
Abstract
A comprehensive understanding of the pathology of spinal cord injury (SCI) in non-human primates may facilitate greatly the development of new strategies to promote recovery in humans with SCI. Relatively few studies, however, have been conducted to systemically examine pathological changes in the monkey, a non-human primate, after SCI. We report axonal, glial, and fibrotic responses in the spinal cord of monkey Macaca fascicularis after a thoracic (T) 8–9 lateral hemisection. We examined these changes at three regions—i.e., the lesion epicenter, the peri-lesion area, and the lateral white matter of the intact, contralateral hemicord at 7 (subacute) and 30 (early chronic) days post-injury. The lateral hemisection resulted in a marked axon and myelin loss, along with tissue loss, at the lesion epicenter at both time points. Unexpectedly, axonal loss and myelin degeneration, along with reactive gliosis and microglia/macrophages activation, were also observed in the contralateral spared hemicord, indicating a spread of the initial damage to the contralateral side. In addition, activated microglia/macrophages were found both within the injury epicenter and the peri-lesion area, indicating that they participate in injury-induced immune responses that may exacerbate the secondary damage. In contrast to rodents, substantial reactive astrocytic responses at the lesion border were not observed in the monkey. Conversely, a deposit of robust fibrotic scar was observed at the injury epicenter, which filled the space originally created by the hemisection. Thus, understanding the pathology of monkey SCI may provide clinically relevant information in designing repair strategies targeting specific problems associated with human SCIs.
Introduction
D
With the presence of these differences, it may be difficult to translate research results directly from rodents to the clinic. Because the anatomy and function of the non-human primates are strikingly similar to humans, a thorough understanding of their pathology after SCI, at both the acute and chronic stages, may greatly facilitate the preclinical investigation of therapeutic interventions. Unfortunately, except for early reports on the observation of axonal pathology at light 9 and electron microscopic level 10 in the monkey, relatively few studies have been conducted to examine systemic pathological changes in the monkey after SCI.
The monkey lateral hemisection model at both cervical 11 –13 and thoracic 14 levels has been used widely to study anatomical and functional changes aftervSCI. Using a T8-9 right-sided hemisection model in a non-human primate species Macaca fascicularis, we previously examined axonal and glial responses in areas of wallerian degeneration (WD) remote to the injury site. 15 We reported that distinct differences in pathological changes exist in area of WD in the monkey compared with the rat, 8 but are similar to those in humans. 16
The objective of the present study was to systemically examine major cellular responses at the primary injury site using a T8-9 lateral spinal cord hemisection model in the adult monkey Macaca fascicularis. 15 This model produces a spinal hemisection on the right side and leaves the left hemicord largely spared. The fate of axons, myelin, astrocytes, microglia, and activated microglia/macrophages, as well as formation of fibrotic scars at the lesion site was studied. We examined and quantified these changes at three regions—i.e., at the lesion epicenter, the peri-lesion area, and the lateral white matter of the intact hemicord. We examined these changes at 7 and 30 days post-injury, representing two clinically relevant stages—i.e., a subacute and an early chronic stage.
Methods
Animal groups and surgical procedures
A total of nine adult male Macaca fascicularis monkeys (4–6 years, 3.94–6.12 kg, Jinghong Breeding Station, Yunnan Laboratory Primates, Inc., Yunnan, P. R. China) were used in this study. Three monkeys received only laminectomy as sham-operated controls and were sacrificed at 7, 15, and 35 days after surgery. Six monkeys received right-sided spinal cord hemisection and were sacrificed at either 1 week (n=3) or 4 weeks (n=3) after injury. Before the study, the animals had not been used for other surgical procedures or pharmacological experiments, and were bred in an enriched environment. All surgical operations and post-surgery animal care were approved by the Animal Use and Care Committee of Kunming General Hospital of People's Liberation Army, and carried out in accordance with the Guide for the Care and Use of Laboratory Animals (National Research Council, National Academy Press, Washington D.C.).
All surgeries were performed according to our previous report. 15 In brief, the surgeries were performed under sterile conditions. The monkeys were anesthetized with 15 mg/kg ketamine hydrochloride injected intramuscularly (IM). After absence of all responses to cutaneous stimuli, the skin overlying the thoracic vertebral column of each monkey was incised, and the muscles were detached from the vertebrae. A dorsal laminectomy was performed at the T8-T9 spinal cord level. After opening the dura mater, the spinal cord was cut with fine iridectomy scissors, resulting in a right-sided lateral spinal cord hemisection. The sham-control monkeys underwent a laminectomy alone.
After surgery, all animals were maintained in a temperature- and humidity-controlled room and were kept warm with an infrared heater to avoid hypothermia until their recovery from anesthesia. Manual bladder expression was performed three times during the first postoperative day, after which bladder function returned. The monkeys were fed ad libitum and hydrated with saline. Prophylactic antibiotics were used during the first post-operative week.
Perfusion and tissue preparation
The animals were deeply anesthetized with ketamine hydrochloride (IM). After all reflex responses to coetaneous stimulation were verifiably absent, the monkeys were perfused transcardially with 3000 mL of physiologic saline, followed by 3000 mL of 4% paraformaldehyde in physiologic saline. The flow rate was controlled so that the perfusion was completed within 30 min. The entire spinal cords were dissected out of the vertebral columns and post-fixed in the same buffered fixative at 4°C for 24 h, washed briefly with 30% sucrose-phosphate buffered saline (PBS) (0.1 M, pH=7.4), and cryoprotected with the same 30% sucrose buffer at 4°C. Three equal 1.5-cm–long spinal cord segments, including one centered at the injury epicenter (T8-9), as well as one rostral and one caudal to it, were embedded in optimal cutting temperature compound and sectioned transversely and serially with a cryostat at a thickness of 20 μm.
Immunohistochemistry
The procedures of immunohistochemistry staining were performed according to a previously reported protocol. 17 Briefly, sections were rinsed with PBS (0.1 M, pH=7.4) and treated with 3% H2O2 for 1 h to quench endogenous peroxidase. The sections then were incubated in 10% goat serum - 0.3% Triton X-100 for 1 h. Primary antibodies, including mouse anti-SMI-31 (1:1,000, Sternberger Monoclonals Inc., Baltimore, MD) to recognize the phosphorylated neurofilament epitope of axons, mouse anti-glial fibrillary acidic protein (glial fibrillar acidic protein [GFAP]; 1:3,000, Sigma, St. Louis, MO) to identify astrocytes, rabbit anti-Iba1 (1:200, Wako, Richmond, VA) to label microglia, and mouse anti-KP1 (CD68; 1:200, Dako, Carpinteria, CA) to mark activated microglia/macrophages, were used (Table 1).
All antibodies were diluted in 0.1 M Sorenson buffer (pH=7.2), and the sections were incubated with the individual antibodies overnight at 4°C. On the second day, sections were incubated at room temperature with biotinylated goat-anti-mouse IgG or goat-anti-rabbit IgG antibody respectively (1:400, Vector Laboratories, Inc, Burlingame, CA) for 1 h. After incubation with ABC reagent (Vectastain ABC kit, Vector Laboratories), the reaction complex was visualized with 3,3-diaminobenzidine (Dako, Carpinteria, CA). Then all sections were dehydrated with graded alcohols, cleared with xylene, and coverslipped. The specificity of labeling was confirmed by using isotype control of mouse or rabbit primary antibody (Invitrogen, California, CA).
Luxol fast blue staining
Sections was stained with Luxol fast blue (LFB) for spared myelin, as has been previously described. 18 Briefly, the slides were warmed at 37°C overnight. After dehydrating with 70% and 95% alcohol for 2 min each, the sections were immersed with 0.1% LFB solution at 37°C for 4 h. After cooling in a 4°C refrigerator for 20 min, the slides were dipped in 95% alcohol 5 times and dH2O 10 times. The sections were then infiltrated in 0.05% lithium carbonate for 5 min. After washing with 70% alcohol for 5 min and dH2O for 1 min, the sections were dehydrated, cleared, and sealed.
Collagen stains
Aniline blue solution was applied to examine changes of collagen and fibrous connective tissue in the lesion area under the manufacturer's instruction. Briefly, sections were mordanted in preheated Bouin Solution (HT10-1, Sigma-Aldrich, St. Louis, MO) at 56°C for 15 min, followed by staining in Aniline blue solution (HT154, Sigma-Aldrich, St. Louis, MO) for 10 min. Then the slides were immersed in 1% acetic acid for 2 min, washed in tap water, dehydrated in graded alcohol, cleared in xylene, and mounted with a cover glass using DPX mounting medium (EMS, Hatfiled, PA).
Quantification and statistics
For assessments of axons, microglia, macrophages, and astrocytes, three sections per spinal cord segment at the injury epicenter (T8-T9) were selected at random. At each section, three areas of interest were defined: the lesion area (defined as Area 1 or A1), the penumbra or peri-lesion area (defined as Area 2 or A2) with a band of 300 μm width immediately adjacent to the lesion area, and the lateral margin of the white matter on the contralateral side (defined as Area 3 or A3; 300 μm width). Within each area, four to six photomicrographs were taken at random for assessments.
Quantifications of immunoreactive cells/axons were performed within a rectangle of each 40X photomicrograph (≈64,687.5 μm2) and were averaged. The number of axons, glial cells, and macrophages were counted manually based on their morphological characteristics. The dark or deep brown tiny dots, excluding the big plaques or dim dots, were recognized as axons. The Iba1-, KP1-, and GFAP-immunoreactive (IR) cells, containing a clear nucleus and prominent processes or cytoplasm, were recognized and their numbers were counted. Areas of LFB-positive staining in each section were measured and averaged with the aid of ImagePro Plus software (Media Cybernetics, Inc., Bethesda, MD). The area of LFB-staining was considered positive if the labeling intensity is stronger than the threshold—e.g., intensity of the gray matter.
The total area of each section at 7 and 30 days after injury was estimated by measuring the area of spared hemicord plus its mirror area on the right. The mean optical density (OD) of aniline blue staining areas was measured by setting the threshold of integrated optical density (IOD) with ImagePro Plus software in each section, which includes both the left hemicord and its right mirror image. The blank field of each slide was set as a threshold for measuring the optical density of aniline blue staining. The number obtained from averaged sections of each animal was used as a basic unit for statistical analyses.
All data collection was performed blindly with respect to group identities. Data were presented as mean±standard deviation (SD). One-way analysis of variance (ANOVA) with Tukey post hoc was used to determine statistical significance. A p value of<0.05 was considered statistically significant.
Results
Axons
We first examined axonal degeneration at the injury epicenter after a right-sided spinal cord hemisection (Fig. 1). Axons, IR for SMI-31, were examined in three defined areas—i.e., A1 at the lesion site; A2 at the peri-lesion site; and A3 at the later margin of spared lateral funiculus (Fig. 1G). In the sham-operated spinal cord, dark brown-stained SMI-31-IR axon profiles where clearly seen to distribute evenly in the spinal white matter (Fig. 1A), which can be appreciated at a high magnification (Fig. 1B). At 7 days post-hemisection, tissue loss at the injury site was clearly seen (Fig. 1C). In areas where residual tissue was present (Fig. 1C, dashed line), the number of SMI-31-IR axons was dramatically decreased (Fig. 1D). In this region, degenerative axonal changes, including axonal swelling, fragmentation, and end bulbs, were clearly seen (Fig. 1D). At 30 days post-injury, only a few residual dystrophic axons were seen in the lesion area (A1).
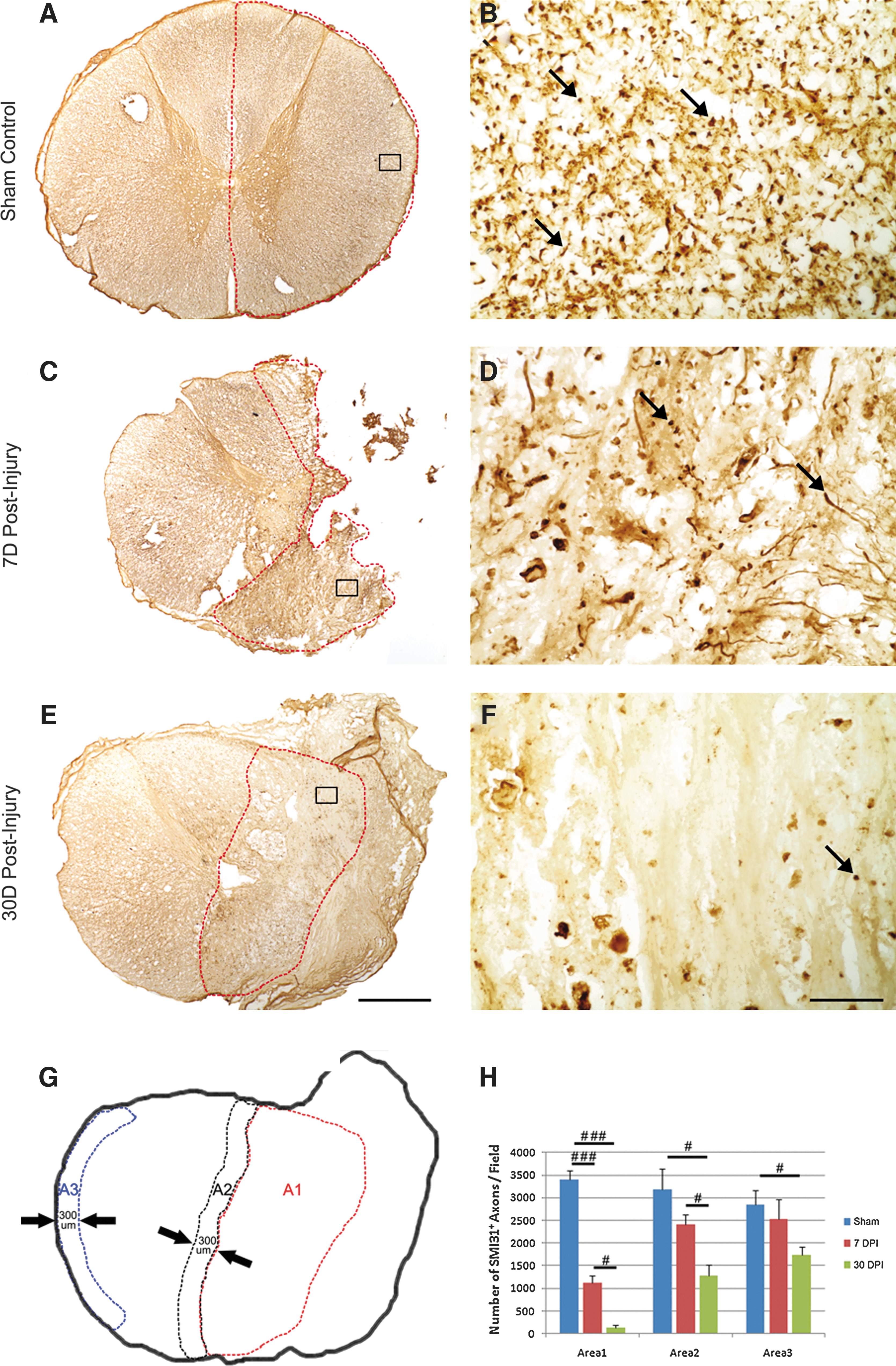
Morphology and qualification of axons at the injury epicenter after T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show distribution of SMI-31+ axons in the sham-operated control
Statistical analyses showed a significant decrease in the number of axons within the lesion site at 7 days post-injury (Fig. 1H, ###, p<0.001, compared with the control). At 30 days post-injury, only a few axons were found at the lesion site (Fig. 1H, ###, p<0.001, compared with the control). As anticipated, a significant decrease in the number of SMI-31-IR axons was found in the peri-lesion area at 7 days post-injury (Fig. 1H, #, p<0.05, compared with the control group), and there was a further decrease at 30 days post-injury (Fig. 1H, #, p<0.05, compared with the 7 days group), likely because of secondary damage to these axons after the initial trauma.
Unexpectedly, the number of SMI-31-IR axons in the lateral white matter of the spared hemicord (Fig. 1G and A3) also showed a significant decrease at 30 days post-injury, indicating that axons remote to the injury site could also be affected by a unilateral spinal cord injury (Fig. 1G and H; #, p<0.05, compared with the control).
Myelin
We next examined the spared myelin in sections stained with LFB at three levels—e.g., at the injury epicenter and at levels 1.5 cm rostral and caudal to it. To calculate the percentage of spared myelin, the LFB-stained area was transferred to black (Fig. 2G). The percent myelin area was represented as the ratio of black area to the whole section area, described above. In sham-operated animals, myelin was evenly distributed throughout the entire white matter (Fig. 2A), which could be further appreciated at high magnification (Fig. 2B). Significant myelin loss (Fig. 2H, ##, p<0.01, compared with the control), along with tissue loss, was observed at the injury epicenter, and no statistically significant difference in percent myelin area was found between the 7-day and 30-day groups (Figs. 2C, 2E, 2H).
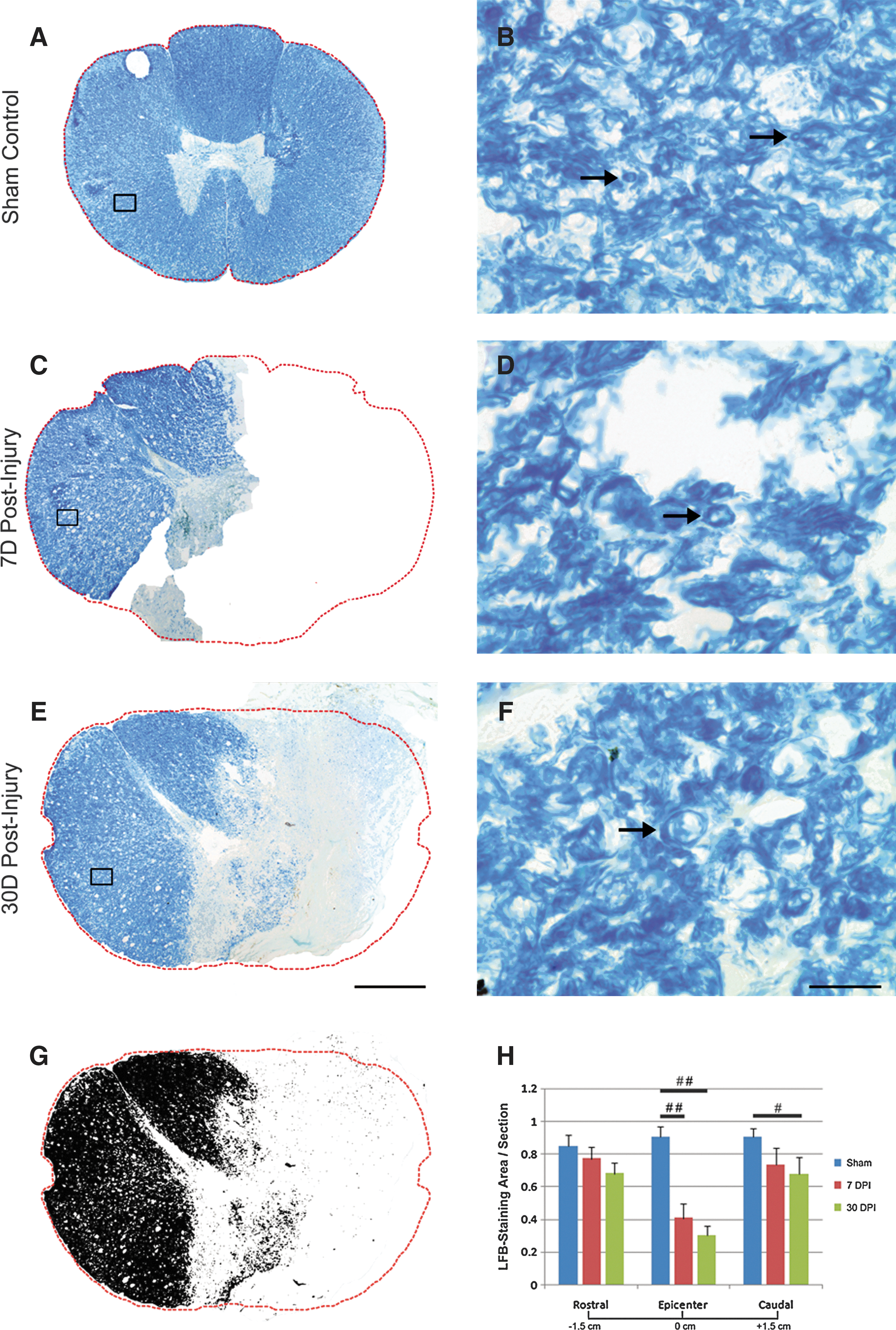
Morphology and quantification of myelin after a T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show distribution of myelin stained with Luxol fast blue (LFB) in the sham-operated control
At 1.5 cm caudal to the injury epicenter, the LFB intensity decreased tardily after injury and was statistically significant between the sham and 30-day group (Fig. 2H; #, p<0.05, compared with the control), indicating that demyelination also occurred caudal to the injury. There was no significant difference in LFB staining, however, between the sham and both injury groups at 1.5 cm rostral to the injury epicenter, indicating that the myelin remained less affected at this level, although a trend of myelin decrease was found at 30 days post-injury (Fig. 2H). Corresponding to axonal decrease in the contralateral spared white matter, described above, characteristics of myelin pathology such as large onion bulbs, thinner myelin, and myelin debris were found in the contralateral spared white matter (Figs. 2D, 2F).
Microglia/macrophages
Microglia and macrophages are implicated in participating in the secondary injury process including axonal degeneration and, later, myelin clearance after SCI. 15,19 To determine the response of microglia and macrophages in monkeys after SCI, antibodies against Iba1 and KP1 were used to identify resident microglia and activated microglia/macrophages, respectively. We found that resident microglia with ramified processes were evenly distributed throughout the entire gray and white matters in the sham-operated monkeys (Figs. 3A, 3B).
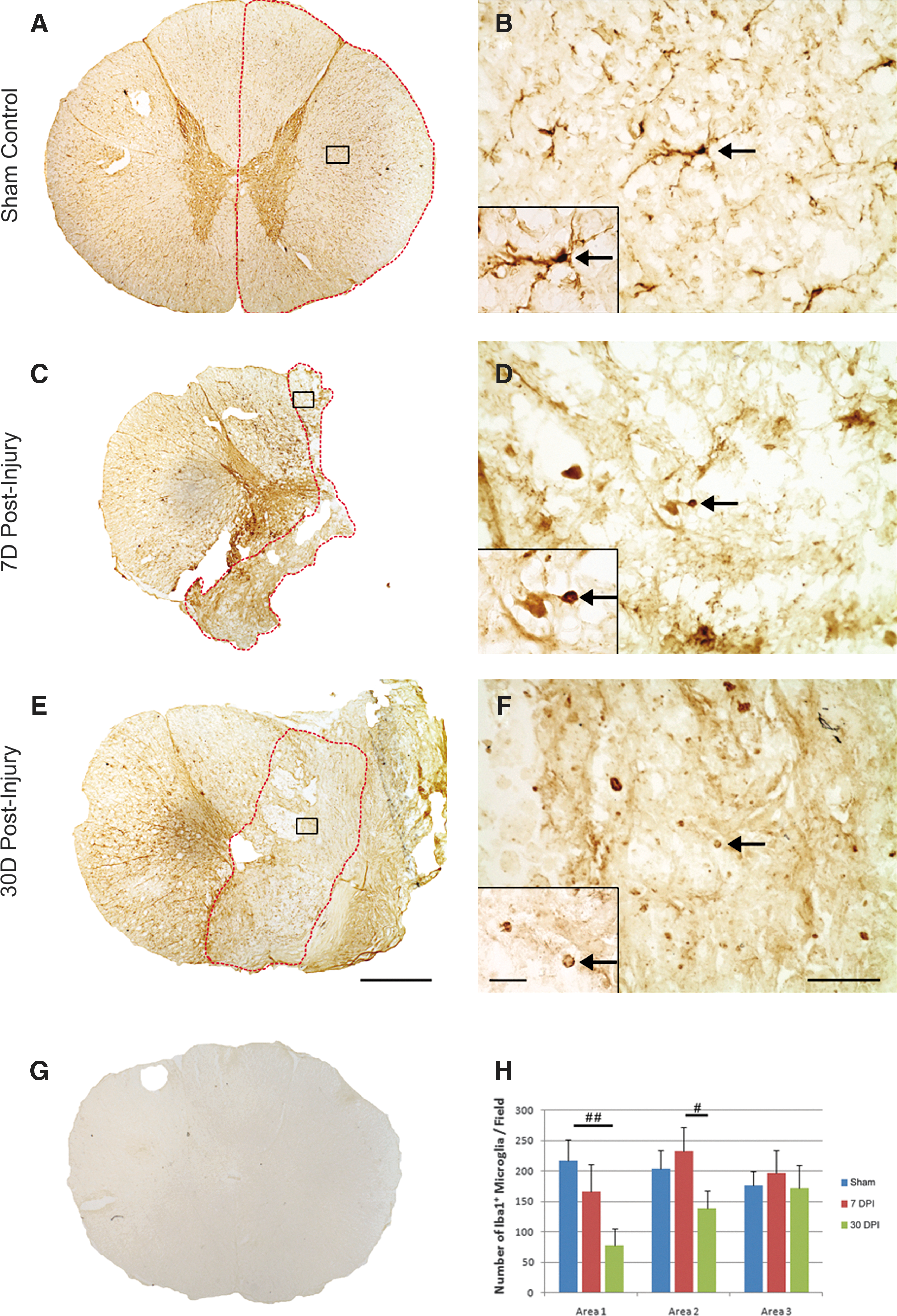
Morphology and quantification of microglia after a T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show distribution of Iba1+ microglia in the sham-operated control
At 7 and 30 days post-injury, microglia became activated and showed rounded cell body and shortened processes (Figs. 3D, 3F). Except for the morphological changes, the number of Iba1-IR microglia/field in both the epicenter (A1) and penumbra (A2) regions did not change between the sham-operated and 7-day post-injury groups (Fig. 3H). At 30 days post-injury, however, a significant decrease in the number of Iba1-IR microglia was found in both regions, indicating a decrease in the number of activated microglia at the early chronic stage (30 days) in both the lesion epicenter and penumbra regions as to the subacute stage (7 days, p<0.05) or the sham-operated control (p<0.01). In regions away from the injury (A3), no statistical significant difference in the number of Iba1-IR microglia was found among all three groups (Fig. 3H).
With the specific KP1 staining, only activated microglia and macrophages were labeled and, therefore, no labeling was found in the sham-operated control (Figs. 4A, 4B). After SCI, microglia with amoeboid-like morphology and strong immunoreactivity at the lesion site were found indicative of their activation (Figs. 4C, 4D). Blood-born macrophages with round-shaped morphology, labeled with KP1, were also observed and contribute to the overall KP1-positive cells (Fig. 4D).
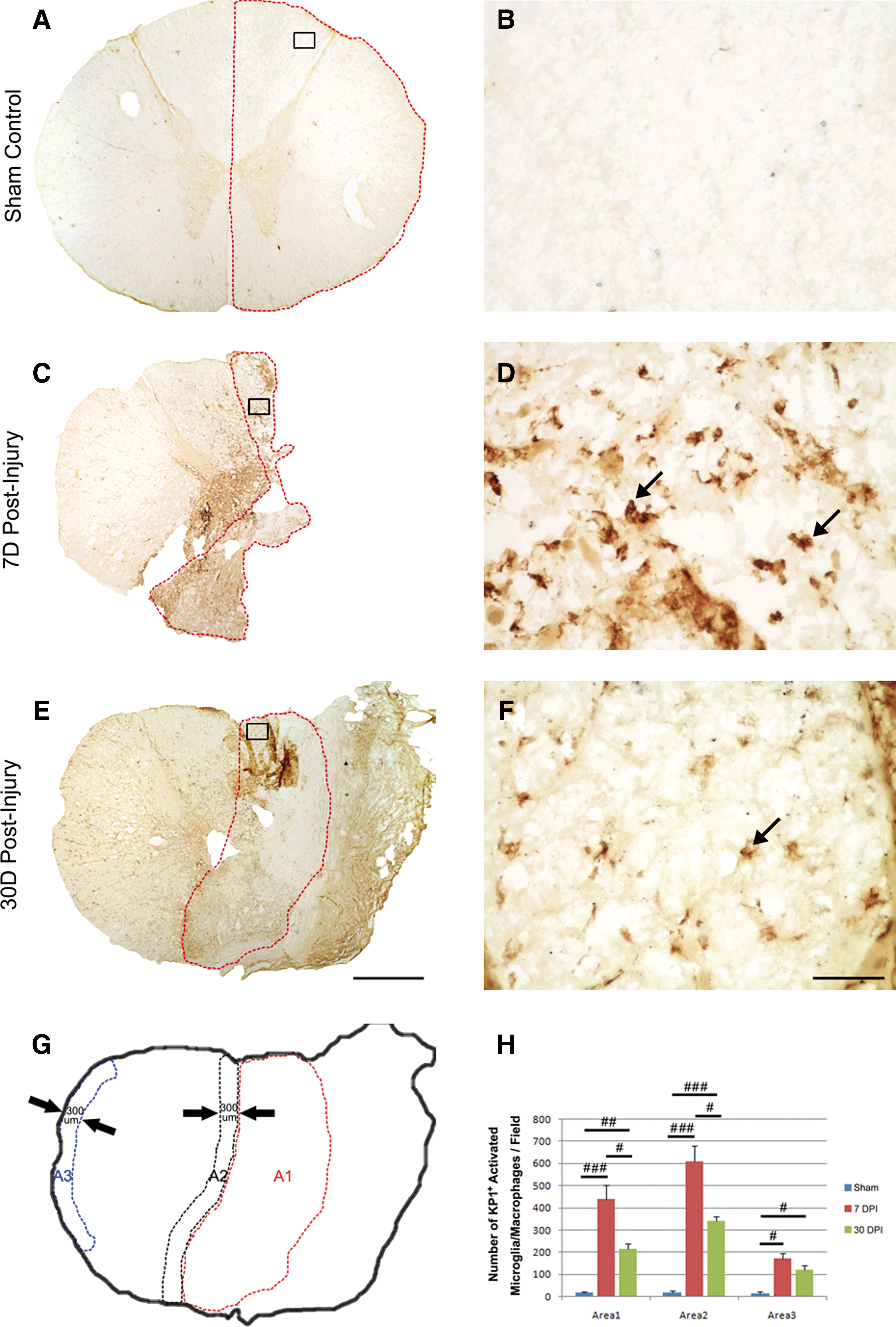
Morphology and quantification of activated microglia and macrophages after a T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show distribution of KP1+ activated microglia and macrophages in the 7- day
Statistical analysis showed that the number of KP1-IR activated microglia/macrophages at the injury epicenter and injury penumbra region was significantly increased after SCI, research the highest at 7 days (P<0.001, compared with the sham-operated control; p<0.05 compared with the 30-day group), and declined but remained statistically significantly high at 30 days post-SCI compared with the sham-operated controls (p<0.01). Notably, a significant increase in the number of KP1-IR activated microglia/macrophages was found in the lateral white matter of spared hemicord (A3, p<0.05, Fig. 4H) compared with the sham-operated controls, indicating a remote activation of microglia/macrophage in this region, possibly from a spread of inflammatory response from the injury. Such activation in the remote region, however, was at a much less extent than in areas at or close to the injury.
Astrocytes
Astrocytes are another cell type that are thought to play a key role in the response to central nervous system (CNS) injuries. 20 We examined the fate of astrocytes at the injury site at 7 and 30 days post-injury. In the sham-operated control, GFAP-IR astrocytes with stellate morphology were seen and distributed evenly throughout the spinal cord (Figs. 5A, 5B). The processes of the astrocytes were fine and distributed in a predominantly radial pattern in transverse sections of the spinal cord. Unlike what was observed in the rat spinal cord, dramatic astrocytic glial scar formation was not found around the lesion site at both 7 and 30 days post-injury (Figs. 5C, 5E). At the penumbra region (A2) of the 7-day injured group, astrocytes became reactive with cytoplasmic hypertrophy and thickened processes (Figs. 5C, 5D). No statistically significant change in the number of GFAP-IR astrocytes was found in this region, however, although there was a trend of increase in the number of GFAP-IR cells over time in this region (Fig. 5H, A2, p<0.05).
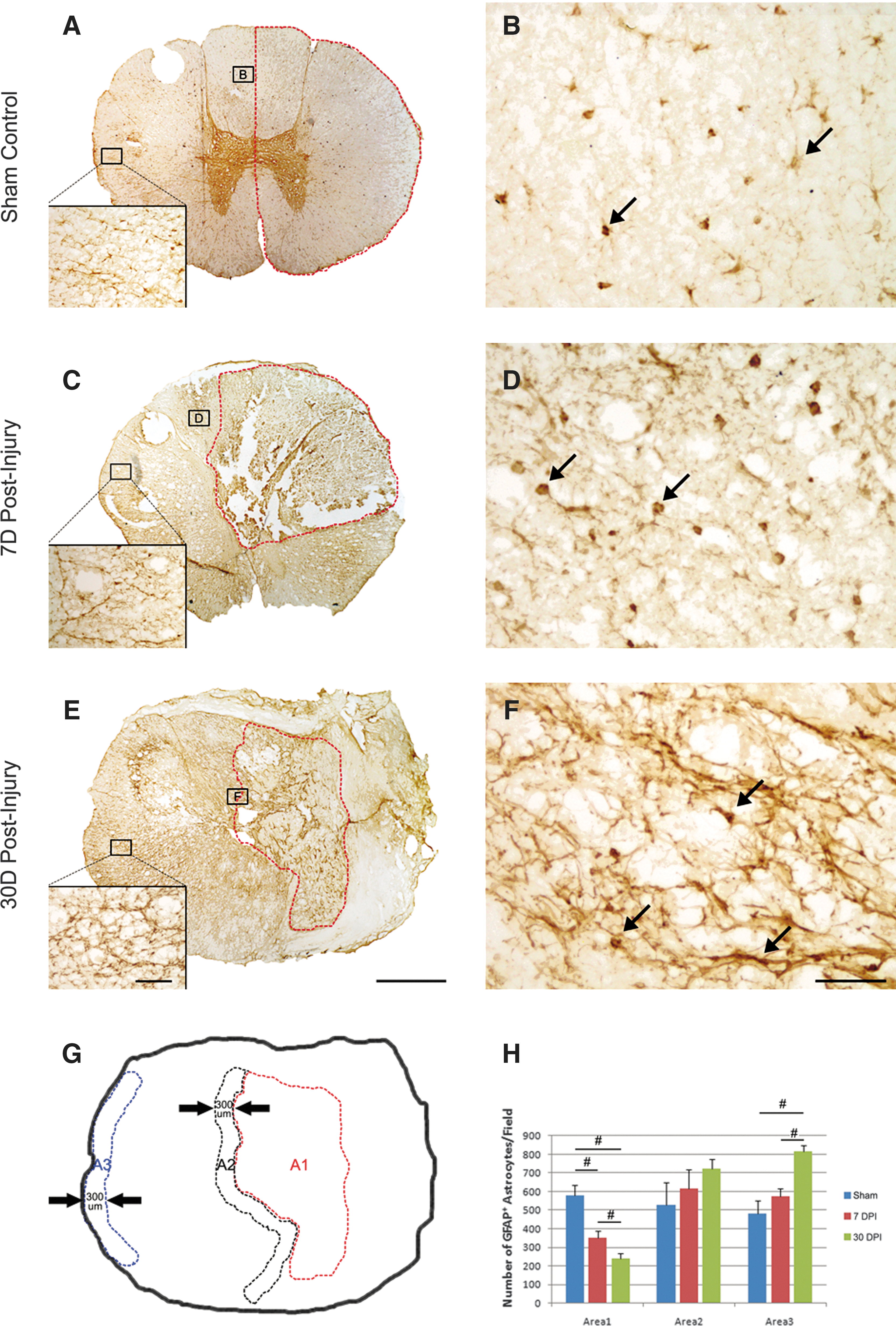
Morphology and quantification of astrocytes after a T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show distribution of glial fibrillary acidic protein (GFAP)+ astrocytes in the sham-operated control
At the lesion epicenter, a steady and significant decrease in the number of GFAP-IR astrocytes was found at 7 and 30 days post-injury (Fig. 5H, Area 1, p<0.05). Interestingly, astroglial reactivity was also observed in the spared contralateral white matter, remote to the site of hemisection (Figs. 5C, 5E, inserts) compared with the sham-operated control (Fig. 5A, insert). In this region, astrocytes appeared to be clustering and starting to cross-link (Figs. 5C, 5E, inserts, A3). The number of GFAP-IR astrocytes in this region (A3) increased over time and reached a statistically significant level at 30 days post-injury compared with both the 7-day and sham-operated groups (Fig. 5H, A3, p<0.05). The primary damage at the injury epicenter may lead to these secondary astrogliotic changes, even in regions remote to the lesion site after SCI in the monkey.
Collagen
We also examined the formation of fibrotic scar in the hemisection model in monkeys. Fibroblasts are the major cellular component of the fibrotic scar. 21 They are prominent in the deposition of collagen matrix, which can be detected by trichrome staining. 22 Collagen detected by aniline solution showed blue color with a slight baseline staining localized mainly in the gray matter region of the sham-operated controls (Fig. 6A). One week after injury, increased collagen staining was observed, particularly in regions surrounding the injury site (Figs. 6C, 6D), but no obvious fibrotic scar was observed. By 4 weeks post-injury, however, remarkably strong fibrotic connective tissue was formed to fill the space of the primary lesion (Figs. 6E, 6F). The fibrous scar tissue may be mainly derived from meningeal fibroblasts migrated into the lesion site after the hemisection when a durotomy was performed.
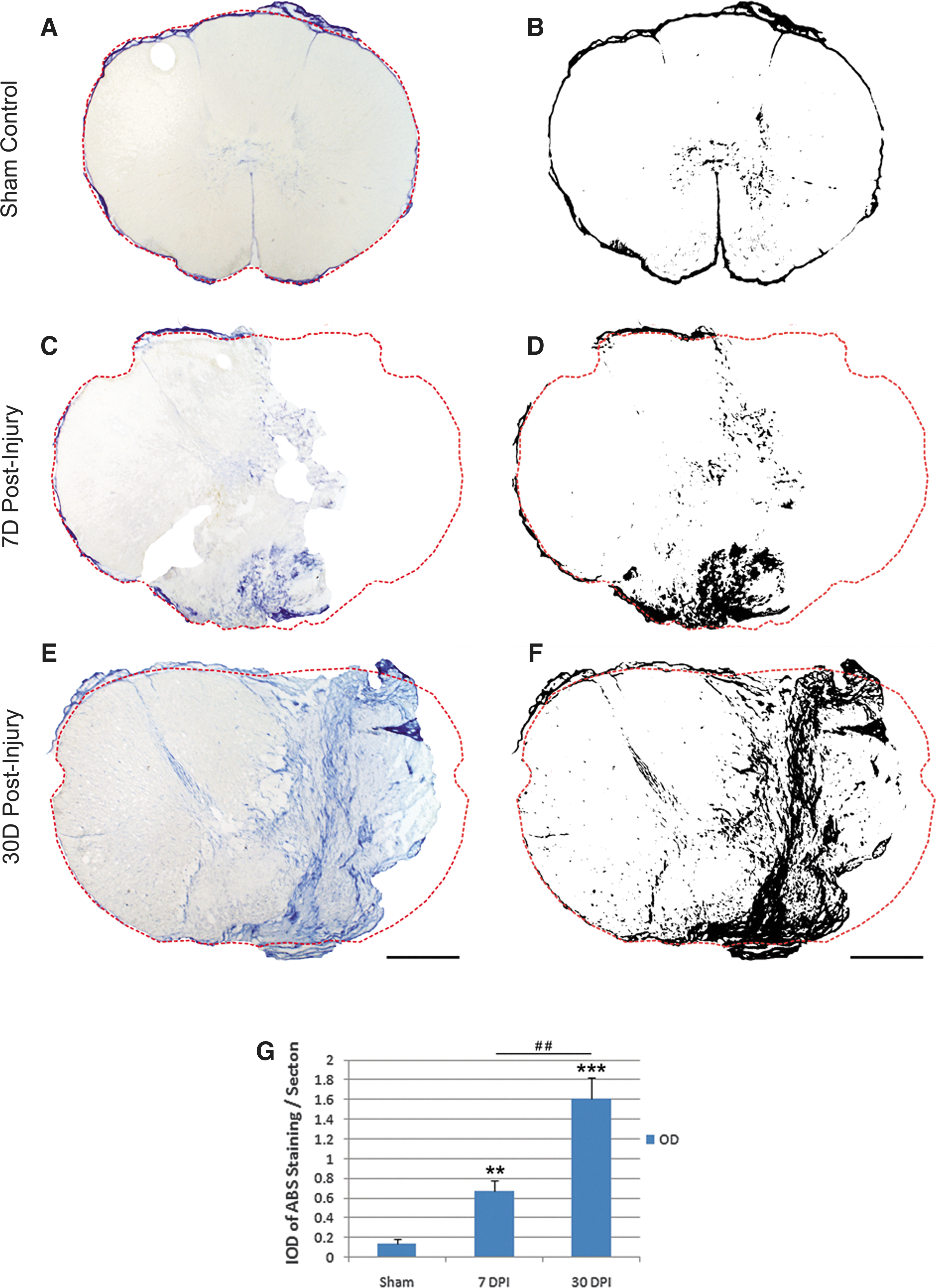
Characterization and quantification of collagen staining after a T9 right-sided hemisection in the monkey spinal cord. Representative photomicrographs show aniline blue staining in the sham-operated control
Discussion
In the present study, we performed a spinal cord hemisection at T8-9 in the monkey in which the right hemicord was either completely or partially transected at the injury epicenter. In the latter case, some ventromedial white matter was spared, but the dorsolateral white matter, containing major descending functional pathways, such as the corticospinal tract (CST), was interrupted. 15 This animal model mimics human penetrating SCI 2 and Brown–Séquard syndrome caused by the lateral hemisection of the spinal cord. This lesion model has been used for studying the responses of supraspinal neurons to axotomy, 13,23 and their plasticity and regeneration 11 in non-human primates. It should be noted that a clean hemisection is not easy and has been rarely documented in the literature. Therefore, we used “contralateral spared hemicord” to define the hemicord left on the contralateral side after surgical hemisection.
Compared with rodents, the monkey model of SCI has been significantly understudied. Although great progress has been achieved to improve treatment for SCI in rodents, 6,17 interspecies differences will need to be overcome in the transition from preclinical research to clinical trials. 24 Because of the difference in physiology and anatomy existing between rodents and humans, the monkey SCI model will be a valuable transitive model to verify promising findings observed in rodents before moving to the clinic.
Axonal response to a spinal hemisection
We examined axonal profiles in three areas: the lesion area (A1), the peri-lesion or penumbra area (A2), and the lateral white matter of the spared hemicord (A3), which is relatively distant from the injury site. After SCI, damaged axons at the distal end experience wallerian degeneration, 8,15,22 whereas those at the proximal end undergo a series of changes including swelling and dieback. 25 These axons, however, do not regenerate in the damaged CNS. Our results are similar to those of a recent study showing the decline and lack of axons in the monkey C7-8 unilateral cervical lesion. 26 The presence of some abnormal axonal profiles at the injury sites indicates that these cellular components underwent degenerative changes. The lack of robust axonal labeling in the lesion site at 4 weeks post-injury indicates that new axons failed to grow or sprouted into the lesion site, likely because of the presence of non-permissive CNS environment at the site of injury 6,27 or simply to the absence of permissive substrates needed for axonal growth. 28
To our surprise, in addition to the damage observed at the lesion and peri-lesion sites, significant axonal degeneration was also found in the lateral white matter of the spared hemicord, which is relatively distant from the injury site at 30 days post-injury. Such axonal degeneration is likely caused by a secondary injury process induced by a cascade of injury insults following SCI. 29,30 Alternatively, axonal loss in A3 at the level of injury could be caused by lesioning of crossed axons that contribute to the contralateral ascending pathways such as the spinothalamic tract. The presence of secondary axonal degeneration in the uninjured hemicord may provide an additional therapeutic target for neuroprotective and regeneration strategies.
Myelin response to a spinal cord hemisection
Loss of myelin (demyelination) and abnormal myelin (dysmyelination) in the white matter are main features of pathological changes after SCI resulting in functional deficits after SCI. 1,31,32 In the present study, we observed loss of myelin at the lesion epicenter, along with tissue loss at both 7 and 30 days after spinal hemisection. Compared with rapid loss of axons at the lesion epicenter, myelin breakdown was relatively slow, because myelin segments could still be detected at 30 days post-injury at the epicenter. The presence of a significant myelin loss at 1.5 cm caudal to the lesion epicenter may be a result of wallerian degeneration of long descending pathways, such as the CST.
We did not observe robust demyelination in the spared tissue on the contralateral side at both 7 and 30 days post-injury, similar to what was reported in human cases. 21 Characteristics of myelin pathology or dysmyelination, however, such as large onion bulbs, thinner myelin, and myelin debris, were found in the spared region of the contralateral white matter (Figs. 2D, 2F), where axonal loss was also observed. This dysmyelination could cause disruption of action potential propagation leading to partial nerve conduction block. 33,34 Whether dysmyelination in the spared white matter can self-repair or eventually leads demyelination is not known and should be further investigated.
Astroglial response to a spinal cord hemisection
Astroglial scar is considered as a physical and molecular barrier to regeneration. 35,36 Reactive astrocytes are a major source of the glial scar, and they secrete inhibitory extracellular matrix molecules such as chondroitin sulfate proteoglycans (CSPGs). Local administration of chondroitinase ABC (ChABC), an enzyme to digest CSPG, increased axonal regeneration after SCI. 37 In experimental rodents, SCI induced a rapid and pronounced increase in astrocytic response at the injury site 38,39 or along the area of wallerian degeneration. 8 These early and robust pathological changes of reactive astrocytes eventually form the glial scar surrounding the lesion in rodents combined with secretion of CSPGs and other extracellular matrix molecules. 27,36
Although astrocytes at the border of the lesion underwent hypertrophy, a robust response of reactive astrocytes to injury was not obvious in the monkey. We did not observe clear deposit of astroglial scar tissue along the border of the lesion up to 30 days after a T8-9 hemisection. Such a mild astrocytic response to injury observed at the lesion site in the monkey was similar to those described in human SCI. 21 It was reported that the extent of glial scarring in humans is modest at best and that a thick wall of astroglial processes is not a component of human SCI. 21
It is possible that the time windows for development of astroglial scar are different between monkeys and rodents. Human SCI pathology studies, however, support the lack of instead of delay in strong astroglial scar formation. In patients with SCI, a dense network of astroglial processes was never seen, with the injury to death interval ranging from instantaneous death to 51 years. 21
Further studies need to be conducted to determine the long-term astroglial responses to injury and the difference in astroglial responses to injury between rodents and primates. The lack of extensive astrocytic responses to SCI in the present study highlights the importance of using non-human primates to model human SCI, particularly when a promising strategy needs to be examined before moving to the clinic, because the pathology of SCI in the monkey is the closest to that in humans.
Interestingly, while the response of astrocytes at the edge of the lesion was mild, their reactivity was clearly increased in the spared contralateral white matter, remote to the site of hemisection (Fig. 5E, insert) compared with the sham-operated control (Fig. 5A, insert). These astrocytes exhibited hypertrophy and their processes were cross-linked, which can be appreciated particularly at 30 days post-SCI. This secondary astrogliotic response, occurring in regions remote to the lesion site in the monkey, was likely caused by the primary damage to the injury epicenter. The observation of increased astrocytic response in this remote tissue region is in agreement with axon loss and dysmyelination observed in the same region as discussed above.
Fibrotic response to a spinal cord hemisection
The scar tissue, which forms a major barrier to axonal regeneration, consisted of reactive astrocytes, meningeal fibroblasts, and blood-borne cells that invade the center of the lesion. 40 The scar tissue also contains extracellular matrix molecules such as collagen (types I, III, and IV), laminin, fibronectin, and proteoglycans secreted by these cells. 41 The scar tissue can be subclassified into astrocytic and fibrotic scars. The astrocytic scar, discussed above, is formed along the edge of the lesion. After injury, the astrocytic scar forms a new glial limitans that isolates the lesion from the remaining viable tissue, 21 which could be beneficial as a wound healing process after SCI. 42
The fibrotic scar, on the other hand, is formed by fibrous connective tissue and collagen that fills the lesion cavity/gap. 21 Such scars are particularly prominent in two classes of human SCIs—i.e., cord maceration and cord laceration, described by Bunge and colleagues. 1 Hemisection of the spinal cord in our case damaged the meningeal sheet, which might lead to the proliferation and penetrating of meningeal fibroblasts deep into the lesion. Such fibrotic scar, evidenced by a dense collagen meshwork that filled the primary lesion space, was particularly prominent at 30 days post-injury.
A similar fibrotic scar at the lesion epicenter, accompanied with robust surrounding astroglial scar, was documented in rat and mice models of penetrating SCI. 43,44 Although we observed that, in adjacent sections, astroglial scar and fibrotic scar occupied their own territories, scattered GFAP+ astroglial profiles were also found within the fibrotic scar indicating some overlap between the two.
In humans, the formation of fibrotic scar is not beneficial and clearly represents an impediment to axonal growth. 21,45 The lack of axons in the scar tissue supports this notion. Reducing or removing the formation of a fibrotic scar, therefore, would be a feasible and clinically relevant strategy aimed at promoting axonal regeneration.
Inflammatory response to a spinal cord hemisection
Spinal cord injury induces an inflammatory reaction that is a complex phenomenon involving the activation of resident microglia and the infiltration of neutrophils, monocytes/macrophages, and lymphocytes into the lesion from the systemic immune system. 30,46 –48 At the lesion site, macrophages are derived from both microglia and hematogenous macrophages and are referred to as microglia/macrophages because they cannot be distinguished by morphology or antigenic markers.
Activated microglia/macrophages in the injured rodent cord are considered to be detrimental by releasing pro-inflammatory cytokines, reactive oxygen species, nitric oxide, and proteases, 47,49 leading to tissue damage, demyelination, and neurological dysfunction after SCI. 50 Attenuation of the early inflammatory response to SCI may therefore limit the extent of tissue injury and promote recovery. 51
Although these evidences indicted that microglia/macrophages result in secondary tissue damage in CNS injuries, other studies have reported protective effects of macrophages under some conditions. 52 They participate in the removal of injured tissue debris and the release of protective cytokines that promote neuronal regeneration, wound healing, and tissue repair. 53 Hence, the development of effective treatments for human SCI requires a precise knowledge of the action and time course of entry of various inflammatory cells into the spinal cord and activation of resident microglia. In contrast to the significant amount of information available from experimental rodents, limited information is available concerning inflammatory responses in the monkey after SCI.
In the present study, we observed a significant decrease in the number of microglia at 30 days after SCI in both the lesion epicenter and penumbra regions compared with either the 7-day or sham-operated group. We further observed a significant increase of KP1-IR microglia/macrophages at the injury epicenter and penumbra regions at both 7-day and 30-day time points with a peak observed at the 7th day. Notably, a significant increase in the number of KP1-IR microglia/macrophages was found in the lateral white matter of spared hemicord, indicating a remote activation of microglia/macrophage, possibly from a spread of inflammatory response to the spared side of the spinal cord.
This observation is consistent with previous reports that resident microglia convert to the macrophage phenotype at subacute or chronic stages in human SCI. 54 The presence of activated microglia, with morphology in transition from resting microglia toward the macrophage phenotype, contributes to phagocytic macrophages in the lesion. These microglia/macrophages can cause the secondary injury by extending into more intact tissue adjacent or even remote to the lesion core, and potentially worsening neurological dysfunction for weeks to months after the injury.
It seems that processes of microglia and macrophage reactions are similar among rodents, non-human primates, and humans. The time course, however, can be different. For example, in rats, microglia become activated within minutes to hours after SCI and are transformed into macrophages. 49 Meanwhile, the blood-borne monocytes/macrophages infiltrate the lesion, achieve the peak at 5–7 days, and persist for weeks to months. 47,55 In humans, the distribution of activated microglia/macrophages occurs in the contused spinal cord, with post-injury duration from 5 days up to months. 56,57
Conclusion
We systemically examined neuropathology at the injury site in the monkey after a T8-9 spinal lateral hemisection at two clinically relevant time points—i.e., the subacute and early chronic stages. We not only examined the lesion epicenter and the peri-lesion or penumbra area, but also the remote region of spared, contralateral hemicord. In general, the pathology and cellar responses to injury in the monkey are closer to humans than rodents. Unexpectedly, degenerative changes including axonal loss, dysmyelination, reactive gliosis, and activation of microglia/macrophages were also observed in the remote region of the spared hemicord, particularly at the chronic stage, suggesting that even the spared tissue can be an important target for therapeutic intervention. The lack of substantial reactive astrocytic response at the lesion border and presence of robust fibrotic scar within the lesion center further emphasize the importance of using non-human primates in testing repair strategies aimed at addressing specific problems associated with human SCIs.
Footnotes
Acknowledgments
The authors thank the generous support provided by the PLA Clinical Center for Spinal Cord Injury, Kunming General Hospital of PLA, Kunming Tongren Hospital, and Jinghong Breeding Station, P.R. China (to ZH), the Major State Basic Research Development Program of China (973 Project 2003CB515302) (to PL), and the Mari George Hulman Endowment (to XMX).
Author Disclosure Statement
No competing financial interests exist.