
Abstract
Whole-body vibration (WBV) is a relatively novel form of exercise used to improve neuromuscular performance in healthy individuals. Its usefulness as a therapy for patients with neurological disorders, in particular spinal cord injury (SCI), has received little attention in clinical settings and, surprisingly, even less in animal SCI models. We performed severe compression SCI at a low-thoracic level in Wistar rats followed by daily WBV starting 7 (10 rats) or 14 (10 rats) days after injury (WBV7 and WBV14, respectively) and continued over a 12-week post-injury period. Rats with SCI but no WBV training (sham, 10 rats) and intact animals (10 rats) served as controls. Compared to sham-treated rats, WBV did not improve BBB score, plantar stepping, or ladder stepping during the 12-week period. Accordingly, WBV did not significantly alter plantar H-reflex, lesion volume, serotonergic input to the lumbar spinal cord, nor cholinergic or glutamatergic inputs to lumbar motoneurons at 12 weeks after SCI. However, compared to sham, WBV14, but not WBV7, significantly improved body weight support (rump-height index) during overground locomotion and overall recovery between 6–12 weeks and also restored the density of synaptic terminals in the lumbar spinal cord at 12 weeks. Most remarkably, WBV14 led to a significant improvement of bladder function at 6–12 weeks after injury. These findings provide the first evidence for functional benefits of WBV in an animal SCI model and warrant further preclinical investigations to determine mechanisms underpinning this noninvasive, inexpensive, and easily delivered potential rehabilitation therapy for SCI.
Introduction
S
Whereas large injuries often lead to severe permanent disabilities, smaller lesions are followed by some degree of spontaneous functional recovery, 3 –5 which can be enhanced by rehabilitative therapies. The benefits of combined locomotor training with pharmacological means, and/or spinal cord electrical stimulation paradigms, on the recovery of sensory and locomotor function after SCI have been demonstrated in animal models 6,7 and SCI patients, 8,9 but the underlying mechanisms are still poorly understood. 10
Focal vibration following SCI has been successfully used in humans to treat sexual and bladder dysfunction 11 –13 and, more recently, has been shown to elicit involuntary step-like behavior in people with complete and incomplete SCI 14 as well as to decrease spasticity. 15
Whole-body vibration (WBV) has attracted increasing attention as a potential therapy during the past decade. Many WBV studies aim to improve some aspect of neuromuscular performance or to increase bone mass or density (see Refs. 16 and 17 for recent review). WBV delivers mechanical stimuli that are conveyed to the entire human body via the feet when standing on a vertically oscillating platform. Nowadays, vibrating platforms are commercially available and typically utilize low-frequency and low-amplitude vibration, thereby delivering strong stimuli to the skeletal muscles of the lower body. 18 WBV is being extensively used to enhance sports training 19,20 by improving muscle power and strength, 21,22 and force-velocity relation 23 with positive training effects being achieved during a relatively short time period. 24,25 Recently, WBV therapy has been reported to improve blood flow velocity and neuromuscular activity in Friedreich's ataxia. 26
After spinal cord injury in humans, WBV has been shown to reduce muscle spasticity and improve spinal reflex modulation, 27,28 walking function, 29 as well as increase bone mineral density. 30 Surprisingly, we identified only one experimental basic research report that examined vibration-induced stepping in spinal cats. 31
The purpose of this study therefore was to determine whether WBV over a period of 12 weeks could improve outcomes after incomplete compressive SCI in rat using behavioral, electrophysiological, and anatomical measures of locomotor and bladder function.
Methods
Animals
A total of 40 adult (175–200 g) female Wistar rats (strain HsdCpb:WU, Harlan) were used. Ten animals were used as intact controls and the remaining 30 were randomized into three groups, each consisting of 10 rats. Animals were fed standard laboratory food (Ssniff, Soest, Germany), provided tap water ad libitum and kept in a conditioned animal room (23°C, 12-hour artificial light-dark cycle). All experiments were conducted in accordance with the German Law on the Protection of Animals, and procedures were approved by the local Animal Care Committee.
Conditioning
All 40 rats were daily trained to walk on a wooden beam and to climb an inclined ladder for 2 weeks.
Those animals who were designated to receive WBV after SCI were additionally trained to remain still in the rat Galileo chamber for 2 min at a WBV of 15 Hz and for 1 min at 30 Hz daily for 2 weeks. Low frequencies were selected, because these vibration trials were well tolerated by the rats. Animals rapidly became accustomed to this procedure within 2–3 days and did not show any signs of stress, such a freezing or trying to bite, weight loss, or lack of grooming; rather, animals readily cooperated.
Spinal cord injury
Severe spinal cord compression followed a technique 32 that we have previously established in our lab. 33,34 Rats were anesthetized by 1.8 vol % isofluorane (Forene, Abbott, Germany), 0.6 L/min O2 (Conoxia, Linde, Germany), and 1.2 L/min N2O (Niontix, Linde, Germany), and laminectomy was performed at T8 under a surgical microscope. The exposed spinal cord was compressed using electromagnetically controlled watchmaker forceps (Dumont #5, Fine Science Tool, Heidelberg, Germany). The degree of forceps closure was 50% of spinal cord diameter, which produces a severe contusion SCI in adult rats. 34
The forceps and electromagnetic drive were mounted on a metal block attached to a stereotaxic frame, allowing precise positioning of the forceps tips either side of the spinal cord adjacent to the dura mater. A vernier-calibrated screw was used to advance the closing pin so as to bring the moving arm of the forceps next to the spinal cord and thus measure its diameter prior to injury (i.e., the distance between the forceps tips; see Figure 1 in Apostolova et al. 35 and in Semler et al. 34 ) The degree of forceps closure (i.e., compression: severe=50% of spinal cord diameter) was controlled by another vernier-calibrated limiting pin passing through a drilled hole in the stationary arm of the forceps. The spinal cord was compressed by the moving arm of the forceps (contusion velocity was 100 mm/sec) for 1 s using a timed current delivered to the electromagnetic drive. The skin was closed using 6-0 nylon sutures (Ethicon, Norderstedt, Germany).

Setup of the rat Galileo-device for whole-body vibration (WBV) training. During conditioning
Operated rats were kept at 37°C for 12 h to prevent hypothermia. Afterwards, they were housed individually in standard cages and the bladder was voided manually three times a day. The three groups comprised: sham (no therapy, N=10), WBV7 (begin WBV-therapy on day 7 after SCI, N=10), and WBV14 (begin WBV-therapy on day 14 after SCI; N=10). Postoperative animal losses were minimal, with final numbers at 12 weeks being 8 in the sham and WBV7 groups and 9 in the WBV14 group.
Whole-body vibration
We used a custom “Galileo-device” designed specifically for rats (Fig. 1A) by Novotec Medical GmbH (Pforzheim, Germany), an established designer and manufacturer of WBV equipment for human patients. To reduce stress imposed on the animals by spatial restriction, the dimensions of the chamber were selected by the company so that animals entered it and stood in it voluntarily (i.e., without unwillingness or resistance). Once within the chamber, rats could move around and even turn around; thus, apart from lying along the long axis of the chamber, there was no standardized posture required of the rats. Nevertheless, the vibrating floor of the chamber is constructed (see below) so that potential small differences in posture (e.g., head to the right or head to the left) would not influence the delivery of vibration to the whole body.
The chamber had a vibrating floor that consisted of two plates that were moved by electric motors on both sides (left and right). The floor plates moved in an alternating up and down fashion. Thus, their movements were in opposite directions, with one descending while the other ascended and were also out of phase by 180° so that the travel of each plate reached its maximum (and opposite) simultaneously with the other one. Thus, when, for most of the time during a training session, the animal's body is positioned along the long axis of the chamber, each vibration cycle starts with an upward movement of, for example, the forelimbs accompanied by downward movement of the hindlimbs and then followed by reversal of the movement directions in the second phase of the cycle. However, because the animals were free to move, thereby reducing stress, the design did not always ensure that the left-sided hindlimbs were on one plate while the right-sided hindlimbs were on the other. As a result, the term “whole body vibration” is used to describe the procedure. However, this does not imply that muscle and body vibration was achieved exclusively via weight-bearing extremities.
The movements of the floor plates caused alternating rocking-like vibrations to the rat's whole body with a maximum amplitude of 1.5 mm and at frequency of 15 or 30 Hz. The acceleration can be calculated using the formula:
where a=acceleration [m/s2], f=frequency [1/s], and A=amplitude [m].
In the experiment, we used the same amplitude throughout (1.5 mm) but with two different frequencies (15 Hz and 30 Hz). When using 15 Hz, the platform acceleration was 13.3 m/s2 (1.45 g); at 30 Hz, acceleration was 53.3 m/s2 (5.8 g). These accelerations produce forces, depending mainly on amplitude of movement and tissue stiffness, which do not exceed forces during normal weight-bearing locomotion. In addition, the low frequencies used were selected because these vibrations were well tolerated by the rats (see above, section “Conditioning”) and the frequencies are within the normal range of motoneuron discharges.
By displacing the rostral to the caudal portion of the rat body and vice versa (but not the left-sided hindlimbs to the right-sided ones), this rocking movement mimicked the normal walking pattern of healthy intact animals (i.e., when a forelimb flexes and extends before stepping, the contralateral hind paw remains on the ground). Thus, no special precautions ensuring that the hindlimbs were on different plates (left or right) were necessary.
Alternating, out-of-phase displacements of the rostral portion of the rat body with respect to the caudal portion (but simultaneous, in-phase movements of the fore-, respectively, hindlimbs) enable rocking movements that are unique for the Galileo-device, as all commercially available vibrating devices produce simultaneous up/down (jump-like) oscillations of the whole body using one platform. However, the importance of the specific design of the Galileo device cannot currently be evaluated since there are no other WBV rodent studies using a single vibrating plate and comparisons with otherwise designed apparatus was beyond the scope of this investigation.
Starting at 1 or 2 weeks after SCI, rats were subjected to WBV once daily for 5 days/week for 12 weeks with the amplitude, frequency and duration being controlled electronically (Fig. 1A). Every WBV session comprised five sequential trials, each lasting for 3 min. Each trial consisted of 1-min 15-Hz vibration followed by and 2 min at 30 Hz.
Locomotor rating
Locomotor functions were evaluated using the Basso, Beattie, Bresnahan (BBB) rating scale. 36 Scoring was undertaken by two independent investigators (S. Angelova and F. Wirth) blinded to the treatment (Sham, WBV7, WBV14) groups and using video recordings of beam walking (see below) observed at slow playback speed. 34,35 Assessment was performed prior to injury (0 weeks) and 1, 3, 6, 9, and 12 weeks after SCI (Fig. 2).

Time course and degree of functional recovery in rats with spinal cord injury (SCI) subjected to WBV from day 7 or 14 on (WBV7 and WBV14, N=8 and 9, respectively) or no WBV (Sham, N=8). Shown are mean values±standard errors of mean (SEM) of locomotor rating scores (Basso, Beattie, and Bresnahan [BBB]
Single-frame motion analysis (SFMA)
Locomotor tests were performed as described
34,35
using a walking platform (“beam,” Supplementary Fig. 1A; see online supplementary material at
Beam walking
The runway bar was made of a wooden plate (1500 mm long, 120 mm wide, and 20 mm thick) fixed on 200-mm high feet (Supplementary Fig. 1A). Prior to SCI, rats were video-recorded from the left and right side during beam walking using a Panasonic NV-DS12 at 25 frames per sec. The video recordings were repeated 1, 3, 6, 9, and 12 weeks after SCI. Animals were not trained to walk between recording sessions. Per time point and animal, at least three walking trials were recorded per side (left- and right-side views). The video sequences were examined using VirtualDub 1.6.19, a video capture/processing utility written by Avery Lee (free software available at
The FSA is the angle at which the hind paw is placed on the ground at the beginning of the stance phase. The angle is defined by a line parallel to the dorsal surface of the paw and the horizontal line and is measured using video frames in which the paw is seen in initial firm contact with the ground (Supplementary Fig. 1B). In intact rats, the stance phase is well defined and the angle smaller than 20° (Supplementary Fig. 1B). After spinal cord injury, rats drag their hind limbs with dorsal paw surfaces facing the beam surface and the angle increases to 130°–140° (Supplementary Fig. 1C). In severely disabled rats, “step cycles” were defined by the movements of the forelimbs. Video frames in which the angle appeared to have its lowest values for individual “cycles”, typically following a visible attempt to flex the extremity, were used for measurements. In less severely disabled rats which performed stepping of variable quality (dorsal or plantar), the angle was measured upon dorsal or ventral placement of the paw on the ground after a swing phase or after a forward sweep of the paw over the beam surface. The FSA has been defined as an objective measure of stepping (plantar or dorsal), a major attribute in the BBB score. Per time point of observation, three to six measurements were made for each hindlimb (i.e., 6 to 12 “step cycles”), defined according to the stepping ability of the animal as described above, were analyzed per animal. The values for the left and right paw of each animal were averaged (Fig. 2B).
The RHI is defined as height of the rump (“a” in Supplementary Fig. 1B, C), that is, the vertical distance from the dorsal aspect of the animal's tail base to the beam, normalized to the thickness of the beam measured along the same vertical line (“b” in Supplementary Fig. 1B, C). For each animal and trial, the maximum rump height index was estimated using video frames of three to six “step cycles”. The RHI is a numerical estimate of the ability to support body weight, an attribute in the BBB rating scale (Fig. 2C).
Ladder climbing test
After the beam-walking test, rats were video recorded during climbing up an inclined ladder, as described previously for spinal cord injured mice. 35 The rat ladder (Supplementary Fig. 2A) is made of a plastic frame (1280 mm long, 190 mm wide, with central incision of 1200×120 mm) and has 19 round wooden rungs (6 mm in diameter) at equal intervals (60 mm) and 150-mm high side-walls. The ladder is fixed in an inclined position (55°) using a plastic platform and metal feet. The rats were placed at the bottom rungs of the ladder and climbing was video recorded from a position “below” the ladder, viewing the ventral aspect of the animals (Supplementary Fig. 2B–F). After initial training (conditioning), rats learned to climb up to the top of the ladder and even severely disabled animals after SCI could do this using their forelimbs and the body weight support provided by the inclined position of the ladder. The video recordings were observed at slow-speed playback and the CLS (correct placing of the hind paw and sustained position until the next forward move, Supplementary Fig. 2D, E) over 16 rungs were counted. The ladder climbing analyses were performed by one investigator (K. Wellmann or F. Wirth) blinded to the treatment (Sham, WBV7, WBV14) groups (Fig. 2D).
Recovery indices
The recovery index
35,37
is an individual estimate of recovery for any given parameter and is calculated in percent as:
where Y0, Y1 and Y1+n are values prior to operation, 1 week after injury, and a time point n weeks after the first week, respectively. This measure estimates gain of function (Y1+n – Y1) as a fraction of the functional loss (Y0 – Y1) induced by the injury. The RI may have 0 or negative values if no improvement or further impairment occurs during the observation time period. The only situation where the index cannot be calculated is if the injury causes no impairment [i.e., when (Y0 – Y1) is 0]. Overall recovery indices can be calculated, on an individual animal basis, as means of recovery indices for individual parameters. The overall indices are more complex estimates of recovery than the individual parameters and represent “scores” for individual animal performance based on objective assessment of different aspects of locomotion (Fig. 2E).
Evaluation of bladder functional status
The functional status of the bladder in our experiments was assessed immediately prior to each manual voiding (i.e., three times daily, using the following scoring method: 38
completely empty (no urine could be expressed, 0 points)
half empty bladder containing several drops of clear urine (10 points)
wet perineum, filled bladder with clear urine (20 points)
half empty bladder with turbid urine (30 points)
wet perineum, full bladder with turbid urine (40 points)
half empty bladder with bloody urine (50 points)
wet perineum, full bladder with bloody urine (60 points)
As described previously, 39 a subset of data, collected mid-week (Wednesday) during every week of the experiment (surgery on a Wednesday) was presented in a graphical form (see below)
H-reflex recordings
We selected the plantar reflex for analysis. Previous work 40 has shown that after SCI the plantar reflex changes were similar to the reflexes elicited in other hindlimb muscles (e.g., the gastrocnemius and the anterior tibial muscles). Therefore, we assumed that the changes in the plantar H-reflex after spinal cord injury were characteristic for the hindlimb musculature as a whole.
Rats were anesthetized using the oxygen-nitrous oxide-isoflurane mixture as described above, but with the concentration of Isofluran reduced to 0.8–0.9 Vol %. The forelimbs and hindlimbs of the rat were fixed onto a plastic plate with tape. The hindlimbs were carefully positioned so as to avoid unnecessary pressure and stretch, which could affect the electrophysiological responses of muscles and nerves. To maintain body temperature, the plastic plate was placed on a warm water pad (37°C) driven by a temperature regulating pump system (TP472T/pump, Gaymar Industries, Orchard Park, NY, USA). The eyes of animals were protected from drying with an eye cream (Bepanthen, Bayer, Leverkusen, Germany).
Experiments were performed in a Faraday cage made of copper metal mesh. All conductible devices used in the cage were grounded. Tibial nerve stimulation was performed by means of two needle stimulating electrodes (stainless steel, diameter: 0.4 mm) placed direct on the medial aspect of the tibial nerve close to Achilles tendon (Supplementary Fig. 3A). The distance between the electrodes was about 0.5 cm (the cathode was red and always proximal; the anode was blue and always distal). A ground needle electrode (stainless steel, diameter: 0.4 mm, black in Supplementary Fig. 3B) was applied at the base of the animal's tail. For recording, a reference stainless steel electrode (green in Supplementary Fig. 3B) was fixed to the skin between the first and second digit and an active recording electrode (tungsten, diameter: 0.25 mm, yellow) was inserted between the second (intermediate) and third (lateral) cuneiform bones. Electrode positioning was performed under a stereo-microscope.
Measurements were performed using LabChart instrumentation (Isolated Stimulator, Chart und Scope View, ADInstruments, Spechbach, Germany) 12 weeks after SCI. The H-reflex was elicited by delivering bipolar electrical pulses of 0.2 ms duration to the tibial nerve close to the Achilles tendon. Stimulus intensity was gradually increased starting at 0.1 mA until both M- and H-waves with latencies of approximately 2 and 5 ms, respectively, were visible. After the threshold measurement, stimulus intensity was further increased until maximal and stable H-responses were elicited (suprathreshold intensity). Thereafter, stimulation continued at the defined suprathreshold level at frequencies of 0.1, 0.3, 0.5, 1, 2, 3, and 5 Hz. Ten consecutive responses were recorded at each frequency.
The amplitudes of M- and H-waves were measured in mV as peak-to-peak values, averaged (excluding the first response at each frequency) and used to calculate H/M ratios. The maximum M- and H-waves and H/M ratios were measured from recordings at baseline stimulation rate (0.1 Hz, Fig. 3A) and used as estimates of the H-reflex excitability. 41 The M- and H-wave amplitudes at higher frequencies (0.3 to 5 Hz) were normalized to the respective amplitudes at baseline frequency (set to 100%) and used to estimate the rate-dependant depression of the H-reflex (Fig. 3B), a parameter providing information about the intraspinal modulation of the H-reflex. 41 The latencies of the responses were measured as time in seconds elapsed between trigger and peak of each waveform. Data were analyzed using the LabChart software.
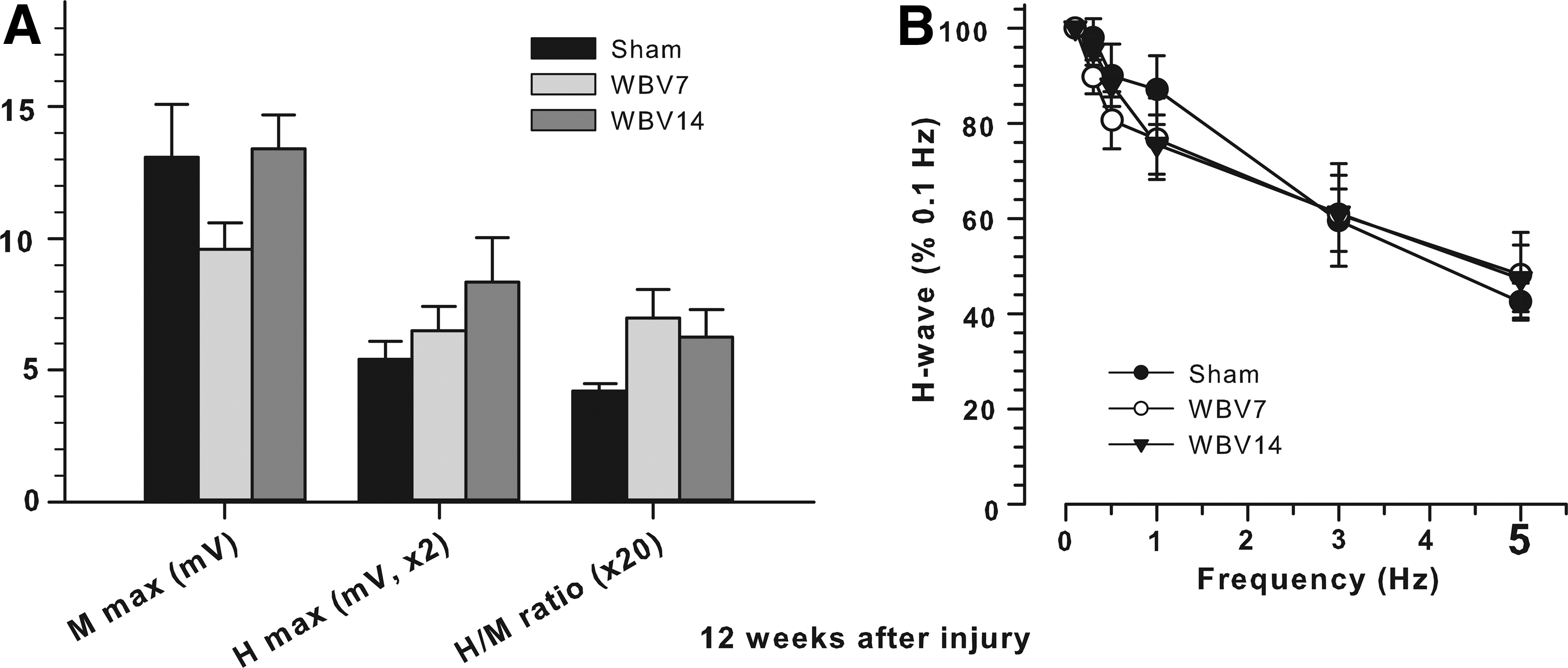
Plantar H-reflex 12 weeks after SCI.
Spinal cord lesion volume
Twelve weeks after SCI, rats were deeply anesthetized with ether and perfused transcardially with phosphate buffered saline (PBS) for 30 sec, followed by 4% formaldehyde in 0.1 M phosphate buffered saline (PBS), pH 7.4, for 20 min. The spinal cord between T7–T9 was dissected free. Following cryoprotection (20% sucrose in PBS), longitudinal sections (25 μm) were cut on a cryostat and mounted on SuperFrost Plus slides (Roth, Germany).
A fractionator sampling strategy 42 was used and each fifth longitudinal section (a total of at least 10 equidistant sections through the spinal cord) was stained with cresyl violet (Nissl) stain; Fig. 4A). A Zeiss microscope equipped with a CCD Video Camera System (Optronics Engineering Model DEI-470, Goleta, CA, supplied by Visitron Systems, Puchheim, Germany) combined with Image-Pro Plus 6.2 software (Media Cybernetics, Silver Spring, MD, USA) was used to quantify the areas (in μm2) of the lesions in each tissue section at a primary magnification of×2.5. The total lesion volume was calculated using the Cavalieri method. 42 –44 Measurements were performed by two observers (J. Ankerne and M. Ashrafi) blinded to the treatment (Sham, WBV7, WBV14) groups.
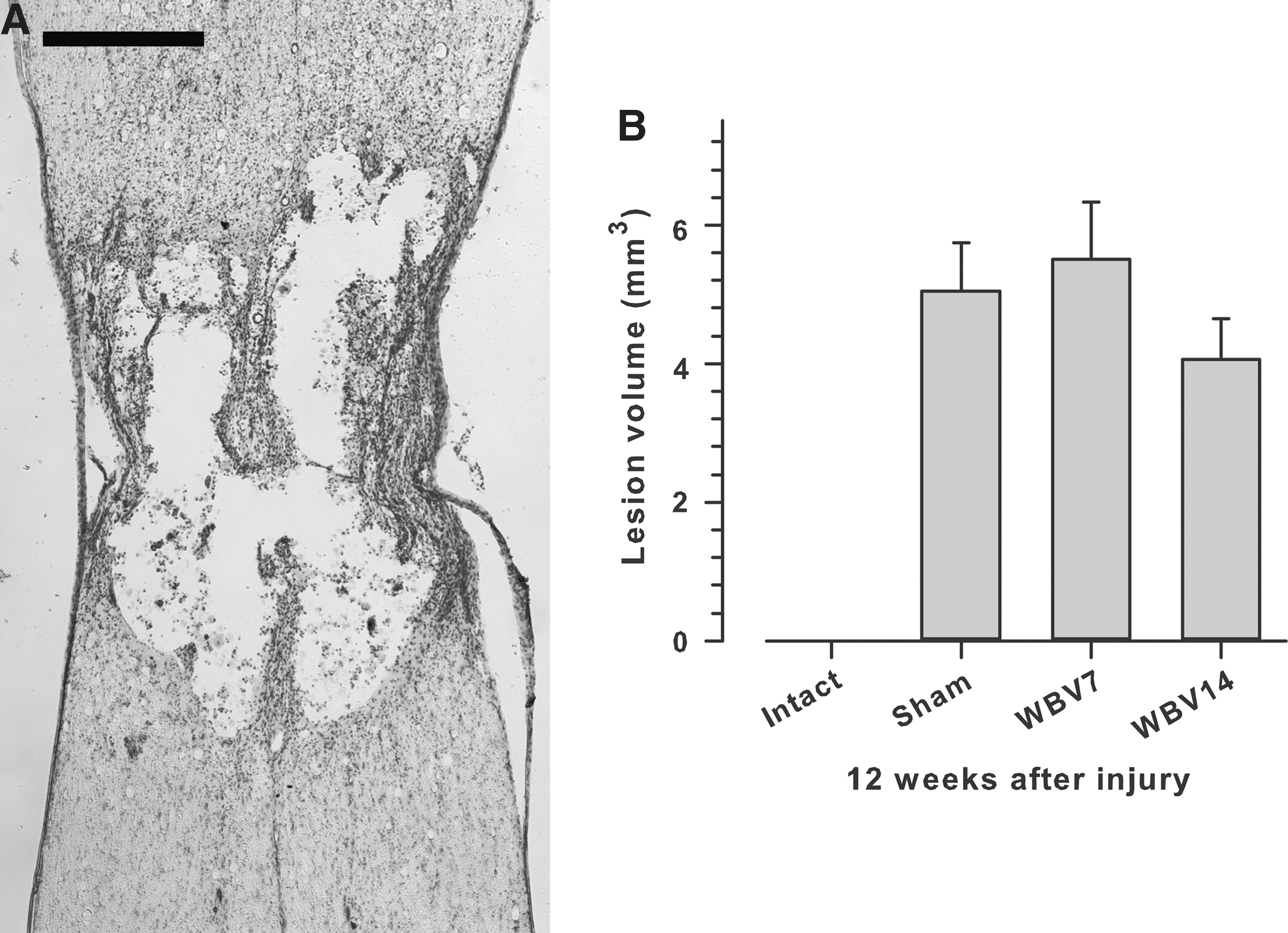
Lesion volume 12 weeks after SCI. Lesion volume was estimated from spaced-serial sections using the Cavalieri principle.
Immunohistochemistry and evaluation of axonal and synaptic terminal densities in the spinal cord
After perfusion fixation (see above), the second lumbar (L2) segment of the spinal cord was harvested (Supplementary Fig. 4) cryoprotected in 20% sucrose in PBS overnight and cut in 25 μm thick transverse sections.
For immunohistochemistry, the following commercially available antibodies were used at optimal dilutions: (1) goat anti-choline acetyltransferase antibody (ChAT) (1:100; Chemicon, Hofheim, Germany); (2) mouse anti-vesicular glutamate transporter 1 (VGLUT1) (1:500; Synaptic Systems); (3) rabbit anti-serotonin (5-HT) (1:400; ImmunoStar Inc., Hudson, WI, USA); (4) rabbit anti-synaptophysin 1 (1:200; Synaptic Systems, Göttingen, Germany).
The staining protocol has been described previously.
37
Water-bath antigen demasking was performed in 0.01 M sodium citrate solution, pH 9.0, for 30 min at 80°C for all antigens.
45
Nonspecific binding was blocked using 5% v/v normal serum from the species in which the secondary antibody was produced, dissolved in PBS, and supplemented with 0.2% v/v Triton X-100 and 0.02% w/v sodium azide for 1 h at room temperature (RT). Incubation with the primary antibody, diluted in PBS containing 0.5% w/v λ-carrageenan (Sigma) and 0.02% w/v sodium azide, was performed for 3 days at 4°C. After washing in PBS (three times for 15 min at RT), the appropriate secondary antibody diluted 1:400 in PBS–carrageenan solution was applied for 2 h at RT. The following cyanine 3 (Cy3)-conjugated secondary antibodies were used: goat anti-rabbit, goat anti-mouse, donkey anti-goat [Jackson ImmunoResearch (West Grove, PA) via Dianova, Hamburg, Germany]. After a subsequent wash in PBS, cell nuclei were stained for 10 min at RT with NeuroTrace 500/525 (1:200; Molecular Probes, N21480, via Life Technologies GmbH
All analyses were performed blinded to the treatment (Sham, WBV7, WBV14) groups.
Motoneuron soma size and quantification of perisomatic puncta
Analyses were performed using the Stereo Investigator 8.1 software (MicroBrightField Europe, Magdeburg, Germany) and a fluorescence microscope (Axioskop 2 mot plus, Zeiss, Oberkochen, Germany) equipped with a motorized stage (Zeiss) and a CX 9000 digital camera (MicroBrightField).
All ChAT-immunostained motoneurons with discernible nuclei were analyzed in every tenth transverse section of the spinal cord. Each motoneuron, visualized at 100x magnification, was focused at the level of its largest cell body cross-sectional area and soma area, perimeter, and number of perisomatic terminals were measured using the Stereo Investigator software. Frequency of perisomatic ChAT+ terminals was calculated as number of perisomatic terminals per unit perimeter length. Mean values of individual animals were used to calculate group mean values.
Fluorescent image analysis of the overall amount of serotonergic fibres and synaptic terminals in the ventral horn
Fluorescent images of 5-HT-, VGLUT1-, and synaptophysin-stained sections were analyzed for pixel brightness as described by previously. 46,47 Fluorescent images were compared using the 8 BPP gray scale format. Thereby each pixel contains 8 bits of information encoding brightness, which ranges in a scale from 0 to 255. The scale for pixel brightness, or pixel gray value, is constructed so that the higher numbers indicate greater pixel brightness. Digital images were captured with a slow scan CCD camera (Spot RT, Diagnostic Instruments) as described previously. 46,47 For quantification of pixel brightness, images were captured using a 16X objective and the Image-Pro Plus Software Version 6.0 (Media Cybernetics, Inc., Silver Spring, MD, USA). Exposure time was optimized to ensure that only a few pixels were saturated at 255 gray value. All images were taken under same conditions of exposure. Interactive threshold was used to detect the pixel brightness of the minimum fluorescence. Threshold values ensured the inclusion of the entire signal range in the sample. This threshold value was further used to extract and compare the pixel number between all animals of the same group and between experimental groups.
Serotonin (5-HT)
Since at a gray value over 205 just a few pixels representing labeling were present (Fig. 5A, B), only pixel gray scale ranges with the maximum change in fluorescence were considered. This threshold was set to a pixel gray value of 220. Accordingly, the range for measuring pixel number and brightness was set at 121–220.
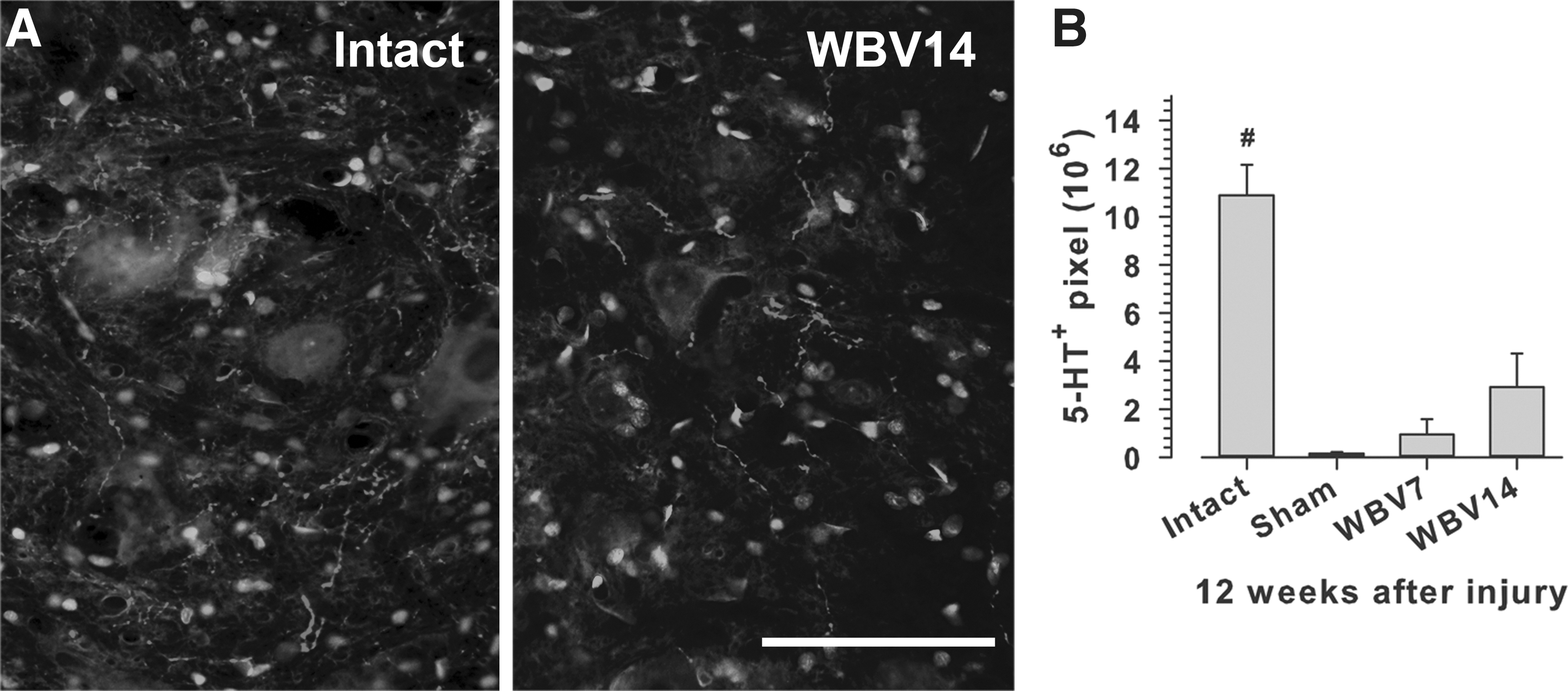
5-HT axon densities in the lumbar spinal cord 12 weeks after SCI.
ChAT
The frequency of perisomatic ChAT+terminals was calculated as number of perisomatic terminals per unit perimeter length. Mean values of individual animals were used to calculate group mean values (Fig. 6).

Frequencies of cholinergic perisomatic terminals and soma areas of lumbar motoneurons 12 week after SCI.
VGLUT1
The background intensities were identical from image to image around a pixel gray value of 105 (Fig. 7A). We therefore selected a threshold level for measuring pixel number and brightness of 107 and a range of 107–156.

Densities of VGLUT1-positive synaptic terminals in the lumbar spinal cord 12 weeks after SCI.
Synaptophysin
The background intensity had a pixel gray value of 130 (Fig. 8A). Accordingly, we set the threshold level at 135 and the range for measuring pixel number and brightness at 136–185.

Synaptic terminals density in the lumbar spinal cord at 12 weeks after SCI.
Bladder weight and histology
After fixation (see above), bladders were carefully dissected free under an operating microscope, paying special attention to remove the entire “neck” which contains the urethral sphincter muscle. If the bladder was full, it was voided after dissection. Bladders were rinsed in buffer, suspended until almost dry, 48 photographed (Fig. 9A), and weighed on a precision electronic balance Sartorius CP423S-OCE (Sartorius AG, 37075 Göttingen, Germany).

Bladder function and bladder innervation.
Histopathology of the bladder wall
After weighing, bladders were cut in half along the mid-sagittal plane and cryoprotected (20% sucrose in PBS). Sagittal sections (25 μm) encompassing the entire bladder, that is, the neck, ventral and dorsal walls, as well as the fundus were cut on a cryostat and mounted on SuperFrost Plus slides (Roth, Germany). In intact rats, one half of the bladder comprised approximately 60 sections; after SCI, the number of sections was ∼100. Employing the fractionator section selection strategy, 42 equidistant sections (every sixth in the intact rats and every tenth in animals with SCI (i.e., ∼10% of sections in both groups) were stained with the polyclonal anti-neuronal class III β-tubulin, which detects all axons (Covance, Richmond, CA, USA, No. PRB-435P) and anti-rabbit IgG Cy3 conjugate. Staining controls (omission of the primary antibody or of the secondary biotinylated antibody) yielded blank (dark) sections. All sections used to compare the intensity of fluorescence between intact and lesioned rats were incubated simultaneously using identical solutions. Sections were photographed using a Zeiss Axioskop 50 epifluorescence microscope equipped with a “rhodamine” filter (excitation BP 546/12, beamsplitter FT 580, emission LP 590) at 1.25X and at 25X magnification (Fig. 9B).
Fluorescence images were compared using an 8 bits per pixel (BPP) gray scale format in which each pixel contains 8 bits of information encoding brightness, which ranges from 0 to 255. The scale for pixel brightness, or pixel gray value, is such that higher numbers indicate greater pixel brightness. Threshold values ensured inclusion of the entire signal range in the sample. Since at a gray value over 140, only a few pixels representing labeling were detectable, only pixel gray scale ranges with the maximum change in fluorescence were considered. This threshold was set to a pixel gray value of 150 for tubulin. Accordingly, the range for measuring pixel number and brightness was set at 102–201.
A large variation in the dimensions of sagittal bladder sections among different animal groups necessitated objective selection of a region of interest (ROI) for analysis that encompassed as large as possible area of the ventral wall which was selected for the following reasons. Acetylcholinesterase staining in the intact bladder has previously shown that postganglionic nerves from the pelvic ganglion reach the point of entry of the ureter as several nerve trunks on each side, slightly ventral and caudal to the ureteral orifices, and branch within the bladder wall, extending to all the regions of the musculature. 49
These nerves run in the ventral longitudinal muscle layer as well-defined trunks with a pattern that is consistent between animals. 50 The ventral wall was also easily detectable by numerous mucosal foldings. After software calibration (Image-Pro Plus, Version 6.2.), we determined the size of the ventral bladder wall in μm2. Fluorescence intensity in each ROI (in pixels) was then calculated per μm2.
We used a fractionator sampling strategy (see above) to analyze at least ten equidistant sections through the bladder. Using Image-Pro Plus 6.2, the projection areas (in μm 2 ) containing Cy3-positive axons were quantified in each tissue section. Measurements were performed by two observers (O. Ozsoy and U. Ozsoy) blinded to the treatment.
Statistical analysis
Statistical analyses were performed using the Sigma Plot 11 software (SPSS, Chicago, IL, USA). Data were analyzed for distribution and equal variance and appropriate parametric tests were used. The threshold value for acceptance of differences was 5%. Details of the statistical methods used are given in the text and figure legends. Data are presented as mean values with standard errors of mean (SEM).
Results
Delayed WBV training improves body weight support in spinal-injured rats
We first analyzed locomotor functions in the spinal-injured rats using the BBB rating scale. At 1 week after SCI, before initiation of WBV, the mean BBB scores in the three experimental groups (sham, WBV7 and WBV14, see below) were similar (1.2±0.29, 1.6±0.49, and 1.4±0.34, respectively, p=0.73, one-way ANOVA), indicating that the groups were equivalent in terms of lesion severity. These BBB scores (range 0–4) show that the spinal cord compression had produced, as intended, a severe injury with complete hindlimb paralysis or only slightly movements in 1–3 joints in all groups. During the following weeks, a statistically significant improvement was observed in rats placed daily in the vibration apparatus but not subjected to vibration (sham group): at 12 weeks, the average BBB score reached 6.6 (range 2–10, Fig. 2A). Only 3 out of 8 animals could occasionally perform plantar stepping (score 10) meaning that, as a whole, this group did not regain functionally meaningful walking abilities. Rats with WBV initiated at 7 days after SCI (WBV7 group) showed a similar degree of recovery to sham rats at 3 to 12 weeks after SCI (Fig. 2A). A trend for better recovery at 3–12 weeks was seen in rats with WBV initiated at 14 days post-injury (WBV14 group, Fig. 2A). Although repeated measures ANOVA did not reveal a significant overall effect of treatment on BBB scores, it should be mentioned that at 12 weeks four out of nine WBV14 rats reached scores between 13 and 16, meaning that these animals were capable of consistent plantar stepping and forelimb-hindlimb coordination, that is, the rats could walk even if specific “fine” aspects of locomotion such as toe-clearance, paw and tail position, and trunk stability were abnormal. In contrast, none out of eight sham-operated rats reached a score >10 at 12 weeks (p=0.08 versus the WBV14 group, two-tailed Fisher's exact test).
Analysis of stepping using the foot-stepping angle (FSA) revealed, similar to the BBB scores, similar degrees of impairment in the three groups 1 week after SCI (Fig. 2B). In the sham group, the angle changed from an average of about 20° prior to injury to 130° at 1 week and only gradually and slightly decreased thereafter to an average of 93° at 12 weeks. Angles larger than 90° indicate that the toes or the dorsal paw surface rather than the plantar surface touch the ground during ground locomotion. Thus, the results show that the sham rats did not recover plantar stepping within the 12-week observation period. The results for the WBV7 group were similar to sham animals with a mean FSA of about 100° at 12 weeks (Fig. 2B). As for BBB scores, we observed a trend for better recovery of plantar stepping in WBV14 rats at 3–12 weeks (p=0.10–0.12 compared with sham rats, t test, Fig. 2B). At 12 weeks, the mean FBA reached 46°, indicating transition to plantar stepping.
The second SFMA parameter used for functional assessment, the RHI, estimates the ability of the hindlimbs to support body weight during ground locomotion. This ability was significantly impaired 1 week after SCI in all animal groups (Fig. 2C). Rats moved along the beam using the forelimbs while the hindlimbs were dragged behind and could not elevate the trunk above the beam surface. In the following weeks, the RHI improved moderately, and to the same degree, in the sham and WBV7 groups. At 12 weeks, the rump could be lifted to about one-fourth of the preoperative rump height. Compared with these two groups, higher RHI was found in WBV14 rats at 6–12 weeks (Fig. 2C). At 12 weeks, the WBV14 rats were able to lift their trunk on average to more than half the normal rump height (p<0.05).
The third numerical parameter analyzed was the number of correct ladder steps during inclined ladder climbing. Proper placement of the paws and maintenance of a stable position of the paw on the rungs to support body weight during climbing require higher levels of motor and sensory control than overground locomotion. One week after SCI, all animals were unable to step on the rungs and the improvement with time after injury was minimal, reaching on average one correct step in the sham and WBV7 groups and two correct steps in the WBV14 group 12 weeks after SCI (Fig. 2D).
We also analyzed functional recovery using normalized value recovery indices (RI). The recovery index is an individual measure normalizing “gain” of function after the first week to “loss” of function between 0 and 1 week for each parameter. The recovery indices allow also calculations of more complex measures, overall indices based on several numerical parameters. Here we calculated overall recovery indices using the data for the foot-stepping angle, rump-height index and number of correct steps. The results showed that the overall recovery reached a maximum of about 25% in the sham and WBV7 groups at 12 weeks after injury (Fig. 2E; p<0.05). At 6–12 weeks, recovery was superior in WBV14 rats, reaching a maximum of over 50% at 12 weeks (Fig. 2E; p<0.05).
From these observations, we conclude that initiation of WBV training 1 week after SCI does not affect, in a positive or negative way, recovery after severe compression SCI. However, WBV initiated 2 weeks after injury has an overall beneficial influence on the functional outcome with most pronounced effect on the ability to support body weight during ground locomotion.
WBV does not alter motoneuron excitability
Previous work showed that, in rodents, the plantar H-reflex is a useful tool to assess motoneuron excitability after SCI that correlates with severity of injury and locomotor outcome. 41,51 Specifically, recovery of better locomotor abilities after incomplete SCI is associated with enhanced H- but not M-wave and attenuated frequency dependent depression (rate depression) of the H-reflex. We analyzed the H-reflex at 12 weeks after SCI. Several variables were measured at baseline stimulation frequency (0.1 Hz): maximum M- and H-wave amplitudes and H/M ratios (Fig. 3A), as well as M- and H-wave latencies (data not shown). None of these parameters showed significant differences among the treatment groups. However, it should be mentioned that there was a trend (p=0.10, one-way ANOVA) for higher H/M values in WBV-treated rats compared with sham rats. In addition, we analyzed the alterations of the M- and H-waves upon incrementally increasing the stimulation frequency from 0.1 to 5 Hz. In contrast to the M-wave, which showed no frequency-dependant depression (not shown), a marked rate depression was found for the H-wave (Fig. 3B). This depression was, however, similar in the three experimental groups. These findings indicate that WBV did not influence the spinal reflex excitability as estimated here for the plantar motoneuron pool.
WBV does not lead to alterations in lesion volume
We next tested whether WBV affects the lesion volume in the thoracic spinal cord, a parameter known to affect the functional outcome of SCI. 52 At 12 weeks after injury, we did not find significant differences in lesion volume among the three experimental groups, sham, WBV7 and WBV14 although, we should mention, the mean volume in WBV14 rats was 20% lower than in sham animals (Fig. 4B).
No alterations in serotonergic innervation of the lumbar spinal cord after WBV
The serotonergic innervation of the lumbar spinal cord is a parameter known to affect the functional outcome of SCI. 52 Similar to lesion volume, the 5-HT innervation of the lumbar spinal cord, as estimated by gray value measurements of 5-HT immunofluorescence, revealed no significant differences among the SCI groups, despite a trend for higher mean values in rats with WBV, in particular WBV14 animals, compared with sham-treated rats (Fig. 5B). In all three SCI groups, the 5-HT values were lower than in intact rats, indicating that the compression SCI causes a dramatic decline in, but not complete loss of the serotonergic innervation of the lumbar spinal cord at 12 weeks post-injury. Overall, with respect to WBV, these findings are in agreement with the observed trends, but no significant effects of WBV on locomotor parameters like BBB scores and foot-base angle.
WBV does not influence cholinergic and glutamatergic inputs to lumbar motoneurons
The compression SCI model used here causes a significant loss of cholinergic C-terminals, 53,54 targeting the cell bodies of lumbar motoneurons and, in mice, the degree of this loss is correlated with locomotor impairment and spinal reflex excitability. 35,41,52 Here, we analyzed whether the frequency (number per unit length) of these terminals is also affected by the injury in rats and, in addition, by WBV (Fig. 6A). Similar to mice, we found a large decline (-36%) in the frequency of C-terminals in sham rats compared with intact animals at 12 weeks after injury (Fig. 6B). However, no effect of WBV on terminal frequency was observed (Fig. 6B). The analyses of cholinergic terminal frequency also produced data on motoneuron profile area, which showed no differences among the SCI rats and intact animals (i.e., there was no effect of SCI or WBV on motoneuron size) (Fig. 6C).
We also analyzed a major glutamatergic input to spinal motoneurons, namely VGLUT1-positive terminals in lamina IX of the lumbar spinal cord (Fig. 7A). These synaptic terminals belong to medium- to large-sized neurons in the dorsal root ganglia that convey mechano- and proprioceptive information to the spinal cord 55 and are crucial for functional recovery of hind limb locomotion after SCI. 56 We found a remarkable loss of VGLUT1-positive terminals, as indicated by gray level measurements in the lumbar spinal cords of spinal cord-injured rats compared with intact animals but no effect of WBV (Fig. 7B). Similar results were obtained for a second target of VGLUT1-positive terminals, the Clarke's column, which convey proprioceptive information along the spinocerebellar tract (Fig. 7C). The overall conclusion from these results is that WBV does not induce alterations in two major inputs to spinal motoneurons, modulatory cholinergic terminals, and excitatory primary afferents, which are essential for locomotor recovery after SCI. This failure may explain the lack of pronounced treatment effects on specific locomotor parameters like stepping ability and BBB scores, and spinal reflex excitability.
WBV14 restored synaptic input in the ventral horn
Since we found no alterations in the synaptic coverage of the motoneuronal somata (by cholinergic terminals; see above), we decided to determine the overall amount of synapses in the ventral horn. We immunostained equidistant transverse sections through the lumbar enlargement for synaptophysin (an integral membrane glycoprotein of the presynaptic vesicles) and visualized the reaction product with Cy3 conjugated anti-rabbit IgG (Fig. 8A).
Analyzing the intensity of Cy3-fluorescence according to a gray scale, we found that, at 12 weeks after SCI, the ventral horn of the animals subjected to WBV14 had significantly more pixels in the selected range (136–185) than the sham and the WBV7 groups. Though smaller, the amount of pixels did not differ significantly from the value obtained for intact animals (Fig. 8B).
We conclude that WBV14 restored the total amount of synapses in the ventral horn.
WBV does not counteract bladder hypertrophy but improves bladder function
In addition to locomotor recovery, we analyzed changes in bladder function during the observation time period using a rating scale. Prior to operation, the score was zero, indicating that the bladders were normally emptied (i.e., normally functioning). At one week after SCI, all injured rats were unable to void the bladder spontaneously and the average scores increased to over 40 in the three SCI groups (43±1.6, 54±3.2, and 44±5.3 in sham-, WBV7-, and WBV14-treated rats, respectively; no significant differences, two-way repeated measures ANOVA with Holm-Sidak post-hoc tests, Fig. 9C). Thereafter, in sham and WBV7 rats, bladder function showed no improvement throughout the 12-week observation period (Fig. 9C). In contrast, we found a recovery of function between the first and the sixth week in the WBV14 group with mean scores significantly better than in sham and WBV7 rats at 6 to 12 weeks (Fig. 9C). Macroscopical observations and weight measurements at 12 weeks after injury revealed severe bladder hypertrophy in all SCI groups compared with intact rats (Fig. 9A, D). There were no statistical differences in bladder weight among the SCI groups (Fig. 9D). Despite a significant functional improvement, there was only a trend for smaller bladder mass in the WBV14 group (-17% compared with sham rats; Fig. 9D). In search of an explanation for improved function in WBV14 rats, we analyzed axonal density at 12 weeks after injury using beta III-tubulin staining of axons in the bladder wall (Fig. 9B). We found no statistical differences among the sham, WBV7, and WBV14 groups, despite a trend for lower density in the WBV14 group compared with sham rats (-24%; Fig. 9E). Axonal densities in the SCI rats were significantly increased compared with intact rats.
Discussion
The results of the present study provide for the first time experimental evidence for some beneficial effects of WBV treatment on recovery after SCI. WBV improved body weight support during locomotion, urinary bladder function, and the synaptic input to the ventral horn of the distal spinal cord.
Therapeutic timeframe for WBV
We found positive effects of WBV when it was initiated at 2 weeks after SCI. Treatment begun at 1 week had no measurable effects; furthermore, initiation of WBV at 1 day after injury appeared harmful (preliminary data not shown). These results suggest that WBV might be a feasible and effective therapy only when initiated subacutely after SCI. Several reasons may be considered as essential for the failure of early-onset WBV treatment. One possibility is that wound healing and the recovery of vertebral column stability and function of the spinal musculature may require approximately two weeks. Second, within the first 2 weeks after contusion/compression SCI, the blood-spinal cord barrier is severely disrupted 57,58 and extravasation of inflammatory blood cells is prominent; 59 (own unpublished observations). Third, it is possible that some remodeling of intraspinal and supraspinal circuitries (i.e., axonal sprouting and new synapse formation), 5,6,60 a process that takes several weeks, must first occur to enable positive effects of WBV on the outcome of SCI.
Effects of WBV in humans
It is generally assumed that the effects of WBV on muscle performance are elicited through increased discharge of the primary muscle spindle (Ia) afferent fibers and subsequent facilitation of homonymous alpha motoneuron output. 18,20,23 Motoneuron firing frequency and synchronization are improved, 23 leading to increase in muscle activity and force. 61 However, a strong discharge in homonymous Ia fibers due to local application of vibration to a muscle or its tendon results in depression of that muscle's stretch reflex and H-reflex. 62 Thus, vibration causes, in addition to enhancement of motor discharge, a concurrent reflex depression, a phenomenon known as the “vibration paradox”. 62 This phenomenon helps to explain data demonstrating that exposure to WBV enhances muscle performance 18 –20,23 and, at the same time, decreases stretch reflex excitability. 27,63
Numerous studies have documented increased muscle strength after WBV in healthy individuals, 21,64 –68 counter-movement jump, extension strength of lower extremities with an isokinetic horizontal leg press, and a sit-and-reach test for flexibility, 21,65,69 as well as other performance measures such as jump ability. 66,67,70 –72 WBV has beneficial effects on motor functions also in elderly healthy humans, 66,73 –75 as well as in patients with stroke, 76 –78 spastic diplegia, 79 and multiple sclerosis. 80 For individuals with SCI, pilot studies have indicated positive effects on walking ability and spasticity, 27,29,81 as well as arterial blood flow and EMG activity. 82 In addition to neuromuscular effects, vibration increases bone density in noninjured people and has been proposed as a therapy to prevent osteoporosis in individuals with SCI. 82 –84
WBV in humans is typically applied by using a vibrating platform on which the subjects stand (i.e., with body weight support of the lower extremities). 72,75,84 –86 In the present study, rats lie rather than stand on the platform during WBV (Fig. 1) (i.e., the body weight load on the extremities is limited). We cannot currently judge the importance of weight- versus nonweight-bearing WBV training for the treatment outcome. However, from human studies, it is apparent that positive effects can be achieved without weight support during training. Focal vibration treatment of limb segments, for example lower limb, or even single muscles (quadriceps) in humans with SCI have shown effects without body weight support including increased muscle activity, 87 decrease in spasticity, 15 and decrease in spinal reflex excitability, and modulation of the mRNA expression levels genes associated with synaptic plasticity. 88 Moreover, continuous vibration of lower limb muscles (quadriceps and hamstring muscle groups) in healthy individuals, as well as in persons with complete and incomplete SCI, is a sufficiently strong stimulus for spinal cord-autonomous stepping to induce involuntary cyclic locomotor-like movements when the legs are supported in a gravity-neutral position. 89
The Galileo vibration device for small animals, which we used here, works in a way comparable to the Galileo devices for human application. 16,17 There is a 180° phase shift between the two platforms and, in humans, positioning of the left and right lower extremities on separate platforms allows mimicking the physiological alternation in neuromuscular lower limb activity during walking. In the rat, alternation in the vibration phases was achieved between fore- and hindlimbs/portions of the body and not between left and right extremities.
The acceleration of the small animal stimulation system is comparable to WBV devices used for humans: side-alternating systems such as Galileo are using an amplitude up to 5.0 mm and a frequency of 5–30 Hz, and vertical systems are using an amplitude of 1.0 or 2.0 mm and a frequency of 25–50 Hz. These combinations of amplitude and frequency may yield an acceleration up to 15 g. Whether the difference between the mode of training in the rat and human represent a limitation of the animal model is not clear. However, the positive results achieved by our training definitely encourage further use of rodent models to better understand physiological, cellular, and molecular responses to vibration in CNS injury paradigms.
Effects of WBV on motor recovery and potential morphological substrates
Using the BBB locomotor rating scale, we found a trend (p=0.08, Fisher's test) for better outcome in WBV-treated rats (WBV14 group) versus sham-operated animals at 12 weeks after injury. This result appears to be unencouraging, even if we speculate that failure to confirm statistically significant differences is due to moderate sample size (8–9 animals per group) rather than absence of training effects. However, importantly, the BBB rating scores after severe SCI primarily reflect only one aspect of locomotion, namely ordinary plantar stepping as opposed to post-paralytic dorsal stepping, with limited contribution to the scores of other aspects of locomotion such as trunk stability or limb coordination. 34 Using an objective measure for plantar stepping, the foot-stepping angle, we found, similar to BBB score ratings, no significant effect of WBV. The good agreement between the two outcome measures, BBB score and foot-stepping angle, is not surprising since previous work has documented high degrees of correlation between these parameters in rats and mice with SCI. 34,41 The conclusion from these results is that WBV does not improve stepping after severe compression SCI. In line with this notion are the findings that the treatment does not lead to significant changes at 12 weeks after injury in the primary afferent input to motoneurons (VGLUT1+ terminals), in the perisomatic motoneuron coverage by cholinergic synapses (ChAT+ terminals) and the H/M ratio, variables previously shown to correlate well with plantar stepping abilities in rodents with compression SCI. 41,52
The use of the SFMA approach allowed us to estimate numerically, by the rump-height index, another aspect of locomotion, namely the ability to support body weight. We found an apparent improvement in this parameter at 6–12 weeks in rats with WBV (WBV14 group) compared with sham-operated rats. This is an important finding from two points of view. First, the result clearly indicates that the simultaneous use of different outcome measures (BBB rating and SFMA in this study) and estimates of specific aspects of locomotion (e.g., plantar stepping and body weight support ability) allows more insightful conclusions about treatment effects and reduces the possibility to oversee such effects compared with a single measure of locomotor abilities (see also Lee 51 ). Second, the WBV training is expected, deduced from data on humans (see above), to improve muscle strength and synchronization. The ability to support body weight depends on coordinated force output of different muscles in the lower limb joints. Thus, we can conclude that our WBV treatment has achieved an effect anticipated from the kind of rehabilitation training employed. This notion concurs with previous experimental work showing not surprisingly that the type of training, for example, stepping versus walking, determines the type of changes in spinal circuitries and the respective functional alterations (i.e., functional outcome) in a task-specific manner. 6 We do not know the morphological variables that influence the ability to support body weight in our experimental model. Still, the finding that WBV initiated at 2 weeks after injury partially restores the total synaptic input to the ventral horn of the spinal cord suggests that synaptic rearrangements in the distal spinal cord might be important for improving body weight support. It is also conceivable that local changes in the vibrated muscles lead to better performance. Our preliminary data do not suggest differences in the extent of muscle fiber atrophy in WBV- and sham-treated rats, but such a finding would not preclude the presence of other favorable alterations such improved blood supply or preservation of contractile elements.
In addition, we can speculate that WBV14 attenuates in some unknown way the late, post-acute tissue damage and “scarring” at the lesion site. Although lesion volume at 12 weeks after SCI in WBV14 versus sham-treated rats was not significantly reduced, the difference between the mean values of the two groups was considerable (-20% in WBV14 animals). Such a “marginal” effect at the lesion site could underlie the observed tendency for higher BBB scores in the WBV14 group which is present already 3 weeks after SCI (Fig. 2A) since BBB scores are well linearly correlated with lesion volume. 34 However, it appears unlikely that the main, well pronounced effect of WBV—the enhancement of body weight support—is related to changes in lesion volume since the rump-height index in WBV14 rats improved, as compared with sham rats, late after SCI (6 weeks) and the rump-height index is less well correlated, in a nonlinear fashion, with lesion volume than the BBB scores. 34
Using SFMA, we also estimated precise stepping during inclined ladder climbing but found no effect of WBV. Precise stepping requires high levels of supraspinal motor control and a severe compression injury of the thoracic spinal cord in rats completely abolishes this ability with virtually no spontaneous recovery. 34 It is unknown whether WBV induced adaptive changes that could favor precise locomotion, for example, axonal regrowth beyond the injury site or axonal sprouting and formation of novel functional circuitries, although reduced dieback and increased synaptogenesis has been shown to be induced by treadmill training in SCI mice. 90
The overall conclusion from the motion analysis data is that WBV, as performed here, is a treatment leading to improved ability to support body weight after SCI. The improvement might be related to neuromuscular training exerted both in the muscles and in spinal circuitries promoting muscle activation and coordination.
Possible mechanisms of improved bladder function after WBV
Improvement of bladder dysfunction is of high priority for humans with SCI. 91 and, therefore, the finding that WBV initiated at 2 weeks post-injury improves voiding in rats is of special interest. Similar improvements have been documented in human SCI. 11,13 In search of possible mechanisms that might underlie this improvement, we have to briefly consider the control of lower urinary tract (LUT) function (urine storage and release) in non-injured rats. 92 During urine storage, the urethral sphincter—under the influence of the lumbar sympathicus and the mesenchymal periaqueductal gray (PAG)—is responsible for outlet resistance. Release of urine (micturition) is produced by coordinated contractions 93,94 of the smooth musculature of the bladder (m. detrusor vesicae) in combination with intermittent contractions of the external urethral sphincter. Effective voiding thus requires synchronous interactions between 1) the sacro-spinal autonomic reflex [(controlled by the supraspinal pontine and cortico-frontal (gyrus frontalis medius and the rostral part of gyrus cinguli) micturition centres to coordinate urine expulsion] and 2) the low frequency contractions of the somatically innervated external urethral sphincter. 95 –97
Currently, we can envisage two possibly complementary mechanisms leading to improved bladder function after WBV. First, although it is considered that loss of coordinated activity of the detrusor and sphincter is, as a rule, permanent after SCI, 98,99 some recovery of voiding may spontaneously occur after incomplete thoracic SCI. 99,100 It is possible that WBV stimulates mechanoreceptors by rhythmic fluctuations in the pressure on the bladder wall and thus facilitates spinal mechanisms that could favor voiding in the course of spontaneous recovery from SCI. Second, the training conditions might have affected the limbic system which has a strong impact on voiding. The SCI in our model is incomplete and it is possible that, compared to sham treatment, WBV enhanced mechanoreceptor signaling from the Gert's nucleus (a cell group in the sacral cord that receives bladder filling information via α-δ fibers from the bladder), via the core of the periaqueductal gray matter to the pontine micturition center (PMC). PMC induces a parasympathetic outflow to the sacral parasympathetic nucleus (stimulating the detrusor) and to the Onuf's nucleus which relaxes the external sphincter. 101,102 In addition, stimuli from other modalities (auditory, visual) might influence the activation patterns in the PMC. Such “ascending” and “descending” stimuli may lead to improved synchronized activities of detrusor and sphincter muscles. However, neither hypothesis (spinal or supraspinal mechanisms) readily explain why bladder function was improved in the WBV14, but not in the WBV7 group. We can hypothesize that bladder “training” is effective only if remodeling of intraspinal and supraspinal circuitries occurs prior to initiation of WBV, which could be the case at 2 weeks after SCI. In contrast, initiating WBV at 1 week might interfere with such remodelling, leading to maladaptations and training inefficacy.
In conclusion, the results of this study provide evidence that WBV treatment improves recovery from spinal cord injury in the adult rat. Applying established measures for the functional outcome after compression injury, we found that, compared with sham-treated rats, WBV14 but not WBV7 significantly improved body weight support during ground locomotion by 6–12 weeks after SCI. Morphological data suggest that enhanced functional improvement is related to enhanced posttraumatic rearrangement of synaptic connections. Improved bladder function after WBV might be especially promising for clinical translation.
Footnotes
Acknowledgments
The helpful comments of N. Brazda and H.W. Müller (Düsseldorf), G.A. Brook and R. Deumens (Aachen), E.A. Joosten and G.C. Koopmans (Maastricht) are highly appreciated.
Funding: This study was financially supported by the Köln Fortune Programm, the Jean-Uhrmacher Foundation, the Scientific and Technological Research Council of Turkey (TUBITAK) and Akdeniz University Research Fund. SAD is a Principal Research Fellow of the National Health and Medical Research Council, Australia.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.