
Abstract
The similarities and differences between acute nonconvulsive seizures (NCS) and other epileptic events, for example, periodic epileptiform discharges (PED) and intermittent rhythmic delta activities (IRDA), were characterized in rat models of penetrating and ischemic brain injuries. The NCS were spontaneously induced by either unilateral frontal penetrating ballistic-like brain injury (PBBI) or permanent middle cerebral artery occlusion (pMCAO), and were detected by continuous electroencephalogram (EEG) monitoring begun immediately after the injury and continued for 72 h or 24 h, respectively. Analysis of NCS profiles (incidence, frequency, duration, and time distribution) revealed a high NCS incidence in both injury models. The EEG waveform expressions of NCS and PED exhibited intrinsic variations that resembled human electrographic manifestations of post-traumatic and post-ischemic ictal and inter-ictal events, but these waveform variations were not distinguishable between the two types of brain injury. However, the NCS after pMCAO occurred more acutely and intensely (latency=0.6 h, frequency=25 episodes/rat) compared with the PBBI-induced NCS (latency=24 h, frequency=10 episodes/rat), such that the most salient features differentiating post-traumatic and post-ischemic NCS were the intensity and time distribution of the NCS profiles. After pMCAO, nearly 50% of the seizures occurred within the first 2 h of injury, whereas after PBBI, NCS occurred sporadically (0–5%/h) throughout the 72 h recording period. The PED were episodically associated with NCS. By contrast, the IRDA appeared to be independent of other epileptic events. This study provided comprehensive comparisons of post-traumatic and post-ischemic epileptic profiles. The identification of the similarities and differences across a broad spectrum of epileptic events may lead to differential strategies for post-traumatic and post-stroke seizure interventions.
Introduction
T
This clinical dilemma of post-traumatic/ischemic NCS management requires a better understanding of the mechanism of these acquired NCS in order to develop safer drugs and establish clinical guidelines to treat or prevent silent seizures. In this respect, animal models of endogenously triggered seizures bearing clinical relevance to brain injury are critically important. Traditionally, seizure research has depended heavily on animal models of either chemically (e.g., pentylenetetrazole [PTZ], pilocarpine, or kanic acid) or electrically (e.g., maximal electrical shock, or kindling) induced seizures. Historically, these models have played an important role in leading discoveries of novel AEDs, but they are not necessarily appropriate for studying the spontaneous yet random occurrence of post-traumatic/ischemic NCS, because of their different seizure dynamics; namely, 1) these models are all convulsive in nature, and 2) the convulsions/seizures occur within a relatively restricted and well-defined period of time after induction. In this regard, the most clinically relevant animal models to be used for studying brain injury-induced NCS are brain injury models themselves, which are capable of triggering NCS in the absence of a secondary trigger.
Currently, many animal models of brain trauma and stroke exist and, in their own capacity, have played pivotal roles in enhancing our understanding of the underlying mechanisms of the injury and neuroprotection. The most widely used severe TBI models are the rodent fluid percussion (FP) brain injury model, controlled cortical impact (CCI) brain injury model, and the most recently developed penetrating ballistic-like brain injury (PBBI) model. The most studied stroke models are the middle cerebral artery occlusion (MCAO) model of permanent or transient focal cerebral ischemia and the two vessel or four vessel occlusion of global cerebral ischemia. However, their role in studying post-traumatic and post-ischemic NCS has been surprisingly scarce. A possible obstacle for experimental studies of this kind is likely the inability to conduct long-term cEEG monitoring of animals in most research laboratories. Nevertheless, with this long-established capability in this arena, both post-traumatic and post-ischemic NCS have been detected and comprehensively studied in the rat unilateral frontal PBBI model and the permanent MCAO (pMCAO) model, respectively. 8,9 We have demonstrated that in both models the electrographic seizures and other EEG epileptic activities, such as periodic epileptic discharges (PED) and intermittent rhythmic delta activities (IRDA), occurred spontaneously as a result of the respective injuries, and that these events can only be detected from cEEG recordings. Critically, the EEG waveforms of these rodent epileptic phenomena closely resemble human clinical manifestations; therefore, the PBBI and the pMCAO model are considered as valid and clinically relevant animal models for studying post-traumatic and post-ischemic NCS. In the present study, we compared the EEG profiles of the nonconvulsive epileptic activities associated with these two very different brain injuries. The purpose of these analyses was to construct a clinically useful matrix that may improve our understanding of the underlying etiology of this medical problem, and aid in seizure prediction, diagnosis, and intervention.
Methods
Male Sprague–Dawley rats (275–350 g, Charles River Labs, Raleigh, VA) were individually housed in a well-ventilated vivarium under a 12 h light/dark cycle (lights on 0600–1800). Food and water were provided ad libitum before and after brain injury surgeries. All surgical procedures were performed on isoflurane (2% vapor) anesthetized but spontaneously breathing rats. The core body temperature was maintained at 37.0°C using a heating blanket (Harvard Apparatus, Holliston, MA). All experimental procedures were approved by the Institutional Animal Care and Use Committee of Walter Reed Army Institute of Research. Research was conducted in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals, and adhered to the principles stated in the Guide for the Care and Use of Laboratory Animals. The animals' housing facility was accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International.
EEG electrode implantation
Stainless steel screw EEG electrodes were used to detect NCS from the surface of the brain of each animal. The EEG electrode was made of a piece of insulated Nichrome wire (0.2 mm in diameter, uninsulated at each end) soldered to a 080-3 mm stainless steel screw (Component Supply Company, Fort Mead, FL). The anesthetized rat was secured on the stereotaxic device, and a midline incision was made longitudinally to expose the surface of the rat skull. Capillary bleeding was cauterized to keep the skull surface clean. Four epidural EEG electrodes were symmetrically screwed on the skull through tightly fit burr holes over bilateral parietal regions of the brain (1 mm anterior and 4 mm posterior to bregma,±3.5 mm lateral to midline, Fig. 1). The reference electrode was placed posterior to lambda over the transverse sinus. The free end of each electrode wire was soldered to a Dale multi-pin connector (March Electronics, West Hempstead, NY) in the sequence of A, B, C, and D, which corresponded to the left frontal, left parietal, right frontal, and right parietal mono-referential recording montage, respectively. The screws were secured to the skull with 3M Vetborne tissue adhesive (St. Paul, MN) and insulated with a layer of dental acrylic. The whole assembly was then firmly attached to the rat skull with an adequate amount of dental acrylic. (For the animals in the PBBI group, the skull region over the right frontal cortex was kept free of dental acrylic to allow the cranial window for the PBBI probe insertion). All rats were allowed 5 days to recover from the EEG surgery before receiving PBBI or pMCAO injury.
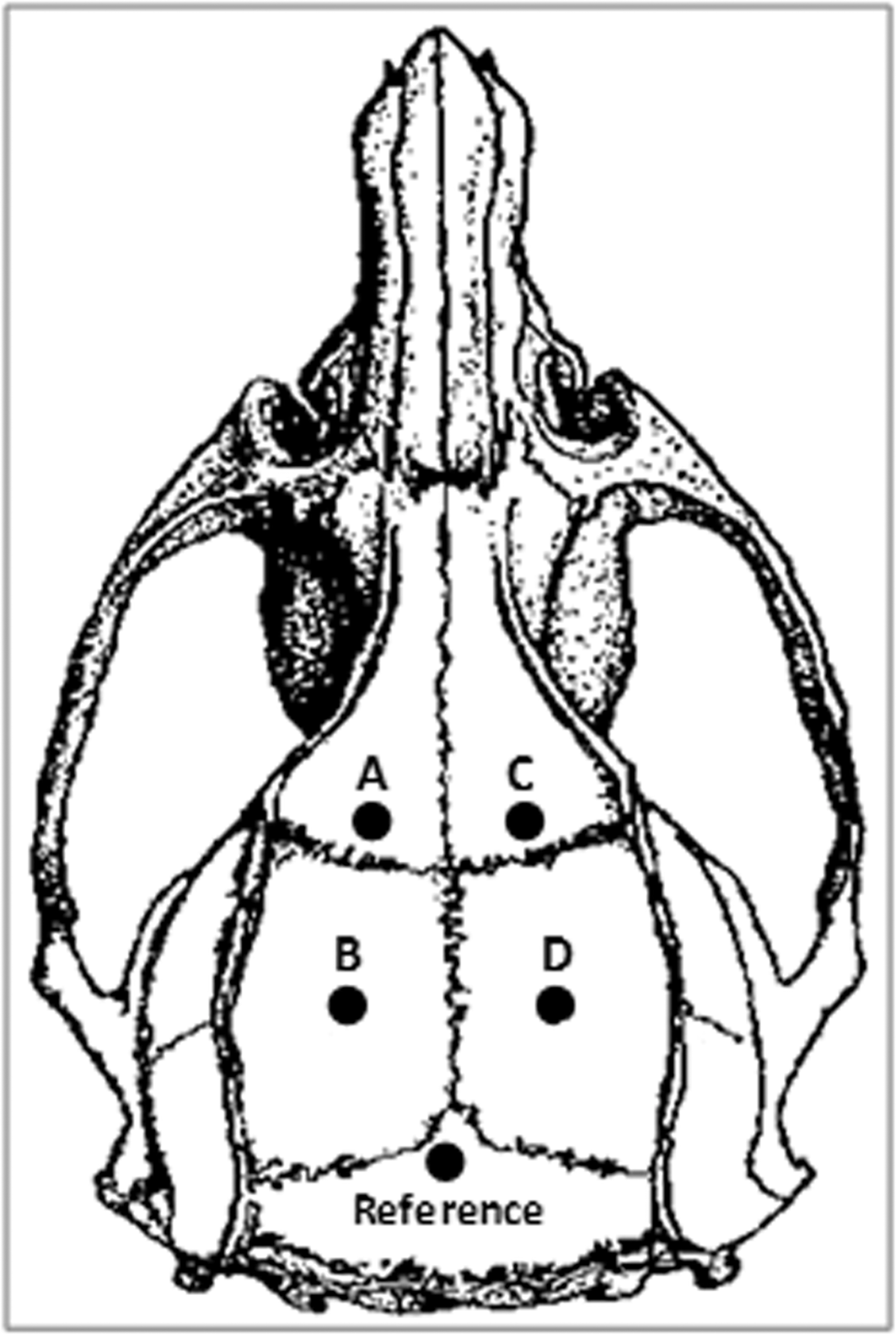
Illustration of the dorsal surface of a rat skull and the electroencephalographic (EEG) electrode placements. The letters A, B, C, and D, represent EEG electrodes over the left frontal, left parietal, right frontal, and right parietal regions of the brain, respectively.
Post-traumatic NCS induced by PBBI
The 10% right frontal PBBI model was used to induce post-traumatic-NCS (n=44). This model has been well characterized 10,11 and successfully used in studies of neuroprotection. 12 As previously described, the PBBI apparatus consists of a computer-controlled pressure generator operated by a cylinder impact system filled with water (Mitre Corp, McClean VA) and a specially designed PBBI probe made of a 20G stainless steel tube. One end of the probe had lines of perforated holes tightly sealed by a piece of elastic tubing (1 cm in length). The other end of the probe was connected to the pressure generator. During the surgery, the anesthetized rat was secured on a stereotaxic device. A cranial window (4 mm diameter) was created to expose the right frontal pole of the brain. The PBBI probe was secured to the arm of the stereotaxic device at an angle of 50 degrees vertically and 25 degrees laterally (counterclockwise). It was manually inserted through the center of the cranial window (+4.0 mm AP, +2 mm ML from bregma) into the right frontal cortex to a distance of 1.2 cm from the surface of the brain. Upon activation by the computer, the pistol of the pressure generator was released, triggering a rapid inflation and deflation of the elastic tubing on the PBBI probe to an elliptical-shaped water balloon inside the brain, creating a temporary cavity along its path. The probe was then retracted from the brain, the cranial opening was sealed with sterile bone wax, and the incision was closed with sterile wound clips.
The injury severity level of the PBBI was defined by the size of the inflated water balloon. For the 10% PBBI, the balloon size was calibrated to have a diameter of 0.633 cm, yielding a balloon volume equal to 10% of the brain volume. However, the ballistic kinetics of the injury may vary for the same 10% PBBI depending upon the amount of energy dissipated from the balloon inflation, that is, a more rapid balloon inflation produces more energy than a slower inflation to the same balloon size. For example, in this study the 10% PBBI was achieved at a pulse duration of 36 ms for the balloon inflation, which required an input pressure of 65 psi. This 10% PBBI caused a transient (10–20 sec) apnea, evidently generating a stronger ballistic impact than the 10% PBBI used previously at a pulse duration of 40 ms and an input pressure of 45 psi, which did not cause apnea. 13 The ballistic pressure inside the brain was not measured.
Post-ischemic NCS Induced by pMCAO
The intraluminal filament model of MCAO was used to induce post-ischemic NCS (n=28). This MCAO model has been widely used for studying focal ischemic brain injury, including numerous studies by our group. 9,14,15 During the surgery, the anesthetized rat was placed in a supine position. A midline incision was made to expose the right common carotid artery at the level of external and internal carotid artery bifurcation. The external carotid artery and its branches were cauterized and cut at the lingual and maxillary artery branches. A piece of 3-0 monofilament nylon suture (Ethilon, Somerville, NJ) with a round tip was inserted into the internal carotid artery via the stump of the external carotid artery. The filament was advanced ∼21 mm beyond the bifurcation of the internal and external carotid arteries until a slight resistance was felt as it was lodged in the narrowing of the anterior cerebral artery and the origin of the middle cerebral artery was completely blocked. The suture was left in place for 24 h until the end of the experiment, to allow a permanent occlusion of the middle cerebral artery.
EEG recordings
On the day of PBBI or pMCAO surgery, rats were placed individually in custom-designed Plexiglas EEG recording chambers (30 cm long×30 cm wide×50 cm high) equipped with multi-channel gold contact swivel commutators (Dragonfly Inc., Ridgeley, WV). The multi-pin electrode connector on the rat skull was connected to the swivel commutator via a flexible shielded cable, allowing the free movement of the animal during EEG recordings. The swivel commutator was interfaced with an EEG amplifier and the EEG signals were recorded at a sampling rate of 200 samples/sec and digitized by computerized data acquisition systems. For the PBBI group, Stellate amplifier and Harmonie software were used (Natus Medical Incorporated, San Carlos, CA) and for the pMCAO group, Grass Model D15 amplifier and Grasslab software were used (GRASS Technologies, West Warwick RI). The baseline EEG recording was collected from unanesthetized and freely moving animals for at least 30 min prior to PBBI or pMCAO surgery. The recording was interrupted during the surgery but immediately resumed thereafter and continued for 72 h after PBBI and 24 h after pMCAO. Video cameras were installed on top of the EEG cages to monitor animal's behavior during EEG recordings for the PBBI group. For the pMCAO group, animals' behavior was monitored by experimenters for the first few hours after the injury, when the majority of seizures occurred.
EEG data analysis
NCS
As described previously, an NCS event was defined as rhythmic spike discharges occurring at a frequency of 1–4 Hz for at least 10 sec with the amplitude distinctively greater than the background activities. 9,10 These criteria were adopted for both injury models in this study, with modifications to differentiate different EEG patterns of NCS. Generalized NCS were defined as seizures detected from bilateral recording channels, whereas focal NCS were defined as seizures detected from a single or multiple channels of one cerebral hemisphere. Based on these criteria, the following descriptive parameters were included in the statistical analyses: 1) NCS incidence, the number of animals that experienced at least one NCS event; 2) NCS frequency, the number of NCS events experienced by an individual animal; 3) NCS episode duration, the duration (sec) of one NCS event; 4) NCS total duration, the sum of NCS episode durations (sec) for an animal; 5) NCS onset latency, the time (h) lapse between PBBI or pMCAO and the occurrence of the first NCS event, and 6) NCS hourly time distribution, the number of NCS occurring per hour expressed as the percentage of the total number of NCS occurring during the entire recording period for each injury group. These NCS parameters were expressed as the mean and standard errors of the mean in the result section.
PED
In general, PED describes an EEG pattern consisting of stereotyped spikes, sharp or slow wave discharges repeating in a periodic or pseudo-periodic pattern at irregular intervals. 9,16 In this study, PED are defined as five or more consecutive spike or sharp-wave discharges with an inter-discharge interval of 2–30 sec. Like seizure events, the amplitude of PED is significantly higher than the background activities. Unlike seizure events, PED did not occur with a metronomic rhythmicity and the onset and offset of a series of PED could be ambiguous because of their varying inter-discharge intervals. For this reason, the abundance of PED was graded using a rank score system, that is, a score of 1, 2, or 3 was assigned to a mild (isolated), moderate (intermittent), or severe (consistently reoccurring) degree of PED, respectively.
IRDA
IRDA is an EEG pattern originally described by Cobb. 17 It consists of distinct bursts of rhythmic spike- or sharp-wave oscillation in the delta-theta (3–8 Hz) frequency range. For the purpose of evaluating the relationship between IRDA and other epileptic activities, the same rank score system described for PED was used to define mild (score 1), moderate (score 2), and severe (score 3) degrees of IRDA.
Results
Seizure statistics
As shown in Table 1, 70% (31/44) of the PBBI animals and 82% (23/28) of the pMCAO animals developed NCS after the respective injuries, and the NCS activities were more intense following pMCAO than following PBBI. In the PBBI group, a total of 297 post-traumatic NCS were identified during the 72 h post-injury period. Each NCS event lasted for an average of 36.9±3.1 sec (range=16–98 sec). When the seizure-free animals were excluded from the group, the PBBI animals experienced an average of 9.6±1.6 NCS events per rat (NCS frequency), which constituted a NCS total duration of 325.1±60.4 sec per rat. In the pMCAO group, a total of 556 post-ischemic NCS were identified. Although the duration of an individual NCS event was similar to that of the PBBI animals (45.7±4.8 sec/event, range=25–114 sec), the NCS frequency was 24.7±5.1 NCS events per rat (after excluding seizure-free animals from the pMCAO group), and the NCS total duration was 1116.9±206.4 sec per rat during the 24 h post-pMCAO period. Collectively, pMCAO-induced NCS were >2.5 times more frequent and lasted 3 times longer than those experienced by a PBBI animal on average in a 72 h post-injury period (Table 1).
The majority of the PBBI and pMCAO animals exhibited NCS. However, seizure activities after the ischemic brain injury were more intense than those after the penetrating brain injury as reflected in the NCS frequency and duration. Furthermore, NCS occurred more acutely with an average onset latency being <1 h after pMCAO. In comparison, NCS occurred more sporadically after PBBI with a delayed onset latency almost 24 h after the injury.
NCS, nonconvulsive seizures; PBBI, penetrating ballistic-like brain injury; pMCAO, permanent middle cerebral artery occlusion.
One of the most salient features that distinguished the post-PBBI and post-pMCAO NCS was the temporal profiles of the seizures. As shown in the insert panels of Figures 2A and B, which depict the differential profile of the NCS onset latency (expressed as the accumulated percentage of the first NCS events over time), the first NCS occurred within 2 h after the pMCAO, with a mean onset latency of 0.6 h (36 min), ranging between 0.2 and 1.5 h (Fig. 2B insert). However, after PBBI, the first NCS occurred in a random and delayed fashion with a mean onset latency of 23.9 h, ranging between 0.5 and 51 h (Fig. 2A insert). In 78% of the PBBI animals, the first NCS did not occur until 18 h after the injury. Another hallmark disparity between the post-PBBI and post-pMCAO NCS profiles was their temporal distribution after the first NCS occurred. As shown in Figure 2B, after pMCAO, 47% of all NCS events (263/556) occurred within the first 2 h post-injury (20% and 27% during the 1st and 2nd h, respectively). The hourly NCS distribution, expressed as the percentage of the total number of NCS in the group, was 0–4% between 5 and 24 h post-pMCAO. By contrast, after PBBI, 51% of all NCS events (142/279) occurred between 24 and 48 h (Fig. 2A), and the hourly NCS distribution was 0–5%/h throughout the entire 72 h recording period.
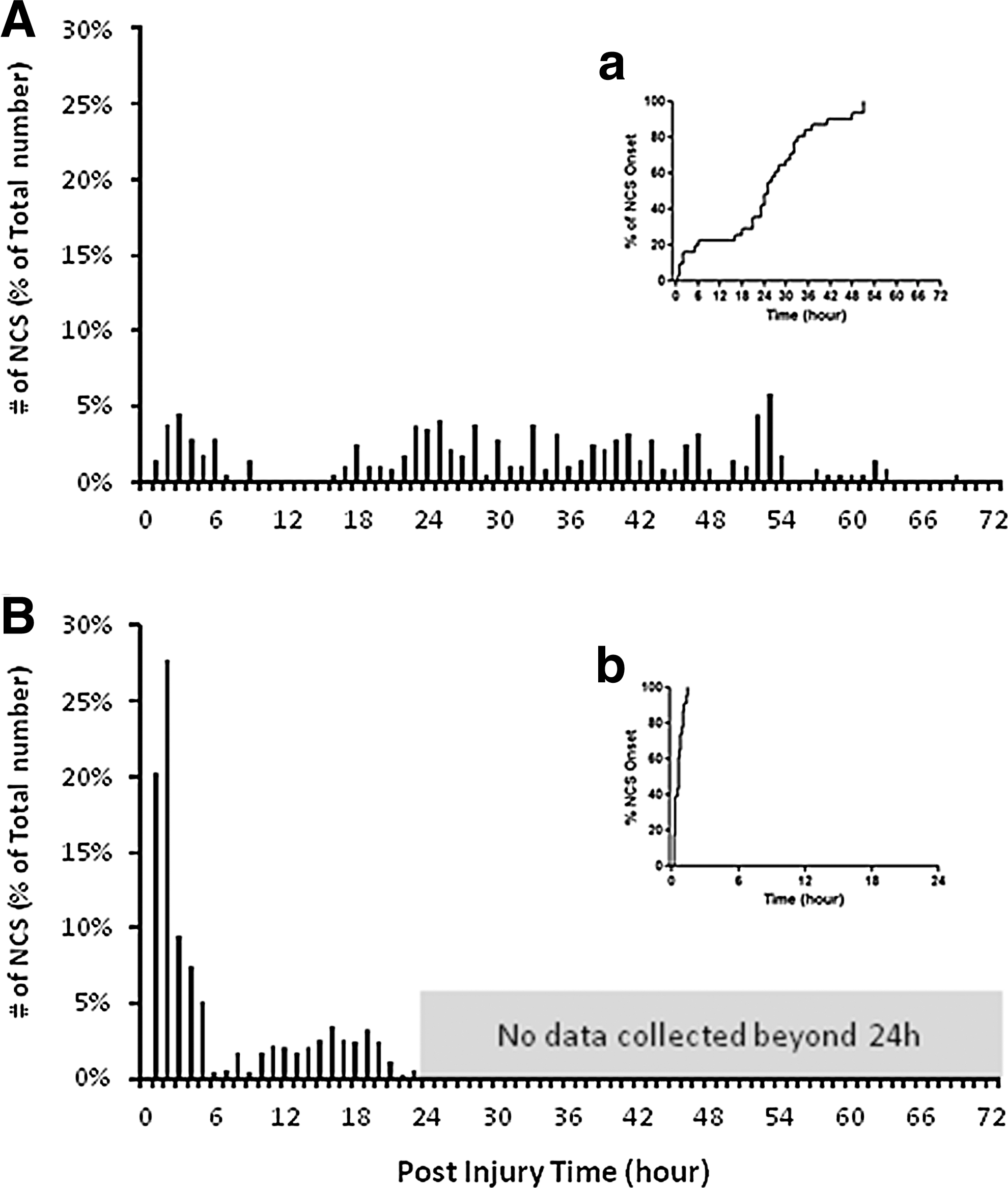
Comparison of differential temporal profiles of nonconvulsive seizures (NCS) between the penetrating ballistic-like brain injury (PBBI)
A circadian analysis showed that 77% of NCS following pMCAO and 60% of NCS following PBBI occurred between 0600 and 1800 h of the day. However, this circadian analysis was biased for the pMCAO animals, because in these animals most NCS occurred soon after the pMCAO surgery, which was always performed between 0900 and 1100 h, within the 0600–1800 time frame.
Video characterization of NCS and other epileptic activities
All seizure events detected in this study were nonconvulsive in nature, verified from the review of video recordings of the PBBI animals and the real-time observation of the pMCAO animals. In most cases, animals remained in one position before, during, and after a seizure. Occasionally they continued engaging in ongoing behavior, for example, eating, drinking, head scratching, or paw licking, when seizures initiated. However, subtle behavioral expressions were often observed accompanying the electrographic seizure episodes. These behaviors included repetitive jaw movement (“chewing” behavior), head nodding, twitching or “wet dog shake,” and attempted but incomplete circling followed by freezing or head nodding. Such behaviors usually often ended abruptly when the NCS stopped, but also sometimes continued for 2–5 more seconds. Sometimes the animal appeared to transition from a sleep state to an awakened state 5–15 sec prior to the onset of an NCS as shown in Fig. 3A–E.
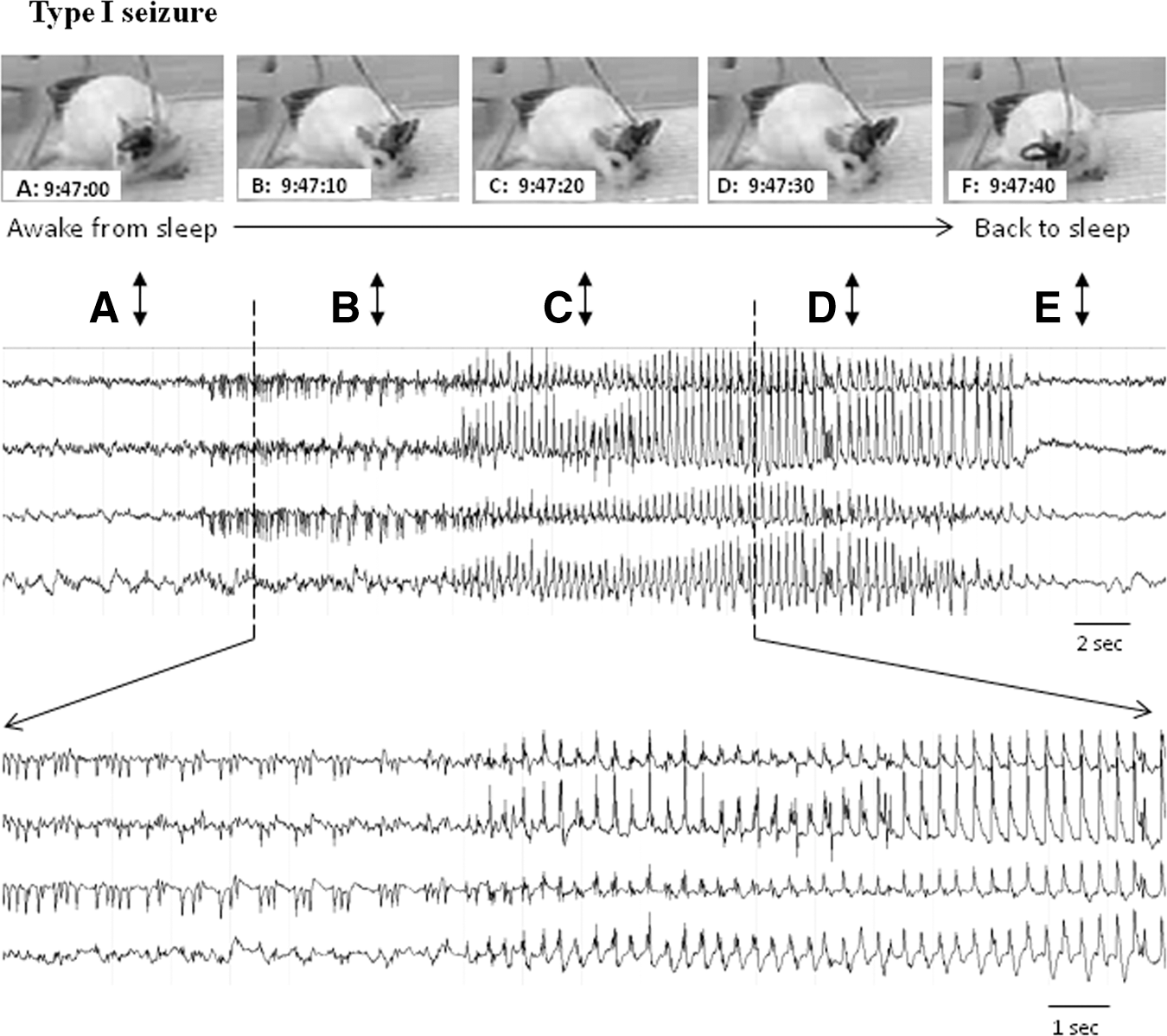
An example of penetrating ballistic-like brain injury (PBBI)-induced generalized type I nonconvulsive seizures (NCS). A series of images (
These subtle but characteristic behavioral features accompanying the NCS events were absent during PEDs or IRDAs, and no other behavioral expressions were identified that were specifically associated with either of these events.
EEG waveform patterns of NCS and other epileptic activities
NCS
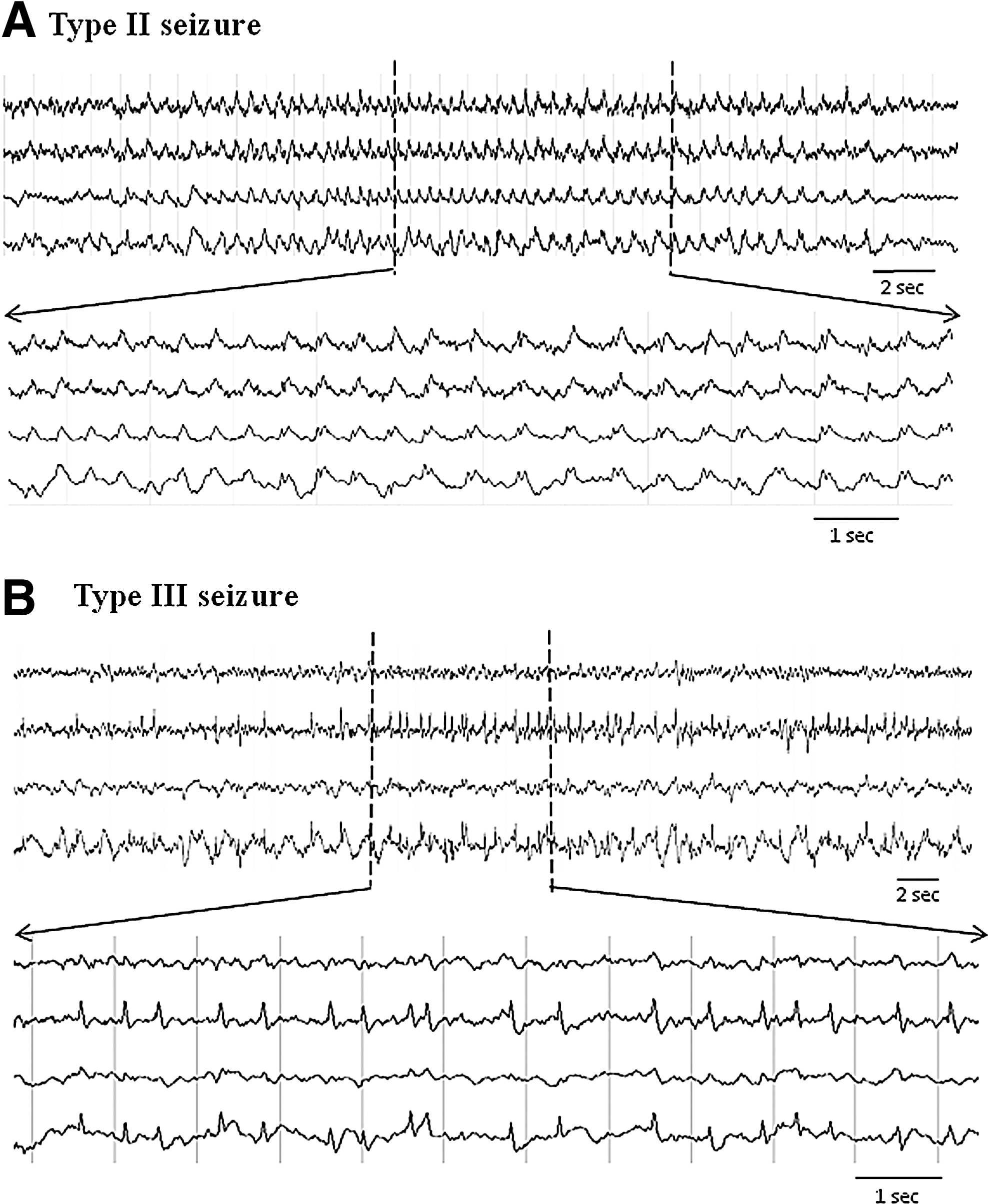
Independent of behavioral expressions, several distinct EEG waveform patterns of NCS were identified. They were characterized as 1) rhythmic spike discharges occurring at a frequency >1 Hz (type I seizures, Fig. 3, middle panel); 2) rhythmic sharp-wave discharges occurring at a frequency of ∼1 Hz (type II seizures, Fig. 4A); and 3) non-rhythmic spike clusters occurring at a frequency 0.5–2 Hz (type III seizures, Fig. 4B). Among a total of 297 NCS detected in the PBBI model and 556 NCS detected in the pMCAO model, the type I seizures accounted for 79% of the post-PBBI NCS and 96% of the post-pMCAO NCS (Fig. 5). The Type II seizures appeared to be very rare (1%) following the pMCAO injury compared with PBBI (14%), whereas the type III seizures accounted for 4–7% of the seizures in both brain injury models (Fig. 5). Whereas there are clear differences between these seizure patterns, sometimes type I NCS evolved from type II or type III NCS, and occasionally different NCS patterns coexisted in one seizure episode detected from different EEG channels.
(
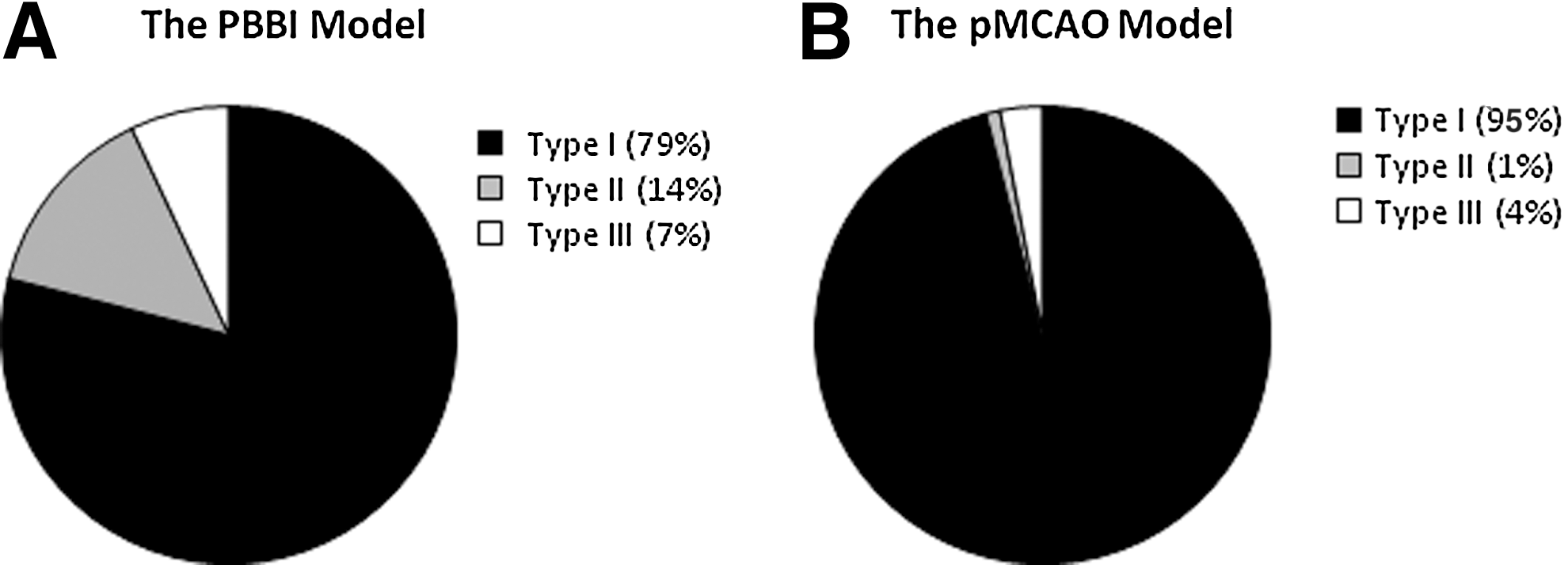
The nonconvulsive seizures (NCS) type distributions following the penetrating ballistic-like brain injury (PBBI)
Among all seizure types, >80% of NCS were generalized seizures in each injury group (86% for the PBBI group and 81% for the pMCAO group). In some cases, a generalized NCS evolved from and/or faded into a focal seizure either ipsilaterally or contralaterally. All animals in each group experienced generalized NCS; in addition to generalized seizures, 45% PBBI subjects and 87% pMCAO subjects also experienced focal seizures.
PED
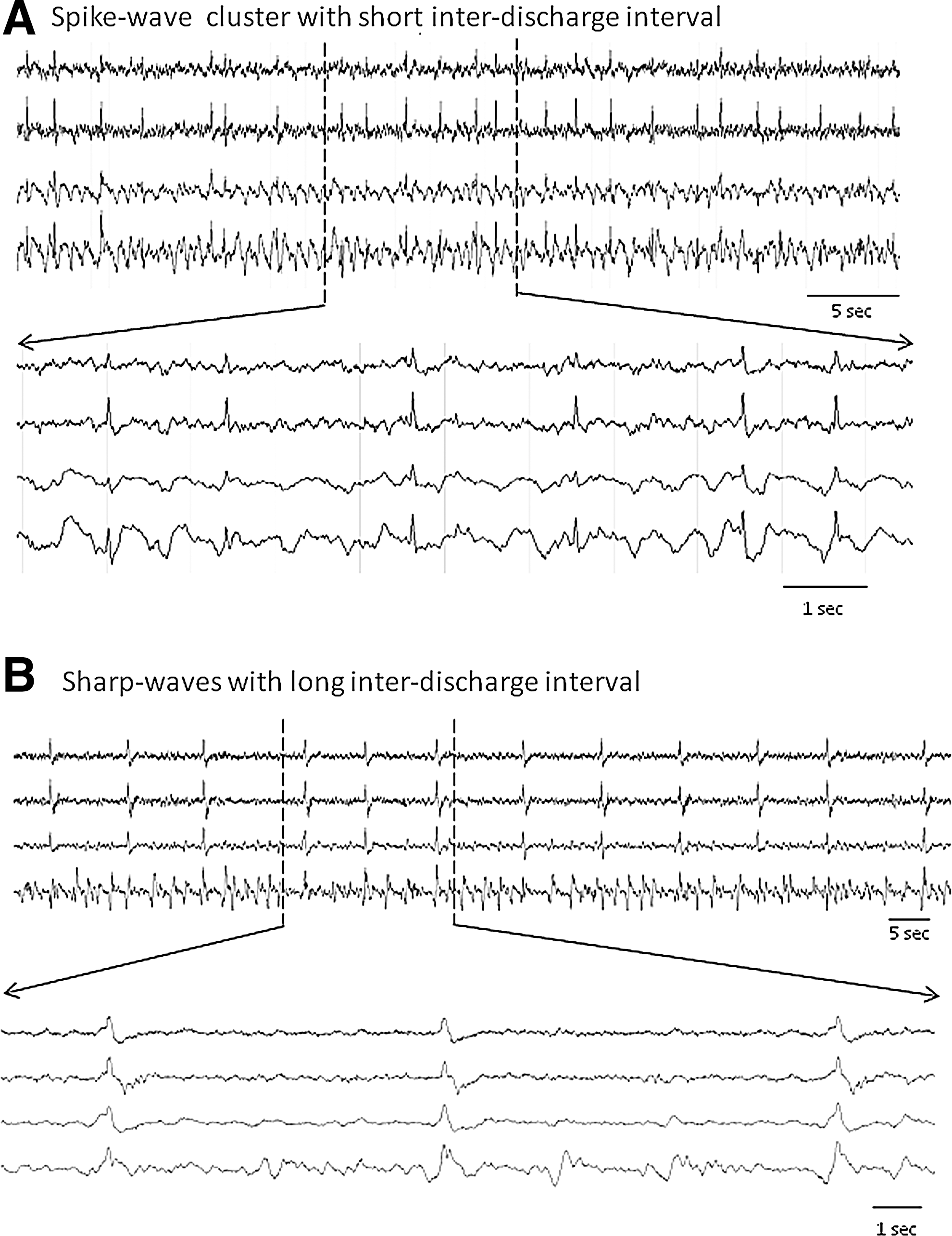
PED of different waveforms were detected in 97% of PBBI animals and 72% of pMCAO animals. The EEG characteristics of PED following the PBBI and pMCAO were previously described in detail. 8,9 Those descriptions also applied to the phenomena observed in this study. In general, the PED detected in the pMCAO model were mostly sharp wave discharges (∼200 ms) occurring at a short inter-discharge interval of 1–4 sec or 4–10 sec, and mainly in the ipsilateral hemisphere. They closely resembled clinical descriptions of lateralized PED (PLED). 18 However, in the PBBI model, EEG patterns of PED were more variable, consisting of spike waves (∼60 ms at zero crossing) with a short inter-discharge interval (1–4 sec, Fig. 6A) and sharp waves (∼230 ms, Fig. 6B) with a long inter-discharge interval (up to 30 sec). Topographically, they occurred ipsilaterally, contralaterally, or generalized, but most frequently they were detected bilaterally from the anterior parietal cortex (channels 1 and 3) or the posterior parietal cortex (channels 2 and 4). These variations were observed either within or between different animals. For both injury models, the majority of PED occurred independent of the NCS activities, only in tandem with 10% of the total NCS activities (10.1% for the PBBI model and 9.7% for the pMCAO model).
IRDA
The IRDA were another distinct abnormal EEG phenomenon detected in most PBBI (96%) and pMCAO (73%) animals. Similar to what has been described previously, 9 IRDA were bursts of rhythmic spike- or sharp-wave oscillations at a frequency 3–8 Hz (most commonly at 5–6 Hz, the low frequency range of the theta band) and had an amplitude distinctively greater than the background (Fig. 7). Each burst lasted 2–4 sec and occurred either as a single isolated event or as a series of events in clusters. Unlike the disparity of EEG characteristics exhibited in NCS and PED, variations of IRDA were minimal, except that after the pMCAO, IRDA emerged and intensified acutely within the first few hours of the injury. Conversely, after the PBBI, IRDA were less active until after 24 h post-injury, and became increasingly intensified during 48 and 72 h post-injury. Interestingly, they occurred exclusively in the contralateral hemisphere in both brain injury models.
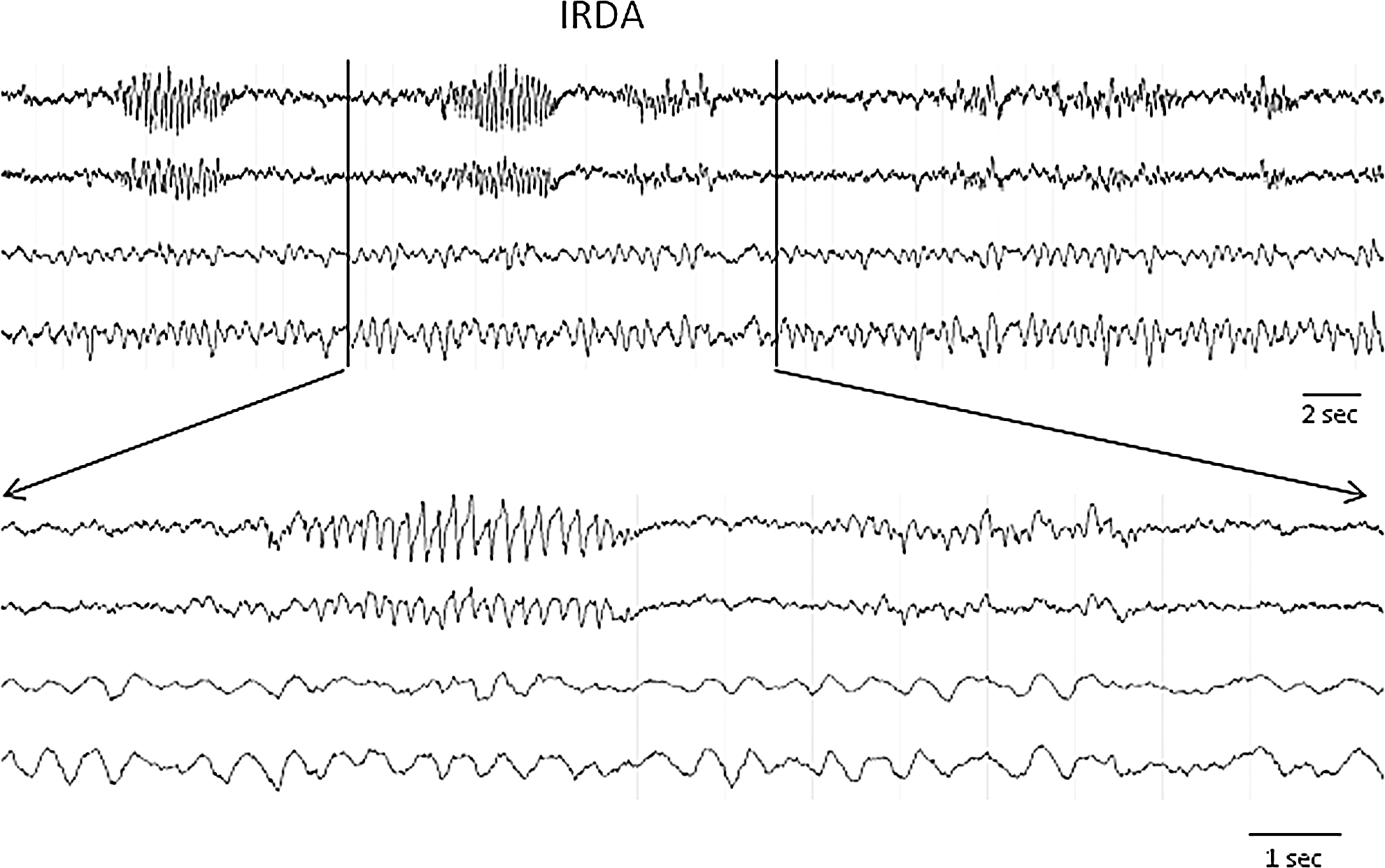
An example of penetrating ballistic-like brain injury (PBBI)-induced intermittent rhythmic delta activities (IRDA) consisting of bursts of rhythmic spike-sharp wave oscillations at a frequency of 5–6 Hz. They occurred in isolation or in clusters.
Correlations analysis
The 10% PBBI produced brain lesions along the injury tract affecting the frontal cortex and the dorsolateral striatum as shown on Fig. 8A, with an average lesion volume equal to 86±4.1 mm3. The pMCAO produced ischemic infarction, which affected the same cortical and striatal regions as the PBBI lesions, but more extensively, including the entire striatum and the lateral parietal and partial temporal cortex (Fig. 8B), with an average infarct volume of 335±19.6 mm3. In the PBBI animals, the lesion volume did not have significant correlations with the NCS severity measured by major seizure parameters (NCS frequency [Fig. 9A], duration, onset latency, p>0.05 for each measure). However, in the pMCAO animals, the infarct volume was significantly correlated with NCS frequency (Spearman ρ=0.42, p<0.05, Fig. 9B), NCS duration (Spearman ρ=0.48, p<0.05), onset latency (Spearman ρ=−0.41, p<0.05), and generalized NCS (Spearman ρ=0.52, p<0.05). Because nearly 50% of NCS in the pMCAO group occurred within the first 2 h post injury, even before the infarction developed, further analysis that separated the acute (0–2 h post-pMCAO) and delayed (3–24 h post-pMCAO) NCS revealed that the number of early seizures did not have significant correlation with the infarct volumes, but that the number of the delayed NCS significantly correlated with the infarct volume.
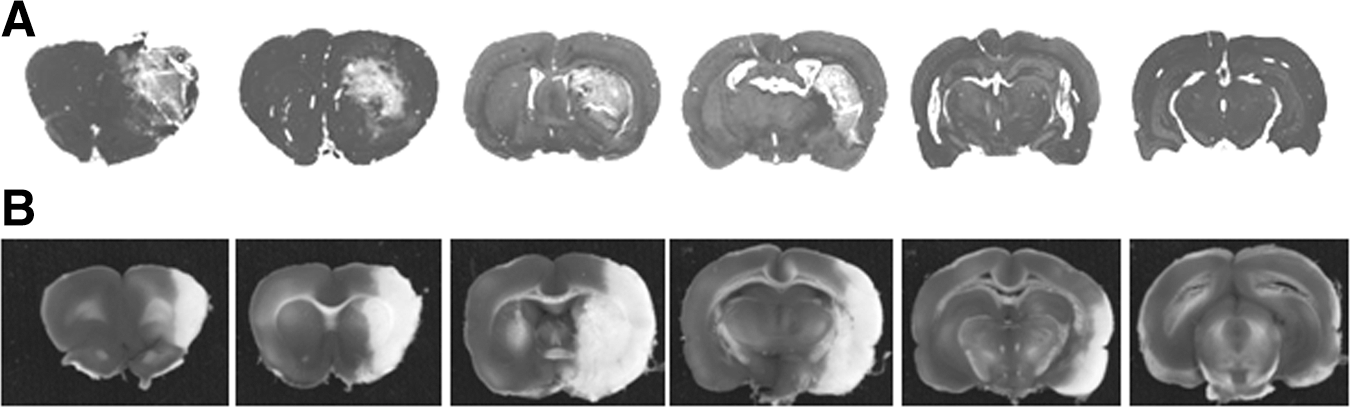
Brain sections of selected animals showing the extent of the brain lesions
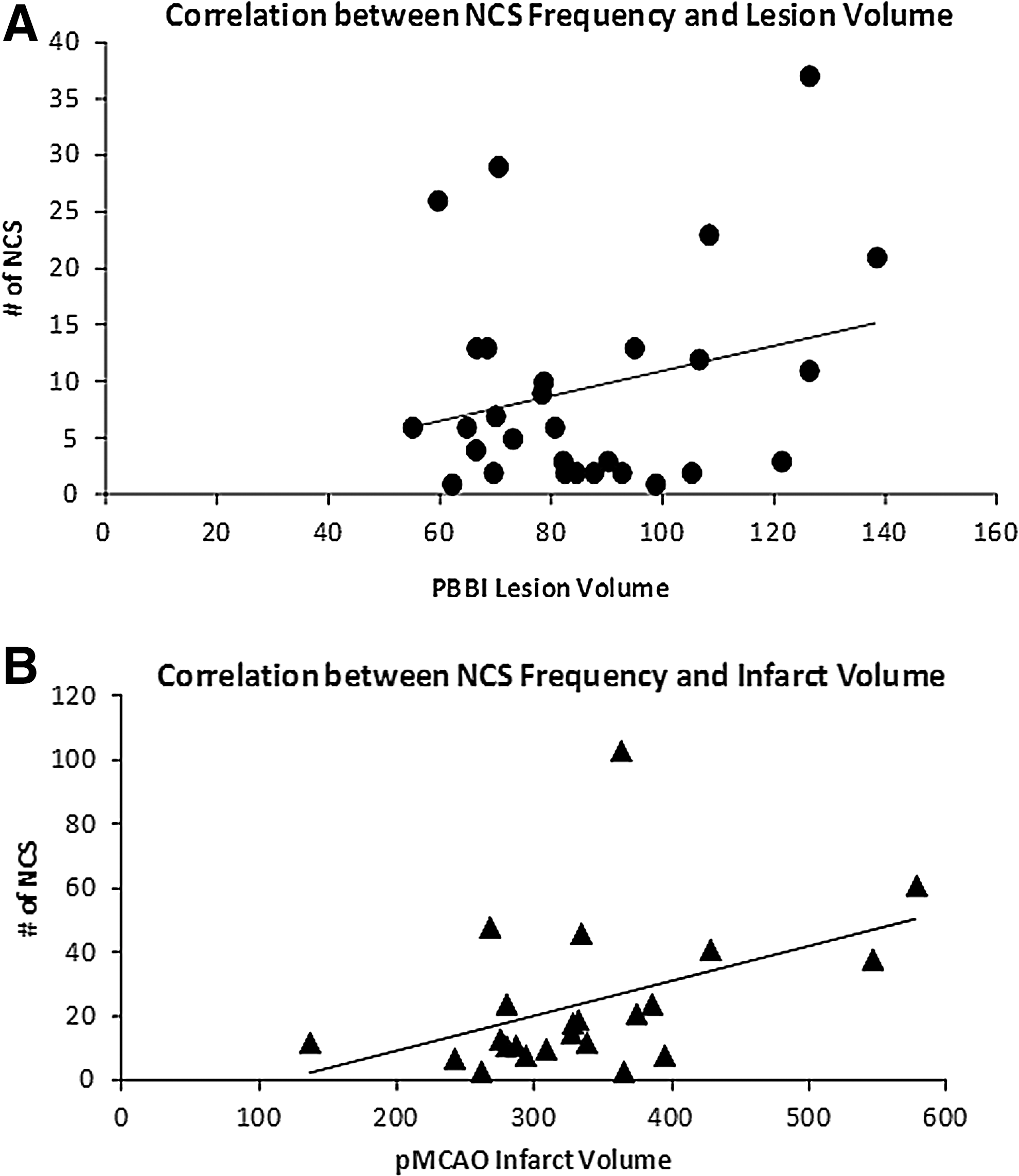
The correlations between the nonconvulsive seizures (NCS) frequency and the brain damages.
The correlative relationships among epileptic activities were examined based on the abundance of their occurrence measured by the NCS frequency and the abundance scores of PED and IRDA described in the Methods section. The nonparametric correlation analysis tests revealed that in the PBBI animals, the NCS frequency was positively correlated with the abundance of the PED (Spearman ρ=0.36, p<0.05) and that this correlation was mainly attributed to the correlation between the type I NCS frequency and the spike PED of short inter-discharge intervals (Spearman ρ=0.37, p<0.05, Fig. 10). By contrast, in pMCAO animals no correlations were identified between the NCS severity and the PED abundance.
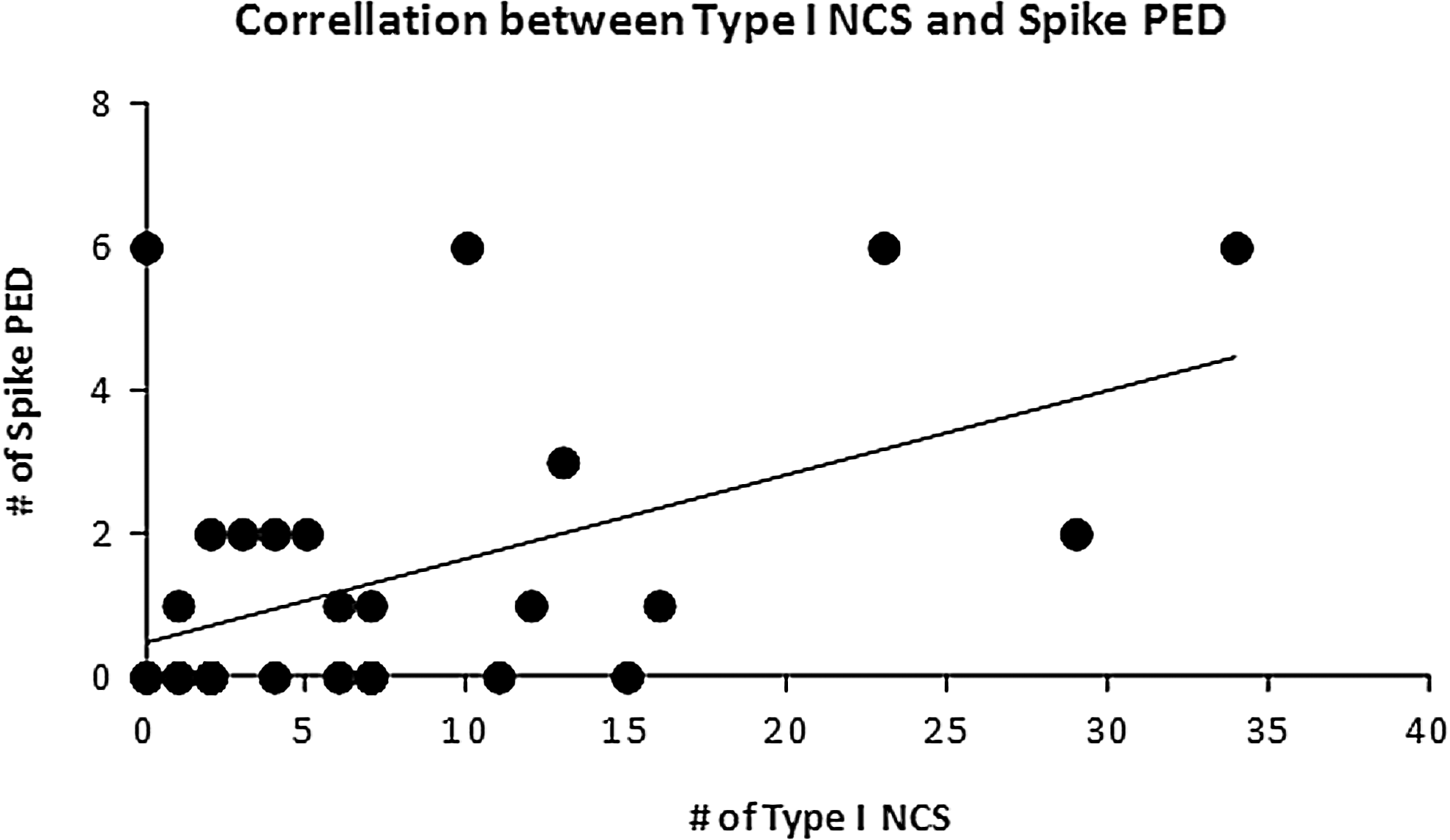
Correlations between the type I nonconvulsive seizures (NCS) frequency and the abundance of the spike periodic epileptiform discharges (PED) of short inter-discharge intervals in the penetrating ballistic-like brain injury (PBBI) model (p<0.05).
For both PBBI and pMCAO groups, the IRDA appeared to be an independent index of epileptic events and not predictive of any other epileptic activities, because it had no significant correlation with either NCS or PED (data not shown).
Discussion
In this study, clinical features of post-traumatic and post-ischemic epileptic activities were characterized in two animal models of severe brain injuries: the rat PBBI and pMCAO models with a special interest in nonconvulsive hyperactivities, for example, NCS, PED, and IRDA. Among several hundred electrographic seizures identified by EEG, no overt convulsive motor activities were observed to accompany the seizure episode, although subtle behavior expressions similar to the “skeletal seizures” or electrically kindled behaviors were observed. The analysis of EEG waveform patterns of these nonconvulsive epileptic activities revealed that both NCS and PED exhibited intrinsic variations that resemble human EEG signals of ictal and inter-ictal events. These variations, although seen in both models and not etiologically specific, were relatively more explicit following the PBBI than following the pMCAO, which allowed further identification of inter-relationships between subtypes of NCS and PED after the PBBI, that is, a stronger correlation existed between type I NCS and the spike-type of PED in the PBBI model. The most salient characteristics that differentiated the post-PBBI and post-pMCAO NCS were the timing and intensity of the occurrence of NCS after the two injuries. As shown in Table 1 and Figure 2, the NCS occurred more acutely and intensely soon after the pMCAO, but more delayed and sporadically after the PBBI. These unique temporal profiles could bear significant implications in determining the correlations between the severity of the NCS and the severity of the injury. Taken together, the EEG pathology observed in these two rodent injury models could be informative for optimizing treatments for patients with TBI and stroke.
Nonconvulsive seizures
Compared to clinical NCS incidence in TBI (∼2–23%) and stroke (∼2–33%) patients, 1,4 –6,19,20 the NCS incidence in PBBI and pMCAO animals was relatively high (≥70% for both models). This discrepancy is likely attributed to the controlled versus the uncontrolled injuries experienced by laboratory animals and NICU patients, respectively. For example, for the post-traumatic NCS, all animals used in this study received the same right frontal PBBI at the same 10% injury severity level, whereas the cohort of TBI patients included in clinical studies represents a heterogeneous population with TBI from various causes (concussive vs. penetrating), severities (mild to severe), and anatomical locations (frontal, lateral, and caudal). For the post-stroke NCS, in addition to the problem of the heterogeneity of the disease (ischemic vs. hemorrhagic, and focal vs. global), the timing of EEG recording may also significantly influence the estimation of the clinical NCS statistics. In our pMCAO model, in which cEEG recording was initiated immediately after pMCAO, >40% of the animals experienced the first seizure in <30 min after the injury, and nearly 50% of all seizures were detected within the first 2 h post-injury. These ultra-acute post-ischemic NCS have never been reported in human stroke patients largely because of the logistical difficulty of performing EEG monitoring so close to the onset of the ischemic insult. Therefore, the inability to collect EEG data during the peak hours of NCS activities might significantly underestimate the clinical NCS incidence in stroke patients.
The initial insults of penetrating and ischemic brain injuries are different in etiology; however, they both elicit similar acute physiological, cellular, and molecular responses in the brain; 21 one can now add epileptogenesis to the list. Because EEG signals of post-traumatic and post-ischemic NCS share similar variations that are not etiology specific, it is a clinical challenge to provide proper seizure managements tailored for patients with different types of brain injuries. Therefore identification of NCS characteristics other than electrographic expressions capable of differentiating post-traumatic and post-ischemic NCS would be helpful for developing injury-specific treatment strategies for critically ill patients.
The results obtained from this study demonstrated that the most salient feature that differentiated post-traumatic and post-ischemic NCS was their temporal distribution following the PBBI and pMCAO. After the frontal PBBI, NCS occurred sporadically and randomly with a late onset; however, after the pMCAO, NCS occurred more acutely and intensely soon after the injury. The implications of these unique temporal profiles of NCS after the penetrating and ischemic brain injuries may be twofold. First, seizure management in NICU is time sensitive. The results from this study suggest that different strategies with respect to the timing of the treatment of post-traumatic and post-ischemic NCS are warranted, for example, post-ischemic seizures should be treated more aggressively during the acute phase of the injury, whereas post-traumatic seizures could be treated more selectively when time allows for referencing other epileptic activities. Second, the temporal differences between post-traumatic and post-ischemic NCS also provide an opportunity to address an important clinical question: that is, to what extent NCS activities exacerbate the underlying injury, and conversely, to what extent the severity of the injury aggravates the NCS activities. In this study the analysis of correlations between NCS activity and the brain damage revealed that the NCS activity significantly correlated with the infarct volume in the pMCAO model, but did not correlate with the lesion volume in the PBBI model. However, further analysis showed that following pMCAO, nearly half of the NCS occurred even before the formation of the ischemic infarction. 22 More importantly, these early NCS (0–2 h post-pMCAO) did not significantly correlate with the infarct volume, but instead, it was the delayed NCS activities (3–24 h post-pMCAO) that significantly correlated with the infarct volumes. These findings suggested that the ultra-early post-ischemic NCS were not a direct consequence of structural damage, but more likely the immediate brain hyperactivity in response to the metabolic derangements caused by a sudden ischemic attack. However, the delayed development of NCS during the maturation of infarction may have contributed to the worsening of the injury. This being the case, the severity of post-ischemic NCS activities would seem to be more likely responsible for exacerbating the ischemic brain injury.
For the penetrating brain injury, the PBBI caused instantaneous structural damage, which expanded and matured into injurious lesions over time. 11 The erratic occurrence of NCS appeared to be independent of the maturation process of the brain lesion. Previously we reported that the severity of NCS activities was positively correlated with the severity of PBBI defined by the temporary cavity created by different sizes of the inflated balloon (i.e., 5%, 10%, and 12.5% PBBI). Increasing the balloon size increased the NCS severity. 8 However, comparing the NCS profile of the current study and the previous study, it is now apparent that increasing the ballistic kinetics of the PBBI could also affect the animals' susceptibility to NCS. For example, in this study, the 10% PBBI with a stronger ballistic impact (36 ms pulse duration) induced NCS in 70% of the animals (9.6 NCS per animal), whereas in our previous study, the 10% PBBI with a lower ballistic energy (40 ms pulse duration) induced NCS in 39% of the animals (3.6 NCS per rat). Therefore, the NCS are sensitive to both the size of the temporary cavity and the ballistic kinetics created by the balloon inflation, suggesting that, contrary to the ischemic brain injury, the severity of the penetrating brain injury might play an important role in aggravating the severity of NCS. Nevertheless, the cause and effect between brain injuries and NCS is not mutually exclusive. Treatment with neuroprotective agents may ameliorate brain injury induced acute seizures, whereas anti-seizure managements may in turn improve patients' prognosis by reducing the energy demand imposed upon the vulnerable brain.
Although the military relevance of the rat PBBI model is well recognized, this model is also highly relevant for studying civilian penetrating brain injury, considering that firearm use accounted for 10% of all TBIs in many states in the United States (Traumatic Brain Injury in the United States: A Report to Congress, Centers for Disease Control and Prevention, 1999). However it is also recognized that the vast majority of both military and civilian TBI victims sustain severe concussive or closed head injuries; therefore, studies of post-traumatic NCS resulting from non-penetrating brain injuries is equally important. Unfortunately, as mentioned, reports of post-traumatic NCS in models of TBI such as FP and CCI brain injury models are rare, and less comprehensive in design. This makes it difficult to compare NCS characteristics across different models within the same injury modality. Nevertheless, evidence of early post-traumatic nonconvulsive epileptic activities and isolated NCS events have been reported in both rodent FP and CCI models. For example, during a study of delayed post-traumatic epilepsy after the FP brain injury, Kharatishvili et al. randomly selected three animals from their subject cohort to receive EEG electrodes implantation prior to the brain injury. 23 Similar to the design of our study, cEEG was collected from these three animals immediately after the injury for 72 h. Scattered epileptiform patterns, for example, spikes (similar to PEDs) and short runs of high amplitude sharp waves (similar to IRDAs), were detected, and one of the animals exhibited electrographic seizure activity (equivalent to type I NCS reported here) at 8 h after the injury. In a separate study, Zweckberger et al. reported EEG epileptiform activities in 67% of the animals within 4 h after the CCI injury. 24 (The authors referred to these epileptic activities as “EEG seizures,” but clearly defined criteria or illustrations were lacking.) The results of these limited studies indicate that the incidence of post-traumatic NCS may vary greatly depending upon the type of TBI, especially when the energy kinetics of the injury becomes a concern; such that the low NCS incidence observed in the closed head injury model may be attributed to the lack of ballistic impact to the brain as exerted by the PBBI model. Therefore, refining the rodent prototype FP and CCI brain injury models may be necessary to enable studies of non-penetrating TBI induced post-traumatic NCS. As for the ischemic brain injury, reports of NCS in animal models other than the intraluminal filament MCAO model are not available. However, in addition to the previous and current studies from our laboratory, 9,14,15 Luckl et al., using the transient filament MCAO model and the same NCS criteria as ours, detected NCS in 60–70% of the injured animals, 25 confirming the high incidence of NCS following focal ischemic brain injury.
Other epileptic activities
Both PED and IRDA are abnormal EEG phenomena indicative of brain function disorders. However, the clinical implication of PED and IRDA in acute brain injuries has not been clearly delineated. PED are widely considered a form of brain hyperexcitation within the continuum of ictal-interictal activities, 18,26 –28 and have been detected in TBI and stroke patients. 29 –34 However, it remains controversial to what extent these PED warrant treatments similar to those recommended for NCS. In general, the EEG characteristics of PED are complicated by their temporal and spatial variations. PED with short and long inter-discharge intervals have been referred to as periodic short-interval diffuse discharges (PSIDDs) and long-interval diffuse discharges (PLIDDs). 18,26,35 Their topographic distributions can also be classified as generalized PED (GPED), bilateral independent PED (BIPLED), or lateralized PED (PLED). In this study, similar to the NCS profiles, the variations of PED patterns in morphology and topography were more prominent after PBBI than after pMCAO. These variations allowed further identification of stronger associations between the subtype of the spike PED clusters and the NCS activities, indicating that recognition of electrographical heterogeneity of epileptic activities is important in determining differential risk factors involved in potentiating post-traumatic and post-ischemic NCS.
PED are usually observed in association with acute and subacute brain structure damage (lesions) with different etiologies. Clinical studies have shown that PED are more commonly seen in stroke patients than in TBI patients (26% for stroke vs. 2% for TBI, 29 and 46% for stroke vs. 2.4% for TBI 34 ). In the current study, the high incidence of PED after pMCAO was consistent with the clinical data, suggesting that the involvement of metabolic disorders may in part be the underlying risk factor. On the other hand, the extremely high incidence (90%) of PED in the PBBI animals was a striking contrast to the clinical reports. This remarkable discrepancy was again probably attributed to the controlled animal model of PBBI used in this study compared with the heterogeneous causes of TBI in patients. However, it may also suggest that PED could be uniquely evoked from a penetrating brain injury rather than from a concussive injury. Importantly, in patients, PED is frequently, but not always, accompanied by motor seizures, 30,36,37 which was not the case in our rat model. Rather, in our study, their direct association with NCS was episodic following either penetrating or ischemic brain injury, as in both models only 10% of all NCS occurred in tandem with PED, and 90% of PED occurred in a random fashion independent of NCS. Nevertheless, PED are clinical markers of a neurological state of increased susceptibility to the development of seizures (convulsive or nonconvulsive seizures). Treatment could be warranted depending upon the intensity of specific types of PED.
Compared with NCS and PED, IRDA seemed to have the fewest characteristics that distinguish post-traumatic from the post-ischemic brain injuries. Clinically, IRDA are often classified based on anatomical origin, that is, frontal, occipital, or temporal IRDA. Although the occipital and temporal IRDA are more commonly associated with seizures than the frontal IRDA, 38 they are not etiologically specific and are relatively less common in TBI and stroke patients. Recently, D'Ambrosio et al. 39 described an electrocorticography (ECoG) phenomenon in a rat lateral FP injury model, which seemed to be very similar to the IRDA observed in this study in the PBBI and pMCAO models. In their study the brief (∼2 sec) bursts of delta or theta spike oscillations were referred to as “epiletiform electrocorticography events (EEE),” and were considered as true post-traumatic ictal activities because of the concomitant behavioral components during each event. Disagreement exists in interpreting such a brief and seemingly ictal event. 40 In our study, no behavioral arrests were observed accompanying the IRDA. They were independent of other epileptic activities in regard to the timing and abundance of their occurrence. Therefore, the clinical implication of this phenomenon in acute traumatic and ischemic brain injury remains unclear. However, it is noteworthy that IRDA originated exclusively in the contralateral hemisphere where no structural damage was induced and the EEG background was relatively stable. The alterations of brain function in the contralateral hemisphere, distant from the injury site, have been called “transhemispheric diaschisis” and were suggested to represent a compensatory defense mechanism through remote inhibition or disinhibition. 41 The transhemisphereic diaschisis may be expressed in the EEG as decreased alpha activities 42 or increased beta activities 43 in the contralateral hemisphere following the ischemic brain injury. To this end, the IRDA observed in this study could be another form of transhemispheric diaschisis relating to epileptic activities.
In summary, the clinical post-traumatic and post-ischemic NCS and PED were replicated in two animal models of penetrating and focal ischemic brain injuries, which made possible a detailed analysis of the similarities and differences of these events for both etiologies. Although the intrinsic EEG variations of these phenomena were indistinguishable between the two types of brain injuries, their unique temporal profiles could serve as a valuable index for determining injury-specific treatment strategies. The identification of differential correlations between these epileptic events and the histopathological outcomes of the respective injuries may also help us to improve our understanding of the underlying mechanisms of these events beyond their seemingly similar manifestations.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Material has been reviewed by the Walter Reed Army Institute of Research. There is no objection to its presentation and/or publication. The opinions or assertions contained herein are the private views of the author, and are not to be construed as official, or as reflecting true views of the Department of the Army or the Department of Defense. Research was conducted in compliance with the Animal Welfare Act and other federal statutes and regulations relating to animals and experiments involving animals and adheres to principles stated in the Guide for the Care and Use of Laboratory Animals, NRC Publication, 2011 edition.