
Abstract
Blast wave-induced traumatic injury from terrorist explosive devices can occur at any time in either military or civilian environments. To date, little work has focused on the central nervous system response to a non-penetrating blast injury. We have evaluated the effect of a single 80-psi blast-overpressure wave in a rat model. Histological and immunochemical studies showed an early inflammatory response, tissue damage and the initiation of apoptosis. With regard to inflammation, polymorphonuclear leukocytes and lymphocytes infiltrated brain parenchyma within 1 h post-blast. Glial-fibrillary protein, cyclo-oxygenase-2ir, interleukin-1β and tumor necrosis factor were present by 1 h and remained detectable at three weeks post-injury. High mobility group box-1 protein was detectable at three weeks. With regard to tissue damage, S100β and 4-hydroxynonenal were present at 1 h and remained detectable at three weeks. Amyloid precursor protein was detectable at three weeks. As for apoptosis, Cleaved Caspase-3 was detectable at three weeks. Morris water maze assessment of cognitive function showed that blast injured animals required significantly more time to reach the platform on day 1 of training and traveled a greater distance to get to the platform on days 1 and 2. Blast-injured animals showed a significant increase in swimming speed (p<0.001), increased total distance traveled (p<0.001) and increased number of entries into the previous quadrant that had contained the escape platform (p<0.05). Magnetic resonance imaging showed hyperintense regions in the somatosensory area within 1 h. T2 relaxation times and apparent diffusion coefficients show increasing trends in both somatosensory and cortical regions. These data indicate an early and lasting response of brain tissue to non-penetrating blast over-pressure injury. This early inflammatory response is indicative of a mild traumatic brain injury. There is evidence of early hippocampal dysfunction.
Introduction
B
Partly because of the difficulty in separating primary injury from secondary and tertiary injury
Blast waves that do not produce visible damage are capable of inducing atypical distribution of phosphorylated neurofilaments in the hippocampus and cortex of rats during the first week following blast. 10 Blast waves induced deficits in physical performance for three days of measurement accompanied by neuronal degeneration and apoptosis in the cerebral cortex at days 1 through 7. 11 Inhibition of inducible nitric oxide synthase reduced or prevented the blast effects on both physical performance and neuronal morphology. 12 We have studied the systemic effects of whole-body blast injury. In animals subjected to three magnitudes of blast wave intensities, graded significant and similar patterns of cardiovascular responses were observed for all three blast forces. 13 There was no evidence of trauma externally at autopsy; yet internally, there was pulmonary hemorrhage, sub-serosal small and large bowel hematomas, and fractured liver. 13,14 There were decreases in cardiac index, hypotension, and bradycardia, which were prevented by bilateral cervical vagotomy and administration of atropine. 14
Recently magnetic resonance susceptibility weighted imaging was used to obtain in vivo measurements of tissue damage, oxidative saturation changes and blood flow changes after experimental TBI in rats.
15
In that study they were able to detect hemorrhagic lesions resulting from TBI. Previously
The cumulative data from this limited number of reports regarding neurotrauma provide evidence that whole body blast wave exposure that does not produce visible damage to the head or loss of consciousness is capable of inducing significant structural, neurochemical, and metabolic changes in the brain, as well as memory and physical performance deficits. This study was undertaken to evaluate the potential for central nervous system injury in response to a non-penetrating blast exposure. In this study we evaluated both body shielded and non-shielded conditions.
Methods
Animals
Thirty male Sprague-Dawley rats, 250–300 g body weight, were used for primary blast wave experiments involving control, shielded and non-shielded groups. Primary blast groups were evaluated at 1 h and 24 h post-injury by MRI. Cognitive function groups were evaluated by Morris water maze (MWM) at five days post-injury to determine chronic cognitive dysfunction. At the conclusion of both MRI and MWM experiments, animals were perfusion fixed and tissues were harvested for histology and immunohistochemistry. Control animals were anesthetized and handled the same as the experimental animals but were not injured. Animals had free access to food and water. Blast wave exposure was carried out under general anesthesia using Forane anesthesia, 4% in 100% medical oxygen. Following induction of anesthesia, animals were maintained via nose cone on Forane anesthesia, 1–2% in a nitrous oxide/oxygen 70/30 mixture. This anesthetic regimen was chosen because it provides a rapid recovery, thereby limiting anesthetic effects due to variable anesthesia times. Once on maintenance anesthesia, animals were placed in the supine position on a foam pad of known elastic properties and positioned under the blast cone. Although the exact properties of the foam are considered proprietary by the manufacturer (Foam to Size Co., Ashland, VA), this material has been designed to support the rat head in a closed head injury weight drop model such that skull fracture is prevented. The animal is placed in a trough cut to accommodate the body and head. The animal is gently secured in place using tape. There is some spatial movement of the animals head in response to the blast wave, but this is minimal (∼1 cm) and the head is cushioned from impact by the foam. The anesthetic nose cone is then removed, in order to prevent unwanted mechanical effects during the blast procedure. The regulator on the nitrogen tank is then opened and the Mylar sheet ruptures at 200 psi. When positioned 4 cm from the tip of the cone, animals receive an overpressure wave of 80 psi lasting 2 msec. Animals are observed for apneic periods immediately following the blast and time is recorded in seconds, where applicable. This time is generally less than 15 sec. This study was approved by the Institutional Animal Care and Use Committee, University of Oklahoma, Health Sciences Center.
Blast
The blast cone has been described in the literature. 13 Basically it consists of a two part steel chamber. A Mylar sheet is placed between the upper and lower sections of the chamber and the device assembled via retaining screws. Compressed nitrogen is delivered to the back side of the Mylar sheet. In accordance with the tensile strength of the Mylar, the membrane ruptures, delivering the blast over-pressure wave to the animal. The magnitude of the pressure wave delivered to the animal is determined by the distance between the front of the cone and the animal. Pressure is measured using a piezo-electric pressure transducer (Piezotronics, Inc., Depew, NY). The probe is calibrated by the company in millivolts/psi. We construct the curve relating distance from cone to psi in bench studies. In addition, we can position the probe within the blast field and measure the pressure amplitude real time. For the purpose of this study, we chose a blast pressure of 80 psi, based on the results of our previous studies. 13,14,22 Figure 1 shows a representative pressure Friedlander wave. The peak pressure is 79 psi with a nearly instantaneous rise to peak value (rise time 0.2 msec) and a duration of 1 msec. A negative pressure of 2 psi immediately follows the waveform.
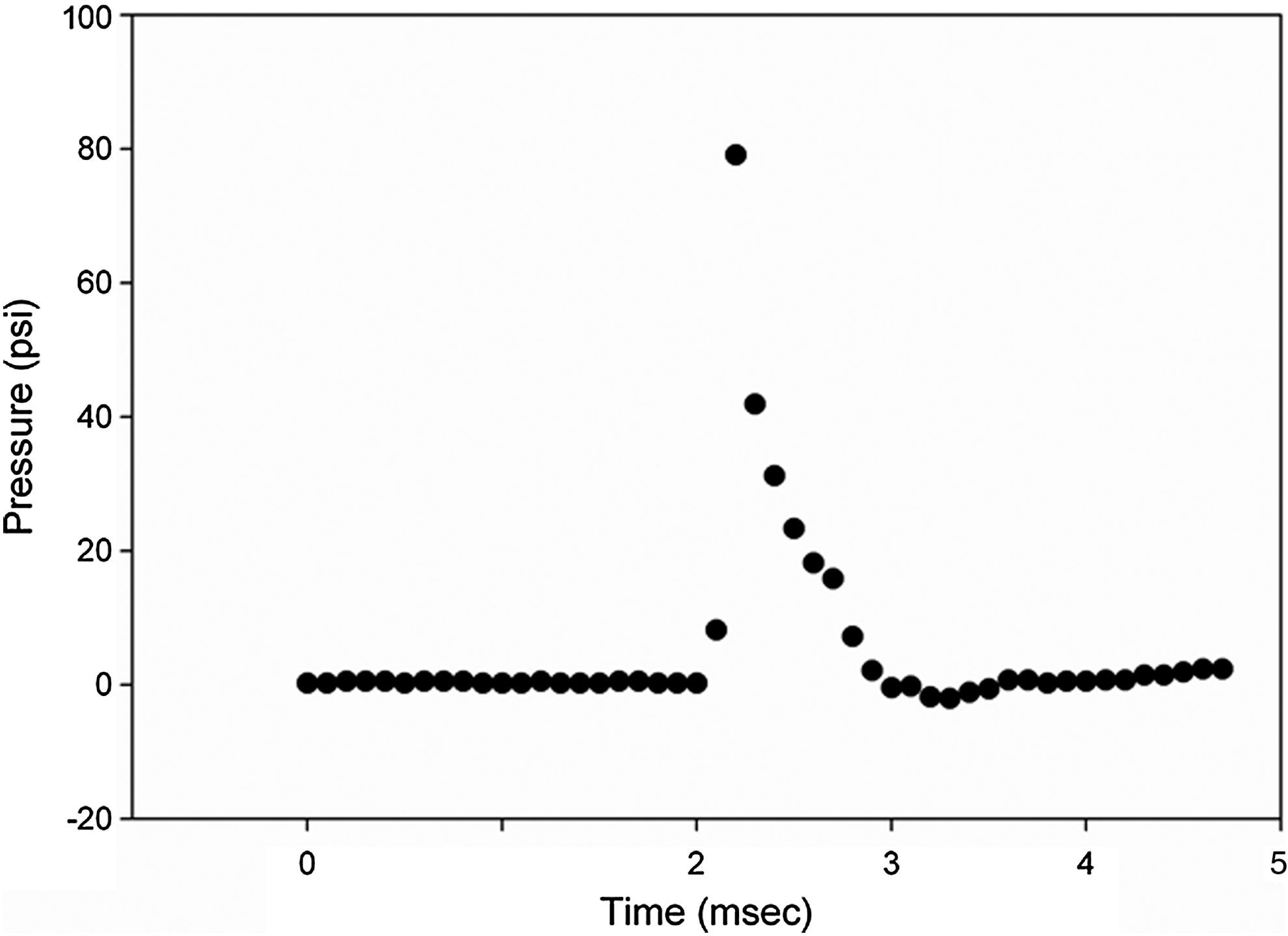
Representative pressure wave form from blast experiment. Peak pressure is 79 psi, rise time 0.2 msec and duration 1 msec. A negative pressure of 2 psi is seen following the wave form.
Histology/Immunohistochemistry
Five micron thick histological sections, embedded in paraffin and mounted on Superfrost®Plus slides (Statlab Medical Products, Lewisville, TX) were rehydrated and washed in Phosphate Buffered Saline/Tween (PBS/T). The sections were processed for Immunohistochemistry using the ImmPress staining system (Vector Laboratories, Burlingame, CA). The ImmPress system reagents provide high sensitivity with very low background staining. According to the manufacturer's directions, sections were treated with peroxidase blocking reagents to inhibit endogenous peroxidase activity followed by washes in PBS/T. If necessary, antigen retrieval was accomplished according to the manufacturer's recommendation for that particular antibody. Following incubation at room temperature or overnight at 4°C, sections were washed and the labeled polymer applied. Visualization was accomplished using the appropriate substrate and counterstaining carried out with ImmunoMaster Hematoxylin (AmericanMaster*Tech Scientific, Inc., Lodi, CA). Controls were incubated with the appropriate isotype antibody.
MWM
The water maze consists of a white, circular tub, 1.5 meters in diameter, containing water that is 30.0 cm deep. The tub is located in a testing room containing numerous distal spatial cues that are visible to an animal within the tub. The position of the hidden escape platform could be determined by reference to these external cues. The water temperature in the tub is maintained at 23°C. A 10.0×10.0 cm escape platform is submerged 2.5 cm below the surface of the water. The water was made opaque by the addition of a non-toxic, water color paint.
On the first test day (five days post-injury) each subject was placed onto the submerged escape platform within the water maze and allowed to remain on the platform of 15 sec before receiving four training trials. For each training trial, the animal was placed into the water maze facing the tub wall at one of four locations selected in a semi-random fashion. The animal was allowed to swim to locate the escape platform. If the animal did not locate the platform within 60 sec, it was guided to the platform. The animal then remained on the platform for an additional 15 sec before being returned to a warmed holding cage for an inter-trial interval of 5 min. Following this initial training day, animals received four training trials per day for a total of seven days. The location of the escape platform remained fixed throughout the seven days of training.
A final probe trial (navigation test) was conducted on the last training day after the four regular training trials. For the probe trial, the escape platform was removed from the tub and the animal was allowed to swim freely for 60 sec. During the training trials and probe trial, data were recorded by means of an overhead video camera and computer software (AnyMaze, Stoelting, CO) that provided numerous measurements of water maze performance. Differences in maze performance between sham controls and blast-injured animals were analyzed with a repeated measures analysis of variance. All analyses were performed by a blinded rater. For cognitive testing, groups included control and body shielded. An un-shielded group was not tested because of the difficulty of separating systemic effects from central nervous system in this physical behavioral test.
MRI
Animals to be used in the MRI studies were maintained on Forane anesthesia for the duration of the imaging experiment, approximately 1 h. Animals in the 24 h group and those for the MWM experiments were allowed to awaken immediately following the blast wave injury. Animals were removed from the foam pad, placed in their housing box and warmed at 37°C with a heating lamp until awake. All animals were observed until they were awake and freely moving about the cage. Animals in the 24 h MRI study and the MWM paradigm were then returned to the housing facility. The MRI study was conducted 24 h later. At the conclusion of both the 1 h and 24 h MRI study, and while under anesthesia, animals were perfusion fixed via an intra-cardiac route and tissues were harvested for histological and immunohistochemical evaluation. MWM animals were allowed three days to recover from the acute effects of the blast injury and were then entered into the training protocol. Following completion of the MWM studies, animals were anesthetized and perfusion fixed, and tissues harvested as described above. Control groups were treated in the same manner.
All experiments described here were performed on a 7T/30 Bruker Avance spectrometer with a cross coil setup. A dedicated head coil was used for some of the images shown here, but the rest were completed with a 2.5 cm flat surface. While setting up imaging parameters using the dedicated head coil, major signal changes were observed in the frontal lobe of the rat brain as shown in Figure 1. On this basis when using the flat surface coil, it was decided to focus the frontal lobe on the center of the coil for maximum sensitivity.
The following parameters were used for the various MRI experiments conducted. For T 2 weighted imaging, a RARE-turbo-spin-echo pulse sequence method was used to collect all images with two or four echo times and t e=12.2, 36.6, 61.0, 1.0, and 85.4 msec with repetition time, t r=0.3 sec. Images were collected in the coronal plane, in 3D mode with a matrix of 256×200×32, field of view FOV=3.83×2.75×1.50, using anti-aliasing in the second phase direction. Accordingly, data were reconstructed to a final matrix of 256×200×32. Diffusion weighted images were collected using a spin echo sequence with diffusion weighted gradients applied in the x-direction only. The following parameters were used; t e=58.8 msec, t r=2.5 sec, gradient separation Δ=50.0 msec, gradient duration, g t=5.0 msec, yielding b values of 95.0, 307.0, and 770.0 sec/mm2. The in plane FOV was as for the T 2 acquisition but with the slice thickness increased to 1.28 mm from 0.75 mm. Five slices were collected in the coronal plane. These parameters could do with further refinement but nonetheless provided acceptable S/N for analysis and calculation of apparent diffusion coefficients (ADC). For chemical shift imaging (CSI) the field of view was set to 2.5×2.5 cm. Pure volume selection, was achieved with a point resolved spatial selection (PRESS) gradient scheme. Thus an additional refocusing pulse extends the original spin echo CSI sequence. The last pulse in this sequence was cycled through+− 180°. The second echo time t e2 of this sequence was incremented 25 msec to 125 msec in 1.6 msec steps to yield 64 spectra.
Results
Histology/Immunohistochemistry
In general, these markers were detectable throughout the three-week study period. Photomicrographs at three weeks are shown as representative of the long-term activation of the inflammatory response to this blast injury. Slides were rated by a blinded observer using the following methodology. Sections were scored for the intensity of 3,3'-diaminobenzidine (DAB) staining (brown color) and were assigned a score of 0 to 3, with 0 representing no staining, 1 representing weak staining, 2 representing moderate staining and 3 representing strong staining. In addition, the percent area of stained cells was determined and assigned a score of 0 to 4, where 0 is 0–2%, 1 is 3–10%, 2 is 11–50%, 3 is 51–80% and 4 is >80%. The intensity and area percent scores are added to give a final score of 0 to 7 in an ordinal scale. These ranked scores are then used to determine statistical significance in a Rank Score procedure using a one way analysis of variance (ANOVA) test in SAS/ACCESS® (SAS Institute, Inc., Cary, NC) as recommended by our statistician.
Coronal hematoxylin and eosin (H&E) stained sections from paraffin-embedded blocks were examined to characterize changes in brain 1 h, 24 h, and three weeks following blast wave exposure. Brains from the 1 h post blast animals, shielded and non-shielded, demonstrated scattered collections of inflammatory cells that seemed to be more constant near the cortical surface but were also present in scattered collections throughout the cortex. Although present, the fewest of these were seen in the 1 h shielded animals. The 24 h non-shielded animals demonstrated the greatest infiltrate of polymorphonuclear leukocytes (PMN) and lymphocytes (Fig 2A, original magnification 40×, arrows denote infiltrate, p<.003). In each of the blast animals there was an infiltrate which was largely periventricular and perivascular and included both PMNs and lymphocytes. The control non-blast animals did not exhibit evidence of inflammatory cell infiltrate. At three weeks post blast (shielded) there were no differences between the control and the shielded blast animals.
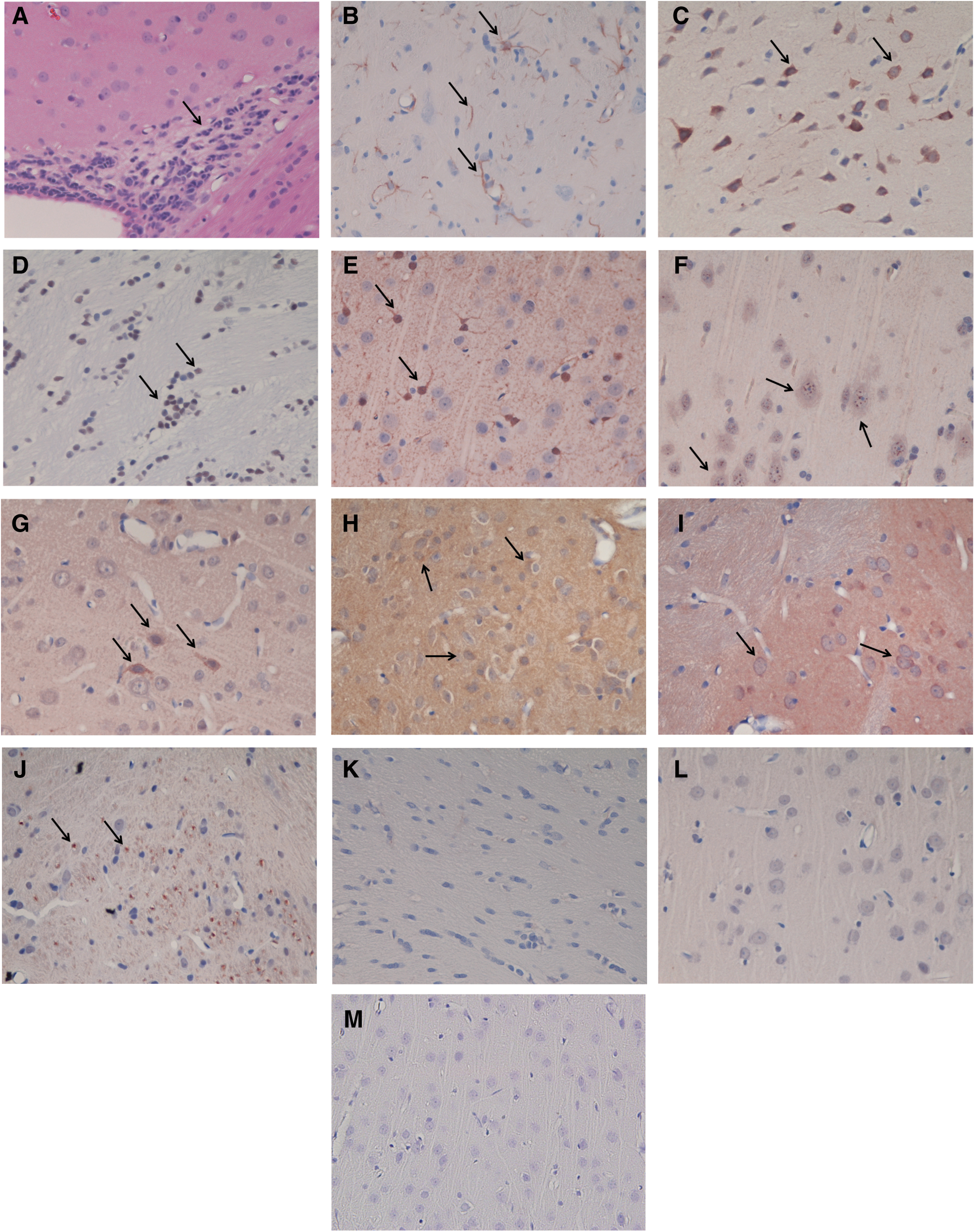
Although expression of glial fibrillary acidic protein (GFAP) was markedly increased in the 24 h blast animals, there was no difference in the pattern between the shielded and unshielded animals Staining in the normal and the 1 h, shielded and non-shielded blast animals was unremarkable. However, three weeks post blast (shielded) there was multi focal staining that involved neurons, astrocytes and oligodendrocytes. (Fig 2B, original magnification 40×, arrows denote positive staining, p<0.003).
Cyclooxygenase-2 (COX-2ir) is an inflammatory mediator involved in the cyclooxygenase pathway. The 1 h shielded and non-shielded animals the intensity of the staining increased to 2+ and involved 60–70% of the neurons in the cortical and para ventricular areas. There was no discernible difference in the two groups. In the 24 h unshielded animals, the intensity of the staining increased to 3+ and involved 80% of the neurons in the two regions. The intensity of staining in the shielded animals was as great as that seen with the unshielded but involved less neurons (60%). The staining pattern for the three week post blast (shielded) animals was similar to that seen with the 24 h shielded group. (Fig 2C, original magnification 40X, arrows denote positive staining, p<.02).
Immunohistochemical localization of the high mobility group box-1 (HMGB1) revealed no staining in either the 1 h or 24 h shielded or non-shielded groups. However, animals in the three week post blast (shielded) groups demonstrated increased reactivity primarily localized in oligodendrocytes. HMGB1 is a nuclear protein that displays extracellular pro-inflammatory properties and is involved as a late mediator in the inflammatory process. (Fig. 2D, original magnification 40×, arrows denote positive staining, p<0.003).
S100β, recognized as a marker of tissue damage in the brain, was also utilized to assess an association between extent of injury and inflammation. The majority of cells stained in the 1 h blast animals were astrocytes with more cells stained in the non-shielded versus the shielded blast animals. At 24 h post blast there was an even divide between neurons and astrocytes. These stained cells spread throughout the brain and were more pronounced near the cortical surface. Animals three weeks post blast (shielded) were markedly positive. (Fig. 2E, original magnification 40×, arrows denote positive staining, p<.003).
Axonal damage was determined with amyloid precursor protein. Rare positive cells were observed in the control animals in contrast to the moderately positive cells in the three week blast (shielded) animals. (Fig. 2F, original magnification 40×, arrows denote positive staining, p<0.003).
Oxidative damage was determined using 4-hydroxynonenal (4-HNE) immuno-histochemistry. HNE is a neuro-toxic by product of free radical-mediated lipid peroxidation. Control animals did not show 4-HNE staining. In both the 1 h shielded and non-shielded were rare neurons stained at their cytoplasmic membranes. At 24 h, a marked increase was observed in both the shielded and non-shielded animals. 4-HNE immuno-reactivity in animals three weeks from blast injury demonstrated lipid peroxidation in both neurons and astrocytes. (Fig. 2G, original magnification 40×, arrows denote positive staining, p<.003).
Localization of the pro-inflammatory cytokines Interleukin 1β (IL-1β; Fig. 2H; p<0.003) and tumor necrosis factor-α (TNF-α; Fig. 2I; p<0.003) in the 24 h shielded and non-shielded animals was more pronounced than the three week animals, however the staining pattern in the three week animals was more diffuse. In general the areas nearest the ventricles were positive and the ependymal cells lining the ventricles were very positive for both TNF-α and IL-1β. There were no differences in staining between the shielded and unshielded animals. Apoptosis was determined in the three week animals using Cleaved Caspase-3 (p<.003; Fig. 2J) immuno-reactivity. Immunohistochemical staining showed cytoplasmic localization of CC-3 in apoptotic cells scattered throughout the tissue. Representative control slides are shown in Figure 2K (GFAP), Figure 2L (4-HNE), and Figure 2M (TNF-α). There is clearly no reaction in the control slides.
MWM, cognitive studies
Data were collected and analyzed using the AnyMaze software. Rating was performed by a blinded observer. Blast-injured animals exhibited more vocalizations during handling and more defecation during training trials than did sham controls, suggesting perhaps an increased level of anxiety. This also was evident in a trend, though not statistically significant, toward increased swim speed in the blast-injured animals (Fig. 3A) and suggesting a lack of motor impairment in these animals. Significant group differences were observed in several measures of place learning across the early training sessions. Blast-injured animals required significantly more time to reach the escape platform during the first training day (p<0.01; Fig. 3B) and traveled a greater distance in the water maze (p<0.05; Fig. 3C) during the first and second days. Both groups showed significant improvement in performance during the seven-day training period, and blast-injured animals did not differ from sham controls (p>0.05) on any response measures by the fifth training day.

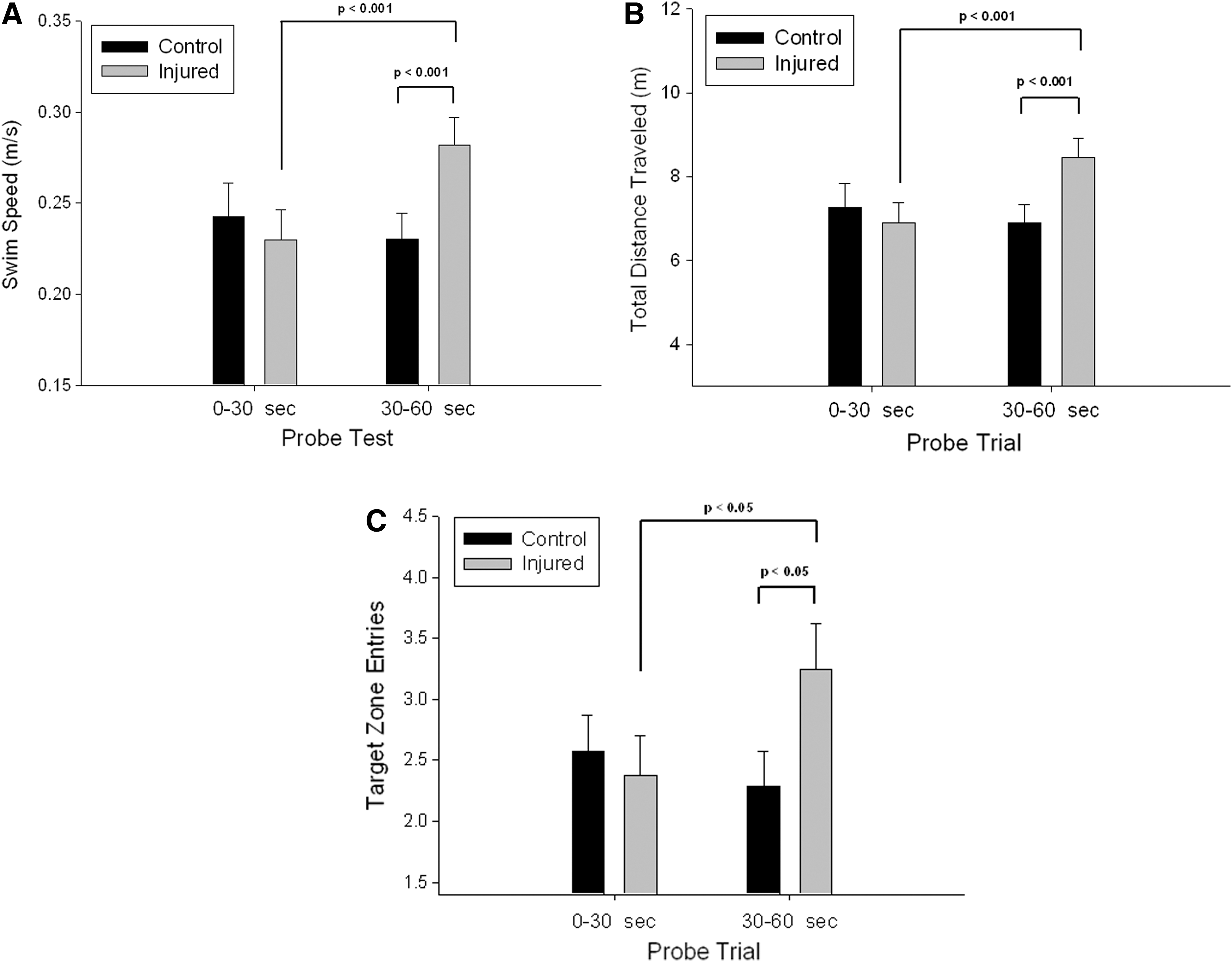
Analysis of the probe trial performance also indicated several significant group differences. Sham controls and blast-injured animals exhibited similar swim speed performance during the initial 30 sec of the 60 sec probe trial (p>0.05; Fig. 4A). However, during the latter half of the probe trial period, blast-injured animals showed a significant increase in swimming speed (p<0.001; Fig. 3A), increased their total distance traveled (p<0.001; Fig. 4B), and significantly increased the number of entries to the previous quadrant that contained the escape platform (p<0.05; Fig. 4C). While there are several possible interpretations of these findings, these data could indicate an increase in response perseveration that is sometimes observed with hippocampal dysfunction. 19 These preliminary data indicate subtle, though detectible, differences in water maze acquisition with relatively mild blast injury and suggest possible hippocampal dysfunction.
MRI
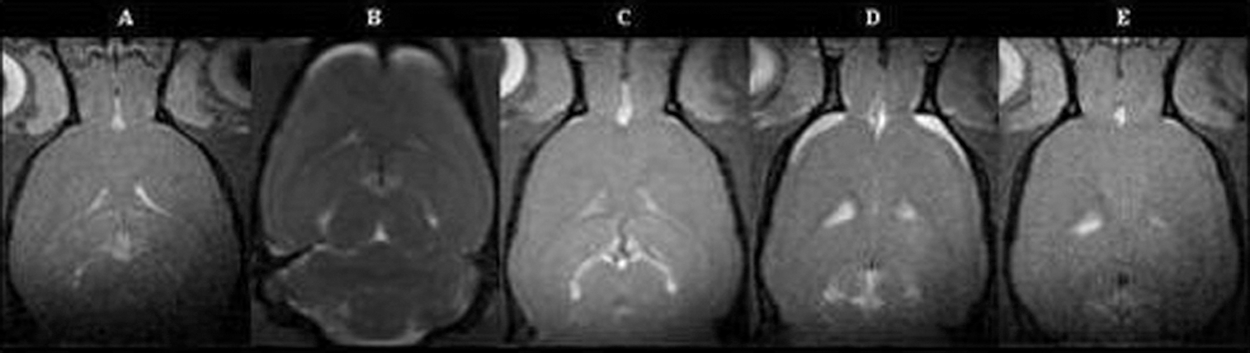
Clearly within 1 h, hyperintense regions were seen in the frontal region as in Figure 5 (regions B and D) and are not evident 24 h later as noted from Figure 5 regions C and E, in both shielded and non-shielded cases. This hyperintense region was seen in all experimental animals (n=5). Control (Fig. 5, region A) shows no hyperintense region. No hyperintense region was seen in any control animal (n=5). This hyperintense region probably represents a subdural effusion of CSF. It is similar to the signal generated by CSF in the ventricles (Fig. 5, regions A and D). This phenomenon is seen in the clinical arena and is referred to as a hygroma. This can be a significant event, although it is usually just observed and will spontaneously resolve. The hygroma does occasionally require surgical drainage. Since this phenomenon is seen at 1 h and is absent by 24 h post injury, it is not likely to be detected in the clinical setting, where patients would present at some remote time point. The significance of this hygroma is yet to be determined in the overall picture of mild traumatic brain injury described herein.
T2 weighted images.
Results of calculated relaxation times T 2 are summarized in Figure 6.

Calculated T2 relaxation times. No significant differences were obtained because of the large standard errors.
The regional T 2 relaxation data correlated with the increases in pure signal intensity in the morphological images within the frontal and ventricle regions. Although the number of animals used was low (n=3 for each group) an increasing trend in T 2 relaxation is observed for the 24 h non-shielded rats. Statistical significance was not reached owing to large standard errors. ADC values also show an increasing trend in the frontal and cortical region, particularly in the non-shielded rats after 24 h In so far as the mechanism is concerned, a great deal more needs to be done to associate the T 2 and ADC changes to some physicochemical process.
We have begun PRESS localized projection spectra collection on these groups (data not shown). Preliminary results indicate that a 2D frequency domain/2D spatial domain chemical shift image method is feasible and extends the breadth of in vivo spectroscopy by yet another dimension.
Discussion
This study was undertaken to evaluate the potential for central nervous system injury in response to a non-penetrating blast exposure. Both the body shielded and non-shielded conditions were examined. While the real-world situation cannot be studied directly, animal models allow separation of primary blast effects from secondary and tertiary. This study shows that even in the case of shielding of air filled organs (i.e., lung, liver, intestine, etc.), a significant CNS response is elicited by a moderate blast over-pressure wave (80 psi).
Histology/immunohistochemistry
Based on observations from the H&Es, known markers of inflammation or injury in the brain were selected for immunohistochemical analyses to help further characterize the inflammatory changes seen by routine screening (Table 1). Preliminary data was generated using specific immunohistochemical stains selected for their ability to detect protein activity associated with injury and inflammation. GFAP is a marker of neuro-inflammation and signals activated astrocytes which are a source of pro-inflammatory cytokines; S-100β is a marker of injury and outcome and is associated with calcium homeostasis primarily expressed in glial cells; ß-amyloid precursor protein antibody is an indicator of the extent of axonal injury; 4-HNE is a cytotoxic by-product of lipid peroxidation and therefore an indicator of free radical activity; HMGB1 is a nuclear protein that displays extracellular pro-inflammatory properties and is involved as a late mediator in the inflammatory process; COX-2ir is an inflammatory mediator involved in the cyclooxygenase pathway; TNF-α and IL-1β are pro-inflammatory cytokines which regulate the interaction of immune and inflammatory cells; and CC-3 detects endogenous levels of activated caspase-3 associated with apoptosis.
GFAP, glial fibrillary acidic protein; HMGB1, high mobility group box-1; COX-2, Cyclooxygenase-2; APP, amyloid precursor protein; 4-HNE, 4- hydroxynonenal; TNF-α, tumor necrosis factor α; IL-1β, interleukin 1β; CC3, Cleaved Caspase 3.
To our knowledge, a comprehensive panel of immunohistochemical antibodies designed to identify proteins associated with injury and inflammation as described above has never been applied to blast wave induced traumatic brain injury. This information will identify mediators and pathways and will facilitate understanding mechanisms involved with blast wave induced brain injury leading to determination of therapeutic targets and biomarkers for detection and diagnosis. These changes may be associated with elevations in pro-inflammatory cytokines and may play a strong role in regulation of the inflammatory processes seen following blast wave exposure.
Water maze
Blast wave induced traumatic brain injury is often associated with cognitive and memory disturbances and neuropathological changes in brain regions that mediate these functions. 23 –27 Hippocampal damage and function are particularly sensitive to mild traumatic brain injury and traumatic brain injury is often reported to produce hippocampal damage in animal models. 9,10,25,28,29
In the current study, the acquisition of a spatial navigation task (MWM) was investigated as a measure of a hippocampus-dependent learned behavior. 30,31 This task is frequently used in animal studies of memory and learning and of cognitive effects of traumatic brain injury. 27,32 –37 This task is sensitive to alterations in hippocampal function. 30,31 In this study, blast-wave injury resulted in delayed acquisition of a spatial navigation task during the first two days of training, but injured animals exhibited normal performance during subsequent trials. This finding is similar to other reports of deficits in early acquisition of spatial navigation tasks with improvements in performance with additional training. 27
In addition, the present findings indicate alterations in performance during a memory probe trial that suggest increased perseveration or inability to suppress inappropriate responses following blast-wave injury. Injured animals showed normal behavior during the first half of a probe trial where the escape platform has been removed from the water maze, showing good retention of the original platform location. But during the latter half of the probe trial when control animals begin searching new locations for the platform, blast-wave injured animals spent significantly more time in the vicinity of the original platform location. Similar increases in response perseveration have been reported following specific injury to the hippocampus. 38,39
MRI
It is generally accepted that MRI technologies are extremely useful for non-invasive investigation. It is clear that early diagnosis is the key to successful treatment and management of mTBI. The use of these technologies for early diagnosis of mTBI will require modification of the MRI protocol to accommodate rapid implementation. The duration of 1 hwas chosen for this study as the shortest time possible to obtain MRI data in this model. This timing will clearly need to be modified in the clinical and/or field setting. Translation to the clinical setting will benefit from techniques such as multiple spin-echo experiments generating T1 and T2 contrast. The application of this methodology is relatively rapid and provides the basic data sets for further investigation using other magnetic resonance methods, such as spectroscopy, perhaps the most promising method in early diagnosis. A PRESS localized CSI method and a stimulated echo acquisition mode (STEAM) are available for this purpose. While the techniques may seem well established, it is generally accepted that methods such as PRESS and STEAM are limiting. While CSI is also in use, additional information in an experimental setting can only be achieved by extending these techniques further. Recently, 2D in vivo spectroscopy has been reported from clinical spectrometers.
Even though there appears to be little MRI data available related to blast wave induced brain injury, MRI is the imaging modality of choice for assessing brain pathology. Extensive studies using MRI to assess TBI produced by classical methods such as weight drop and fluid percussion have been published over the last decade. It is essential to note that these classical methods produce focal injury rather than the diffuse injury produced by blast waves. These studies range from morphological assessments 2,4,6,10 , to obtaining functional (measurement of water diffusion, contrast agent perfusion, and cerebral blood volume) 15,16,27 and metabolic (magnetic resonance spectroscopy of neuronal metabolites, such as N-acetyl aspartate, creatine and choline) 28 information.
The development of non-invasive tools to permit early diagnosis and the ability to track the disease process will certainly reduce the extent of injury, resulting in attenuation of cognitive dysfunction experienced by blast wave brain injury patients. This approach will yield MRI data relating organic changes to cognitive function, histology, immunohistochemistry-derived injury and inflammatory biomarkers, and cognitive function data from similar if not identical brain tissue samples.
Early diagnosis of blast wave induced brain injury will provide the information necessary to establish procedures to initiate therapeutic interventions.
Statistical analysis
H&E stained sections were used for identification of infiltrating inflammatory cells and were scored using the 4-point scoring system of Mann, et. al., 2012, 40 where 0 is within normal limits, 1 is minimal, 2 is slight, 3 is moderate and 4 is severe. Standardized nomenclature for tissue location of positive staining was according to Kaufmann and colleagues. 41 Grading of response in immunohistochemical staining was done according to the methodology of Regitnig and colleagues. 42 Positive regions were assigned an intensity score of 0 to 3, where 0 is no intensity, 1 is weak, 2 moderate, and 3 strong staining (DAB reaction, brown color). In addition, the percentage of stained cells was determined and assigned scores of 0 to 4, where 0 is 0%, 1 is <10%, 2 is 10–50%, 3 is 51–80% and 4 is >80%. The intensity and percent area scores are added to give a final score of 0–7 representing negative, weak, moderate and highly positive categories. All grading was performed by a blinded rater. These data are Ordinal in nature and as such can be analyzed using a Ranks test one-way ANOVA. Scores for each animal were compared across groups to the Control Group using the npar1way ANOVA test for Rank Scores. Significance was set at 0.05.
Acknowledgments
These studies were approved by the Institutional Animal Care and Use Committee, University of Oklahoma, HSC.
Footnotes
Author Disclosure Statement
No competing financial interests exist.