measures ANOVA: F(2,39)=4.06, p=0.025) and contralateral (two-way repeated measures ANOVA: F(2,39)=8.56, p<0.001) foot-faults than did their sham-operated counterparts on days 1 and 2 post-injury (Fig. 2D). By day 3, the performance of the injured animals did not differ from the sham group.
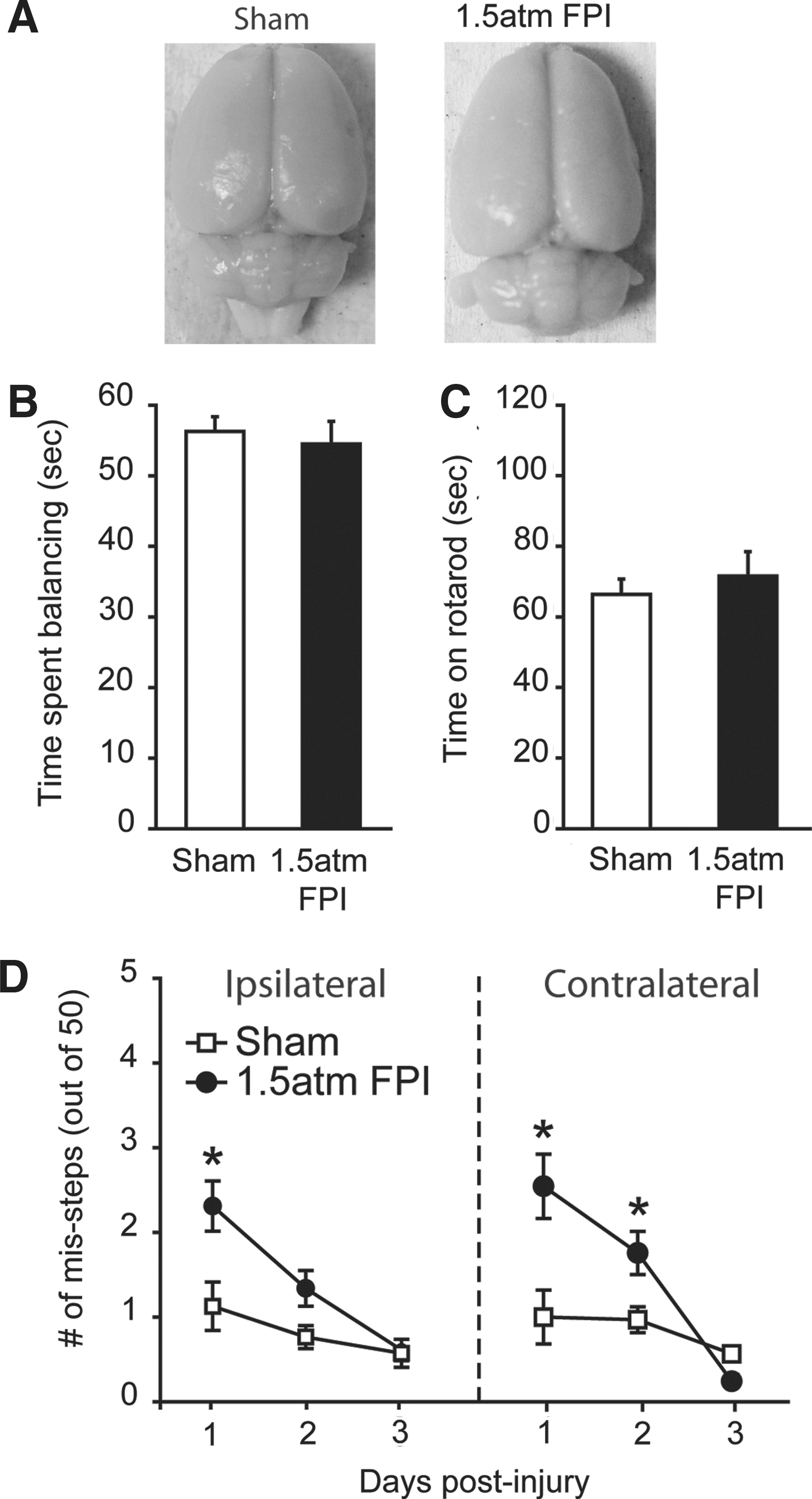
FPI of 1.5 atm caused transient motor coordination deficits.
1.5 atm FPI caused spatial learning and short-term memory dysfunction
To test if 1.5 atm FPI causes learning and memory deficits, sham and injured animals were trained and tested in the abbreviated version of MWM task on day 5 post-injury. During training, there was a significant difference in the latency to find the hidden platform between the two groups (two-way repeated measures ANOVA: F(1,20)=25.25, p<0.001; Fig. 3A). The representative traces shown in Figure 3B indicate that, although sham-operated animals initially swam along the perimeter of the tank looking for a means of escape, by the end of training, they had learned the location of the hidden platform and took a direct path to its location. In contrast, 1.5 atm animals appeared to continue to spend more time in the periphery of the tank and had a less focused search pattern. Quantification of the swim paths revealed that over the course of training, 1.5 atm FPI injured animals entered (F(1,20)=22.84, p<0.001) and spent more time (F(1,20)=21.55, p<0.001; Fig. 3C) in the periphery of the tank. Further, 1.5 atm FPI rats spent significantly more time in the opposite half of the tank throughout training (F(1,20)=9.71, p=0.005; Fig. 3D). No significant differences in number of right or left turns were found between the groups, indicating no inherent circling bias.

FPI of 1.5 atm caused significant spatial learning deficits.
Short-term memory was assessed by a probe trial administered 30 min after the completion of training. Figure 4A shows representative traces from a sham and a 1.5 atm FPI rat, indicating the paths taken to first platform crossing and during the remainder of the 60 sec probe. The traces show that the sham-operated controls had a more localized search pattern, requiring less time to cross the platform location and resulting in more platform crossings than that observed in the trace from the 1.5 atm FPI animal. Consistent with this, latency (Student's t-test: p=0.016; Fig. 4B) and path length (Student's t-test: p=0.011; Fig. 4C) to first platform crossing were found to be significantly different between the sham and FPI groups. Throughout the 60 sec probe trial, 1.5 atm FPI animals crossed the platform fewer times than did sham controls (Student's t-test: p=0.025), indicating a less focused search pattern. Further, sham-operated controls spent more time searching in the quadrant in which the platform was located (one-way repeated measures ANOVA: F(3,30)=7.46, p<0.001), whereas the 1.5 atm FPI animals spent an equivalent amount of time exploring three of the four quadrants (one-way repeated measures ANOVA: F(3,30)=2.70, p=0.063; Fig. 4D). No differences in swim speed (sham, 26.91±0.88 cm/sec; 1.5 atm FPI, 28.04±1.10 cm/sec; Student's t-test: p=0.435) or latency to a visible platform (sham, 5.79±0.82 sec; 1.5 atm FPI, 8.77±1.54 sec; Student's t-test: p=0.103) was observed between the two groups.
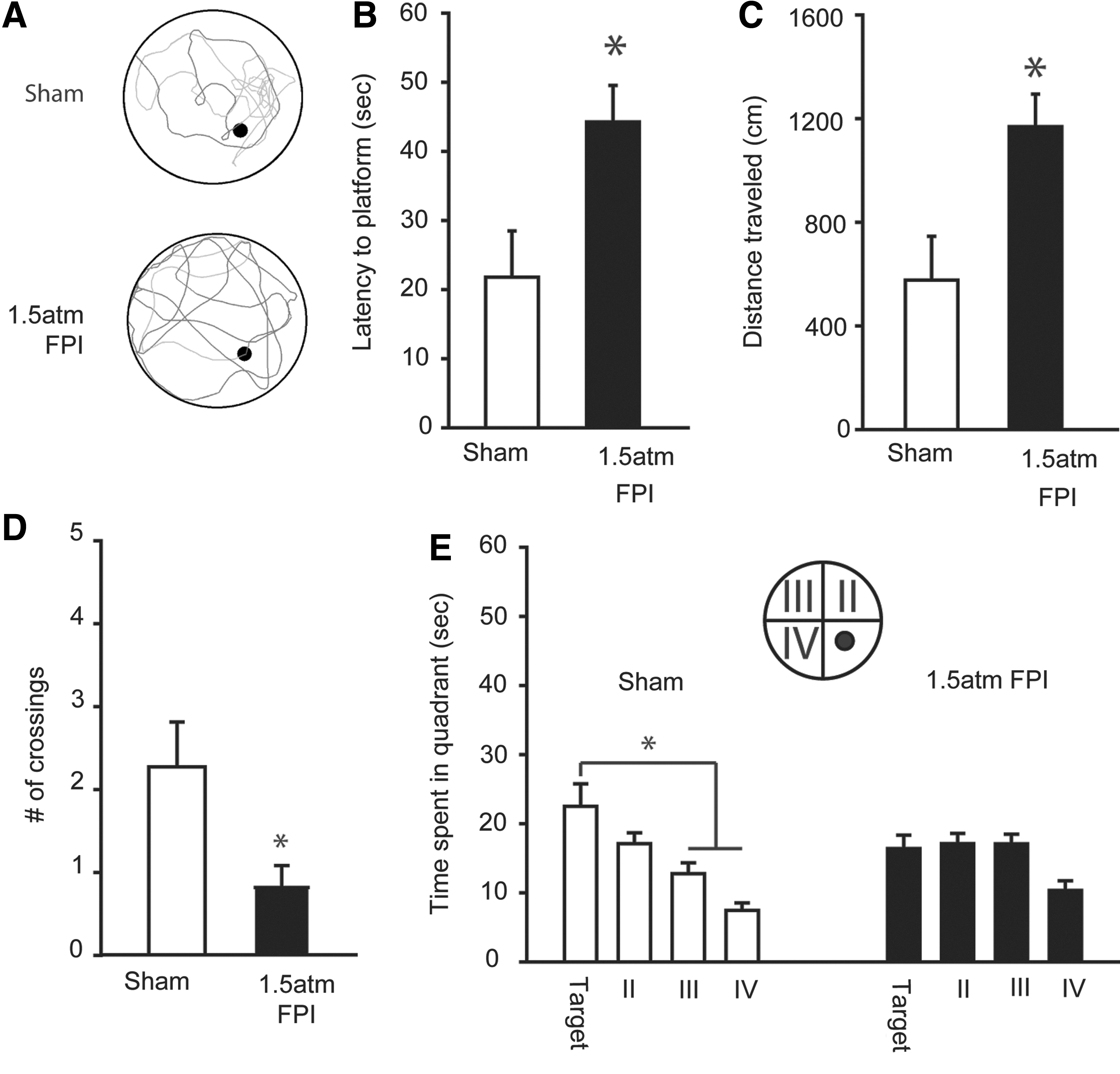
FPI of 1.5 atm caused significant short-term memory deficits.
1.5 atm FPI transiently altered cerebral perfusion
To examine the consequences of 1.5 atm FPI on cerebral perfusion, a Periscan PIM3 Laser Doppler Blood Perfusion Imager was used as a noninvasive means of monitoring microvasculature blood perfusion. Due to the low intensity of the laser and its inability to penetrate the skull, imaging was done through the craniotomy prepared for hub implantation. Sham animals received the crainiectomy in order to allow cerebral imaging to be performed. Scans were performed prior to injury (baseline), 3 min post-injury and every 10 min thereafter for the first hour after injury. Figure 5A shows representative color-coded images from a sham and a 1.5 atm FPI animal. An apparent decrease in cerebral blood perfusion can be observed 13 min after injury that returns to baseline by 24 h post-injury. The summary data in Figure 5B show that, compared to sham-operated controls (n=4), cerebral perfusion significantly decreases in the 1.5 atm FPI animals (n=6) (two-way repeated measures ANOVA: F(8,57)=14.337, p<0.001). The decrease could be detected as early as 3 min after injury, and persisted throughout the 1 h monitoring period. When examined the following day, cerebral perfusion had returned to baseline values.

FPI of 1.5 atm caused transient disturbances in cerebral perfusion. Cerebral perfusion at the site of injury was monitored using a Perimed PIM3 Laser Doppler Blood Perfusion Imager.
1.5 atm FPI did not cause overt cell loss or dendritic damage
Patients with uncomplicated mild TBI are typically negative for findings on computerized tomography (CT) or MRI scans. To examine if histopathological changes associated with 1.5 atm FPI can be detected using imaging, the animals used for behavioral studies were transcardially perfused on day 15 post-injury with 4% paraformaldehyde and brains removed. Ex vivo Rapid Acquisition with Refocused Echoes (RARE) scans throughout the rostro-caudal extent of the cerebral cortex were obtained from sham and 1.5 atm FPI animals. No overt damage to the cerebral cortex, hemorrhage, or midline shifts was observed in 1.5 atm FPI rats compared to sham-operated controls (Fig. 6A). Examination of NeuN immunostained tissue confirmed that neither 1.0 atm nor 1.5 atm FPI caused changes in cortical or hippocampal neuron density or distribution (Fig. 6B). Likewise, dendritic integrity appeared to be unaffected by the injury, with no reproducible loss or reductions in microtubule-associated protein 2 (MAP2) immunoreactivity observed at the site of injury nor in the ipsilateral hippocampus (Fig. 6C), areas known to suffer dendritic loss following more severe forms of injury. 25,26
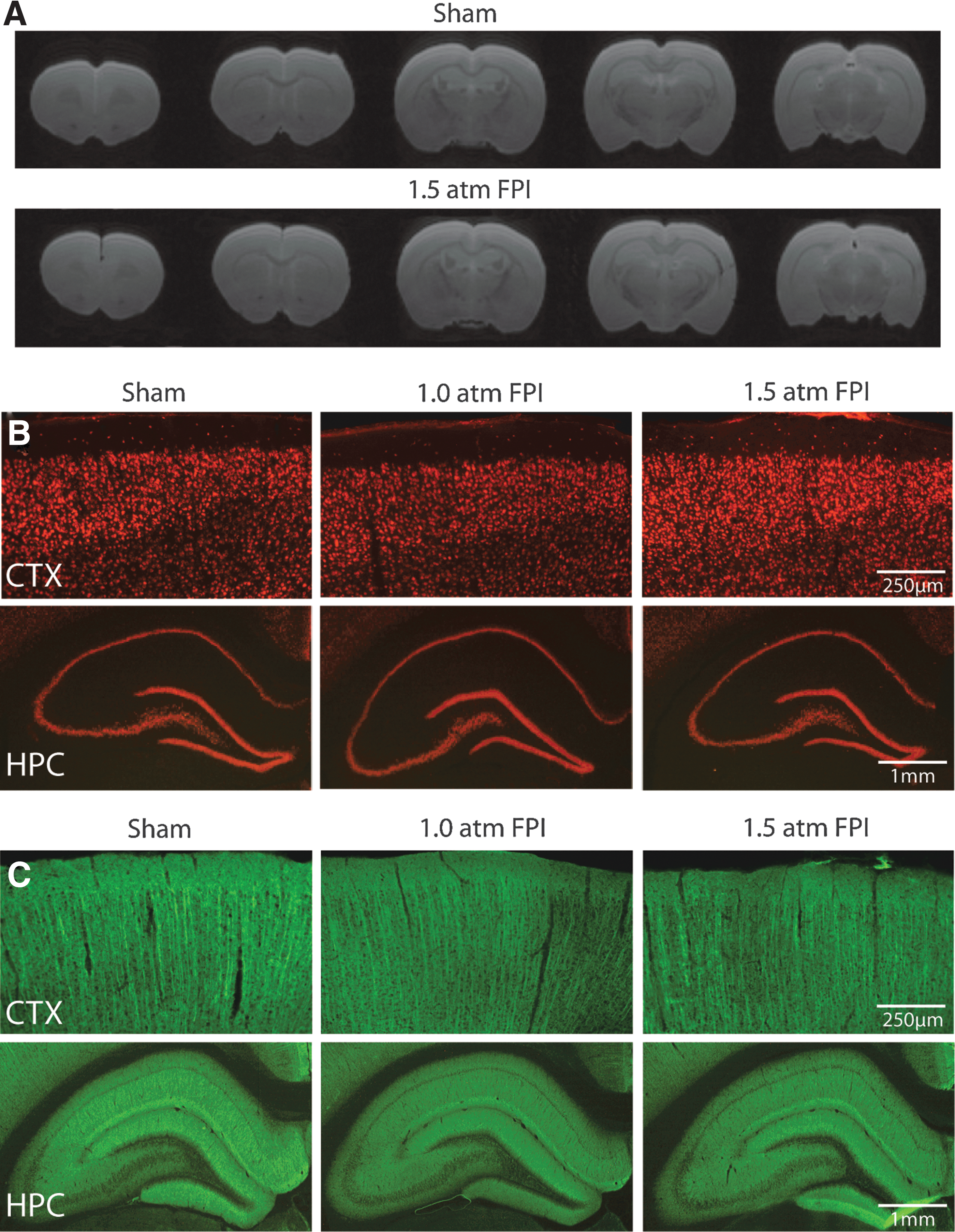
FPI of 1.5 atm did not cause overt cell loss or dendritic damage.
1.5 atm FPI increases indices of axonal damage
Recent imaging studies are beginning to show that mild TBI is associated with abnormal findings detected using Diffusion Tensor Imaging (DTI). 27,28 To examine if 1.5 atm FPI causes axonal disturbances that can be detected using DTI, brains from sham (n=6) and 1.5 atm FPI (n=6) were used for ex vivo imaging. Representative colorized FA maps from a sham and an injured animal are shown in Figure 7A. ROI analysis revealed that 1.5 atm FPI caused a significant decrease in FA values in the cingulum (Fig. 7B). The representative tract tracings shown in Figure 7C demonstrate a reduction of the fibers with FA values over a threshold in the ipsilateral cingulum of the 1.5 atm FPI rat. When tissue sections from sham and FPI animals were used for silver staining, dramatic increases in silver impregnation were observed in the corpus callosum (Fig. 7D), the cingulum (cg; Fig. 7E), and the internal (IC) and external (EC) capsule (Fig. 7F) of 1.5 atm FPI rats. Although to a lesser extent, enhanced silver impregnation was also observed in these areas when silver staining was carried out using tissue sections from 1.0 atm rats (Fig. 7D–F).
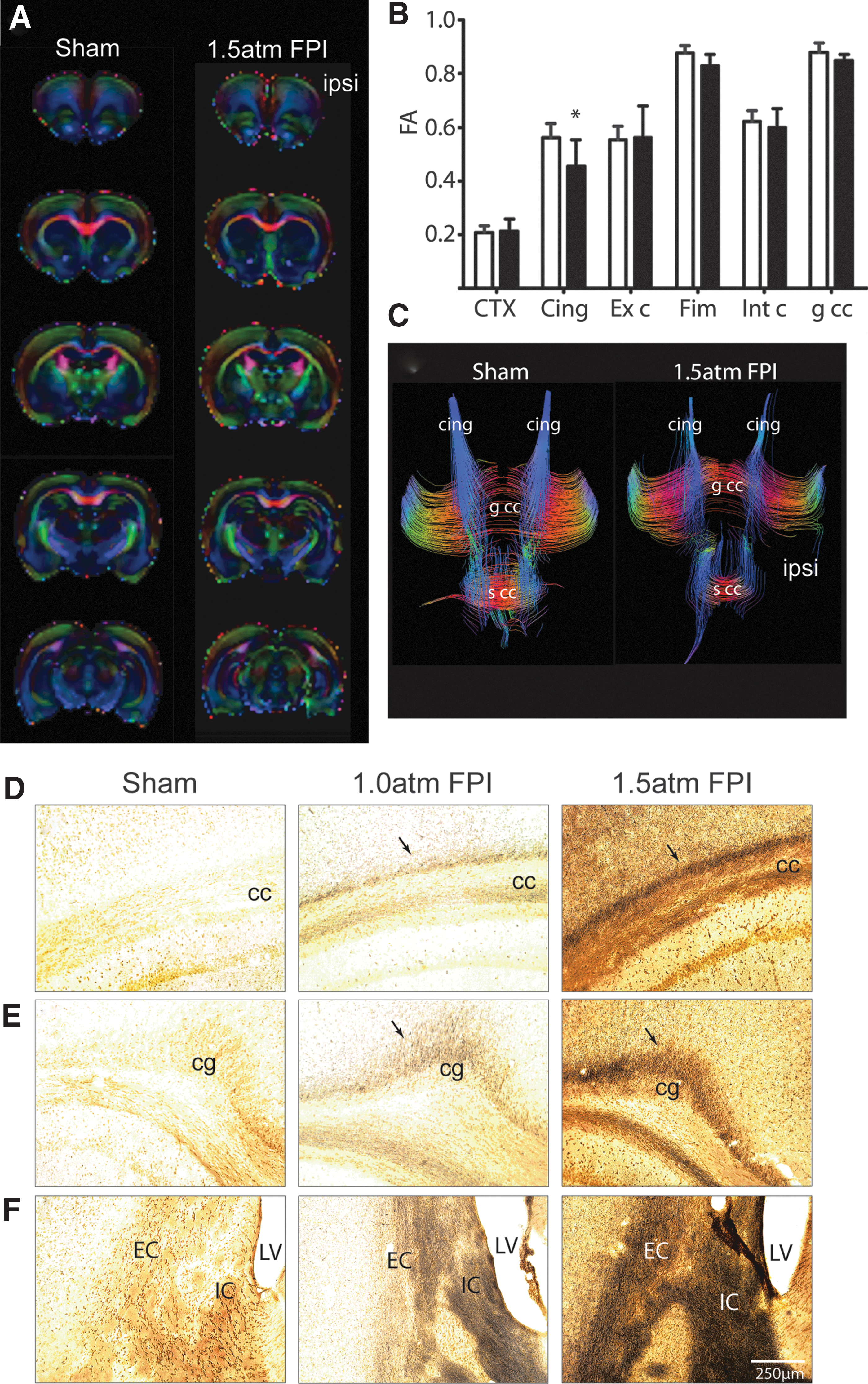
Mild FPI increased markers of axonal damage.
Further supporting a conclusion of axonal damage, we detected an increase in amyloid precursor protein (APP) immunoreactivity in the corpus callosum (cc) ipsilateral to the injury of 1.5 atm FPI animals (Fig. 8A). As it had been previously reported that unmyelinated axons in the corpus callosum are more vulnerable to TBI, 29 we examined if the increased APP immunoreactivity we observed was localized to myelinated (Fig. 8B) or nonmyelinated axons using double-label immunohistochemistry. In contrast to our expectations, the representative confocal pictures shown in Figure 8C demonstrate that APP immunoreactivity (red) was predominantly located in MBP-positive (green) myelinated axons.
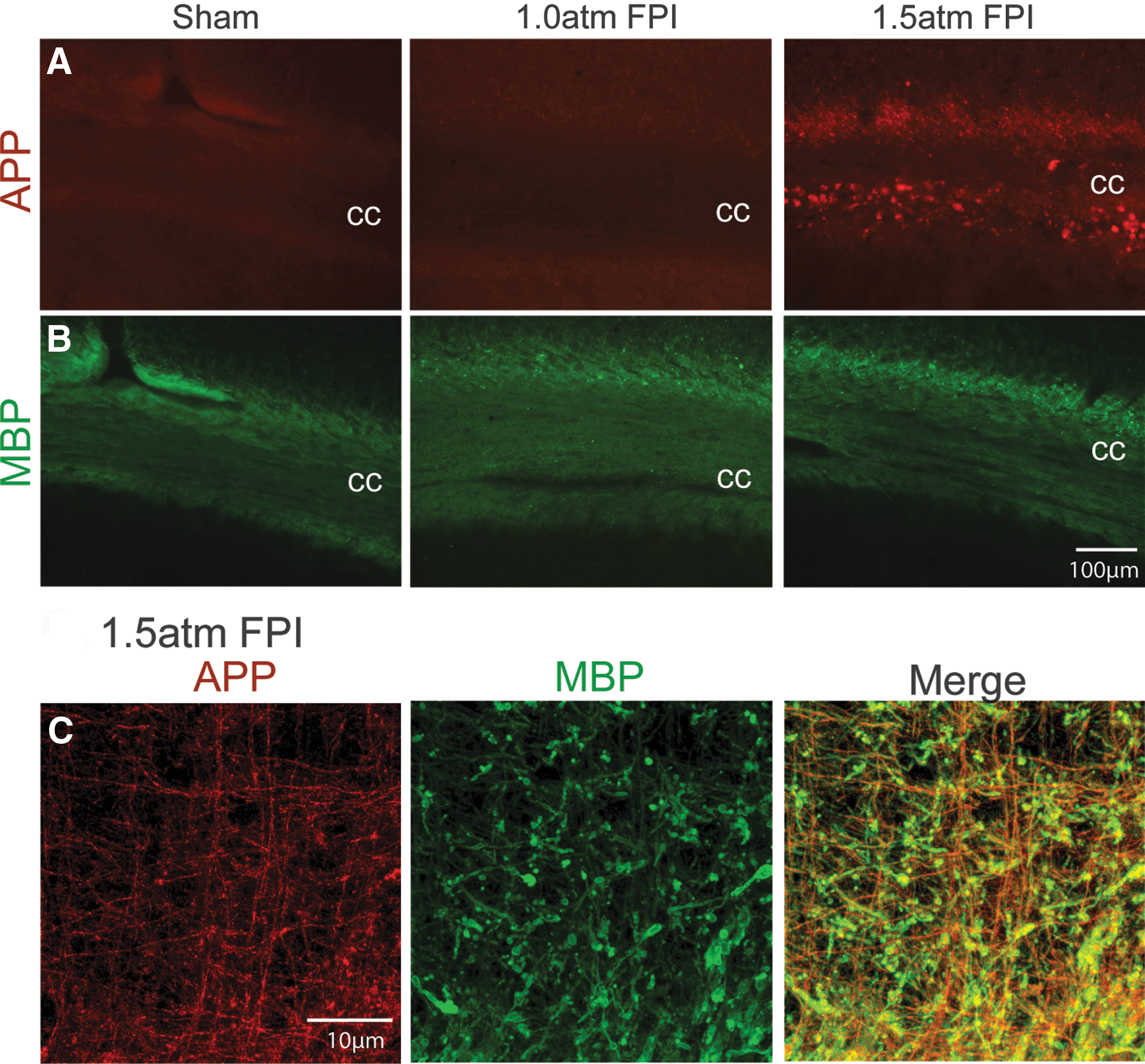
Mild FPI enhanced APP immunoreactivity.
1.5 atm FPI increased markers of inflammation
To examine inflammation after mild FPI, we investigated if enhanced Iba-1 (a marker of microglia) and GFAP (a marker of astrocytes) immunoreactivity, could be observed at 15 days after the injury. Figure 9A shows that enhanced Iba1 and GFAP immunoreactivity can be seen in the corpus callosum of both 1.0 atm FPI and 1.5 atm FPI rats compared to that seen in sham-operated controls. Similarly, a robust increase in Iba-1 and GFAP can be seen in the thalamus (Fig. 9B). These observations suggest that mild FPI caused an inflammatory response that persisted for at least 2 weeks after injury.
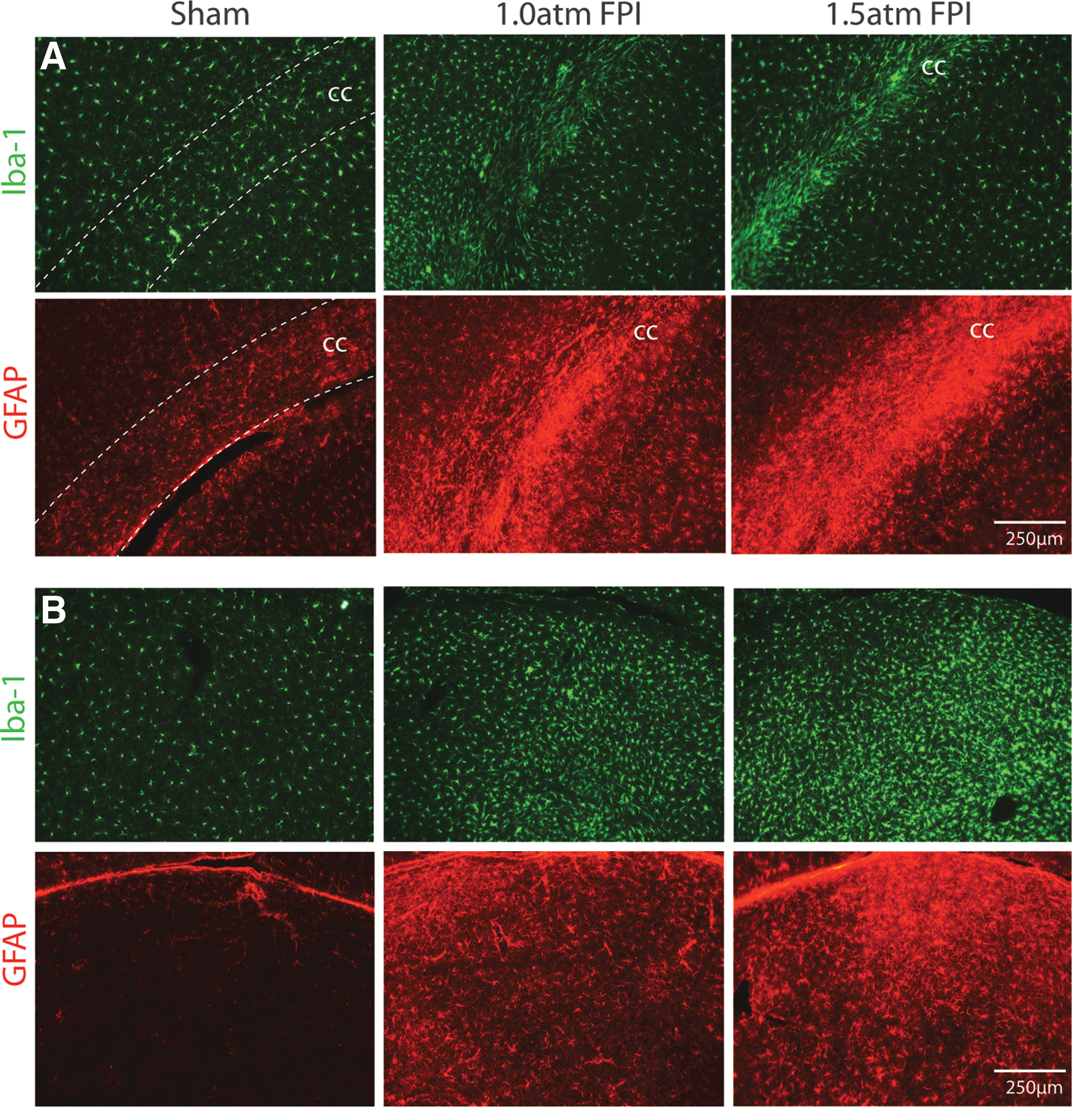
FPI of 1.5 atm increased Iba-1 and GFAP immunoreactivity. Representative photomicrographs of tissue sections taken 15 days after sham surgery, 1.0 atm FPI or 1.5 atm FPI are shown.
Discussion
In the present study, we employed 1.0 atm and 1.5 atm FPI levels to investigate the neurobehavioral and histopathological changes triggered by mTBI. Our results show the following five key findings: 1) Neither 1.0 atm nor 1.5 atm mFPI caused visible contusion injury to the brain; 2) Neither injury caused visible hippocampal or neocortical neuronal loss, a finding consistent with previous reports; 3) 1.5 atm, but not 1.0 atm, FPI caused transient motor and significant learning and short-term memory impairments; 4) Both 1.0 atm and 1.5 atm FPI caused a local inflammatory response in the corpus callosum and the thalamus that persisted for at least 2 weeks after the injury; and 5) Both 1.0 atm and 1.5 atm FPI caused axonal damage in the corpus callosum, and internal and external capsules, findings consisted with clinical findings.
One of the defining features of mTBI is the temporary loss or alteration of consciousness. While consciousness is difficult to assess in rodents, investigators have employed the duration of suppression of righting response as a rodent correlate for the period of loss of consciousness. Using this measure, we found that 1.0 atm and 1.5 atm FPI injury resulted in durations of suppression of righting responses of 2.8 min and 5.8 min over sham, respectively. This period of suppression is consistent with a classification of mild from other studies. 13,16 In addition to loss of consciousness, some mTBI patients display problems with balance and motor function in the subacute phase of injury. Although neither 1 atm FPI nor 1.5 atm FPI caused vestibulomotor deficits, 1.5 atm FPI caused temporary motor deficits on both the ipsilateral and contralateral sides. Although our lateral FPI was centered on the right parietal cortex, motor deficits on both sides are consistent with the reported diffuse nature of injury caused by FPI. These deficits were related to visually coordinated behavior, as indicated by increased foot fault errors, but not gross locomotion as both injured and sham animals performed equally well on a rotarod task.
Previous studies in rats, cats, and pigs documented that FPI caused alterations of regional blood flow. 31 –33 For example, Ginsberg et al. 32 reported that in neocortical regions ipsilateral to the trauma, blood flow was depressed by 44% in moderately-injured FPI rats as compared to that measured in sham-operated controls. Using a scanning laser-doppler device, we observed that mild FPI also results in a significant reduction in ipsilateral cerebral perfusion within minutes of injury that lasted throughout the 1 h monitoring period, returning to normal values by 24 h post-injury. At present, it is not clear if this suppression was due to impaired cerebral autoregulation or systemic physiological changes such as decreased arterial blood pressure, changes in blood carbon dioxide or oxygen levels, or decreased heart or respiratory rates. Previous studies that have monitored the systemic response to FPI have not observed significant influences on cardiovascular parameters. For example, Yamakami and McIntosh 34 observed that regional cerebral blood flow after moderate FPI is reduced and occurs in the absence of changes in mean arterial blood pressure, heart rate, or blood gas levels. Similarly, DeWitt and colleagues observed significant reductions in both global and regional cerebral blood flow, but not in other cardiovascular parameters or blood chemistry, after moderate FPI. 35,36 Although these physiological measures were not carried out in the present study, these findings suggest that local vascular changes may underlie the decreased cerebral perfusion we observed. While the consequences of this reduced cerebral perfusion is not known at present, it has been suggested that reduced blood flow may make the injured brain more vulnerable to a subsequent mild TBI or ischemic insult. Consistent with this, Robertson and colleagues reported that hypotension, when initiated soon after cortical impact injury, results in marked brain damage and contusion. 37
The temporal lobe, especially the hippocampus, is vulnerable to TBI. Consistent with this, mTBI patients often exhibit short-term declarative memory deficits (e.g., recall of a list of words after a short delay) that can be observed for days after the injury, and in some cases much longer. In order to test FPI animals in a short-term task analogous to that used in humans, we employed an abbreviated water maze task to test hippocampal-dependent spatial learning and short-term memory recall. In this task, animals were given a single training session (consisting of 10 trials) on day 5 post-injury, followed by a probe trial 30 min later. Although we report changes in molecular pathways including inflammatory pathways after 1 atm FPI <
In contrast to that seen at 1.0 atm FPI, 1.5 atm FPI caused significant learning and short-term memory deficits when animals were tested in the abbreviated Morris water maze task on day 5 post-injury. This observation raises the possibility that this injury may result in hippocampal neuron loss and/or neuronal dysfunction. In agreement with a previous report by Eakin and Miller, 13 we did not observe visible hippocampal cell loss after 1.5 atm FPI. These investigators, however, did observe that injured animals had a reduction in the number of neurons having spatiotemporal activity, suggesting that deficits in spatial learning and memory as a result of 1.5 atm FPI may result from altered place cell firing rather than loss of hippocampal neurons. As one of our goals was to evaluate the consequences of mild FPI using testing procedures reflective of those typically performed in humans, we did not test memory recall with a longer delay.
Concussions are not typically associated with gross gray matter damage, 39,40 but have recently been shown to cause diffused axonal injury as detected by DTI. For example, mild TBI patients have reduced FA values in both the genu and splenium of the corpus callosum, the internal and external capsule, the left superior cerebellar peduncle, the cingulum, the hippocampus, the thalamus, the dorsolateral prefrontal cortex, and other brain regions. 5,27,41 –47 Interestingly, depending on the time points examined, either increases or decreases in FA values have been observed, a finding thought to be related to the development of axonal swelling that occurs early after injury. 48 Quantification of FA values showed a significant decrease in the cingulum of 1.5 atm FP brains 15 days post-injury. However, differences in FA values did not reach significance in other regions of interest (Fig. 8). Using more sensitive histopathological analysis, we observed enhanced APP immunoreactivity in the corpus callosum of 1.5 atm FPI animals, suggesting axonal injury. This was further supported by the detection of silver-impregnated axons in this fiber bundle, as well as in the internal and external capsule, the cingulum, and the thalamus. Previous ultrastructural analysis and electrophysiological recordings have suggested that unmyeliniated axons are more vulnerable to TBI. 29,49 However, Creed et al. 50 reported a reduction of compound action potential (CAP) amplitude after TBI that was restricted to myelinated axons. Consistent with this later study, our double immunostaining showed that the enhanced APP immunoreactivity seen in the corpus callosum after 1.5 atm FPI occurred predominantly in myelinated axons. At present, it is unclear why the more severe form of FPI used by Reeves resulted in unmyelinated axon injury, whereas the lower magnitude of FPI used in this study damaged myelinated axons.
One of the key findings in the present study is that mild FPI, both 1.0 atm and 1.5 atm, resulted in a dramatic increase in Iba-1 and GFAP immunoreactivities within the corpus callosum and the thalamus, suggesting persistent inflammation. This is consistent with our genomic studies <
In summary, we found that both 1.0 atm and 1.5 atm FPI injury caused transient suppression of acute neurological functions that occur in the absence of visible contusion to the cerebral cortex. Further, we show that 1.5 atm injury led to transient motor disturbances, and significant impairments in spatial learning and short-term memory. Consistent with previous studies, 1.5 atm FPI does not cause visible neuronal loss in the hippocampus or in the neocortex, but gives rise to a robust inflammatory response. Measures of axonal injury (e.g., DTI, APP immunoreactivity, and silver staining) identified several areas with axonal disturbances, including the cingulum, the corpus callosum, and the internal and external capsules. These findings are consistent with the deficits and pathologies associated with mild TBI in humans, and support the use of mild FPI as a model to evaluate putative therapeutic options.
Footnotes
Acknowledgments
The authors would like to thank Dr. Bruce Lyeth of the University of California, Davis for kindly providing the hubs used to mount the rats to the FPI device. We also thank the Imaging Core of the Mission Connect Mild TBI Translational Consortium under the direction of Dr. Ponada Narayana for their assistance with the MRI. The work performed in the authors' laboratories were made possible by grants from DOD (W81XWH-08-2-0134) and TIRR Foundation.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.